
Abstract
This paper addresses the open philosophical and scientific problem of explaining and defining life. This problem is controversial, and there is nothing approaching a consensus about what life is. This raises a philosophical meta-question: Why is life so controversial and so difficult to define? This paper proposes that we can attribute a significant part of the controversy over life to use of a Cartesian approach to explaining life, which seeks necessary and sufficient conditions for being an individual living organism, out of the context of other organisms and the abiotic environment. The Cartesian approach contrasts with an Aristotelian approach to explaining life, which considers life only in the whole context in which it actually exists, looks at the characteristic phenomena involving actual life, and seeks the deepest and most unified explanation for those phenomena. The phenomena of life might be difficult to delimit precisely, but it certainly includes life's characteristic hallmarks, borderline cases, and puzzles. The Program-Metabolism-Container (PMC) model construes minimal chemical life as a functionally integrated triad of chemical systems, which are identified as the Program, Metabolism, and Container. Rasmussen diagrams precisely depict the functional definition of minimal chemical life. The PMC model illustrates the Aristotelian approach to life, because it explains eight of life's hallmarks, one of life's borderline cases (the virus), and two of life's puzzles. Key Words: Nature of life—Definition of life—Program-Metabolism-Container (PMC) model—Rasmussen diagram. Astrobiology 10, 1011–1020.
1. Why Care about Minimal Chemical Life?
A
Why should scientists care about how to define minimal chemical life? Most biologists can safely ignore the question and happily focus their attention on certain model organisms. However, those studying the origin of life on Earth must have some prior conception or working hypothesis about what life is, because they need some way to choose which kinds of chemical systems to study. Similarly, astrobiologists who are seeking signs of “weird” life on Mars, Titan, or other planets and moons need some working hypothesis about what life is (National Research Council of the National Academies, 2007). The wide presumption that liquid water is necessary for life might be wrong (Benner et al., 2004), and extraterrestrial life might lack many familiar molecular signatures of life on Earth, such as DNA, ribosomes, or proteins (Rasmussen et al., 2003). In addition, those attempting to create synthetic life in the laboratory must have a working hypothesis about what life is (Szostak et al., 2001; Deamer, 2005; Luisi, 2006; Rasmussen et al., 2009a).1 How can synthetic cell scientists working to create life from nonliving materials choose what kind of chemical systems to create in the laboratory unless they are guided by some working hypothesis about life?2
2. Divergent Accounts of Life
One of the striking empirical facts that those interested in the nature of life must acknowledge is that there is no consensus about what life is, even among those scientists and philosophers who care about the question. Most natural candidate definitions have straightforward counterexamples (see, e.g., Sagan, 1970; Farmer and Belin, 1992; Luisi, 1998; Bedau and Cleland, 2010).3 So giving an account of life involves giving some explanation for the pluralism of different accounts of life.
Some would explain the pluralism of accounts for life by saying that living things share only a Wittgenstinian family resemblance. Different forms of life might share various characteristic properties, but each individual property in the cluster could be missing in some members of the family. The typical properties possessed by living organisms are not strictly necessary or sufficient. The cluster conception provides a natural explanation for borderline cases, because all clusters have vague borderlines. Cluster conceptions also provide a natural explanation for why life comes in various degrees, for things can have more or fewer of the properties in the cluster. But cluster conceptions cannot explain why forms of life are unified by one set of hallmarks rather than another. The cluster view must simply accept the hallmarks as given and then identify the cluster with those hallmarks. Thus, cluster views can identify life's hallmarks only post hoc; they cannot predict or explain the hallmarks. Those who think that there should be an explanation for life's hallmarks will therefore find the cluster conception unsatisfying.
Keller (2002) goes further and doubts that life is the sort of scientific kind that has a scientific definition, and she concludes that life is a human kind, rather than a natural kind. The view that life is a human kind easily explains borderline cases. Since the concept of life changes with the progress of science and technology, one should expect its boundaries to change, thus creating borderline cases. The view also provides some general ammunition against life's puzzles, for it is unsurprising (and also uninteresting) if a mutable human construct is confusing. New concepts generated by scientific and technological progress can be expected to violate older taxonomies, including the distinction between life and nonlife (Keller, 2002).
A direct and constructive response to Keller would be to figure out the biochemical universals that would apply to any form of life in the known universe. The physical prerequisites of minimal forms of life were first mapped out by Erwin Schrödinger (1946). Concerns from astrobiology have recently reinvigorated this kind of biochemical definition of life, when scientists attempt to specify the biochemical properties that any form of life must have (Pace, 2001; Benner et al., 2004). The biochemical universals of life include its thermodynamic limits, energetic limits, material limits, and even geographical limits. A biochemical definition always presupposes a prior account of life and describes the physical, chemical, and biological possibilities for any biochemical system that meets that prior account of life. The biochemical “definition” of life is really just a description of the biochemical consequences and constraints that any actual example of life must obey.
