
Abstract
The presence of nonprotein α-dialkyl-amino acids such as α-aminoisobutyric acid (α-AIB) and isovaline (Iva), which are considered to be relatively rare in the terrestrial biosphere, has long been used as an indication of the indigeneity of meteoritic amino acids. However, recent work showing the presence of α-AIB and Iva in peptides produced by a widespread group of filamentous fungi indicates the possibility of a terrestrial biotic source for the α-AIB observed in some meteorites. We measured the amino acid distribution and stable carbon and nitrogen isotopic composition of four α-AIB-containing fungal peptides and compared this data to similar meteoritic measurements. We show that the relatively simple distribution of the C4 and C5 amino acids in fungal peptides is distinct from the complex distribution observed in many carbonaceous chondrites. We also identify potentially diagnostic relationships between the stable isotopic compositions of pairs of amino acids from the fungal peptides that may aid in ruling out fungal contamination as a source of meteoritic amino acids. Key Words: Filamentous fungi—Amino acids—α-aminoisobutyric acid—Contamination—Isotopes. Astrobiology 11, 123–133.
1. Introduction
T
Most meteorites studied in the laboratory are “finds,” and for the majority of these samples little is known about the length of time they have resided on Earth, during which they were exposed to terrestrial environments (Jull, 2006). Even samples recovered as observed meteoritic “falls,” in which meteorite fragments are retrieved relatively quickly (days to weeks), are still exposed to potentially contaminating terrestrial environments from their landing site or during collection and curation. For example, extensive terrestrial amino acid contamination from the impact site environment has made it difficult to determine whether indigenous amino acids are present in the Allan Hills (ALH) 84001 and Nakhla martian meteorites (Bada et al., 1998; Glavin et al., 1999), and the amino acids initially observed in the Tagish Lake meteorite appear to be primarily the result of terrestrial contamination from the lake ice meltwater (Kminek et al., 2002). In addition, infiltration by terrestrial microbiota is a potential problem. For example, the Nakhla martian meteorite, which fell in 1911 and was collected within a month of the fall, has been shown to be contaminated with terrestrial microbiota and terrestrial amino acids (Glavin et al., 1999; Toporski and Steele, 2007). This contamination includes infiltration by filamentous organisms, potentially fungi or bacteria (actinomycetes). The carbonaceous chondrite Orgueil also contains terrestrial contamination of its free organic components (Watson et al., 2003). Environmental samples from the fall or find sites, such as ice, soil, or sediments, can help to elucidate the potential contamination, but these samples are not always made available for analysis.
There are several approaches for determining a terrestrial or extraterrestrial origin of meteoritic organics. Among these are (1) comparing meteoritic molecular distributions and abundances with those found in the biosphere; (2) using stable isotope ratios to search for terrestrial or extraterrestrial signatures; (3) analyzing samples from environments in which the meteorite has resided on Earth (e.g., soil from the fall site); and (4) determining enantiomeric ratios of chiral compounds such as amino acids.
Amino acids are particularly interesting meteoritic compounds, as they are ubiquitous molecules of life as the monomers of proteins. The suite of identified meteoritic amino acids is far more diverse than the 20 α-hydrogen protein amino acids widely found in the terrestrial biosphere, with over 80 amino acids identified in the Murchison meteorite alone (see Martins and Sephton, 2009 for a review). Many of these amino acids are either rare or unknown in the biosphere, and many of the chiral amino acids in the meteorite were found to be racemic (Kvenvolden et al., 1971). Terrestrial biology utilizes the L-enantiomer of amino acids preferentially but not exclusively (Martínez-Rodríguez et al., 2010), while abiotic formation processes in the absence of a chiral influence produce racemic mixtures (equal ratios of enantiomers). Thus, both the racemic nature of meteoritic amino acids and the presence of rare compounds have been used to argue for the extraterrestrial origin of these compounds. In particular, the presence of the α-dialkyl-α-amino acids 2-amino-2-methylpropanoic acid (α-aminoisobutyric acid or α-AIB) and 2-amino-2-methylbutanoic acid (isovaline or Iva) in many carbonaceous chondrites has been widely viewed as a marker for amino acids of apparent extraterrestrial origin, since these compounds were believed to be rare in life on Earth (Kvenvolden et al., 1970; Cronin et al., 1979; Ehrenfreund et al., 2001; Martins et al., 2007). However, investigations of filamentous fungi have revealed that many species of these fungi have both α-AIB and Iva in their peptides (e.g., Kenner and Sheppard, 1958; Daniel and Rodrigues, 2007; Brückner et al., 2009). These α-AIB-containing fungal peptides are designated
The abundance of fungal taxa that are potential producers of peptaibiotics raises the concern that the α-AIB and Iva detected in meteorites may be the result of terrestrial contamination. Consequently, reliable analytical methods that can distinguish between abiotically and biotically synthesized α-AIB and Iva are necessary. Analyses of meteorites that have resided on Earth for various periods of time show that contamination by bacteria or fungi can occur relatively quickly and can continue over several years (Oró and Tornabene, 1965; Glavin et al., 1999; Kminek et al., 2002; Toporski and Steele, 2007). Some terrestrial contamination was also deduced from isotope data of protein amino acids in the Murchison and Murray meteorites (Pizzarello and Cooper, 2001) and from nonracemic enantiomeric ratios for some protein amino acids in the CM2 carbonaceous chondrites Murchison, LEW 90500, and ALH 83100 (Glavin et al., 2006).
In addition, α-AIB has been detected in a sample of blue ice collected from La Paz, Antarctica, a region where many meteorites have been retrieved (Glavin et al., 2006; Botta et al., 2008). Isovaline, however, was not detected in this ice sample. The α-AIB found in the La Paz ice was presumed to have leached from micrometeorites or other extraterrestrial particles present in the ice, either after falling to Earth or during the ice extraction procedure (Glavin et al., 2006). A terrestrial source of the α-AIB found in Antarctic ice cannot, however, be ruled out. Filamentous fungi (including yeasts) have been found in permafrost and entombed in glacial ice in Antarctica, as well as the Arctic and Greenland (e.g., Castello and Rogers, 2005; Onofri et al., 2007). Included among the fungal taxa isolated from these environments are several that are known to produce peptaibiotics: Emericellopsis spp. produce antiamoebins, emerimicins and zervamicins; Acremonium crotocinigenum produces acretocins; Trichoderma longibrachiatum makes longibrachins; and Geotrichum candidum produces neoefrapeptins that also contain D- and L-Iva in the same peptide sequence (Leclerc et al., 2001; Fredenhagen et al., 2006; Krause et al., 2007; Brückner et al., 2009). Thus, as more is learned about the potential for contamination from α-AIB-producing and D- and L-Iva-producing fungi, the need for additional proof beyond the presence of these amino acids and the enantiomeric ratio of Iva becomes mandatory in order to confirm a lack of terrestrial contamination of meteorites.
