
Abstract
Microbial life on Earth uses a wide range of chemical and energetic resources from diverse habitats. An outcome of this microbial diversity is an extensive and varied list of metabolic byproducts. We review key points of Earth-based microbial metabolism that are useful to the astrophysical search for biosignature gases on exoplanets, including a list of primary and secondary metabolism gas byproducts. Beyond the canonical, unique-to-life biosignature gases on Earth (O2, O3, and N2O), the list of metabolic byproducts includes gases that might be associated with biosignature gases in appropriate exoplanetary environments. This review aims to serve as a starting point for future astrophysical biosignature gas research. Key Words: Exoplanet—Biosignature—Microbial redox reactions. Astrobiology 12, 61–82.
1. Introduction
T
Earth analogs by definition are potentially habitable 1 . For the present, however, detection and spectroscopic observations of Earth analogs are beyond technological reach. Astronomers are instead close to finding a different kind of habitable planet: big Earths orbiting small stars (Tarter et al., 2007, and references therein). Selection effects favor their discovery and characterization compared to Earth analogs in almost every way (Nutzman and Charbonneau, 2008). Follow-up observations will be possible for a subset of exoplanets orbiting small stars—transiting exoplanets, those that go in front of their stars as seen from Earth. Challenging atmospheric observations of super-Earths transiting small stars will be possible in the next decade with the launch of NASA's James Webb Space Telescope (e.g., Deming et al., 2009; Seager and Deming, 2010).
Despite a short-term focus on super-Earths orbiting in the habitable zones of low-mass stars, it is only natural that investigators desire to discover and characterize another planet like our own, a true Earth analog. The ability to study true Earth analog atmospheres is still feasible in the longer term. Highly specialized space telescopes with mirror diameters in the 4–8 m range will be able to directly image Earth analog planets (Cash, 2006; Trauger and Traub, 2007; Soummer et al., 2009, and references therein).
Most discussions of exoplanetary biosignature gases have been limited to O2 (and its photolytic product O3), N2O, and CH4. These are the important, global biosignature gases on Earth (see Fig. 1). (See Section 2.2 for a discussion of more minor terrestrial biosignature gases considered in the literature.) Yet the microbial world on Earth is incredibly diverse, and microorganisms yield a much broader range of metabolic byproducts. In an environment different from Earth's, these metabolic byproducts may accumulate to produce biosignature gases different from those on past and present Earth.
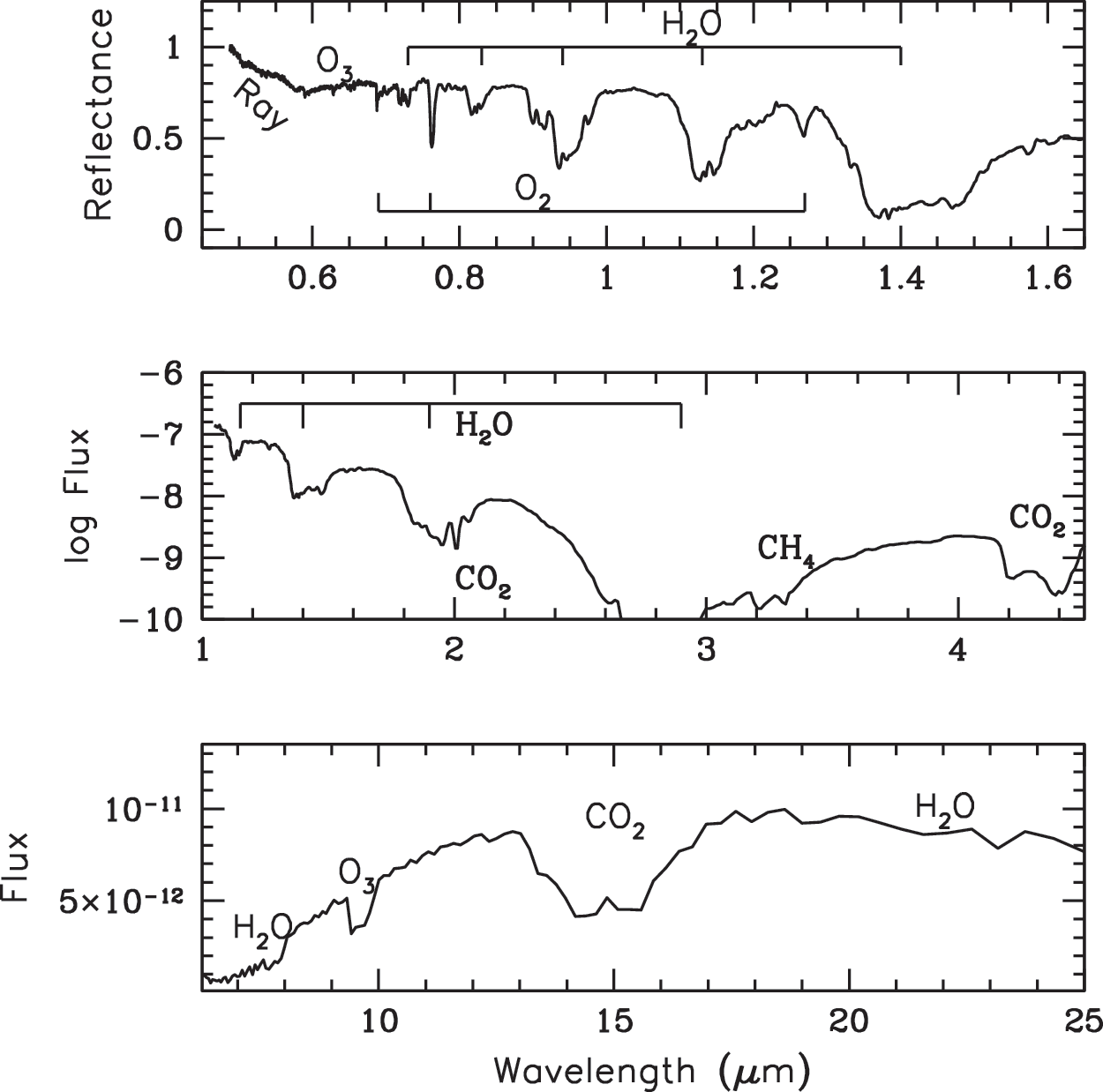
Earth as an exoplanet: Earth's hemispherically averaged spectrum. Top: Earth's visible wavelength spectrum from Earthshine measurements plotted as normalized reflectance (Turnbull et al., 2006). Middle: Near-IR spectrum from NASA's EPOXI mission with flux in units of W m−2 μm−1 (Robinson et al., 2011). Bottom: Earth's mid-IR spectrum as observed by Mars Global Surveyor en route to Mars with flux in units of W m−2 Hz−1 (Christensen and Pearl, 1997). Major molecular absorption features are noted including Rayleigh scattering (noted as “Ray”). Only strongly absorbing, globally mixed molecules are detectable.
The goal of this paper is to review the chemical outputs of life on Earth. The motivation is to provide a starting point for the systematic discussion of biosignature gases on exoplanets beyond Earth. To do this, we review Earth-based metabolic reactions and byproducts, with the hope that, with further research into their planetary context, some of the byproduct gases might be viable biosignature gases on other planets.
This paper is structured as follows. In Section 2, we present background for biosignature gases created from metabolic reactions. We discuss terrestrial metabolic inputs and outputs in Sections 3 and 4, deferring discussion of their planetary context to Section 5. In Section 3, we review the major redox reactions performed by terrestrial life, and in Section 4 we review “secondary” metabolism byproducts, gases produced by life but not as byproducts of their central chemical functions. In Section 5, we discuss why most of Earth's inorganic metabolic byproducts are not unique biosignature gases, and we consider implications for exoplanetary biosignature gases.
2. Background to Earth-Based Biosignature Gases
With our astrophysical view, we examine the metabolic byproducts generated by microorganisms. By focusing on net metabolic “inputs” and “outputs,” we strive to achieve a viewpoint that is relatively independent of the complex set of evolutionary factors that dictate extant life's underlying physiology, genetic makeup, and chemical structure. This enables us to focus on a “top down” approach of a biosignature framework. In the top-down approach, we do not focus on what life is but instead on what life does. Life metabolizes and, for our purposes, generates metabolic byproducts. In this way, we leave the “bottom up” approach—the details of the internal mechanisms and origins of life—to others. (For an overview of how life generates energy from diverse chemical reactions, and the internal transactions of energy metabolism, see, e.g., Nicholls and Ferguson, 2002).
Our motivation is that a review of metabolic byproducts may eventually lead to new ideas for biosignature gases on other worlds. For example, a particular metabolic byproduct gas that plays only a minor role in Earth's atmosphere may accumulate to detectable levels on an exoplanet with different sources and sinks from Earth's.
We divide biosignature gases into two categories to provide a framework for future, quantitative analysis of the feasibility of gases as biosignatures in exoplanetary atmospheres. The first class of biosignatures is gases generated by redox chemistry. (Redox chemistry is defined below and in the Appendix.) Redox reactions are used by life both for converting environmental chemical gradients into usable metabolic energy and for converting environmental chemicals to biomass. The two different uses for redox reactions might be considered as separate classes of use, but we will treat them as one class for this review. By contrast, the second class of metabolic byproducts we define is produced for diverse reasons by life, in amounts that do not correlate with either the chemistry of the molecules or the mass or number of organisms producing them. We refer to these as “secondary” metabolic byproducts.
We note that both primary and secondary metabolism byproducts can be transformed abiotically. The resulting product might also not be naturally occurring in Earth's atmosphere and therefore also a sign of life (e.g., O2 photochemically converted to O3). These abiotically altered products can be considered as a separate subclass of each of primary and secondary metabolism byproducts.
2.1. Biosignature gases from redox reaction byproducts
Central to all life on Earth is reduction-oxidation (redox) chemistry (e.g., Metzler, 1977). Redox reactions are those in which electrons are transferred between molecules or compounds, or both. The Appendix describes redox reactions in more detail and expands on the more specialist terminology used in this paper. Redox reactions are used by life to exploit chemical energy gradients and are also a key part of chemistry of metabolism, which includes production of energy-storing materials. Some of the products in redox reactions end up as “waste” or “byproducts” and can accumulate in the atmosphere as biosignature gases. Canonical examples for exoplanets are CH4 from methanogenesis and N2O from nitrate reduction (see Section 3 and Des Marais et al., 2002).
