
Abstract
We revisit the validity of the presence of O2 or O3 in the atmosphere of a rocky planet as being a biosignature. Up to now, the false positive that has been identified applies to a planet during a hot greenhouse runaway, which is restricted to planets outside the habitable zone (HZ) of the star that are closer to the star. In this paper, we explore a new possibility based on abiotic photogeneration of O2 at the surface of a planet that could occur inside the HZ. The search for such a process is an active field of laboratory investigation that has resulted from an ongoing interest in finding efficient systems with the capacity to harvest solar energy on Earth. Although such a process is energetically viable, we find it to be a very unlikely explanation for the observation of O2 or O3 in the atmosphere of a telluric exoplanet in the HZ. It requires an efficient photocatalyst to be present and abundant under natural planetary conditions, which appears unlikely according to our discussion of known mineral photochemical processes. In contrast, a biological system that synthesizes its constituents from abundant raw materials and energy has the inherent adaptation advantage to become widespread and dominant (Darwinist argument). Thus, O2 appears to continue to be a good biosignature. Key Words: Biosignature—Oxygen—Ozone—Photosynthesis—Artificial photosynthesis—Photocatalysis. Astrobiology 11, 335–341.
1. Introduction
T
In other words, the qualification of biological photosynthesis as the only possible generator of significant O2 concentrations has been pivotal to most of the proposed projects to search for life outside the Solar System [e.g., the Darwin/Terrestrial Planet Finder Interferometer (thermal IR), the Terrestrial Planet Finder Coronagraph, and the New World Observer (visible and near-IR)], and this should be carefully studied. In the present paper, we summarize and discuss the arguments for proposing O2 and O3 as biosignatures (Section 3), explore the process of photocatalysis of water splitting as a source of O2 (Section 4), and contrast the characteristics of photocatalysis to those of biological systems (Sections 5 and 6).
2. What Is a Biosignature?
Presently, there are no definitive biosignatures. Our best current definition is pragmatic and temporary. It is “an observable feature of a planet, such as its atmospheric composition, that our present models cannot reproduce when including the abiotic physical and chemical processes we know about.” This does not exclude the idea that, in the future, new models may explain the observed features without the need of a biological system. This definition also makes the search for any possible false positives particularly important. Once detected, a biosignature will thus have to be put to the test by the scientific community.
3. The Case of O2 and O3 as Biosignatures
3.1. The need for an abundant production of O2
The presence of O2 as a biosignature was proposed for the first time in a pioneer communication by Tobias Owen (1980). The possibility of detection of (primitive) life on an exoplanet by remote sensing opened a new scientific field with a major potential for the future and the possibility to provide a scientific answer to a question raised two millenniums prior (Epicure, 300 B.C.), “Are we alone in the universe?” At least, this could be the beginning of an answer in that a biosignature can only address the question of primitive (photosynthetic) life, not that of intelligent life, which would require the search for technomarkers.
Since 1980, the proposal of O2 as a biosignature has been revisited by a number of authors whose intent was to qualify or falsify the notion (Kasting et al., 1993; Rosenqvist and Chassefière, 1995; Kasting, 1997; Selsis et al., 2002). The main objection that has been considered so far is the possibility of abiotic production of O2 by UV photolysis of H2O (Rosenqvist and Chassefière, 1995).
Qualitatively, water photolysis can occur efficiently only if H2O rises en masse and remains in the stratosphere. On a temperate planet like Earth, such an ascent does not happen because most of the water vapor produced at the planetary surface by evaporation or sublimation of H2O cools upon ascending to a temperature at which it condenses and then returns to the surface as rain or snow. If the atmosphere is warm enough so that, at the coldest location (tropopause), the saturating vapor pressure is sufficient to allow a significant flux through it, an abundant photolysis can occur, which would produce a large amount of abiotic O2 and allow the corresponding hydrogen to escape. Likely, this is what happened on Venus.
Quantitatively, Kasting et al. (Kasting et al., 1993; Kasting, 1997) studied this greenhouse runaway and concluded that it is expected only if the planet is close enough to its star, namely, closer than the inner boundary of the habitable zone (HZ) so that its tropopause is hot enough. The risk of observing a planet “in the act” of losing its O2 exists only when it is located closer to the star than the HZ. Therefore, if the planet is inside the HZ of its star, an O2 biosignature can still be claimed.