Isotopic measurements of meteoritic compounds are also used to characterize a terrestrial versus extraterrestrial origin. Extraterrestrial organic compounds typically are enriched in the heavier isotopes (e.g., 13C vs. 12C, 15N vs. 14N, D vs. H), which reflect the isotopic ratios of the interstellar precursors of these molecules formed in cold environments (Pizzarello et al., 1991; Sandford et al., 2001; Messenger, 2002; Pizzarello and Huang, 2005). Analyses of meteoritic amino acids have shown δ 13C values typically in the +20‰ to +40‰ range (Pizzarello et al., 2004; Martins et al., 2007; Pizzarello and Holmes, 2009). Amino acid δ 13C values measured for a suite of amino acids in a diverse group of terrestrial organisms show a range from −70.5‰ to +11.25‰ (Scott et al., 2006). The difference between the isotopic values of biologically produced amino acids and those seen in meteorites is strongly indicative of an extraterrestrial origin for the meteoritic organics. However, the δ 13C value for α-AIB from fungal peptides has not been previously measured, so it is not known how distinct their value is from the meteoritic values.
Here, we report on the molecular distribution and stable carbon and nitrogen isotopic ratios for amino acids found in the total hydrolysates of four biologically synthesized peptaibiotics. They comprise alamethicin, paracelsin, trichotoxin, and antiamoebin, and represent natural, microheterogeneous mixtures isolated from fermentation broths of common species of Trichoderma and Emericellopsis. Molecular distribution analyses were performed via liquid chromatography with fluorescence detection and time-of-flight mass spectrometry (LC-FD/ToF-MS). Compound-specific stable isotopic measurements were carried out with gas chromatography coupled with quadrupole mass spectrometry and isotope ratio mass spectrometry (GC-MS/IRMS). We compared these measurements with those from the CM2 carbonaceous chondrite Murchison and three Antarctic CR2 carbonaceous chondrites. We discuss the results in the context of understanding the peptaibiotics as a potential source of meteoritic contamination.
2. Materials and Methods
2.1. Fungal peptides
Four isolates of peptides from fungi obtained as pure cultures from culture collections were grown in suitable culture broths and analyzed. Accession numbers of strains and currently accepted names (in parentheses) are given. Table 1 shows the amino acid sequences of these peptides.
Amino acids in parentheses indicate some major amino acid exchange positions. Chiral amino acids and amino alcohols are of the L configuration with the exception of D-Iva. Abbreviations used are standard amino acid nomenclature, with the following special abbreviations: Ac, acetyl; Aib, α-aminoisobutyric acid; D-Iva, D-isovaline; Pheol, phenylalaninol; Valol, valinol.
Alamethicin
Alamethicins F50 peptides were isolated from Trichoderma viride (Trichoderma cf. brevicompactum) NRRL 3199. The malt extract medium contained 30 g malt extract (Serva, Heidelberg) and 3 g soy peptone (Oxoid) in 1 L of tap water adjusted to pH 6–6.5. The crude mixture of alamethicins was isolated from 80 L of culture broth by using XAD-adsorber resin and purified by silica gel chromatography.
Paracelsin
Paracelsins A–D were isolated by organic solvent extraction from the wet mycelium of the mutant strain Trichoderma reesei (Hypocrea jecorina) QM9414 produced in 80 L modified Raulin-Thom medium. The Raulin-Thom medium was prepared with D-glucose (50.0 g), L-tartaric acid (2.7 g), diammonium tartrate (2.7 g), (NH4)2SO4 (0.16 g), (NH4)2HPO4 (0.4 g), K2CO3 (0.4 g), basic magnesium carbonate (0.33 g), FeSO4 · 7H2O (0.046 g), ZnSO4 · 7H2O (0.046 g) dissolved in 1 L of tap water and adjusted to final pH 4.0. The crude peptide mixture was purified by Sephadex LH-20 chromatography and crystallized from hot methanol/water.
Trichotoxin
Cultures of Trichoderma viride (T. asperellum) NRRL 5242 were grown statically in modified Raulin-Thom medium in Fernbach flasks. The crude peptide mixture was isolated from the wet mycelia by extraction with methanol and dichloromethane. Peptides were purified by Sephadex LH-20 chromatography, and microheterogeneous groups trichotoxins A40 and A50 separated by silica gel chromatography.
Antiamoebin
A mixture of peptides composed mainly of antiamoebin I (ca. 85%) and antiamoebin II (ca. 15%) was isolated from the culture broth of Emericellopsis synnematicola CBS 176.60 grown in 80 L malt extract medium. The crude peptide mixture was purified by chromatography and crystallized from methanol/water.
2.2. Meteorite samples
The CM2 carbonaceous chondrite Murchison (USNM 5453, mass 6.3 g) was provided by the Smithsonian National Museum of Natural History, Washington DC. The Antarctic CR2 carbonaceous chondrite Elephant Moraine 92042 (specific 58, parent 0, mass 0.6 g) was provided by the Antarctic meteorite curator at the NASA Johnson Space Center in Houston, Texas. The other Antarctic meteorites discussed in this paper were analyzed by other research groups and are reported elsewhere (Martins et al., 2007; Pizzarello and Holmes, 2009).
2.3. Extraction and processing procedures
All glassware and sample handling tools were rinsed with Millipore Direct Q3 UV (18.2 MΩ, <5 ppb total organic carbon) ultrapure water, wrapped in aluminum foil, and then heated in a furnace in air at 500°C overnight. The four vacuum-dried fungal peptide residues analyzed in this study were shipped to NASA Goddard Space Flight Center from the University of Giessen in capped microcentrifuge vials. Each vial contained an estimated 2–5 mg peptide in solid or crystal form. One empty sterile microcentrifuge vial (procedural blank) was carried through the identical extraction procedure as the peptides. To each microcentrifuge vial 0.5 mL of methanol (Optima Grade) was added and then mixed with a vortex for 5 min. After mixing, a 0.1 mL aliquot of each methanol extract was transferred into separate clean glass test tubes (10 × 75 mm), dried under vacuum, and then hydrolyzed under 6 M HCl vapor by flame-sealing each small test tube inside a larger test tube (20 × 150 mm) containing 1 mL double-distilled 6 M HCl and then by placing the large sealed tubes in an oven at 150°C for 3 h (Glavin et al., 2006). After HCl vapor hydrolysis, the large tubes were broken open and the small interior tubes removed and dried under vacuum to remove any residual HCl from the hydrolyzed sample. One milliliter of Millipore water was added to each sample (acid-hydrolyzed extract), and the vials were stored at −20°C.