In our “astrophysical view” of Earth-based life, both “light energy” (phototrophy) and “chemical energy” (chemotrophy, which can also be called “dark energy”) reactions can be viewed under the same framework. Both light and dark energy reactions involve chemical energy gradients, as follows. Photosynthesis converts light energy into chemical energy by exciting an electron from a pigment-protein complex, which donates that electron to reduce subsequent acceptor molecules along an electron transport chain. The oxidized complex regains its electrons by oxidizing molecules from the environment (such as H2O or H2S). The donor and acceptor molecules form a redox pair. This redox pair is functionally equivalent to an energy gradient—two separate molecules that, if allowed to come together to react, will release energy—but one that is internal to the cell. Dark energy exploits chemical energy gradients that exist naturally outside the cell, in the environment, and chemotrophy is the process that exploits the dark energy.
Redox reactions that capture energy (used for metabolism and building biomass) and redox reactions that capture biomass, acquiring and transforming the materials necessary for an organism's growth and repair (e.g., Smil, 1991), are closely intertwined in Earth-based biochemistry. In this review, we focus on energy-generating reactions and not on biomass building.
2.2. Biosignature gases generated from “secondary” metabolism processes
“Secondary” metabolism byproducts are gases produced by life but not as byproducts of their central chemical functions. Primary metabolism (including redox reactions described above) captures energy from the environment, builds the basic molecules of life, and (in the case of autotrophic organisms) captures carbon and turns it into a form suitable for living things to use (e.g., see the textbook by Alberts, 2008). The basic reactions of primary metabolism are common to wide groups of organisms, and the core reactions for transforming chemical energy and assembling the basic building blocks of life—amino acids to make proteins, sugars, and bases to make DNA—are common to all life.
Secondary metabolism, in contrast to primary metabolism, is the synthesis of compounds that are not directly involved in growth of an organism but may influence organism fitness or be byproducts of stress. Secondary metabolism products appear to be special to particular species or groups of organisms. Secondary metabolism compounds include, for example, those produced under heat stress or those utilized for ecological interactions such as defense or attraction of pollinators. The term “secondary metabolism” is sometimes used specifically to designate the production of secreted chemicals such as antibiotics by microorganisms or the production of complex, aromatic molecules, alkaloids, and pigments in plants. We discuss some of these plant chemicals below. In this paper, we use the more general definition of “secondary metabolism” to mean the production of chemicals that are not part of primary metabolism and are not produced as part of energy metabolism. If a chemical produced by secondary metabolism is a gas, it could be an astrophysical biosignature. We term such chemicals “secondary metabolism byproduct gases” or “secondary metabolism biosignature gases.” Secondary metabolism, for example, produces the chemicals that confer unique colors on flowers, smells on herbs, and tastes on vegetables, each species making a combination of molecules not made by any other.
Secondary metabolism potential biosignature gas molecules previously described in the literature include dimethyl sulfide (CH3·S·CH3) and methanethiol (CH3·SH) (Pilcher, 2003) and methyl chloride (CH3Cl; Segura et al., 2005). A review of secondary metabolism products is worthwhile because some are present in sufficient quantities to materially alter local or even global atmospheric chemistry (Andreae and Crutzen, 1997; Stavrakou et al., 2012).
The relevance of secondary metabolism byproduct gases in an astrophysical context, however, is uncertain. The role of many secondary metabolism byproduct gases is not known (e.g., isoprene, methyl chloride), while others are clearly the result of particular evolutionary history and competitive needs. The chemical nature of the secondary metabolism byproduct gas, and the amount that is released into the atmosphere, is not determined by the local chemistry and thermodynamics, so we cannot point to an environment and argue why a particular organism makes one chemical rather than another. In other words, we know the chemicals are made biotically but cannot always rationalize them environmentally.
A good example of a secondary metabolism byproduct gas is dimethyl sulfide (DMS). DMS is made by marine phytoplankton, and the oceans release between 15 and 60 Tg of DMS per year into the atmosphere (Watts, 2000). Once in the atmosphere, DMS was believed to play a significant role in the modulation of climate, but this is now considered unlikely (Quinn and Bates, 2011). However, the organisms spend energy and lose carbon to make DMS. The reason that DMS is made and released by marine phytoplankton is a complex web of causes that are, apparently, quite specific to the phytoplankton and the organisms that prey on them in Earth's oceans (summarized below). There is no a priori reason that we should expect DMS to be a biosignature gas on an exoplanet, although if we saw it in another planet's atmosphere we would suspect that it had been produced by life. Geochemistry is unlikely to generate DMS spontaneously; DMS is thermodynamically unstable to oxidation to dimethyl sulfoxide (DMSO) and to reduction by hydrogen to CH4 and H2S. Any plausible geochemical source would be oxidizing or reducing. See Pilcher (2003) for further discussion of DMS as a biosignature gas and Domagal-Goldman et al. (2011) for further discussion of sulfur compounds as potential biosignature gases.
3. Redox Metabolic Byproducts
In this and the following section, we describe the major net chemical transformations performed by life on Earth and the product gases. We leave discussion of sources and sinks of the metabolic byproduct gases until the Discussion Section 5.
3.1. Phototrophy
Phototrophy is the process that converts light energy into chemical energy in living organisms. The term derives from the roots photo (light) and troph (eater). Vegetation, algae, and photosynthetic bacteria use light energy to power the “fixation” of carbon 2 compounds in a process known as photosynthesis. In our astrophysical view, phototrophy harvests solar radiation to create chemical energy gradients across membranes inside the cell, which are then used for carbon and nitrogen fixation.
3.1.1. Oxygenic photosynthesis
We begin with oxygenic photosynthesis, as the generation of oxygen by photosynthesis has yielded the clearest evidence of life's impact upon Earth's atmospheric chemistry.
Cyanobacteria, algae, and plants perform oxygenic photosynthesis via
[See, e.g., Hall and Rao (1999) for a general introduction to photosynthesis, with an emphasis on the oxygen-generating light reactions.] Here, CH2O represents carbohydrates in a general form, which are then converted into numerous, diverse biomolecules.
The above reaction includes light harvesting (photo-) and biomass building (synthesis) via carbon fixation. In the astrophysical view, we disentangle these reactions because we are primarily interested in the metabolic process that harvests the energy and the eventual metabolic byproducts (or outputs). We therefore describe oxygenic photosynthesis as
In Reaction 1, separating the generation of electrons (Reaction 1a) from the synthesis of carbohydrate (Reaction 1b) represents biochemical reality and is not an arbitrary reformulation. The two processes, light capture and biomass building, are mechanistically distinct. A complex of proteins and pigments, including the characteristically green chlorophyll, are responsible for electron generation independent of carbon fixation (e.g., Babcock, 1987). The oxygenic phototrophs produce oxygen as a metabolic byproduct, using H2O as the electron donor. The protons and electrons that result from the splitting water reaction are channeled into intermediates to create a chemical energy gradient. The constituents of the reaction, H2O, H+, and e−, are common to the myriad of redox reactions carried out by microorganisms and are, by and large, channeled into intermediary metabolism—that is, the biomass-building reactions (Reaction 1a and 1b).
For the purposes of this review, a clearer perspective as to the net chemistry of metabolism could be represented in the following schematic, which separates inputs from outputs.
Note that in this review we do not attempt to write or balance formal chemical reactions; metabolism carries out an internal network of very complicated reactions, usually with many inputs and outputs. Here, we are interested in the net change caused by one reaction type. For examples of the real-world, complex stoichiometry of bacterial net metabolism see, for example, Heijnen (1994).
Oxygen dominates Earth's biosignature gases in part because of the vast amount of sunlight and essentially unlimited supply of water available to surface-dwelling life. Although on early Earth there were plenty of photons, H2O, and CO2, there was no O2 signature, because life had not yet the capacity to split H2O as a source of electrons for fixing CO2.
On modern Earth, organisms that can split water to generate electrons dominate the biosphere and fix at least 1000 times as much carbon as all other forms of life (Des Marais, 2000).
The easy availability of reactants in Reaction 1c means that O2 is produced in large quantities. O2 accumulates in the atmosphere because a small fraction of the carbon fixed by Reaction 1 is trapped in living organic matter and buried in sediments, so it is not available for reverse reaction with O2. Furthermore, O2 is a good biosignature gas because its atmospheric lifetime against photochemical sinks is long enough to accumulate to detectable levels but short enough to combine with other molecules; thus extant O2 would indicate replenishment and highly suggest a biotic origin.
O2 is considered Earth's most robust biosignature gas because no known geophysical or photochemical process can generate such large quantities of O2 (Des Marais et al., 2002), and false-positive cases can most likely be identified by other atmospheric diagnostics. For example, dissociating water in a runaway greenhouse with H escaping to space could lead up to detectable O2 levels. This situation could be identified by detection of an atmosphere heavily saturated with water vapor. O2 could also accumulate in a dry, CO2-rich planet with weak geochemical sinks for O2, a case which could be identified via weak H2O features (Selsis et al., 2002; Segura et al., 2007).
Oxygenic photosynthesis is an ancient process on Earth. There is fossil evidence of organisms resembling present-day cyanobacteria from 2.7 billion years ago (Brocks et al., 1999; Summons et al., 1999) and widespread global evidence of their impact—via oxygenation—by 2.45 billion years ago (Farquhar et al., 2000). (The evidence for the rise of atmospheric oxygen is the end of mass-independent fractionation of sulfur isotopes in sedimentary fossils.)
Oxygenic photosynthesis is not the only mechanism by which life captures light energy as chemical energy. Before oxygenic photosynthesis evolved, organisms used a variety of other abundant chemical sources of electrons in photosynthesis (Canfield et al., 2006), and some still use these alternative chemistries today.
3.1.2. Anoxygenic photosynthesis
There is a diverse set of organisms that do not produce oxygen as a byproduct of photosynthesis, known as the anaerobic photosynthetic bacteria (for a review see, e.g., Pfennig, 1977). Instead of using H2O as the source of electrons, these organisms use inorganic sulfur compounds (H2S, S°), ferrous iron (Fe2+), nitric oxide (NO), hydrogen gas (H2), or organic compounds. Like oxygenic photosynthesis, anoxygenic photosynthesis results in compounds that are more oxidized than the inputs. The resulting more oxidized byproducts of anoxygenic photosynthesis include compounds such as elemental sulfur, sulfuric acid (H2SO4), ferric iron (Fe3+), and other less abundant compounds [e.g., arsenate (
An example of anoxygenic photosynthesis is that of green sulfur bacteria with the reaction
where H2S is split by photons to yield an electron donor, CH2O represents the carbohydrates incorporated into the microbe, and S and H2O are the metabolic byproducts. On ancient Earth, hydrogen-based photosynthesis was probably also important (Olson, 2006),
Both the hydrogen and the hydrogen sulfide for these reactions are produced today in mid-ocean ridge volcanism. On early Earth, hydrogen was probably the more important electron donor (Tice and Lowe, 2006).