Specifically, the condition is that of a surface temperature colder than 50°C, below which a hot runaway process cannot start. In fact, there is no lower limit on the surface temperature, because even on a snowball planet volcanic gas would pierce the ice layer and reduce the small amount of O2 produced by photolysis of the water that reaches the stratosphere, which would remove oxygen from such an atmosphere. It must be added, though, that a biological production of oxygen on a planet with a very low mean temperature is also unlikely, unless the planet has large seasonal variations from a large obliquity, because the type of life we are looking for needs liquid water.
On a planet in the HZ, the minimum amount of O2 above which biological activity would need to be invoked is P O2 ∼ 10 mbar at the surface (Rosenqvist and Chassefière, 1995).
A large production is needed to reach such partial pressures because O2 is a reactive gas that promptly oxidizes the Fe2+ contained in the fresh lava provided by volcanism, mainly that of oceanic ridges, and produces Fe3+ compounds. The mere visual color of rocks is an indicator. Fresh volcanic rocks are dark (Fe2+), whereas altered rocks, as in deserts, are red (Fe3+). The different volcanic sulfides are additional reducing materials, but on Earth their output rate is ∼ 15 times less than that of iron (1.1 × 1011 mol yr−1 of S release, versus 1.7 × 1012 mol yr−1 of Fe; Bach and Edwards, 2003).
On Earth, the abiotic production of O2, for example, by UV photolysis of water, is less than 1 ppm of the photosynthetic production (Walker, 1977).
If the O2 production by biological photosynthesis on Earth were to stop, its abundance in our atmosphere would decrease in a short time (2 × 107 yr; Léger et al., 1993) compared to the age of our planet (4.6 × 109 yr). Therefore, the presence of O2 in most cases would reveal a biological activity at the present time. This is assuming that fresh Fe2+ is continuously brought to the surface from the planetary mantle. It would be the case, on an Earth-like planet, if plate tectonics is active and volcanic ridges provide abundant lava upwellings. In the absence of plate tectonics, this argument is no longer valid, as indicated by the oxidizing surface of Mars (Benner et al., 2000). It is then essential to confirm, or falsify, the suggestion by Valencia et al. (2007) that plate tectonics is likely to be maintained in massive Earth-like planets 1 . A recent study by Korenaga (2010) goes in that direction and suggests that massive-Earth analogues with water would indeed have plate tectonics.
It should be noted that missions with the capacity to detect O2 in the visible part of the planetary spectrum would measure the absorption of the stellar light by the planetary atmosphere (twice) in the A and B bands of O2. These are sensitive to the total column density of O2 and cannot distinguish between tropospheric and stratospheric oxygen. In practice, the observations would be fully dominated by tropospheric O2 because the troposphere has the highest amount of material, by far, for a gas uniformly mixed in the atmosphere. The detection of O2 bands would allow the measurement of an abundance that corresponds to P O2 = 10 mbar if a 3% precision can be obtained in a visible spectrum at ∼ 75 resolution (Des Marais et al., 2002), a difficult task indeed.
The case of O3 as a marker of O2 is somewhat subtle. On one hand, it has the major advantage of being active in the IR 2 and being a logarithmic tracer (Léger et al., 1993; Des Marais et al., 2002), which implies that even a small amount of O2 could be discovered this way. On the other hand, this logarithmic dependence upon the O2 content would make a precise estimate of the abundance of the latter difficult. Clearly, the desirable situation would be an access to both spectral domains, the IR (O3) and the visible (O2).
3.2. O2 and H2 production by H2O abiotic photosplitting
In the HZ, the most abundant power available at the surface of a planet, once it has solidified its magma ocean (a few million years after a giant impact), is by far the stellar irradiation. On Earth, this corresponds to 1.3 kW m−2 outside the atmosphere and ∼350 W m−2 when averaged over latitudes and the day-night cycle (Rutgers, 2010), compared to only 0.09 W m−2 for the geothermal flux (Pollack et al., 1993; Schubert et al., 2001). Consequently, if abiotic water photolysis is possible on a significant fraction of the planetary surface, with the assistance of a catalyst, it can be an abundant source of O2.