The extraction protocol and amino acid analyses of the EET 92042 meteorite sample are described elsewhere (Glavin and Dworkin, 2009); isotopic analyses of the amino acids in this meteorite were performed and reported by Martins et al. (2007). The Murchison meteorite sample was crushed into a fine powder and homogenized with a mortar and pestle. The sample was then split into three portions (each ∼2 g), and each portion was sealed in a glass test tube with 1.5 mL Millipore water and extracted at 100°C for 24 h. The water supernatants were then subjected to the acid vapor hydrolysis procedure described above. The resulting acid-hydrolyzed extract was desalted by using cation-exchange resin (AG50W-X8, 100–200 mesh, hydrogen form, Bio-Rad), and the amino acids were recovered by elution with 2 M NH4OH. The desalted, hydrolyzed extracts from the three portions were then combined for amino acid analysis. A procedural blank was created by performing the same extraction, hydrolysis, and desalting procedures as the meteorite and fungal peptide samples in parallel but in the absence of any sample.
2.4. LC-FD/ToF-MS amino acid measurements
Most of the chemicals and reagents used in this study were purchased from Sigma-Aldrich. A stock amino acid solution (10−6 M) was prepared by mixing individual standards (97–99% purity) in Millipore water. The o-pthaldialdehyde/N-acetyl-L-cysteine (OPA/NAC) reagent used for amino acid derivatization was prepared by dissolving 4 mg OPA in 300 μL methanol (Optima Grade) and then adding 685 μL 0.1 M sodium borate buffer (pH 9) and 15 μL 1 M NAC. A 0.1 M hydrazine (NH2NH2) solution was prepared by vacuum distillation of concentrated anhydrous hydrazine (98% purity) and subsequent dilution in Millipore water. The hydrochloric acid (HCl) was double distilled, and the ammonium formate buffer used in the LC-FD/ToF-MS analyses was prepared by ammonia titration (95% purity) of a 50 mM formic acid solution to pH 8.
Prior to LC-FD/ToF-MS analyses, each 1 mL acid-hydrolyzed extract was diluted by a factor of 1000 by taking a 1 μL aliquot of the extract and adding 999 μL of Millipore water. Then, 10 μL of each diluted sample (corresponding to 1/100,000 of the hydrolyzed peptide, or ∼0.004 to 0.01 micrograms of the original nonhydrolyzed peptide) was combined with 10 μL of 0.1 M sodium borate buffer in a glass autosampler vial, the solution dried under vacuum, the residue brought up in 20 μL of Millipore water and then derivatized by adding 5 μL of OPA/NAC (Zhao and Bada, 1995). The derivatization reaction was quenched after 15 min at room temperature with 75 μL of 0.1 M aqueous hydrazine, and 10 μL of each derivatized extract was injected into a Waters ACQUITY Ultra Performance liquid chromatograph with the fluorescence detector coupled in series to a Waters LCT Premier time-of-flight mass spectrometer that uses positive electrospray ionization. The details of the ToF-MS settings and the amino acid quantification methods used for these analyses are described elsewhere (Glavin et al., 2006). Each derivatized sample was analyzed by using tandem LC column conditions optimized for amino acid separation and characterization (Glavin et al., 2010a). The first column was a Waters BEH C18 column (2.1 × 50 mm, 1.7 μm bead) followed by a second Waters BEH Phenyl-Hexyl column (2.1 × 150 mm, 1.7 μm bead). The conditions for separation of the OPA/NAC amino acid derivatives at 30°C were as follows: flow rate, 150 μL/min; solvent A (50 mM ammonium formate, 8% methanol, pH 8.0); solvent B (methanol); gradient, time in minutes (%B): 0 (0), 35 (55), 45 (100). For comparison, the hydrolyzed hot water extracts from the Antarctic CR2 meteorite EET 92042 and the CM2 Murchison meteorite were analyzed under the same conditions. Additional results from the amino acid analyses of these meteorites are reported elsewhere (Glavin et al., 2006, 2010b).
2.5. Isotopic measurements
For the isotopic analyses, aliquots of each hydrolyzed peptide sample (20 μL for carbon analysis, 100 μL for nitrogen analysis) and of the Murchison meteorite sample (10% of the total combined extract for carbon analysis, 30% for nitrogen analysis) were dried under vacuum by a LabConco CentriVap centrifugal concentrator. Samples were esterified with isopropanol and the isopropyl esters reacted with trifluoroacetic anhydride (TFAA) with established methods (e.g., Elsila et al., 2009). Derivatized samples were dissolved in 5 μL of ethyl acetate (Fisher Chemical, Optima Grade).
The δ 13C and δ 15N values of the TFAA-isopropanol derivatized samples were analyzed by GC-MS/IRMS, which provides compound-specific structural and stable isotopic information from a single sample injection. The GC-MS/IRMS instrument consists of a Thermo Trace gas chromatograph whose output is split, with approximately 10% directed into a Thermo DSQII electron-impact quadrupole mass spectrometer that provides mass and structural information for each eluting peak. The remaining ∼90% passes through a Thermo GC-C III interface, where eluting amino acids are oxidized and reduced to form CO2 and N2. These gases are passed into a Thermo MAT 253 isotope ratio mass spectrometer (IRMS) for stable isotopic measurement. For δ 15N analysis, a liquid nitrogen trap is used to remove the CO2 prior to entry into the IRMS.
Derivatized extracts were injected in 1 μL aliquots into the gas chromatograph. For the fungal peptides, the gas chromatograph was outfitted with a Lipodex-E cyclodextrin column (Macherey-Nagel, 50 m, 0.25 mm i.d.), and the following program was used: helium carrier gas flow 2.3 mL/min; oven temperature 70°C for the initial 5 min, ramped at 2°C/min to 100°C, ramped at 4°C/min to 200°C, ramped at 10°C/min to 225°C, and held at 225°C for 5 min. For the Murchison extract, a 5 m base-deactivated fused silica guard column (Restek, 0.25 mm i.d.) and four 25 m Chirasil L-Val columns (Varian, 0.25 mm i.d.) were connected with Press-Tight connectors (Restek), and the following program was used: initial oven temperature 50°C, ramped at 10°C/min to 85°C, ramped at 2°C/min to 120°C, ramped at 4°C/min to 200°C, and held at 200°C for 10 min. Six pulses of CO2 (δ 13C = −24.234‰) or N2 (δ 15N = −0.621‰) that had been precalibrated against commercial reference gases (Oztech Corporation) were injected into the IRMS for computation of the δ 13C or δ 15N values of the eluting compounds. Analysis of the MAT 253 data was performed with Thermo Isodat 2.5 software.