In our astrophysical view, we can break up the anoxygenic photosynthesis reactions to disentangle these reactions in analogy with oxygenic photosynthesis (Reactions 1a and 1b):
Because we are primarily interested in the metabolic process that harvests the energy and the eventual metabolic byproducts (or outputs), we therefore describe H2S-based anoxygenic photosynthesis as Reaction 2a. Together with other anoxygenic photosynthetic metabolic energy-yielding processes we have the Reactions 2 (e.g., Garlick et al., 1977; Padan, 1979; Madigan et al., 2003; Canfield et al., 2006; Olson, 2006).
Oxygen atoms that appear in product molecules are usually obtained from water. We do not show water in the input molecule list because in these reactions the oxidation state of the oxygen and hydrogen in water is not changed. Omitting water from the input list is intended to make clear that energy from light (hν) is used to change the oxidation state of the listed molecule, not of water.
The anoxygenic photosynthesizers live in a variety of anoxic environments. For some anoxygenic photosynthetic organisms, oxygen is actively poisonous. Some organisms can switch between oxygenic and anoxygenic photosynthesis to take best advantage of available nutrients, and usually the presence of oxygen inhibits anoxygenic photosynthesis (Garlick et al., 1977). The most common habitats for anoxygenic photosynthesizers are illuminated but anoxic, including freshwater lakes and ponds, hot springs, sulfur springs, and some marine waters. The sources of electron donors (e.g., H2S) can be either geological (in sulfur springs) or biological (produced by sulfate-reducing bacteria), depending upon the setting. In evolutionary terms, anoxygenic photosynthesis is believed to have preceded oxygenic photosynthesis and to have appeared on Earth more than 3 billion years ago (Des Marais, 2000). One interesting aspect of anoxygenic photosynthesis is that some photosynthetic bacteria are able to deposit the solid sulfur (S) inside the cell, where it is stored for later use if required, as would be the case if H2S and
3.1.3. Geothermal energy as a radiation source for photosynthesis
While starlight is the most obvious source of light energy, recent studies suggest that photosynthetic green sulfur bacteria can even be sustained by the dim glow of geothermal blackbody radiation emitted by hot volcanoes on the seafloor. While the overall net metabolism of these organisms is similar to anoxygenic photosynthesizers near the Earth's surface, these photosynthetic bacteria live and photosynthesize 2.5 km beneath the ocean surface (Beatty et al., 2005). The energy supply is limited, due to the low numbers of photons at red wavelengths (at ∼1011 photons cm−2 s−1 sr−1 over the 600–1000 nm range), and the organisms contain specialized photopigments adapted to these wavelengths of light.
With such small numbers of photons compared to solar, geothermal radiation-based photosynthesizers likely have very slow growth rates. For example, a Black Sea green sulfur bacterium living at 80 m water depth in the Black Sea—and receiving a water-attenuated solar radiation intensity similar to the geothermal radiation described above—is estimated to have an in situ cell division time on the order of 2.8 years, compared to hours to days for most surface-dwelling organisms (Overmann et al., 1992). The example of deep-sea anoxygenic photosynthetic organisms show that life can grow by using light sources orders of magnitude fainter than the light provided by the Sun at Earth's surface.
3.2. Chemotrophy
Chemotrophy describes the process whereby potential energy in the form of chemical disequilibria is channeled into cellular metabolism. Specifically, chemotrophic organisms couple energetically favorable pairs of oxidation and reduction half-reactions. The disequilibria can involve either completely inorganic compounds or make use of organic matter. In fact, the only clear limitations upon the types of reactions used are that they have a negative Gibbs free energy 3 and that life can make the reactions occur faster than the rate of nonbiological reactions. In other words, Earth-based metabolic pathways exploit chemical energy potential gradients in the form of chemical reactions that are thermodynamically favorable but kinetically inhibited. (e.g., Madigan et al., 2003 for more details).
In the forthcoming sections, we will describe the most common chemotrophic bioenergetic reactions used on Earth. We emphasize that our list of common chemotrophic reactions is not an exhaustive one; organisms can specialize such that they derive nutrients from most unexpected, and limited, environments, for example, from the chemicals in stainless steel containers of spent nuclear fuel (Chicote et al., 2005), from phenol (the active ingredient in traditional “carbolic” antibacterial soap) (Aneez Ahamad and Mohammad Kunhi, 2011), and the chemicals in “plaque” from blocked human arteries (Mathieu et al., 2007). Certain species of Pseudomonas (a bacterium) are able to metabolize over 100 different compounds (Madigan et al., 2003). We do not address such exotic metabolisms in this review, since they are either limited to human-made materials or their astrophysical significance is likely limited.
3.2.1. Aerobic chemotrophy
If oxygen itself is a sign of life, why review aerobic (oxygen-consuming) metabolisms? A motivation is that oxygen may be available in an exoplanetary atmosphere as an electron acceptor for energy-generating redox reactions, even if the oxygen is not at high-enough levels to be a remotely observed spectral feature. For example, we can imagine a super-Earth massive enough to hold on to a hydrogen-rich atmosphere that would create a very long oxidation time. In this situation, hypothetically, oxygen-consuming life-forms may use oxygen (created by other life), even without a significant excess of oxygen accumulation to oxidize the atmosphere and produce an oxygen gas biosignature. This scenario is somewhat similar to the mildly reducing atmosphere on early Earth just before the rise of oxygen. This kind of hydrogen-rich atmosphere hypothetical scenario motivates us to review the metabolic byproduct molecules produced by aerobic chemotrophs.
(Chemo)-heterotrophy. The aerobic oxidation of organic matter by life (termed heterotrophy) is probably the most pervasive and well-known bioenergetic reaction on Earth after photosynthesis. While the oxidation of glucose-like intermediates and complex organic compounds is more commonly recognized, microorganisms also catalyze the oxidation of a number of small organic molecules, such as short-chain alkanes (methane, ethane, and propane) and small carboxylic acids (e.g., formate, acetate), which may be produced abiologically.
Although the oxidation of organic matter can produce a number of intermediate compounds, the ultimate products of aerobic respiration are CO2 and H2O. CO2 is already present in terrestrial planetary atmospheres (at the 97 percent by volume level on Venus and Mars), and H2O vapor is usually taken as an indicator of liquid water oceans and not as a biosignature.
The astute reader will notice that Reaction 3 is the reverse of Reaction 1. The net effect of the two reactions together (Reactions 1 and 3), however, does not consume all the atmospheric oxygen. Oxygen accumulates in the atmosphere because a small fraction of the carbon fixed by Reaction 1 is buried in sediments and so is not available for use in Reaction 3. We discuss sources and sinks further in Section 5.
Hydrogen oxidation. Among the completely inorganic “fuels” used for biosynthesis, the aerobic oxidation of hydrogen has the largest free energy yield. Direct oxidation of H2 by O2 by microorganisms is called the Knallgas reaction. Hydrogen itself can be derived from numerous reactions, including the breakdown of preformed organic matter (through fermentation) from geothermal sources or from water-rock reactions (serpentinization). Although oxygen is the most widely used oxidant, hydrogen peroxide can also be used to respire hydrogen to water. The most common habitats for organisms that use the Knallgas reaction are those at interfaces between anoxic sources for hydrogen and oxygenated fluids. For example, many types of hydrogen oxidizers have been characterized from terrestrial hot springs and deep-sea hydrothermal vents. The byproduct of aerobic hydrogen oxidation is water, which makes its net geochemical impact difficult to detect in aqueous environments.
Oxidation of sulfur compounds. Following hydrogen, sulfides (specifically H2S) yield the most energy when oxidized by oxygen as fuel for chemotrophic growth. Because sulfur compounds can have a range of oxidation states, some of the compounds can be both reduced and oxidized by different groups of organisms.
Sulfur compounds used in energy-generating redox reactions by life in order of increasing oxidation state are
From left to right the chemical formulae are for hydrogen sulfide, elemental sulfur, thiosulfate, sulfite (and sulfur dioxide), and sulfate.
H2S, elemental sulfur (S), and S2O3 are all oxidized by microorganisms that use oxygen as an electron acceptor. Aerobic sulfur-oxidizing microorganisms are also responsible in large part for the weathering of metal sulfide minerals (e.g., FeS2), such as weathering that occurs in acid mine drainage (Rawlings, 2002). The source of H2S can be either biological (anaerobic bacterial sulfate reduction) or geochemical (such as sulfide springs and volcanoes). The other intermediate sulfur compounds used as electron donors are commonly generated as byproducts of sulfur cycling.
Oxygen atoms appearing in product molecules, as noted before, are usually obtained from water. We do not show water in the input molecule list because in these reactions the oxidation state of the oxygen and hydrogen in water is not changed. Omitting water from the input list is intended to make clear that chemical energy is derived from changes in oxidation state of the input molecule.
The sulfur compounds used in microbial metabolism can encompass a range of phases. Elemental sulfur, S, is relatively insoluble in water and thus usually occurs as a solid. Some bacteria deposit S inside the cell, and some deposit it outside, both for later use.
Iron oxidation. When combined with oxygen, following sulfur, ferrous iron (e.g., Fe2+) is the next most energy-yielding fuel. Ferrous iron either occurs in the solid form (metal sulfides) in acidic aqueous solutions such as acidic mine drainages (Rawlings, 2002), or in highly reduced fluids such as those at deep-sea hydrothermal vents.
The generalized metabolic reaction for aerobic iron oxidation (under neutral or alkaline conditions) is
Iron oxidation is an energy-yielding process but provides only a small amount of energy (Emerson et al., 2010). The bacteria therefore must use significant quantities of ferrous iron (Fe2+) to grow and as a result produce large amounts of ferric iron (Fe3+). Where the Fe2+ is found as sulfides (FeS and FeS2), other bacteria in the ecosystem reduce the Fe3+ back to Fe2+ by oxidizing sulfide to sulfate (Rawlings, 2002). If there is no mechanism to recycle the Fe3+, the byproduct of iron oxidation is typically insoluble FeO(OH) “rust.” Rust is a solid. The growth sites are geological deposits of iron sulfide minerals where H2O and O2 are present and are typically acidic. At neutral pH, Fe2+ is rapidly oxidized chemically by oxygen to Fe3+, but only slowly at acid pH (Emerson et al., 2010). Iron oxidation is another interesting example of how life exploits reactions that are just slightly kinetically inhibited.
Ammonia oxidation “nitrification.” Nitrification is the process of converting ammonia into nitrate involving the following steps.