Water splitting is summarized as
Using the formation enthalpy of the different species (Johnson, 2002), one finds a reaction enthalpy of ΔH 1 = −2.52 eV for water vapor and ΔH 2 = −2.98 eV for liquid water, which correspond to photons with wavelengths of λ < 490 nm (vapor) and λ < 420 nm (liquid), respectively. However, the term 1/2 O2 indicates that intermediate species are produced with higher enthalpies, for instance the O* radical, so that the preceding values are minimum enthalpies in absolute value. The high enthalpy of the intermediates can either be reached by adding the energies of several successive photons (biological photosynthesis) or by significantly decreasing it with the action of a suitable catalyst that provides stabilization through interactions at its surface (photogeneration of O2 and H2 through water splitting that is usually, but somewhat incorrectly, called “artificial photosynthesis;” Barber, 2009).
Then, energetically, it is possible that abiotic water splitting can be produced by the absorption of a single photon at the surface of a planet orbiting a solar-type star (F, G, or K types). However, this photolysis would need a photocatalyst so that the system would have the capacity to convert almost the whole energy of one photon into chemical energy. For comparison, the direct photolysis of water molecules in the high stratosphere is performed by photons that are much more energetic, hν > 6.9 eV, λ < 180 nm (Fig. 1).
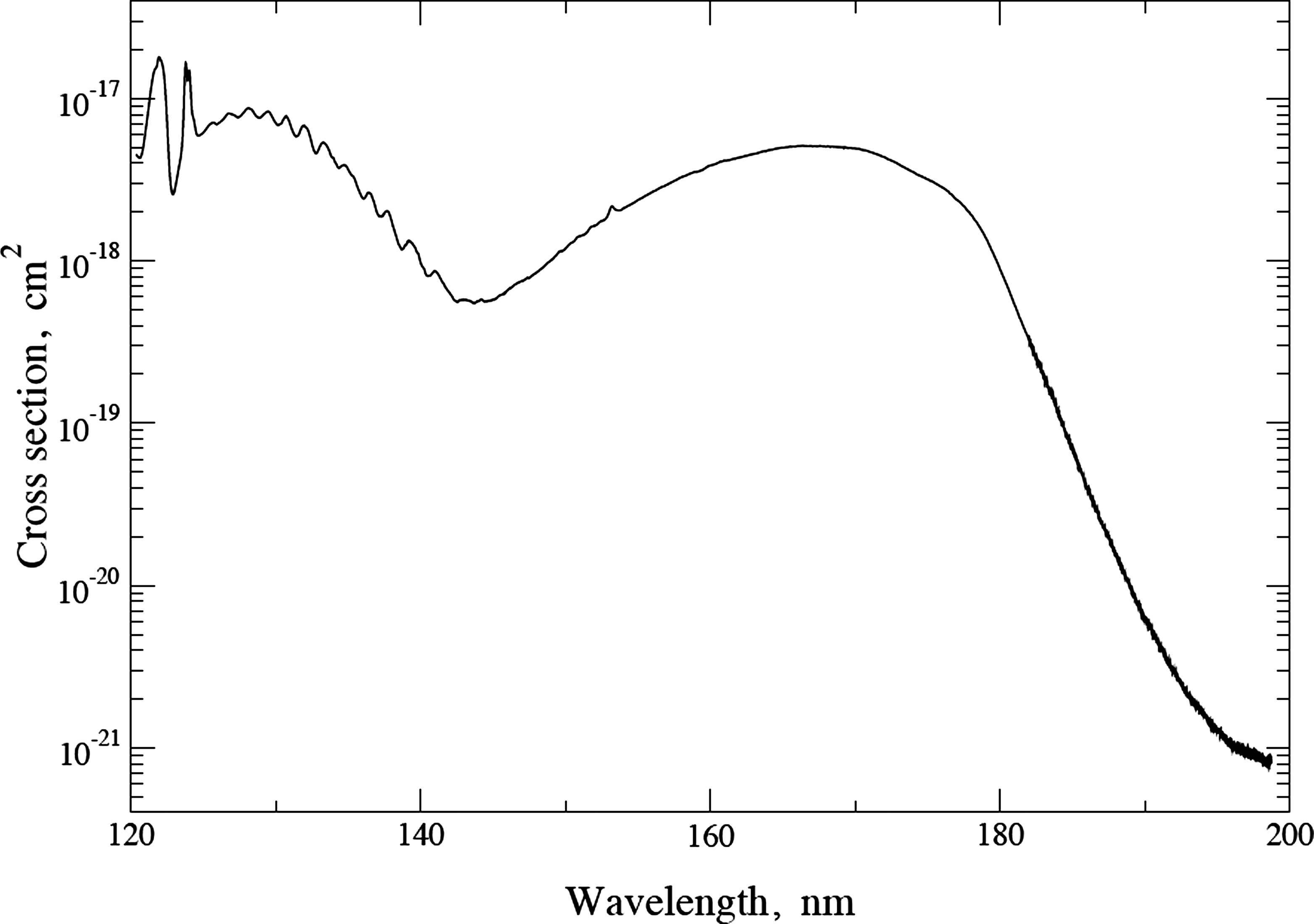
Cross section of H2O photoabsorption, according to Parkinson and Yoshino (2003).