Stock solutions of the amino acids of interest were combined to make a standard mixture that was carried through the derivatization process and run daily on the GC-MS/IRMS. A stock solution of L-alanine with a known δ 13C value (−23.33‰, IsoAnalytical) was also derivatized and injected daily for calibration purposes. The individual, underivatized stock solutions or solid pure standards were also analyzed on a Costech ECS 4010 combustion elemental analyzer connected to the MAT 253 IRMS; this was necessary to correct for the carbon added during derivatization. The final δ 13C values of the amino acids in the samples and their precision were calculated as described elsewhere (Docherty et al., 2001; O'Brien et al., 2002; Elsila et al., 2009). The δ 15N values of the derivatized amino acids were assumed to be identical to those for the underivatized acids, as no nitrogen is added during the derivatization process used in this study.
3. Results
3.1. Amino acid distribution
Liquid chromatography–fluorescence detection and time-of-flight mass spectrometry analysis revealed a range of amino acids present in the fungal peptides (see Fig. 1 for representative chromatograms and Table 2 for calculated abundances). The relative abundances of the detected amino acids agreed with the expected values calculated from the sequences in Table 1 within experimental error and taking the natural microheterogeneity (i.e., amino acid exchange in certain positions) of the peptides analyzed into account. In all cases, α-AIB was present as the most abundant amino acid. D-Iva was present in two of the samples (as a minor component of trichotoxin A40 and a major component of antiamoebins); L-Iva was not observed in any of the samples. In addition, low levels of certain D-amino acids (e.g., D-Glu, D-Leu, and D-Val) were observed. Their formation can most likely be attributed to partial amino acid racemization of the corresponding L-amino acids as a result of the acid vapor phase hydrolysis procedure, since no D-amino acids were detected in a separate analysis with conventional hydrolysis in aqueous 6 M HCl.
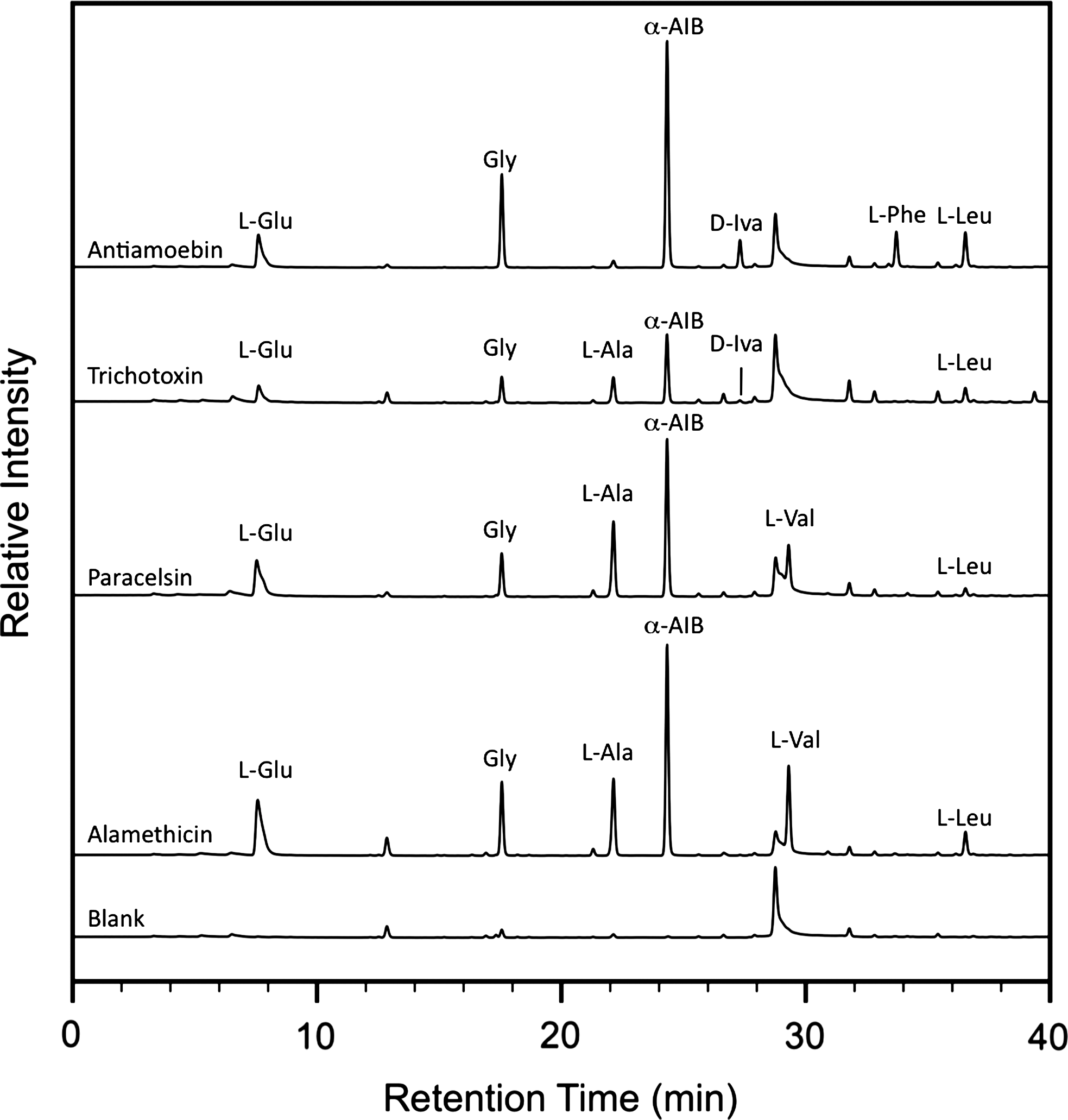
Fluorescence traces from the LC-FD/ToF-MS analysis of the acid-hydrolyzed fungal peptide extracts and a procedural blank.
Concentrations are calculated from the fluorescence chromatograms.
Pro/Hyp and amino alcohols not determined.
n.d., not detected in this peptide.
3.2. Isotopic measurements
Table 3 summarizes the measured isotopic data for the four fungal peptides and the Murchison sample, as well as two CR2 Antarctic meteorites. The δ 13C isotopic ratios were measured for glycine (Gly), α-AIB, and L-alanine (L-Ala) in all samples. D-Iva was measured in the two peptides in which it appeared (trichotoxin A40 and antiamoebin). In addition, δ 15N values were measured for α-AIB and Gly in all samples, for L-Ala in all samples except antiamoebins, and for D-Iva in antiamoebins.