There is one more explanation that is sometimes given for the lack of consensus about what life is: the sample size of one. Biology generalizes about possible forms of life, and those generalizations rest on what we know about the forms of life that actually exist. Biologists study a number of different model organisms, like Escherichia coli (a common bacterium), Caenohabditis elegans (a nematode), and Drosophila melanogaster (a fruit fly). Picking model organisms that are as different as possible best illustrates the possible forms that life can take, and thus enables the widest generalizations about all life. But the evolution of all life on Earth still is only one large, interconnected evolving biosphere. Because all forms of life are part of the same evolving biosphere, generalizations about all life rest on a sample size of one.
The implications of the sample size of one can be overblown, though. Our one sample of life is not just “one” thing; it is a vast biosphere consisting of uncounted billions (or more) of species, each of which comprises uncounted millions (or more) of individual organisms. Furthermore, comparing these individuals with each other reveals complex similarities and differences, as there is quite a variety of forms of life. So this “one sample” of life is extremely heterogeneous and affords many opportunities to generalize. It is not as if we must generalize about all forms of life after examining just one single organism. Furthermore, progress in artificial life and synthetic biology could repeatedly increase the sample size. Anytime we can synthesize a new kind of system that exhibits life's characteristic properties, whether in wetware or hardware or software, we can expand our empirical understanding of what life is, at its most general. Similarly, astrobiology holds out the promise of expanding the sample size of life.4
3. The Aristotelian Approach
The deadlock of divergent accounts of minimal chemical life is striking. One recommendation for how to break this deadlock and adjudicate rival accounts of life is to focus on organisms in the natural context in which they carry out their lives, interact with their environment, and cooperate with other forms of life (Dupré and O'Malley, 2009), and to ask what underlying phenomena generate and explain the characteristic phenomena of life. Using broad historical license, we may call this recommendation the Aristotelian approach to explaining what life is.
Today, most people who wonder about what life is do not follow the Aristotelian approach but instead follow what (using historical license, again) we may call the Cartesian approach to explaining what life is. To figure out the essential properties of life, the Cartesian gathers a large and diverse set of paradigm instances of living beings (i.e., organisms) and asks what properties uniquely identify all and only living beings. The organisms in the set are considered in isolation from all other beings, out of the normal context in which they live their life. The Aristotelian addresses a different question: How should we explain the characteristic phenomena of life? Explaining life involves much more than defining what it is to be an individual living organism, for life's characteristic phenomena involve a significant number of different hallmarks, borderline cases, and puzzles.
This paper proposes that the Cartesian approach is responsible for the deadlock over accounts of life and that the Aristotelian approach provides more traction. According to the Aristotelian, accounts of life should be evaluated on their ability to explain the characteristic phenomena involving life (Bedau, 1998, 2007, 2010), and this amounts to doing at least three things: explaining life's hallmarks, explaining life's borderline cases, and resolving the puzzles about life. According to the Aristotelian approach, any account of minimal chemical life that can explain the deepest and most characteristic phenomena involving life is worthy of respect and probably has a role to play in any comprehensive account of life.
To ward off a possible misunderstanding, we should note that the Aristotelian and Cartesian are not concerned with the meaning of the word life or its translations in other languages. Anyone who does not know the meaning of the word can look it up in a dictionary, and everyone who is reading this paper knows the meaning of that word. Furthermore, the Aristotelian and Cartesian are not analyzing the conceptions of life currently held by various people around the globe today. Our concepts involve many preconceptions, and on a long enough timescale they continually change. So we have good reason to think that the concepts of life believed and presupposed today are also changing and developing. Thus, the Aristotelian and Cartesian are not trying to describe how we actually do think about life, but how we should think about it.
Let us dwell a bit further on what question is at stake. Some investigations think the key test for any account of life is to fit it with our pre-theoretic intuitions about which things are alive and which are not (e.g., Boden, 1999). But why should we defer to such intuitions? A good theory of life might prompt us to reconceptualize and recategorize life. This might change our attitudes and intuitions about exactly which things are alive. Thus, although our pre-theoretic intuitions carry some weight about the nature of life, the best account of life might reject and explain away some intuitions.
Scientific essentialism, originating from Kripke (1980), is the philosophical view that the essence of natural kinds like water and gold is their underlying causal powers, which are discovered by empirical science (see Bealer, 1987). The Aristotelian could adopt a scientific essentialist approach to life, too, and let empirical science explain life's underlying causal powers. It is in this spirit that this paper proposes the PMC model of minimal chemical life as an explanation of some of the underlying causal powers of minimal chemical life.