This is because, in the direct absorption of a photon, the transition must be vertical in a representation where the electronic energies are a function of nucleus positions. The latter has no time to move during the fast electronic transition induced by the photon absorption. The produced compact antibonding state is more energetic, by a factor of over 2, than the same state when nuclei are far away (Fig. 2).
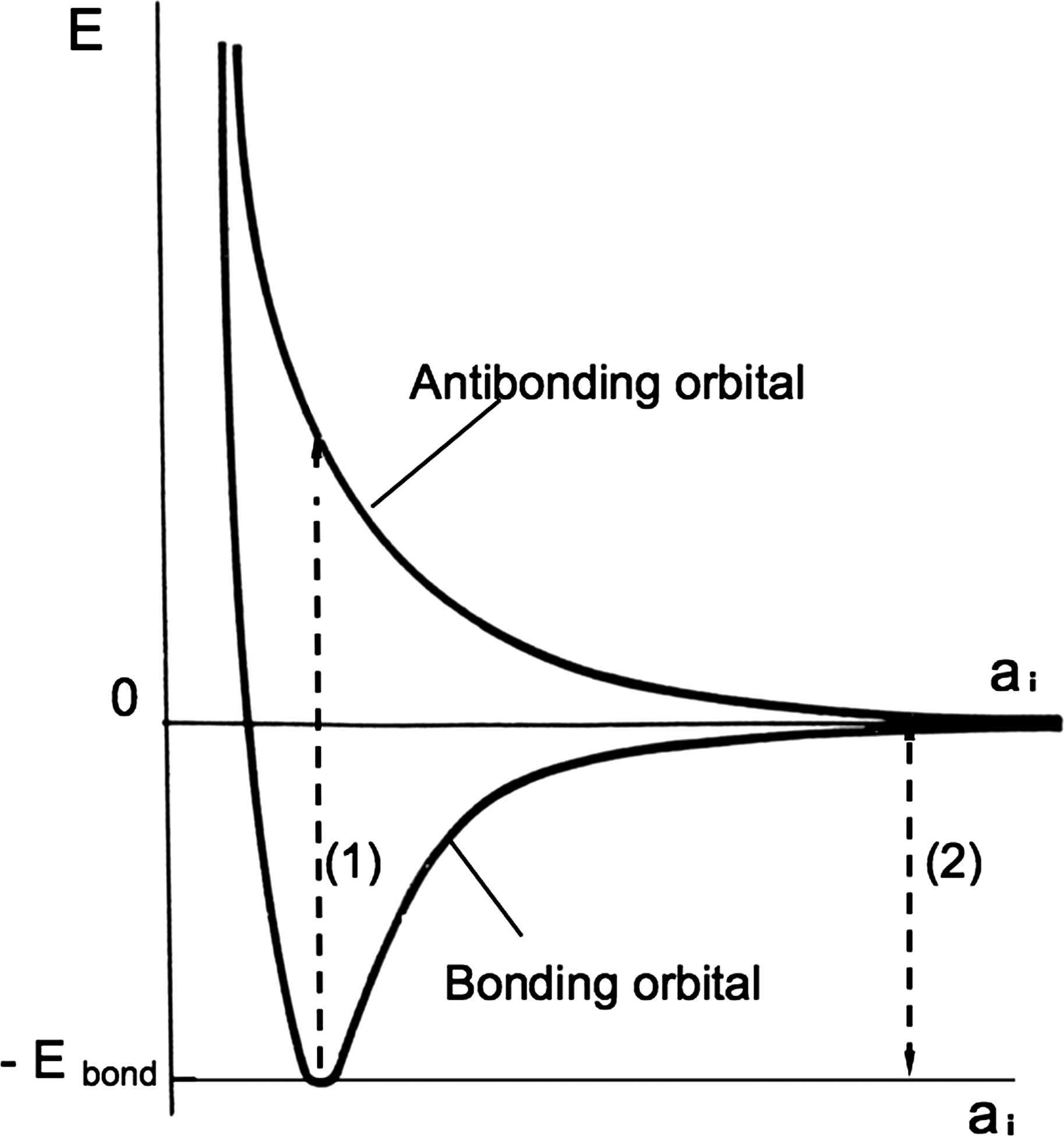
Molecular potential energy curve for a water molecule, versus one of the parameters ai describing the nucleus positions, for bonding and antibonding electronic orbitals. The electronic transition made by a photon absorption is so fast that nuclei have no time to move [ratio of motion times for same kinetic energies: (M nucl/m el)1/2]. In this scheme, it is a vertical transition. The corresponding energy (1) is significantly larger than the bonding energy (2) = E bond that corresponds to the situation where fragments are separated (large ai). The zero point energy is significantly smaller than the electronic energies, e.g. ∼Ebond/25, and is not represented.