NH3 is produced on Earth almost exclusively from biological processes, apart from that manufactured industrially. The common byproducts for ammonia oxidation are nitrogen species more oxidized than NH3 (
Nitric and nitrous acids are strong acids, that is, in seawater the equilibrium toward dissociation into the nitrate and nitrite ion is far toward the formation of the ion. Volatile forms are therefore going to be negligible with no atmospheric biosignature for remote detection.
Oxidation of metals and other inorganic compounds. Microbes can oxidize a variety of metals and other inorganic compounds. Because of their oxidizing ability, microbes are exploited for microbial leaching (concentrating metal ores in mining production) in the acidic environments of mines. Microbes are able to oxidize metal sulfide ores, including iron and copper sulfide ores, and uranium- and gold-containing ores (Madigan et al., 2003). All of these are likely to occur in relatively low quantities on terrestrial planets and are not highly relevant from the astrophysical viewpoint of potential biosignature gases. Furthermore, the products will be nonvolatile metal salts and so will not be suitable biosignature gases even if they are produced in significant quantities.
3.2.2. Anaerobic chemotrophy
During the first half of Earth's history (approximately the first 2.3 billion years), Earth's atmosphere contained virtually no oxygen (<2 ppm) (e.g., Kump, 2008; Koehler et al., 2010). However, microorganisms that did not require oxygen were abundant and thrived. The present-day Earth retains anoxic environments away from the planetary surface (such as in subsurface rocks and ocean sediments). The motivation for reviewing anaerobic metabolism is that, in contrast to Earth, we anticipate that many exoplanetary surface layers will be anaerobic. A life-bearing planet may have an anaerobic surface due to the absence of oxygen-producing organisms or because a small number of oxygen-producing organisms do not dominate the redox state of the atmosphere (e.g., owing to very long oxygenation times for massive planets that lose their putative early H-rich atmospheres slowly). In such a scenario, without an atmospheric sink of oxygen, anaerobic metabolic byproducts might accumulate in an exoplanetary atmosphere as viable biosignature gases.
Earth-based microorganisms living in anaerobic environments, by definition, do not use oxygen for energy-yielding reactions. Instead, anaerobic microbes use a variety of other electron acceptors. This process is referred to as “anaerobic respiration.” On the negative side, anaerobic respiration always yields less energy per mole of reactant than aerobic respiration (see Fig. 2). On the positive side, anaerobic respiration enables life to extract energy from very diverse environments on Earth, where oxygen is absent. We will go through the various metabolic groups of anaerobes based upon the oxidants they use.
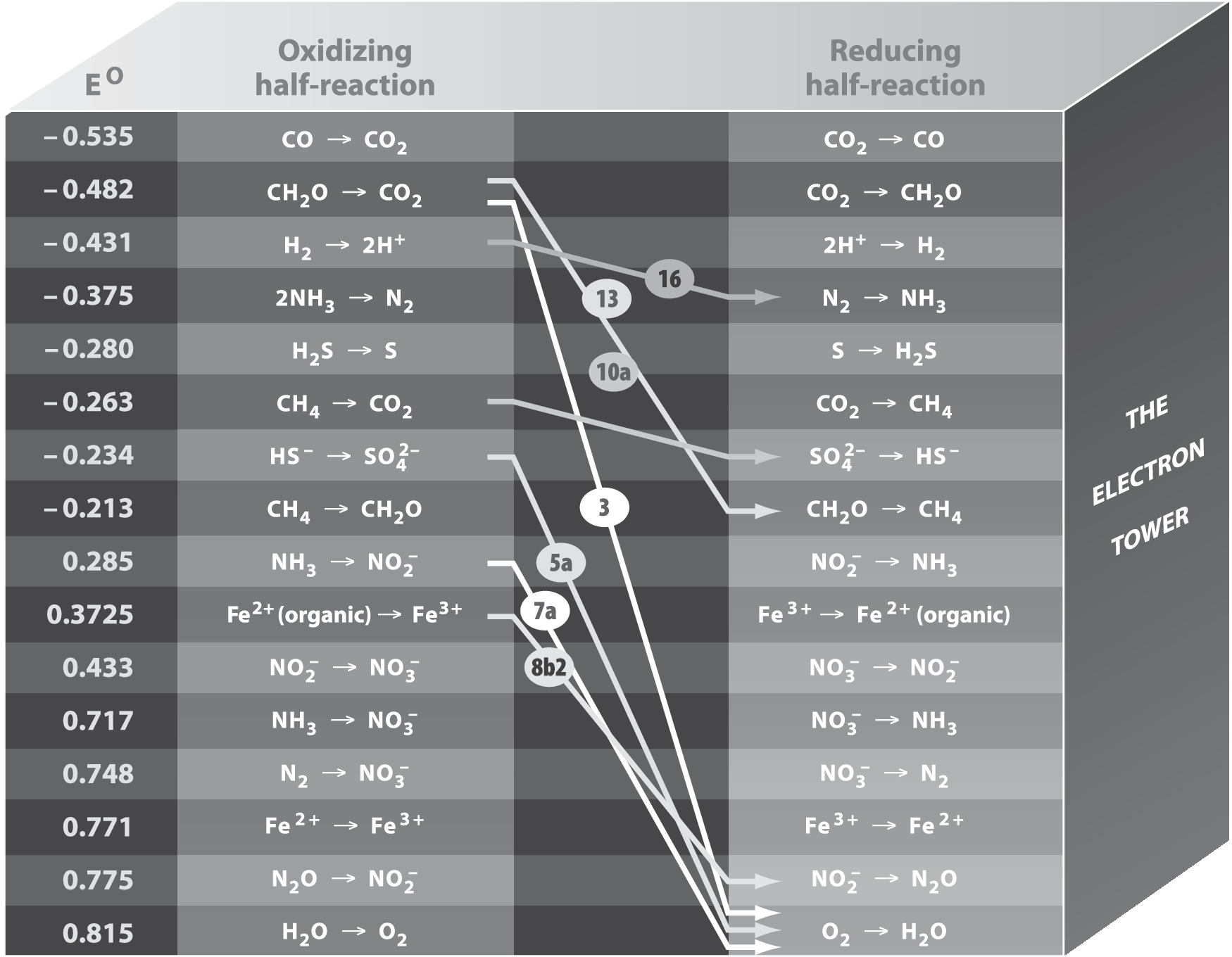
The electron tower diagram for selected microbial redox couples. Redox half-reactions are shown in order of their electrode potential (in volts) at pH 7.0, calculated from the standard electrode potentials for the reactions (Milazzo et al., 1978). Each reaction is shown as an oxidation half-reaction (in the left column) and as a reduction half-reaction (in the right column). If the oxidation of molecule A in the left column is above reduction of molecule B in the right column, then the oxidation of A can be coupled to the reduction of B yielding energy. Thus arrows drawn between half-reactions yield energy if they run downward from left to right.
Denitrification. Denitrification is the conversion of nitrate (
From left to right the chemical formulae are for the nitrate ion, the nitrite ion, nitrogen oxide, nitrous oxide, and nitrogen.
The reduction of these nitrogen compounds can be coupled to the oxidation of various inorganic fuels such as H2 and Fe2+, as well as to the oxidation of organic material such as degrading biomass in sediment. The nitrogen-reducing reactions can result in a variety of byproducts, including both aqueous species such as nitrite (
Nitrate (
The metabolic byproduct of denitrification is N2. Among the metabolic byproducts of denitrification, the gas N2 is naturally occurring and makes up 80% of our atmosphere by volume. It is therefore not a useful biosignature gas. In fact, it is the stability of N2 against destruction, rather than the total atmospheric amount, that is the critical problem for N2 as a biosignature gas. The nitrate (
Iron reduction (Fe3+). Iron reduction is a widespread metabolic capability of anaerobic bacteria, especially bacteria that live in Earth's subsurface. Iron reducers typically couple the oxidation of H2 or organic matter (simple compounds) to Fe3+ reduction. This Fe2+ produced generally ends up precipitating minerals such as metal sulfide (FeS) or the mixed Fe2+/Fe3+ mineral magnetite.
Sulfate and sulfur reducers. As discussed in the section on aerobic sulfur oxidation, the general sulfate and sulfur reduction processes include some molecules of the intermediate redox (in order of decreasing oxidation state),
Sulfate reduction can be coupled to inorganic compounds such as hydrogen, or organic compounds such as methane. For example, 90% of marine sediment methane is oxidized through anoxic mechanisms such as the reduction of sulfate (Strous and Jetten, 2004). The anaerobic sulfate-reducing metabolism is widespread around Earth in marine environments (but is restricted to narrow layers of anoxic sediments in the very narrow zone where there is both sulfate and anoxia), as sulfate is one of the major anions in ocean water (Watts, 2000). The metabolic byproduct of both sulfate and sulfur reducers is H2S, the rotten-egg odor gas. At pH>7, H2S will exist in water largely as HS− ions, which are not volatile. Below neutral pH (pH<7), H2S remains as uncharged molecules in water and can diffuse into the overlying atmosphere readily (Pasiuk-Bronikowska et al., 1992).
Methanogenesis and acetogenesis. Some methanogens produce methane (CH4) by using carbon from CO2. The electrons to reduce CO2 come from completely inorganic sources. In other cases, methane is produced from the metabolism of a range of organic compounds, including methylamines, acetate, and formate.
At the deep-sea floor, H2 is released from rocks by hot water emitted from hydrothermal vents (serpentinization). CO2 is available dissolved in water. In other environments, H2 is also produced as a byproduct of biological metabolism, and CO2 is available as gas in air or dissolved in water. The metabolic byproducts from methanogenesis are CH4 and H2O. The byproduct CH4 is also produced abiotically by volcanism.
Acetogenesis also reduces CO2 by using molecular hydrogen (Ljungdahl, 1986).
Although the chemistry and energetics are similar to methanogenesis, acetogenesis is not as relevant a generator of biosignature gases as is methanogenesis because the product, acetate ions, are very soluble in water. Only at pH<4.7 would acetate form uncharged acetic acid molecules, which are volatile enough to move to the atmosphere at low rates.
Reduction of metals and other inorganic compounds. Microbes can reduce a variety of metals under anaerobic conditions. One example is reduction of manganese from Mn4+ to Mn2+(Lovley and Phillips, 1988). The reduction potential of the Mn4+/Mn2+ couple is high. Other inorganic compounds used as electron acceptors include selenium and arsenic compounds, uranium, and even halogenated compounds (Madigan et al., 2003). All of these are represented in relatively low quantities and are probably not relevant from the astrophysical viewpoint of potential biosignature gases. Additionally, the products will be nonvolatile metal salts, and so will not be suitable biosignature gases even if they are produced in significant quantities.