There is one more point to make about the Aristotelian approach. Aristotle is known in contemporary biology for his view that each species has a fixed, real essence. Contemporary biology typically abandons Aristotelian essentialism and instead embraces Darwinian anti-essentialism. According to Darwinian anti-essentialism, species have no fixed essence, and their members share no necessary and sufficient properties. There are real similarities among the members of a species, but they are only statistical.
The Aristotelian approach to minimal chemical life is consistent with Darwinian anti-essentialism, that is, the denial of Aristotelian essentialism about species. Unfortunate terminology aside, there is no paradox about this. The Aristotelian approach takes no stand on the nature of species; it simply identifies the nature of life with whatever is the best underlying explanation of the phenomena of life. Today, the best explanation of the phenomena of life would deny Aristotelian essentialism about species. Thus, Darwinian anti-essentialism about species is fully compatible with the Aristotelian approach to minimal chemical life.
4. The Phenomena of Life: Hallmarks, Borderline Cases, and Puzzles
The Aristotelian approach is to evaluate an account of life5 by how well it explains the characteristic phenomena of life. We can debate exactly what life's most characteristic phenomena are, but we can probably agree that they include life's familiar hallmarks, borderline cases, and puzzles. A representative list of life's hallmarks is provided by Gánti (2003). Gánti's list includes criteria for life in an individual living organism as well as criteria for populations of living organisms being able to populate a planet and sustain themselves:6
(1) Holism. An organism is an individual entity that cannot be subdivided without losing its essential properties. An organism cannot remain alive if its parts are separated and no longer interact. (2) Mortality. Living systems are mortal. This is true even of clonal asexual organisms, because death can afflict both individual organisms and whole clones. Systems that could never live cannot die, so death is a property only of things that were alive. (3) Active information. A living system stores information and uses it in its development and functioning. Children inherit this information, and copy mistakes can “mutate” the information, so natural selection can shape the resulting genetic variance in a population of organisms. (4) Metabolism. An individual organism takes in material and energy from its local environment and chemically transforms them. (5) Inherent stability. An organism preserves its overall structure and organization while living in a changing environment, by changing and adapting to the environment. (6) Flexible control. Processes in an organism are regulated in such a way that they promote the organism's continued existence and flourishing. This control is adaptive and flexible and often improves with experience. (7) Growth and reproduction. Post-reproductive or sterile animals and plants are living but cannot reproduce, so the capacity to reproduce is not necessary for life. But a population of organisms can survive and flourish only if enough organisms in the population reproduce. (8) Evolvability. Living systems can be members of a population with the capacity to evolve. It is an open question today exactly which kinds of evolving populations have the capacity to generate increasing complexity and differentiation.
Life's hallmarks are strikingly heterogeneous. It is unclear what, if anything, unifies them. Are some central and others peripheral? Can some be combined? Any adequate account of life should give some kind of explanation for life's hallmarks.
In addition to life's hallmarks, the phenomena of life include its characteristic borderline cases. Prominent examples include viruses and prions, which self-replicate and spread even though they lack a metabolism. Dormant seeds or spores are a different kind of borderline case and are more like frozen bacteria or insects. Some clearly nonliving things have some chemical characteristics of life. A candle flame preserves its form while its constituent molecules are constantly changing, so it has something like a metabolism (Maynard Smith, 1986). Populations of microscopic clay crystallites growing, proliferating, and undergoing natural selection are another kind of borderline case; they seem not to be alive, though they evolve by natural selection (Bedau, 1991). So a second important aspect of the phenomena of life is its borderline cases,7 which any adequate account of life should explain.