δ 13C values are shown in ‰ relative to Vienna Pee Dee Belemnite, δ 15N values are ‰ relative to air. Errors calculated as described in Methods section; number of replicate analyses are shown in parentheses.
n.d., not detected in this peptide.
D-Iva isolated and measured in fungal peptides; D,L-Iva measured in meteorites.
Abundance too low to measure δ 15N.
From Pizzarello and Holmes (2009).
4. Discussion
4.1. Amino acid distribution
The detection of nonprotein amino acids such as α-AIB and Iva has often been invoked as proof of an extraterrestrial, abiotic origin. The recent discovery of the widespread nature of peptaibiotic-producing fungi, however, calls this assumption into question. To date, about 850 individual sequences of peptides containing α-AIB have been isolated from over 23 common fungal genera, and the potential for contamination of meteorites from these fungi must be recognized.
Analysis of the complete amino acid distribution pattern of a meteorite, as opposed to focusing only on the presence or absence of α-AIB and Iva, continues to be a potentially powerful tool in distinguishing fungal peptide amino acid contamination from indigenous compounds. Figure 2 shows the distribution of the amino acids containing four or five carbon atoms (as indicated by the single ion mass chromatograms of the OPA/NAC derivatives centered at mass-to-charge ratios 365.1171 and 379.1328, respectively) isolated from a representative fungal peptaibiotic (antiamoebins) compared to the Antarctic CR2 meteorite EET 92042 and the CM2 Murchison meteorite. Similar results were obtained for the other three fungal peptides studied in this work. It is clear that the distribution observed in the purified fungal peptide is much less complex than those observed in the meteoritic extracts, with α-AIB being the primary four-carbon and D-Iva the primary five-carbon amino acid. In contrast, the meteorites show complete structural diversity for the acyclic aliphatic C4 and C5 amino acids, as well as a racemic distribution of chiral amino acids that argues against terrestrial contamination. Recent analyses of several different carbonaceous chondrite groups have confirmed that this diversity of C4 and C5 amino acids is common in carbonaceous chondrites (Glavin and Dworkin, 2009; Glavin et al., 2010b). Thus, although the presence of α-AIB in a meteorite can no longer be used as a definitive extraterrestrial marker, the relative simplicity of the presently known distribution of α-dialkyl-α-amino acids expected from fungal peptide contamination provides strong evidence to support the abiotic, indigenous nature of the complex distributions observed in meteorites.
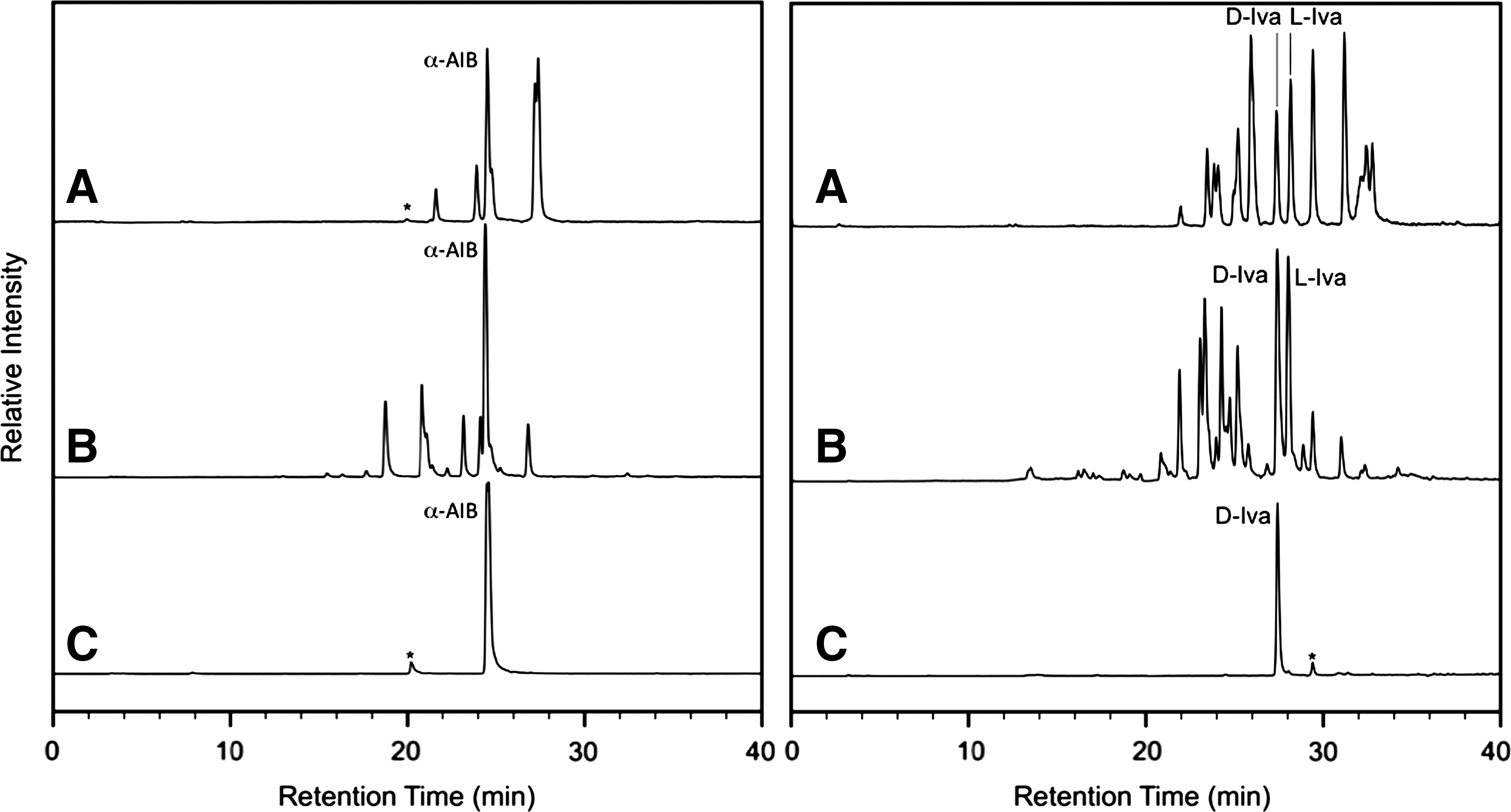
LC-FD/ToF-MS chromatograms corresponding to mass-to-charge ratios 365.1171 ± 0.02 and 379.1328 ± 0.02 (the peak widths at half maximum) of amino acids containing four (left panels) or five (right panels) carbon atoms, respectively. The mass traces shown represent (
4.2. Isotopic analysis
Isotopic analysis represents another important tool for determining the origin of meteoritic organic compounds. Figure 3 shows the δ 13C and δ 15N isotopic values for α-AIB and Gly in the four fungal peptides analyzed in this work compared to values measured from several carbonaceous chondrites. The fungal peptides plot in the lower left regions of the graphs (depleted δ 13C values, δ 15N close to zero), indicating values that fall within typical terrestrial ranges [for comparison, the δ 13C of Gly from a variety of terrestrial organisms has been measured in the −42‰ to +11‰ range (Scott et al., 2006), while δ 15N in terrestrial organics falls in the range −10‰ to +20‰ (Hoefs, 1980)]. By contrast, meteoritic amino acids are enriched in 13C and 15N. Although it is unsurprising that the fungal peptides analyzed here show “terrestrial” values, this is the first isotopic analysis performed that confirms that the α-AIB in these peptaibiotics, produced from fungi grown on a diet of common terrestrial food sources, is not anomalous. The difference between the fungal α-AIB and that observed in meteorites suggests that most of the α-AIB is not the result of terrestrial contamination from such fungi; in other words, it is highly unlikely that the α-AIB detected in the meteorites analyzed in this study is the result of transfer into the meteorites of peptaibiotics produced by fungi in the terrestrial environment in which the meteorite fell or was curated.