On Earth, biological photosynthesis uses a complex protein machinery, for example, chloroplasts in plants, that is able to collect sunlight, store the energy of several photons (usually eight), and convert a fraction of this energy into chemical energy. A critical reaction step is the splitting of two H2O molecules into O2, electrons, and protons (McEvoy and Brudvig, 2006). This allows photosynthesis to be efficient even with low-energy photons, for example, those at the chlorophyll's absorption peak at 650 nm (1.91 eV). Such machinery is the result of a long evolution and is not expected in an abiotic world. In the latter, the storage of photon energy over a period of time long enough to allow the absorption of a second photon (t ∼ 1 μs) seems unlikely because the energy relaxation of excited electrons is a rapid process, even in artificial high-technology materials that are optimized for this process, such as high-purity Si carefully doped (t ∼ 1 ns; Bongiovanni and Staehli, 1992). This is one of the main reasons why the search for artificial photocatalysts is so difficult.
The question raised then is, “Could a plausible abiotic system have the capacity to build up an O2-rich atmosphere (P O2 > 10 mbar) through photocatalytic splitting of water, the system one that is produced in the natural conditions of a planet in the HZ of its star?” The falsification or qualification of O2 as an actual biosignature depends on the answer to this question.
4. Search for an Abiotic Photocatalyst of H2O Splitting
Strong industrial and societal motivations have emerged in recent decades for finding such a photocatalyst, given its potential economic and environmental impact. Starting with the first main success obtained in 1972 by Fujishima and Honda with TiO2 (albeit a low yield), this research field has been very active and has been named somewhat incorrectly “artificial photosynthesis” (Barber, 2009). A photocatalyst requires two components: (i) a photosensitizer unit to collect photons and generate a charge separation (an electron-hole pair) and (ii) a catalyst to capture the pair, minimize charge recombination, and drive the oxidation of water. In abiotic nature, the photosensitizer can be found in a number of semiconducting metal oxides such as TiO2, Fe2O3, WO3, whereas catalysts can be provided by Co3O4 (Frei, 2009) or Mn2CaO4 (Najafpour et al., 2010), with the use of atomic species that are rather abundant on a telluric planet (Table 1). Other materials such as IrO2 or Pt provide better yields but correspond to species with low abundances (Table 1).
However, the abundance problem depends upon the physical state of the considered species. For instance, TiO2 can be dispersed in nanoparticles on a photon-collecting support, which requires much less TiO2 material than a continuous thick layer. Nanoparticles are abundant in the natural conditions of soils (Schrick et al., 2004), so the possibility of the utilization of low-abundance material is not excluded.