Anoxic ammonium oxidation: anammox. Anammox is the anaerobic oxidation of ammonium (
Anammox is a special case of disproportionation 4 , a type of chemical reaction in which a single chemical species takes part in redox chemistry, with one part of the molecule being oxidized and one reduced (Lehninger, 1972). In the case of anammox, ammonium nitrate or ammonium nitrite is disproportionated, with the ammonium being oxidized and the nitrate or nitrite being reduced. This requires no external source or sink of electrons, so an organism generating energy from disproportionation is independent of geochemistry to provide or remove electrons.
Fermentation and disproportionation. Fermentation reactions disproportionate organic matter, which generates energy. The organic chemicals are almost always made by other life; fermenting organisms are usually heterotrophs. There is no need for disproportionation chemistry to generate particular types of end-products. For carbon chemistry, however, it is generally true that smaller, simpler molecules are more energetically favored (i.e., have lower free energy of formation) than large, complex molecules; and molecules with intermediate oxidation state are less favored than molecules that are fully oxidized or reduced (Amend and Shock, 2001; Bains and Seager, unpublished data). Thus fermentation biology tends to take large molecules of intermediate oxidation state and turn them into small, simple molecules that contain carbon that is largely or completely oxidized or reduced. Examples of potentially volatile output molecules are listed in Table 1.
In principle, any organism can produce any combination of one or more oxidized product and one or more reduced product from biomass (see textbook by Metzler, 1977).
An important fermentation example is methanogenic fermentation:
While methanogenic fermentation only yields one-eighth the energy per mole of organic matter as methane oxidation to CO2, it nevertheless provides enough energy to support flourishing microbial ecosystems and more energy per mole than the fermentation of sugars to produce alcohol and CO2 (Schink, 1997).
Some organisms can “ferment” nonbiological carbon if it is provided in an intermediate redox state. Carbon monoxide has carbon in the (−2) state and can be considered the anhydride of formic acid, which can be fermented (Lorowitz and Bryant (1984) according to the overall equations
Microorganisms can also disproportionate sulfur compounds of intermediate oxidation state (Finster, 2008), producing H2S and
4. Secondary Metabolism Byproducts
In this section, we list the major secondary metabolism byproduct classes, that is, the classes of secondary metabolism gases found at highest concentration in Earth's atmosphere. We emphasize the caveat that, unlike the products of primary metabolism, we cannot predict under what circumstances secondary metabolism byproducts might be produced on other worlds. Recall from Section 2 that primary metabolism is the metabolism that generates the energy and structural components for life; the net inputs are constrained by the environment. In contrast, we use secondary metabolism to refer to biochemical reactions that are specific to individual organisms, involve a wide variety of molecules, and are not traceable to environmental conditions.
A number of the molecules produced as byproducts to primary metabolism are also produced by secondary metabolism. These include CH4, NO, H2S, and CO. The secondary metabolism products NO, H2S, and CO are produced through different chemical routes than those used in primary metabolism. For example, NO is synthesized in human blood vessels from the amino acid asparagine via the specialized nitric oxide synthase enzyme family. In contrast, NO is produced in primary metabolism to generate energy by direct action of nitrate reductase on nitrite.
The most interesting aspect of secondary metabolism gas byproducts as a biosignature class is the much more diverse range of molecules as compared to molecules produced by primary metabolism. These diverse molecules may conveniently be divided into “inorganic” molecules (those which lack carbon-carbon bonds) and “organic molecules.” 5 Here, we describe molecules not apparently produced by primary metabolism.
4.1. Inorganic molecules
4.1.1. Sulfur compounds
A range of sulfur compounds are made as secondary metabolism gases on Earth. The major compounds released into the atmosphere are hydrogen sulfide (H2S), carbon disulfide (CS2), carbonyl sulfide (OCS, sometimes written as COS), dimethyl sulfide (DMS), and dimethyl sulfoxide (DMSO: CH3·SO·CH3). Estimates of the amounts released into the atmosphere vary widely (summarized in Table 2) because of uncertainties in translating global atmospheric concentrations into source fluxes. H2S is also a product of primary metabolism (Section 4.2). The sulfur secondary metabolism gases are produced by geological processes as well as biological ones, although in all cases the biological sources have higher fluxes (see references in Table 2). H2S, CS2, and OCS are products of the breakdown of organic material, usually bacteria or fungi, although plants can also release these volatiles. OCS can also be produced abiotically by atmospheric oxidation of CS2. DMSO is the product of oxidation of DMS; some microorganisms can generate DMSO from DMS (Taylor and Kiene, 1989), but nonbiological atmospheric chemistry can also oxidize DMS to DMSO, dimethyl sulfone (CH3·SO2·CH3), and methyl sulfate (CH3·SO3H). Other sulfur-containing compounds produced in small amounts that are made directly by life include methyl sulfide (also called methyl mercaptan: CH3SH), ethyl sulfide (C2H5SH), 2-thioethanol (HS·C2H4·OH), and dimethyl disulfide (CH3·SS·CH3) (Barbash and Reinhard, 1989). See also the work of Domagal-Goldman et al. (2011) for a discussion of sulfur compounds as biosignature gases.
Dimethyl sulfide is the largest biological source of atmospheric sulfur. DMS is produced by marine organisms. It is a breakdown product of the secondary metabolism byproduct dimethylsulfoniopropionate [DMSP: (CH3)2S+(CH2)2·CO2H], which some marine plankton accumulate to intracellular concentrations of up to 2 molar (Barbash and Reinhard, 1989). Why some marine phytoplankton accumulate DMSP is not known, but it may be for stress resistance. When the plankton are attacked by predators or viruses, or otherwise severely stressed, some of the DMSP is released into the ocean where enzymes break it down to DMS and propionic acid. Much of the DMS generated is consumed by other organisms, but some escapes to the atmosphere (Caron and Kramer, 1994).
4.1.2. Nitrogen compounds
Life produces a wide variety of oxides of nitrogen (Section 3) and nitrogen gas but does so primarily for energetic reasons (i.e., primary metabolism). For secondary metabolism, terrestrial organisms produce a wide range of amines, though amines generally have very low vapor pressure and are very soluble in water, so almost none get into the atmosphere in detectable amounts. (Amines are slightly more volatile in alkaline solutions about ∼pH=10.) Exceptions are ammonia and mono-, di-, and tri-methyl amine, which are produced by bacterial action on protein (Veciana-Nogues, 1997) and can sometimes be detected in the air. The most common and volatile of these, ammonia, is only present in Earth's atmosphere at low parts per billion level and is generated primarily by intensive animal farming (Timmer et al., 2005).
4.1.3. Inorganic gases other than sulfur or nitrogen compounds
Life is enormously chemically diverse and can manipulate almost any element found in its environment. In particular, metals and semimetals can be methylated to generate volatile methyl compounds; those known to be produced biologically include methyls of antimony [(CH3)2Sb], arsenic [(CH3)3As], bismuth [(CH3)3Bi], lead [(CH3)2Pb and (CH3)4Pb], mercury [(CH3)2Hg], and selenium [(CH3)2Se] (reviewed in Gadd, 1993; Bentley and Chasteen, 2002), which are all volatile. Even mammals, including humans, can transform arsenite (
Life also makes reactive oxygen species, including hydrogen peroxide (H2O2), through secondary metabolism, as well as several superoxides and peroxide radical compounds. These are made to attack other organisms; for example, human white blood cells can make peroxide ions to attack invading bacteria. In principle, hydrogen peroxide could be released into the atmosphere. Very little of these reactive and toxic molecules is made, however, and their low volatility and short half-life means that negligible amounts reach the air. Essentially, all the hydrogen peroxide, and other reactive oxygen species such as ozone, hydroxyl radicals and superoxide ions, and radicals, that can be detected in the atmosphere is the result of atmospheric photochemistry (Seinfeld and Pandis, 2006).
A final curious inorganic example we present is phosphine (PH3). Phosphine is too reduced to survive for long in Earth's oxidizing atmosphere, being rapidly oxidized to phosphorus oxides (Fluck, 1973). [It is claimed that biologically produced phosphine can spontaneously ignite in air (Burford and Bremner, 1972)]. Phosphine is generated in tiny amounts, which, coupled with its very rapid removal by oxidation, makes its concentration in Earth's atmosphere negligible [e.g., PH3 is present at levels up to 20 ppb above specific landfill sites (Glindemann et al., 2005)]. PH3 is produced under anaerobic conditions and is a measured source from solids and sludges. There is still debate about whether PH3 is generated directly by metabolism (bacterial reduction of phosphate-containing biomolecules) or is a geochemical side effect of metabolism [i.e., bacterial attack on iron particles in the ground which have traces of iron phosphide in them (Roels and Verstraete, 2001)].
Our last potential secondary metabolism biosignature example is not chemical at all, but electromagnetic. Some organisms produce light that is intense enough to glow visibly to human eyes (e.g., Ziegler and Baldwin, 1981; Shimomura, 2006). Detecting such a glow at stellar distances is clearly a task beyond any presently imagined instrument, but it is not inconceivable. The “milky sea” bioluminescence caused by concerted light emission by phytoplankton blooms has been detected from orbit (833 km altitude) in the Indian Ocean (Miller et al., 2005) and can be distinguished from thermal radiation or reflected sunlight by its unique spectral signature.
4.2. Organic molecules
Terrestrial life generates an enormous variety of carbon-based molecules via secondary metabolism. At least 50,000 different chemicals are made by plants alone (Pichersky and Gang, 2000). Many of these carbon-based molecules are volatile to some extent and enter the atmosphere as trace chemicals.
Most terrestrial organisms exude small, volatile carbon compounds (collectively termed volatile organic carbon—VOC). These include methane, methanol, ethanol, acetone, acetaldehyde, methyl ethyl ketone, methyl vinyl ketone, benzene, and toluene (Black et al., 2010, and references therein; see Table 3 for a selected list of VOCs and their formulae). Methane is also produced by primary metabolism via methanogenesis and methanogenic fermentation, as discussed in Section 3. Methane aside, methanol, acetaldehyde, acetone, and isoprene are released into the atmosphere at the highest global average flux (Warneke et al., 1999; Lewis et al., 2005; Sinha et al., 2007) and are made by bacteria, oceanic plankton, fungi, land plants, and animals.
4.2.1. Isoprene and terpenoids
A major class of VOCs by number are the terpenoids, named after a class of hydrocarbon molecule that plants build by joining two or more isoprene units together and then modifying the resulting molecule. Isoprene (2-methyl-1,3-butadiene) itself is a significant trace organic in the air around plants (Isidorov et al., 1985). Over 22,000 naturally occurring terpenoids are known (McGarvey and Croteau, 1995), and any of the lower-molecular-weight ones can be present in the air around plants. Terpenoids include flavor and fragrance compounds such as menthol and pinene (the sharp odor characteristic of pine forests), the characteristic odor components of oranges, lemons, and geraniums. Which specific terpenoid compound is present in an air sample depends on the plants nearby at the time, their state of nutrition, whether they have fungal or insect infections, and the time of day. It is not practical to try to predict which of these molecules will be present in a terrestrial air sample, let alone an exoplanetary one. The one commonality is that many plants release isoprene itself, although it is unclear why (Harley et al., 1999). It is worth noting that isoprene reacts with NOx to produce tropospheric ozone.