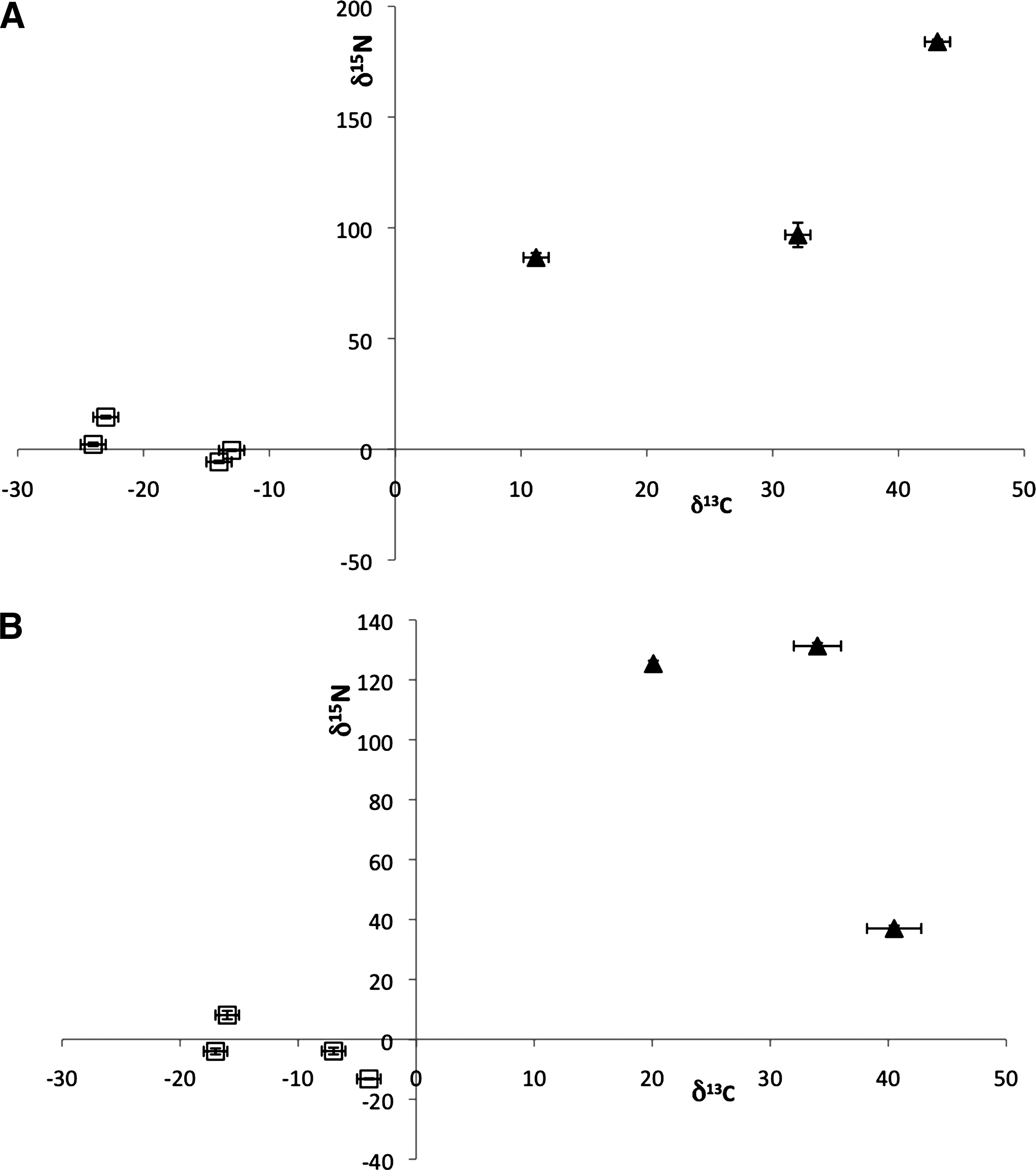
Stable carbon and nitrogen isotopic values measured for (
The δ 13C and δ 15N values of D-Iva were not plotted in Fig. 3 because most meteorite analyses have measured combined D,L-Iva, rather than each enantiomer individually. D,L-Iva typically shows δ 13C values similar to other meteoritic amino acids [e.g., +17‰ to +22‰ in the Murchison meteorite (Engel et al., 1990; Pizzarello et al., 2004)]. However, in some samples the D,L-Iva δ 13C value is significantly depleted in 13C [e.g., δ 13C = −5.1‰ in the LAP 02342 meteorite (Pizzarello and Holmes, 2009)]. For comparison, the δ 13C values measured in trichotoxin and antiamoebin ranged from −16‰ to −25‰. Although the D,L-Iva δ 13C value in LAP 02342 may suggest contamination with fungal Iva, the other protein amino acids in this meteorite possess enriched δ 13C ratios, and the meteorite shows a complex amino acid distribution, which suggests a lack of substantial contamination. Further work is needed to understand the source of the isotopic variability of isovaline in meteorites and to measure their individual D- and L-Iva isotopic values for comparison to terrestrial values.