However, these systems meet several difficulties as a major source of O2 in the natural environment of a planet, namely: (a) To date, their quantum yields are rather low (<5%), but this may change toward larger values in the future, as the result of efforts presently made by chemists and physicists to optimize the active materials (Barber, 2009). Furthermore, an analogue to such a possible optimized system would have to be found in natural conditions not necessary a simple task. (b) Generally, the proposed systems are the result of elaborate chemical technologies. For instance: (i) to produce TiO2 nanoparticles, Suzuki et al. (2007) used natural TiO2 crystals of rutile and heated them to 150°C in a (very) concentrated NaOH solution (10 M); (ii) to obtain an efficient display of Co3O4 nanoparticles, Jiao and Frei (2009) included them in a special physical state of silica, mesoporous silica SB-15; (iii) Najafpour et al. (2010) prepared the CaMn2O4 catalyst in a sophisticated way, which included heating the different ingredients to 400°C and 600°C, respectively. While it is not completely excluded that natural conditions could lead to similar structures, especially given the very long timescales available, it is not apparent that they could. In particular, the active materials needed for the reactions may neither be abundant enough nor spread on a large enough fraction of the planetary surface. (c) The reaction implies that both the photosensitizing and the catalytic materials co-localize and function in concert, since in most cases there is no available material combining the two functions. (d) Most catalysts used in our chemical industry become poisoned after a period of time as the result of adsorption of trace species that bind to, and remain in, the active sites with high energy, which renders the site unavailable. Periodically, our catalysts must be regenerated by a proper treatment, or replaced. An abiotic photocatalyst in a planetary environment would have to work for a long period of time to build up an O2-rich atmosphere. The corresponding durations are of the order of the time required for continental surface modifications by plate tectonics (1–100 million years), which is 6–8 orders of magnitude longer than the lifetime of our industrial catalyst (typically 1 year). (e) Photocorrosion of material, a frequent process, should also be avoided during long durations (million years). Usually, this requires very specific treatments of the catalyst, even for much shorter times (years), for example, of those developed in the US Patent 4574039 (Pyke and Bruce, 1986).
If objection (a) is not prohibitive, it is difficult to imagine a natural solution that would simultaneously circumvent objections (b), (c), (d), and (e). To falsify a large abundance of O2 as a biosignature, it would be necessary to propose a system that can photolyze water with a regeneration process that turns its lifetime from 1 yr to 106 to 108 yr while occurring under natural planetary conditions on a significant fraction of the planetary surface. To date, we are not aware of such a system 3 .
5. O2 Flux Estimates on Earth
The biological photosynthesis production rate of O2 and its different sinks, oxidation of organics, and oxidation of minerals can be estimated on Earth (Léger et al., 1993; Kaltenegger et al., 2010). On an Earth-like exoplanet, an O2 production, either biological or abiotic, would have a similar fate.
The production of organic carbon by photosynthesis on Earth is estimated to be 1.1 × 1014 C-kg yr−1 by Field et al. (1998), which corresponds to an O2 production of 2.9 × 1014 kg yr−1, according to the stoichiometry of the reaction
where (CH2O) is the building block of an organic molecule, for example, a sugar.
Most of this O2 is consumed by the oxidation of organics from dead biomass (e.g., wood, humus, plankton) that was produced by the photosynthesis itself. The only net production corresponds to organic carbon that is withdrawn from contact with O2, by sedimentation in the oceans followed by subduction into the mantle and a possible re-emergence as rocks. On Earth, the main reservoirs of organic carbon are clay and shale sediments that contain ∼1% in mass of organics. The small fraction of O2 that avoids the organic sink is eventually trapped by rock oxidation (Fe2+ → Fe3+) and volcanic sulfide oxidation. The corresponding sink flux is 6.2 × 1010 kg yr−1. Then, the presence of O2 in an atmosphere (1.2 × 1018 kg for present Earth) requires a continuous and abundant production that compensates these sinks. If photosynthesis were to stop, the O2 content of our atmosphere would disappear within a characteristic time, the O2 residence time, of 2 × 107 yr (Léger et al., 1993). These different fluxes are reported in Table 2.
Most of the O2 produced by photosynthesis is used to oxidize the organic compounds it has produced. This translates into a flux of O2 trapped in biomass oxidation almost equal to that of the production by photosynthesis. The totals of the sources and the sinks (biomass oxidation and mineral oxidation) are equal, as expected in a stationary state, but the second sink is so small that it corresponds to a change of the fourth digit in the total flux, which does not appear in the two-digit values of the main fluxes. However, this second sink determines the residence time of O2 in the event that the photosynthesis production stops and so does the biomass oxidation.