4.2.2. Halogenated organics
A bewildering diversity of organic molecules that contain one or more halogen atoms are made as a result of Earth-based biochemistry. It is often controversial whether a specific chemical is made directly by biochemistry or by subsequent geochemical or photochemical transformation of a biologically synthesized molecule. Methyl chloride and methyl bromide are the major halogenated hydrocarbons released on Earth. Methyl chloride is produced at levels of 3.5 Tg year−1 on Earth (Laturnus et al., 2002) and has been suggested as a potential biosignature gas for exoplanets (Segura et al., 2005). Other halogen compounds known include the carbon cores of methane, ethane, propane, and propene substituted with one or more atoms of chlorine, bromine, or iodine (Scarratt and Moore, 1998; Moore and Groszko, 1999; Laturnus et al., 2002). Mixed halides (e.g., CHClBrI) are also biologically generated.
There are few organofluorine compounds made by life. Almost all the organofluorine compounds in the atmosphere are believed to come from human industrial activity, and most of the 12 organofluorine molecules known to be biosynthesized are not volatile (O'Hagan et al., 2002). Only fluoroacetone would enter the atmosphere to any detectable extent. As far as we know it has not been detected in air, and in any case it is not clear whether fluoroacetone is a true metabolic byproduct or a photochemical breakdown product of fluoroacetate.
5. Discussion and Summary
5.1. Redox disequilibrium versus chemical disequilibrium
The canonical concept for the search for atmospheric biosignature gases is to find an atmosphere severely out of thermochemical redox equilibrium (Lederberg, 1965; Lovelock, 1965). This is because abiotic processes are unlikely to sustain a redox disequilibrium. Indeed, Earth's atmosphere has oxygen (a highly oxidized species) and methane (a very reduced species) several orders of magnitude out of thermochemical redox equilibrium. In practice, it could be difficult to detect an atmosphere out of thermochemical redox equilibrium. The Earth as an exoplanet, for example (Fig. 1), has a relatively prominent oxygen absorption feature at 0.76 μm, whereas methane at present-day levels of 1.6 ppm has only extremely weak spectral features. Before Earth was 2.5 billion years old, CH4 may have been present at much higher levels (1000 ppm or even 1%), as possibly produced by widespread methanogenic bacteria (Haqq-Misra et al., 2008, and references therein). Such high CH4 concentrations would be easier to detect, but since Earth had very low oxygen levels (<0.001%: Bekker et al., 2004), the co-existence of O2 and CH4 would not be detectable. Segura et al. (2005) showed that oxygen at 0.2% would be detectable at visible wavelengths, and at 0.02% it would be detectable at IR wavelengths via the O3 absorption feature.
The more realistic atmospheric biosignature gas is a single gas completely out of thermochemical equilibrium with the other atmosphere (and/or surface) constituents. Earth's example again is oxygen or ozone, which are more than 5–10 orders of magnitude higher than expected from equilibrium chemistry on a terrestrial planetary troposphere (i.e., lower atmosphere), depending on the CO2 mixing ratio and the altitude (Kasting and Catling, 2003). O2 and O3 have no known abiotic production at such high levels. The challenge with a single biosignature gas outside the context of redox chemistry becomes one of false positives. To avoid false positives, one must look at the whole atmospheric context. For example, a high atmospheric oxygen content might indicate a planet undergoing a runaway greenhouse with evaporating oceans. When water vapor in the atmosphere is being photodissociated with H escaping to space, O2 will build up in the atmosphere for a short period of time. In this case, O2 can be associated with a runaway greenhouse via very saturated water vapor features, since the atmosphere would almost certainly be filled with water vapor at all altitudes. Other O2 and O3 false positive scenarios (Selsis et al., 2002) are discussed and countered by Segura et al. (2007).
We now continue on to the case of ideal biosignature gases in the context of a single metabolic byproduct gas—one that is orders of magnitude higher than in the chemical equilibrium case.
5.2. The fate of Earth's primary metabolism inorganic byproducts
The same molecules appear again and again as reactants and products in the set of 14 categories of metabolic reactions described in Section 3. Why? The common molecules are those that are readily available; they are made of the abundant elements in Earth's atmosphere, ocean, and subsurface, and formed by chemistry that can happen spontaneously at the temperatures and pressures of Earth's atmosphere, oceans, or upper crust. It makes sense, therefore, that the same molecules produced and cycled by life are also those produced and recycled in geophysical processes.
The ideal biosignature gas is one that (1) does not exist naturally in the planetary atmosphere at ambient temperatures and pressures, (2) is not created by geophysical or geochemical processes, (3) is not produced (or rapidly destroyed) by photochemistry, (4) has a strong spectral signature.
See Fig. 3 for a list of gases from energy-yielding redox reactions described in the four-point framework.

Earth-based inorganic metabolic byproduct pyramid. The pyramid description emphasizes that only a few of Earth's biosignatures can robustly be attributed to life.
Sources and sinks are critical to the formation of biosignature gases. Indeed, points 2 and 3 above are another way of stating that the ideal biosignature gas has no significantly competing sources. Sinks influence whether a biosignature gas can actually accumulate in a planetary atmosphere to detectable levels. Potential biosignature gas sinks include reaction with other molecules and photochemical destruction (and in some cases reactions with surface constituents). Biosignature sinks will vary from exoplanet to exoplanet because the varying atmospheric chemistry depends on the particular planet's atmosphere and the differing UV radiation from different star types. Abiotic sources will also likely vary from planet to planet because surface composition and volcanic outgassing (related to crustal composition) may also vary. We leave a treatment of Earth-based metabolic byproducts in different exoplanetary environments with non-Earth-like sources and sinks for a separate paper.
Regarding point 4, what makes a strong overall spectral signature depends not only on the concentration of a gas but also on the gas absorption cross section. The absorption cross section itself originates from quantum mechanical interactions of molecules and radiation. In general, the greater the difference among the atomic weights of a molecule's constituent atoms, the stronger spectral signature that will result. Homogeneous diatomic molecules (e.g., N2 and O2) have no permanent dipole moment and weak to no IR rotational-vibrational spectral features, and their electronic transitions are usually at UV wavelengths that are hard to access by telescopes. O2 is a fortuitous exception with electronic transitions at visible wavelengths. See the work of Des Marais et al. (2002) for examples of spectra of Earth's atmosphere with different gas concentrations.
Earth's most robust biosignature gas O2 satisfies all four of the above “ideal biosignature gas” criteria. The gas N2O satisfies the first three. While O2 and N2O are also generated by photochemistry (from photolysis of H2O and lightning fixation of N2), the amounts are miniscule. The major sink of both O2 and N2O is destruction by photolysis.
Absent a unique biosignature gas like O2 and N2O on other planets, we must consider the more common gases produced as metabolic byproducts. To understand their potential as biosignature gases, we must consider sources and sinks. Regarding sources, the potential biosignature gas must be much more abundant than can reasonably be produced by abiotic processes. Any biosignature gas sinks must be small enough to enable the biosignature gas to accumulate to detectable levels. In other words, the main criteria for a biosignature gas is that the gas exists in such great quantities that its presence in a planetary atmosphere is well above the amounts that could be produced by the abiotic processes of geology or photochemistry.
We now turn to a discussion of Earth's inorganic metabolic byproducts, including their sources and sinks. The list of inorganic gaseous metabolic byproducts generated from the microbial reactions described in Section 3 are O2, H2, CO2, N2, N2O, NO, NO2, H2S, SO2, CH4, H2O.
The gases O2, (and its photolytic product O3), N2O, and CH4 have been widely discussed in the literature as biosignature gases on planets with Earth-like composition atmospheres (Leger et al., 1993; Des Marais et al., 2002; and references therein). O2 and its photolytic product O3 are the most reliable biosignature gas indicators for life as we know it. O2 is highly reactive and therefore will remain in significant quantities in the atmosphere only if it is continually produced. The major sink of O2 and O3 is surface reactions. There are no known abiological mechanisms that can continually produce large quantities of O2 (Venus and Mars both have very small amounts of O2). Only rare O2 false positives can likely be ruled out by other planetary characteristics (Segura et al., 2007). O3 is a nonlinear indicator of O2; only a small amount of O2 need be present to produce a relatively large quantity of O3 (Leger et al., 1993).
N2O is produced by life—albeit in small quantities—during microbial oxidation-reduction reactions. N2O has a very weak spectroscopic signature. The main N2O sink is destruction by photolysis. Furthermore, because the N2O spectroscopic signature overlaps with H2O absorption bands, it may be more easily detectable on a planet with less water vapor than Earth has.
High concentrations of CH4, such as may have been present on early Earth, could indicate the presence of methanogenic bacteria but could also be from volcanism. The main sink of CH4 on Earth is by reaction with the ·OH radical, itself produced from H2O photolysis. The gases CH4 and N2O could accumulate to higher abundance in a planet whose host star has a much lower UV-radiation output than the Sun's. CH4 and N2O will also accumulate to a higher abundance in an atmosphere with less water vapor (Segura et al., 2005).
It is useful to review why most, if not all, metabolic byproducts are not useful as biosignature gases when Earth is viewed from afar. We will start with gases that are already natural components of Earth's atmosphere. On Earth, the gas N2 makes up 80% of our atmosphere and is essentially primordial (derived from geological sources during planet formation) and therefore not a useful biosignature gas. Furthermore, as a homo-nuclear molecule N2 has no rotational-vibrational transitions and hence no prominent spectral signature in the visible and IR. Like N2, water is already present on Earth in the oceans and atmosphere. As a biosignature gas, H2O is not useful. While not a biosignature, liquid water is a habitability indicator, because all known life needs liquid water.
Like N2 and H2O, CO2 is also already present in Earth's atmosphere and makes up about 0.038% by volume. For Venus and Mars (the only other solar system terrestrial planets with an atmosphere), CO2 makes up more than 97% of their atmosphere. CO2 originates from outgassing and is expected to be present on all small, rocky planets. As such, CO2 is considered a major planetary atmosphere gas that results from planet formation and evolution; hence it is not clearly viable as a biosignature gas.