Although this data suggests that the α-AIB in meteorites was not the result of fungi fed on terrestrial sources, questions remain about the possibility of fungal infiltration of meteorites and the resulting potential for contamination. Extracts and solids from carbonaceous chondrites have been shown to support bacterial and fungal life (Mautner et al., 1997; Mautner, 2002), and filamentous microorganisms have also been detected as contaminants inside the Nakhla meteorite (Toporski and Steele, 2007). Fungi contained within a meteorite may feed on indigenous meteoritic organics; thus they could incorporate the heavy isotope enrichment of this food source into their peptides. Without an understanding of the metabolic pathways responsible for peptaibiotics, it is difficult to predict the extent of any enrichment. Thus, the absolute isotopic values of the fungal peptides analyzed in this study, which were produced by fungi fed on terrestrial nutrients, may not be enough alone to rule out meteoritic fungal contamination.
The relationships between certain amino acids in the peptaibiotics, however, may prove diagnostic. Isotopic relationships between pairs of amino acids from a variety of biological species have previously been evaluated with linear regression analysis, which revealed correlations between compounds linked by biological processes (Scott et al., 2006). To explore the possibility that similar relationships may aid in differentiating fungal peptaibiotics from indigenous meteoritic amino acids, we analyzed the δ 13C and δ 15N values of pairs of amino acids measured from the fungal peptides, using least-squares linear regression. Figure 4 illustrates this data and compares it to similar data from the CM2 Murchison meteorite and the Antarctic CR2 meteorites GRA 95229, EET 92042, and LAP 02342.
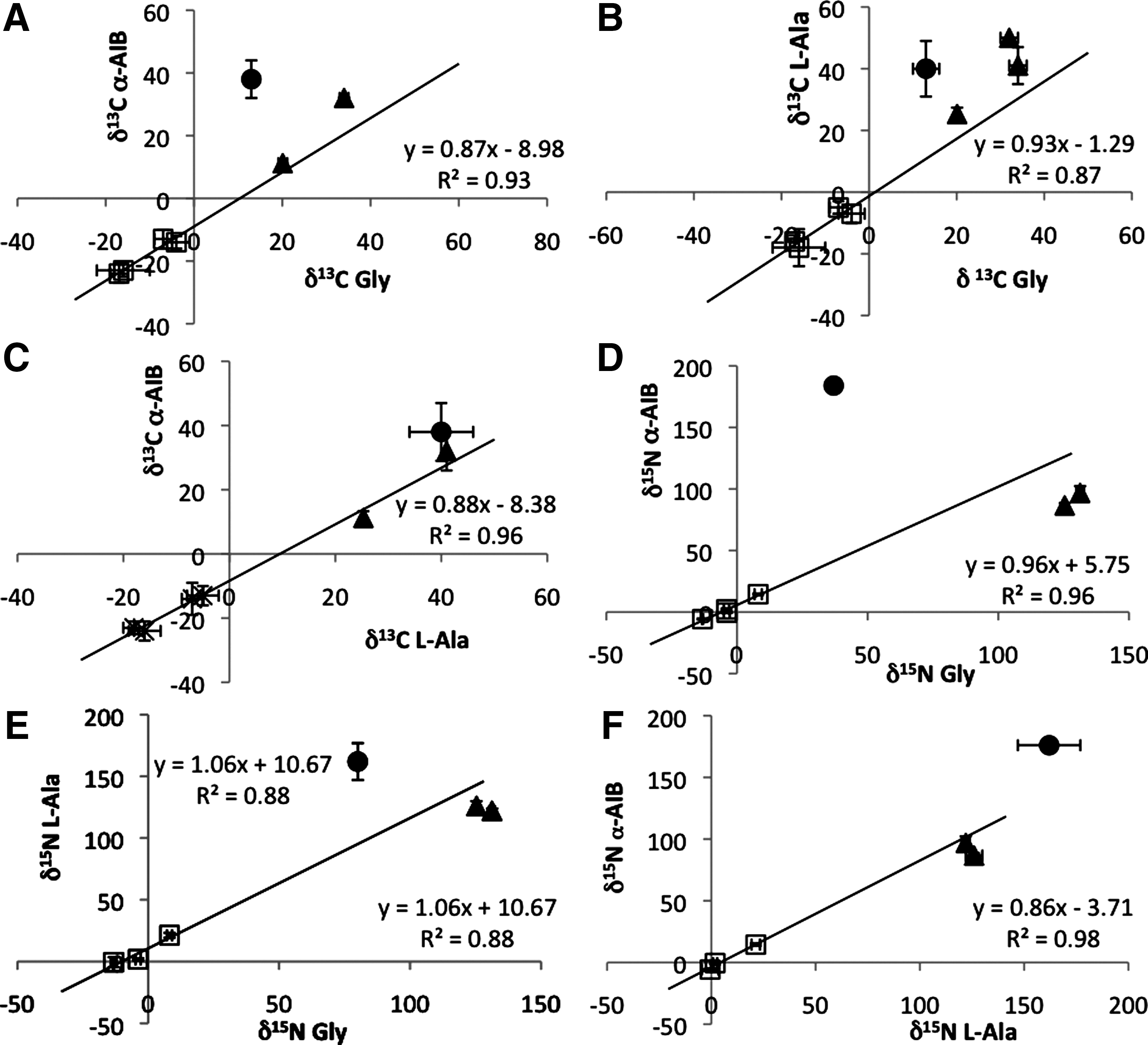
Isotopic ratios and linear regression fits for δ
13C for the pairs of amino acids (
The linear regression through the fungal peptide data in Fig. 4 illustrates that potentially diagnostic relationships do indeed exist between pairs of fungal amino acids. In these fits, a negative y intercept indicates that the amino acid on the y axis is depleted in the heavier isotope compared to the amino acid on the x axis; a positive y intercept indicates enrichment. The data show that α-AIB from the fungal peptaibiotics is consistently depleted in 13C compared to both Gly and L-Ala. α-AIB is also slightly enriched in 15N compared to Gly and slightly depleted in δ 15N compared to L-Ala. L-Ala and Gly have similar δ 13C values, but L-Ala is enriched in δ 15N compared to Gly. D-Iva is not plotted because of the scarcity of data from both fungal peptaibiotics and meteorites, but the δ 13C value of D-Iva is indistinguishable from α-AIB within experimental error in the two peptides in which it was detected. All the analyzed pairs show strong linear correlations (r2 values between 0.87 and 0.96), and the slopes of these lines are within one 95% confidence interval of 1.0.
The observed correlations may speak to the biosynthetic origins of the amino acids. If, for example, α-AIB as a building block for peptaibols is produced by enzymatic methyl addition via adenosyl-methionine to L-Ala (Kubicek et al., 2007), our measurements suggest that this process causes enrichment in the α-AIB relative to the L-Ala, with δ 13C values of α-AIB being enriched by 5–8‰. Presumably, this correlation would hold true regardless of the absolute δ 13C value; that is, α-AIB would be enriched relative to L-Ala even in peptaibiotics produced from fungi feeding on already enriched meteoritic organics.