6. A Darwinist Argument
In the mid-term future, it is possible that our astronomical instruments could detect in the atmosphere of an exoplanet the presence of O2 or O3, which could not be explained by the UV photolysis of H2O or CO2. This would require an O2 production at the planetary surface (Owen, 1980). The competing explanations would entail a biological photosynthesis or, if available at the time, an abiotic photogeneration process. The former exhibits a reinforcing advantage over the latter.
As described in Section 4, within our present understanding, the abiotic processes appear very unlikely. On the contrary, if a local biological photosynthetic system appears on a planet with the ability to synthesize the organic carbon it needs for growing, this would provide a major selective advantage, as this system would use abundant chemical species (H2O and CO2 4 ) and the main source of energy available on the planet. A Darwinist-type selection would allow it to dominate over other forms of life on that planet and progressively colonize most of the accessible niches, as chlorophyll-based life did on Earth. Two main features of a living system would be at work: its ability to evolve and its ability to replicate and invade accessible niches (Fig. 3). This would turn the difficult problem of the needed abundance of the abiotic system into the result of a natural law for the biological one. A comparable “appear and expand” character is difficult, although not impossible, to envision for an abiotic system.

Illustration of biological growth and invasion when in a favorable environment: simulation of bacteria colony growth on a nutriment-rich substrate with a model that fits the experiments (adapted from Lega and Passot, 2004, with the kind permission of the authors). Color version available in electronic file. Color images available online at
7. Conclusion
The qualification of the presence of O2 or O3 as a marker of life in specific physical conditions, for example, T surf < 50°C on a rocky exoplanet, depends on the failure of an alternative abiotic explanation for the observations. This also applies to the presence of chlorophyll-like molecules, the observation of specific changes in the features of a planetary surface with local seasons, and other such indicators. An abiotic catalytic photolysis of H2O and production of O2 is energetically possible on a planet irradiated by a solar-type star and, thus, cannot be excluded. However, an efficient photocatalyst is needed, and thus far the ones that are industrially produced are unlikely to exist under natural conditions on a planet.
On the other hand, a biological explanation for the presence of abundant O2 by invoking photosynthesis due to a living system presents an advantage. It could explain the required large abundance as the result of a feature common to all living systems: their ability to reproduce and colonize niches whenever the conditions are favorable.
Thus, the presence of O2 on a rocky exoplanet in the HZ of its star is at least a good, if not definitive, indication for the presence of life. Its reliability depends on identifying all possible abiotic photocatalysts. Among those known so far, we have found no good candidate for the large production that would be necessary to account for a possible abundant presence of oxygen in the atmosphere (P O2 ≥ 10 mbar).
The discovery of such a signature would be a major achievement, considering the scientific and philosophical implications of the presence of biological activity beyond Earth.
Footnotes
Acknowledgments
The authors are grateful to Luc Arnold, André Brack, Hervé Cottin, Olivier Grasset, Beate Orberger, and François Raulin, for providing several valuable pieces of information, and to Joceline Lega and Thierry Passot for discussions on bacteria colony growth and the kind permission to use their data, as well as two anonymous referees that contributed to significant improvements of the manuscript. They also thank the CNRS Pluridisciplinary Programme “Origine des Planètes et de la Vie” for its support.
Abbreviation
HZ, habitable zone.
1
O'Neill and Lenardic (2007) have started that study, but it cannot be considered as final (Valencia and O'Connell, ![]() ).
).
2
A spectral domain where the star-to-planet contrast is 3 orders of magnitude more favorable (weaker) than in the visible.
3
The micrometeorites (interplanetary dust particles), the most abundant input of extraterrestrial matter on present Earth, may be considered to provide a fresh and proper catalyst. With their flux (106 C-kg yr−1; Chyba and Sagan, ![]() ) and assuming the observed size of interplanetary dust particles (∼1 μm), it would be necessary that they stand the weathering of rains during 5 × 105 yr to produce a continuous covering of the planetary surface, an unlikely resistance to weathering by comparison to weathering timescales of resistant minerals.
) and assuming the observed size of interplanetary dust particles (∼1 μm), it would be necessary that they stand the weathering of rains during 5 × 105 yr to produce a continuous covering of the planetary surface, an unlikely resistance to weathering by comparison to weathering timescales of resistant minerals.
4
Within our present understanding of telluric planet formation, CO2 is expected to be present in all rocky planets that have an atmosphere, as it is the case for Mars, Earth, and Venus.