Out of Earth-based metabolic byproduct gases that are not already part of Earth's atmosphere or one of the three established biosignature gases (O2, N2O, CH4), we are further left with the following as possible biosignature gases to consider: H2, H2S, SO2, NO, and NO2. The gases H2, H2S, SO2, NO, and NO2 are not useful biosignature gases as they are not uniquely biotic in origin and would be difficult to detect from afar in Earth's concentrations. These gases could perhaps be prominent under different exoplanetary conditions of sources and sinks.
H2 is an interesting gas to consider. Earth's atmosphere has negligible H2 because Earth's surface gravity is not strong enough to hold on to hydrogen outgassed early in Earth's history, given the temperature at the top of Earth's atmosphere (i.e., exosphere). On present-day Earth, the abiotic source of H2 is volcanism, and the net sink is atmospheric escape. H2 is tricky as an exoplanetary biosignature gas, because on some planets H2 is expected to be naturally occurring, with no real sinks. Such planets would have a higher surface gravity and a lower exospheric temperature or weaker stellar wind to contend with than Earth does. It would be difficult to ascertain whether a super-Earth with H2 had properties such that it could retain H2 or whether H2 were a biosignature gas. Atmospheric escape is a complicated calculation because many critical parameters (such as the star's present and past extreme UV radiation, the presence or absence of a planetary magnetic field, and the original and present-day mass and composition of the planetary atmosphere) are not measurable, even for planets where the mass, radius, and surface gravity can be determined.
On Earth, SO2 is not a useful biosignature gas for two reasons. SO2 is unstable in Earth's atmosphere because it is quickly oxidized by the ·OH radical. ·OH is generated from H2O vapor after H2O is split apart by solar UV radiation. In the presence of water, SO2 combined with ·OH leads to hydrosulfuric acid, which rains out as acid rain. SO2 can also dissolve directly in liquid water droplets in clouds to form sulfurous acid. In a planet with less atmospheric water vapor than Earth's, SO2 might accumulate in the atmosphere even under abiotic processes, namely, volcanism (Kaltenegger and Sasselov, 2010). Lastly, SO2 is not a net product of terrestrial metabolism but exists only as an intermediary in metabolic redox reactions.
On Earth, H2S is not a biosignature gas because Earth's volcanoes produce significant amounts of H2S. Furthermore, H2S is very rapidly oxidized by nonbiological processes to sulfate by atmospheric oxygen; thus it does not accumulate (Watts, 2000). Phototrophic and chemoautotrophic sulfur bacteria can also be sinks of H2S. H2S is a major end-product of terrestrial metabolism. Ground-level biological sources of H2S are sulfate-reducing organisms. Relevant for exoplanets is that any H2S that makes it into Earth's atmosphere is photolyzed or readily oxidized by atomic O, which is produced from photodissociation of H2O. H2S, produced in high-enough quantities, is therefore a potential biosignature gas on an exoplanet with an oxidized atmosphere and suitable atmospheric chemistry and UV flux to allow it to survive long enough to be detected. H2S would have to appear in abundances many orders of magnitude above that plausibly generated from volcanic emission.
The atmospheric gas NO is generated from N2 naturally by lightning—an enormous amount of energy is needed to break the N2 triple bond. NO also originates as a product of the atmospheric oxidation of organic chemicals, particularly terpenoids and isoprene (and also as anthropogenically produced air pollutants). A major oxidation path for isoprene and terpenoids is via photooxidation, as they are efficient absorbers of UV light. Photooxidation generates free radical species, which react with atmospheric O2 and H2O to generate ·OH radicals, which in turn react with atmospheric nitrogen to generate NOx species.
Small amounts of nitric oxides and nitric acid are also made in volcanoes (Mather et al., 2004). NO is destroyed by oxidation, primarily by O3 and ·OH, respectively, and subsequently dissolves in rainwater to reach Earth as nitrates. The nitrate (
5.3. The fate of Earth's secondary metabolism byproducts
We now turn to a discussion of the sources and sinks of selected secondary metabolic byproducts (see Section 4). We are motivated to discuss secondary metabolic byproducts because the number of primary metabolism byproducts is limited. The overwhelming sources on Earth for almost all Earth's secondary metabolism byproducts are life—either secondary metabolism processes or human industry.
Dimethyl sulfide. On Earth, the majority of DMS is metabolized by marine organisms to sulfide (in anoxic environments) or to sulfate (in oxygenated environments) (Kiene and Bates, 1990). The DMS that escapes is primarily destroyed by attack by atmospheric hydroxyl radicals, which are the product of photochemistry (Watts, 2000; Lomans et al., 2002). The chain of sulfur products is thought to follow progressive oxidation (Table 4).
Dimethyl sulfoxide, dimethyl sulfone, and methyl sulfate are all detectable in the atmosphere. Sulfuric acid is rained out. The atmospheric fate of the carbon atoms in DMS is to be oxidized by O3 and ·OH to CO and CO2. The above oxidation is fast in Earth's atmosphere. For a thorough atmospheric chemistry discussion of DMS, including breakdown to other molecules, see the work of Seinfeld and Pandis (2006). For DMS in the context of exoplanetary Earth-like atmospheres under a variety of stellar UV radiation see the work of Domagal-Goldman et al. (2011).
Volatile organic carbon. A fraction of the VOC released into the atmosphere is re-absorbed by plants or into the soil—the fraction depends heavily on the nature of the VOC species, the plants growing, the soil, and other factors like wind speed and rainfall. An “average” fraction absorbed is not a meaningful number. VOCs that are not absorbed are photolyzed and oxidized. Oxidation of methane by ·OH or O3 is relatively slow, and methane is transported to the stratosphere where it is oxidized by UV photochemistry. Oxidation of alkenes such as isoprene by ·OH or O3 is fast and occurs in the troposphere (reviewed in Calvert et al., 2008). Photoactivation of alkene VOCs is also efficient, as compounds containing carbon-carbon double bonds, and especially those containing multiple carbon-carbon double bonds in conjugated systems 6 , have high absorbances in the UV even at wavelengths that penetrate to ground level (280–300 nm). The products of VOC oxidation depend on the chemistry of the compounds. VOC photooxidation of alkanes (the hydrocarbons in gasoline and diesel fuel) results in efficient formation of peroxides and oxy-radicals, which stimulate formation of hydroperoxide radicals and NO2, which themselves lead to ozone formation. The partially oxidized hydrocarbons aggregate into aerosol particles, which ultimately are completely oxidized to CO2 or rained out (e.g., Finlayson-Pitts, 2010, and references therein). Biogenic VOCs react very fast with ozone and ·OH but do not generate the same smog (Lelieveld et al., 2008).
Methyl chloride is also oxidized by ·OH and O3, as are other partially substituted halocarbons (Cox et al., 1967). However, fully halogenated compounds such as carbon tetrachloride or the synthetic freon compounds are resistant to tropospheric oxidation, and some have half-lives of hundreds or thousands of years (Calvert et al., 2008).
5.4. Solid metabolic biosignatures
Some metabolic byproducts are in solid rather than gas form. One interesting microbial metabolic byproduct is solid sulfur. Sulfur bacteria oxidize H2S to elemental solid sulfur, and the sulfur granules are deposited either inside or outside the cell (depending on the type of sulfur bacteria). The sulfur granules can be reused by the same microbes for further energy extraction upon oxidation of sulfur to sulfate. Hence, solid sulfur may not be a viable biosignature, even it were widespread on Earth's surface. Under the right conditions, solid sulfur may be a potential biosignature but can also be generated abiotically from volcanism, as on Io.
A second solid metabolic byproduct is rust, in the form of FeO(OH). Some studies have provided evidence that large-scale iron deposits in Archean Earth known as banded iron formations were formed by anoxygenic, phototrophic iron-oxidizing bacteria (Kappler et al., 2005). There is significant debate, however, about formation mechanisms for banded iron formations.
From a biosignature perspective, it is interesting to consider whether the accumulation of large quantities of solid compounds (e.g., elemental sulfur or ferric iron) on the surface of a planet or released episodically from the subsurface could ever be detected by remote studies of exoplanets, and additionally whether the solid compounds could ever be robustly interpreted as a biosignature.
5.5. Inverse biosignature gases
The absence of a chemical can also be a biosignature, although it is often considered inferior to the presence of a biosignature gas because absence of evidence is not strong evidence of absence.
The ideal inverse-biosignature gas is one that is absent in spite of being (1) robustly predicted to exist naturally in the planetary atmosphere at ambient temperatures and pressures; (2) possibly created by geophysical, geochemical, or photochemical processes; (3) not rapidly destroyed by photochemistry; (4) known to have a strong spectral signature.
Lower-than-expected levels of acetylene (Coustenis et al., 2010) and apparent flux of hydrogen into the surface (Strobel, 2010) have been found in the atmosphere of Titan and have been suggested as an inverse biosignature. The reduction of acetylene by hydrogen (McKay and Smith, 2005) or its hydrolysis by water “ice” (Bains, 2004) could be sources of energy for life, and their observed absence is therefore considered to be a sign of life. The idea that Titan has surface-dwelling life, however, is highly speculative, despite the apparent inverse biosignature result.
5.6. Beyond energy-yielding inorganic metabolic reactions
5.6.1. Nitrogen fixation
On Earth, there is a metabolic pathway that is energetically unfavorable but necessary to produce a biologically important molecule: nitrogen fixation. Nitrogen is required for all life on Earth because it is an essential component of many important biological molecules (such as amino acids and proteins). Yet most organisms cannot break the strong N2 triple bond. Because NH3 is a form of nitrogen more easily used by many different organisms, nitrogen fixation is a critical chemical pathway. The ability to fix nitrogen evolved early in the history of Earth's life and is widespread among prokaryotes (bacteria and archaea) (Raymond et al., 2004). We emphasize that the nitrogen fixation reaction does not yield energy and therefore does not fit into our focus on chemical reactions that generate energy. Nevertheless because of its importance for making N available to all life we include it here. Nitrogen fixation is the process that converts nitrogen (N2) into ammonia (NH3).
The byproduct NH3 is a good example of a difficulty in trying to predict biosignature gases based on metabolic pathways. Because fixed N is so scarce, NH3 is rapidly taken up by nearby microorganisms and converted into biological molecules.
5.6.2. The challenge for secondary metabolism gas byproducts
Earth's atmosphere contains small concentrations of a number of gases that are generated by life, but not as the products of primary metabolism (Section 4). Life's choice of particular secondary metabolism biosignature molecules seems quite arbitrary. In contrast to redox-generated biosignatures, which can be put into a quantitative framework, it is often unknown why one type of nonmetabolic biosignature molecule is produced by one organism and not another. A few described in Section 4 include OCS, CS2, PH3, CH3Cl, CH3Br, DMS, and isoprene compounds as an example of vegetation biogenic hydrocarbons.