If this hypothesis is valid, and if the correlations observed in the linear regressions from these four peptaibiotics can be extended to other fungal peptides, then linear regression correlation of amino acid isotopic values may provide a diagnostic tool for ruling out fungal contamination as a source of certain amino acids in meteorites. Meteoritic amino acids that deviate from the correlation observed in the fungi are presumed to originate from other synthetic processes. From Fig. 4A and 4C, it appears that the α-AIB in the Murchison meteorite has a different isotopic relationship with both Gly and L-Ala than that seen from the fungal peptides; Murchison α-AIB is more enriched in 13C compared to Gly and to L-Ala than the corresponding fungal peptide amino acids. Figure 4B also shows that L-Ala is more enriched in 13C compared to Gly in Murchison than in the fungal peptides. The δ 15N relationships between Murchison amino acids shown in Fig. 4D–4F also contrast with those in the fungal peptides. These data confirm the dominant interpretation in the meteoritics and cosmochemistry communities that the amino acids in most specimens of the Murchison meteorite are predominantly indigenous and not due to contamination.
The data from the Antarctic CR2 carbonaceous chondrites are less clear. Figure 4B shows that the L-Ala/Gly relationship in the CR2 meteorites differs from the correlation seen in the peptaibiotics, which again suggests that these amino acids are not the result of fungal contamination; the δ 15N data shown in Fig. 4D and 4E also support this conclusion. The meteoritic data comparing δ 13C values of α-AIB with Gly (Fig. 4A) and L-Ala (Fig. 4C), as well as the δ 15N values of α-AIB and L-Ala (Fig. 4F), however, do not differ significantly enough from the fungal trends to rule out contamination. It is important to note, however, that while these meteoritic amino acid pairs show similar relationships to those observed in fungal peptides, this observation alone is not proof of contamination. Our proposed diagnostic relationship can only be used to rule out contamination; there may be multiple pathways, including extraterrestrial abiotic mechanisms, which produce similar relationships. The observed differences between the CR2 meteorites and Murchison could also indicate different synthetic pathways for these amino acids or a difference in aqueous alteration and processing on the meteorite parent bodies.
4.3. α-AIB in Antarctic ice
The existence of peptaibiotic-producing fungi also raises questions about the origin of α-AIB that has been found in environmental samples. α-AIB has been reported, along with other amino acids, in the extract of an ice block from the La Paz region of Antarctica at a concentration of 1–46 ppt (Glavin et al., 2006; Botta et al., 2008), although it was not detected in a sample of Antarctic ice from the Allan Hills region in a previous study (Bada et al., 1998). No isovaline has ever been reported in Antarctic ice. The concentration of α-AIB in the La Paz Antarctic ice is 106 times lower than the concentration of α-AIB found in the CR2 chondrites EET 92042 and GRA 95229, collected in other regions of Antarctica (Martins et al., 2007). The α-AIB detected in the La Paz Antarctic ice was originally hypothesized to have originated from Antarctic micrometeorites (AMMs) present in the ice, which was not filtered prior to analysis (Glavin et al., 2006). However, subsequent analysis of grains collected in filters from several liters of a different block of La Paz ice found no evidence of extraterrestrial particles in the ice (unpublished results). Measurements of AMMs have shown α-AIB abundances ranging from ∼50 to 220 ppb (Matrajt et al., 2004), although one micrometeorite sample was reported to have a much higher abundance of α-AIB (∼280 ppm) (Brinton et al., 1998).
The well-characterized existence of potentially peptaibiotic-producing fungi occurring or entombed in ice and permafrost from Antarctica, the Arctic, and Greenland (e.g., Castello and Rogers, 2005; Onofri et al., 2007) suggests that fungal α-AIB could also leach into Antarctic ice. The detected α-AIB could also result from other abiotic sources of terrestrial contamination, such as industrial or geochemical processes (Mita and Shimoyama, 1998), and these sources could also have distinct isotopic signatures that could help determine the origin of the α-AIB found in the ice. No isotopic analysis of α-AIB extracted from Antarctic ice or AMMs has yet been performed; such analysis, although difficult because of the extremely small sample sizes available, could help to resolve the origin of the α-AIB in Antarctic ice and extracted from AMMs.
5. Conclusions
Although many astrobiologists still consider α-AIB to be rare in the terrestrial biosphere, this belief has been challenged by the discovery of α-AIB in peptides produced by fungi in a variety of environments. The existence of terrestrial peptaibiotics necessitates a revisiting of previously analyzed meteoritic data, in which the existence of α-AIB and other “exotic” amino acids was often used as sufficient proof of an extraterrestrial origin.
Our analyses of four α-AIB-containing fungal peptides showed notable differences with available meteoritic data. The distribution of amino acids is often strikingly complex in meteorites, with the acyclic C4 and C5 aliphatic amino acids showing complete structural diversity, while the corresponding distributions in the fungal peptides consist primarily of α-AIB and Iva. These distinct distributions provide a means of evaluating the likelihood of terrestrial contamination as a source of meteoritic amino acids.
We have also used linear regression analysis to elucidate correlations between the stable isotopic compositions of the amino acids α-AIB, Gly, and L-Ala in fungal peptides. These correlations can be compared with meteoritic data as another tool to rule out terrestrial contamination, although they are insufficient to prove the presence of contamination. The diagnostic correlations proposed here, however, are based on data from only four peptides. Analyses of more peptaibiotics are needed to determine whether these correlations are broadly applicable. Similarly, this work illustrates the need for additional compound-specific isotopic analysis of amino acids in meteorites. Sparse data exist in this area, but the work reported here demonstrates its potential use to determine the source of amino acids in meteorites and help elucidate abiotic formation pathways.
Footnotes
Acknowledgments
We are grateful for funding support from the NASA Astrobiology Institute and the Goddard Center for Astrobiology, and the NASA Cosmochemistry Program. M.P.C. acknowledges support from the NASA Postdoctoral Program at the Goddard Space Flight Center, administered by Oak Ridge Associated Universities through a contract with NASA.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
α-AIB, α-aminoisobutyric acid; Ala, alanine; AMMs, Antarctic micrometeorites; FD, fluorescence detection; GC-MS, gas chromatograph–mass spectrometer, gas chromatography–mass spectrometry; Gly, glycine; IRMS, isotope ratio mass spectrometer, isotope ratio mass spectrometry; Iva, isovaline; LC, liquid chromatography; NAC, N-acetyl-L-cysteine; OPA, o-phthaldialdehyde; TFAA, trifluoroacetic anhydride; ToF-MS, time-of-flight mass spectrometer, time-of-flight mass spectrometry.