5.6.3. Photosynthetic pigments as biosignatures
The one widely discussed biomass biosignature is related to photosynthetic pigments. A critical component of the ecology of photosynthetic microorganisms is the highly evolved pigments that they contain to harvest light of specific wavelength. Each group of photosynthetic organisms has been shown to possess photopigments adapted to their specific light regime (e.g., Seager et al., 2005; Kiang et al., 2007). The chlorophyll of phytoplankton and green plants can be seen from Earth-observing satellites. Unfortunately, the change in brightness is small for a spatially unresolved exoplanet, especially when considering the low reflectivity of Earth's oceans and the small area of plankton blooms compared to a large part of a hemisphere (Seager et al., 2005).
Vegetation exhibits a more promising “red edge” spectral feature (e.g., Woolf et al., 2002). The reflection spectrum of photosynthetic vegetation has a dramatic sudden rise in albedo from around 700 nm to a plateau between 720 and 760 nm by almost an order of magnitude. This strong reflection feature, known as the red edge, originates from near-IR scattering. The near-IR scattering itself occurs due to the lack of absorbing pigments and the index of refraction between leaf cell walls and air spaces inside the leaf. Some have speculated that vegetation evolved the red edge as part of a temperature control mechanism (Gates et al., 1965) in that energy is not absorbed at all wavelengths. Perhaps this cooling mechanism evolved to prevent overheating, which would cause chlorophyll and proteins to degrade. On Earth, the red edge signature is probably reduced to a few percent due to clouds (see, e.g., Woolf et al., 2002; Seager et al., 2005; Montanes-Rodriguez et al., 2006; and references therein), but such a spectral surface feature could be much stronger on a planet with a lower cloud-cover fraction.
5.7. Summary
We have reviewed Earth-based metabolic byproducts (Table 5), separating our description into two key categories. The first is primary metabolism byproducts created by energy extraction from redox reactions. The redox reactions of primary metabolism include energy capture and biomass building, and the redox reactions are directly constrained by environmental chemistry. The second general category is secondary metabolism byproducts that are specific to individual organisms and not associated with energy capture or biomass building, hence are not quantitatively constrained by environmental chemistry.
On Earth, O2 and N2O are the unique biosignature gases. The other dozen or so primary metabolism gas byproducts are either short-lived in the atmosphere or also produced in dominant amounts by geophysical processes. We also reviewed a number of different classes of secondary metabolism byproducts, emphasizing that life's choice of particular secondary metabolism biosignature molecules seems quite arbitrary, that is, the purpose of many of them as well as biosynthetic pathways are still poorly understood. For both classes of biosignature gases, one hopes that different metabolic byproduct gases might accumulate into biosignatures on exoplanets with conditions different from Earth's, yielding more possibilities for biosignatures than terrestrial precedent allows. The overall idea is that we may find that unusual ratios of gases are the most practical way to identify possible signs of life on other worlds, albeit not with 100% certainty. Determining which gases and in which ratios is anticipated to be a profitable research field for the future.
A. Appendix: Redox Reactions
This appendix provides an introduction to and review of redox reactions. This is a primer for an audience not familiar with the terminology of the field. More detailed material can be found in many textbooks (e.g., Alberts, 2008; Metzler, 1977; Madigan et al., 2003) for more background on this and other aspects of biochemistry.
In particular, the redox potentials, Gibbs free energies, and other measures discussed below refer to reactions done at 1 molar concentration (one mole, or 6.023×1023 molecules, per liter), at standard temperature, pressure, and an appropriate pH. None of these conditions holds exactly in terrestrial life, and some, such as pH and concentration, vary widely. Thus the numerical calculation of whether a reaction will occur and what energy might be released by it depends on knowing not just the chemistry but the environment in which the reaction happens.
Redox is short for “reduction and oxidation” chemistry. The terms reduction and oxidation have deep historical roots in chemistry but have come to mean a class of reactions in which electrons are transferred between ions or molecules. A simple redox reaction would be the reaction of sodium metal and chlorine gas to make sodium chloride
The sodium atoms started out uncharged and ended up as positive sodium ions. The chlorine atoms started out uncharged and ended up as negative chloride ions. One electron was transferred between each sodium atom and each chlorine atom. The sodium atom is said to be the electron donor and the chlorine molecule the electron acceptor. Oxidation is the loss of electrons, reduction is the gain of electrons, so sodium is said to be oxidized in this reaction, and chlorine reduced. Chlorine is the oxidant, and sodium is the reductant.
The reaction between sodium and chlorine occurs because sodium atoms have a lower affinity for an electron than chlorine atoms. The tendency for an atom or ion to lose an electron in a reaction in water can be measured directly. Conceptually, we separate the reaction above into two half-reactions
We measure the tendency of each reaction to happen. In essence, we make an electrode out of the chemical concerned, dip it into water, and see what negative charge it accumulates due to its attraction of electrons. This gives us an electrode potential for that element or compound. In reality, the experiment cannot be done in this way for four reasons. First, electrons are not available in water, so a reaction that transfers electrons (as in the reaction between sodium and chlorine above) needs to be studied. Second, to measure voltage a reference electrode needs to be provided, and the elements of that will themselves have an electrode potential, so all electrode potentials are relative. Third, many compounds react with water, are not soluble, or present other technical problems; so the electrode potential must be inferred. Last, as with any chemical equilibrium, the concentration of the solutions involved can alter the balance of the reaction, including the balance of electrons transferred and hence the voltage. A series of indirect experiments, however, can be done to deduce the standard electrode potential (E 0), in units of volts, of a compound or element. E 0 is a measure of a molecule's tendency to gain or lose electrons when reacting with other compounds in water under standard conditions of concentration, temperature, and pressure, compared to the tendency of hydrogen gas to form hydrogen ions under the same conditions (the reference, with E 0 defined as 0).
The electron tower is a graphical representation of this standard electrode potential. In the electron tower diagram, a series of half-reactions are shown, in each case reducing the compound concerned. To be balanced, each has to be matched with a reaction oxidizing a compound. An oxidation reaction is the reverse of a reduction reaction. The standard electrode potential of each half-reaction is shown on the vertical scale. If we link two reactions, then the voltage generated by our conceptual cell will be the voltage of the oxidation reaction minus the voltage of the reduction reaction.
The NaCl example is a good redox example because the electrons are really transferred from Na to Cl. Under most redox reactions, electrons are not fully transferred between atoms in a given molecule and so may not end up having charges like the sodium and chlorine atoms above. Instead, the atoms get partial charges as the electrons are shared unevenly between atoms in the molecule.
Under this scenario, the voltage is a result of the affinity of the atoms concerned for electrons. The voltage is related to the energy released when the atom gains an electron or energy needed to remove an electron from an atom. In other words, the voltage is related to the energy released by the reaction and is described by the Nernst Equation, which relates the Gibbs free energy to the electrochemical voltage by
Here, ΔG is the change in Gibbs free energy, n is the number of electrons involved in the reaction, and F is Faraday's Constant.
Gibbs free energy is a measure of the energy available to do work when a chemical reaction occurs under standard conditions. The Gibbs free energy is measured in kilojoules per mole. (A mole is 6.023×1023 molecules. Molar is a measure of concentration and is one mole per liter.) We note that E 0 changes with temperature and ΔG changes with concentration of the reactants, themselves sensitive to pressure and pH (see, e.g., Amend and Shock, 2001).
The electron tower (Fig. 2) can be used to determine whether any reaction will yield energy or require it. In general, if the oxidative half-reaction of molecule A is above reductive reaction of molecule B in the tower, then molecule A will reduce molecule B, releasing energy. Thus, we can see that reduction of Fe3+ to Fe2+ will drive oxidation of H2S to elemental sulfur. The reverse is the reduction of sulfur to H2S, which, for one example, can be coupled to oxidation of organic matter (here, methane) to form carbon dioxide. Both of these are reactions used by bacteria, as listed in Section 3. As another example, the reduction of molecular oxygen to water, when coupled with the oxidation of almost any other species in the tower, will yield a lot of energy, which is why oxygen is such a good “oxidizer” of organic and inorganic fuel.
Footnotes
Acknowledgments
We thank George Cody, Feng Tian, and Renyu Hu for useful discussions. This work was in part supported by a grant from Foundational Questions in Physics and Cosmology (FQXI).
Author Disclosure Statement
No competing financial interests exist for any of the authors.
Abbreviations
DMS, dimethyl sulfide; DMSO, dimethyl sulfoxide; DMSP, dimethylsulfoniopropionate; OCS, carbonyl sulfide; VOC, volatile organic carbon.
1
We define an Earth analog as an Earth-mass, Earth-size planet in an Earth-like orbit about a Sun-like star.
2
The term “fixation” means converting inorganic, environmental carbon or nitrogen into a form that life can use to build biological molecules. Specifically, photosynthesis reduces environmental carbon dioxide (CO2) into carbohydrate, and nitrogen fixation (see ![]() ) reduces atmospheric nitrogen gas (N2) into ammonia (NH3). Carbohydrates are sugars, such as glucose, or polymers of sugars, such as starch and cellulose, and can be considered to have the general chemical formula (CH2O)
n
. (Note that carbohydrates are quite different from hydrocarbons, such as methane or octane, which have the general formula C
n
H
m
, where m≥n.)
) reduces atmospheric nitrogen gas (N2) into ammonia (NH3). Carbohydrates are sugars, such as glucose, or polymers of sugars, such as starch and cellulose, and can be considered to have the general chemical formula (CH2O)
n
. (Note that carbohydrates are quite different from hydrocarbons, such as methane or octane, which have the general formula C
n
H
m
, where m≥n.)
4
Disproportionation is any chemical reaction of the type A+A→A′+A″, where A, A′, and A″ are different chemical species. In the case of redox reactions, molecules of one chemical species (A) react with each other to form both an oxidized product (A′) and a reduced product (A″).
5
We note that there are several definitions of “organic” molecules, and that according to different authors “organic” molecules are variously those that contain carbon-carbon bonds (as here), carbon-hydrogen bonds, or carbon in discrete molecules. These distinctions are not important, as our purpose is a convenient classification only, so we adopt the definition of “organic” as “having carbon-carbon bonds” and admit that methane is an anomaly in having no carbon-carbon bonds, though it is almost universally classified as “organic.”
6
A conjugated double bond system is one where two (or more) double bonds are molecular neighbors in a structure such as H2C=CH–CH=CH2. Electrons are shared across both double bonds in such structures. Where such structures form rings, such as in benzene, electrons are shared around the ring, and the compounds are called “aromatic,” a meaning distinct from the conventional meaning of “aromatic” as “having a smell.”