The phenomena of life also include a number of characteristic puzzles, which any complete account of life should explain and resolve. Some puzzles might result just from confusion, but others probe deep open questions about a fundamental and fascinating part of the natural world. Here are some examples of puzzles about life that are most relevant to the PMC model. All such puzzles are included in the characteristic phenomena of life. (1) Origin of life. How does life or biology arise from nonlife or pure chemistry? How does a system undergoing merely chemical evolution, in which chemical reactions are continually changing the concentrations of chemical species, differ from one that is alive? (2) Emergence of life. Life is said to “emerge” from nonlife when the properties of life both depend on, and are autonomous from, the properties of nonlife (Bedau, 2003). The life properties arise over time from the nonlife properties; the emergence of life from nonlife is dynamic. The puzzle of emergence is to explain how life emerges from nonlife in a way that combines the scientific state of the art with a philosophically plausible account of emergence. (3) Degrees of life. Can things be more or less alive? Is life a black-or-white Boolean property or a continuum property with many shades of gray? Common sense leans toward the Boolean view: a rabbit is alive, and a rock is not. But there are borderline cases like viruses, which are unable to replicate unless they infect a host. And spores or frozen bacteria remain dormant and unchanging indefinitely but then come back to life when conditions become favorable. Are viruses and spores really alive or only potentially revivable? Furthermore, the original life-forms differed very little from their nonliving predecessors, and some conclude that there is no sharp boundary between life and nonlife but rather a continuum of things that are more or less alive (e.g., Cairns-Smith, 1985; Emmeche, 1994; Dennett, 1996). So is the distinction between life and nonlife a dichotomy or a matter of degree? (4) Matter, form, and function. Each individual life-form is a material object: a cell or organism, or the like. And being alive requires certain kinds of materials and sets various constraints on the material components of life-forms. But in a way the materials seem inessential. They are transitory, the molecules in your body continually cycling and recycling with material in the environment. What matters, apparently, is the functional organization of those molecules. In other words, the defining feature of life is not (only) the material out of which it is constituted, but (also) the form in which that material is arranged and organized. The form of a living system is the functional network in which those materials interact. (5) Meta-puzzle: why is life so puzzling? There is one more puzzle that was raised in section 2 above and deserves mention: a meta-puzzle about why life is so puzzling. Life seems to be a very basic and fundamental part of the natural world, but it is certainly very complex; and there is nothing even approaching a consensus opinion about what life is, by professionals and experts who study and work with life. Any full account of life should clarify why life is so puzzling.
To summarize: the Aristotelian approach to explaining minimal chemical life is to seek the best underlying explanation for the phenomena of life. The phenomena of life include, among other things, Gánti's eight specific hallmarks of life, borderline cases like viruses, and five general puzzles about life. The next step is to describe a concrete model of minimal chemical life—the PMC model. Then we will examine how well the PMC model explains the phenomena of life.
5. The Program-Metabolism-Container (PMC) Model
In the community of scientists who are trying to make “protocells” or artificial cells from nonliving materials, everyone admits how controversial and contentious the best accounts of life are. Nevertheless, minimal chemical life is generally considered to be a certain kind of complex chemical system that integrates three chemical functionalities in such a way that they mutually support each other (e.g., Rasmussen et al., 2004, 2009b). That is, an artificial cell is viewed as any chemical system that integrates three functionalities. The first functionality is a chemical program that influences and directs the important chemical processes in the cell. This program causally influences the system's behavior. The information in this program is stored inside the living system itself, encoded in informational molecules (“genes”). Errors (“mutations”) can corrupt this information and be inherited in successive generations, so a population of these systems can evolve by natural selection. The second chemical functionality is to extract free energy from the system's environment and digest environmental resources in order to regenerate the materials it needs to maintain and repair itself, to grow, and ultimately to reproduce. The third functionality is to maintain an identity over time by localizing all components, concentrating reagents, and protecting proper chemical operations from molecular parasites and poisons, inside a protective container such as a lipid vesicle.
The PMC integrated-triad model stipulates that program, metabolism, and containment support enable each other so that there is functional interaction connecting all three. The triad exhibits holistic autonomy, because the component functionalities (programming, metabolism, and containment) and their chemical integration are created and sustained solely by the operation of the system itself, without the interventions of any external agency. For this reason, the system is responsible for its own continual existence and proper functioning.
In the PMC model, the terms program, metabolism, and container are to be understood functionally and with minimal constraints on their chemical realization. The program might achieve informational control and inheritance without DNA or RNA or even without any other biopolymers (e.g., Sagre et al., 2000). Similarly, a metabolism could harvest redox energy or get energy from light; it could be supplied with complex material precursors or ATP and complex enzymes. Often containers are amphiphilic structures such as a vesicle or micelle (Rasmussen et al., 2009a). Because the PMC model is functional rather than material, it accepts any biochemical material realization of the PMC triad as a genuine example of minimal chemical life. A functional account of life automatically has broad scope, because any chemical system with the right functional structure is an example. The gory nonfunctional chemical details can vary wildly from example to example. The laws of physics and chemistry constrain which kinds of materials can achieve which kinds of functions (Pace, 2001; Benner et al., 2004), but any material realization of the right functional details within those constraints will be alive.8
As indicated above, the functional PMC model is the rough consensus in the protocell research community that is trying to assemble life in the laboratory from nonliving materials (Rasmussen et al., 2009a). This community uses the PMC model as a working hypothesis about what life is, which suggests which kinds of experiments to pursue. No actual manmade chemical construction has yet met the full PMC conditions of minimal chemical life; and, as we learn and construct more systems in the laboratory, the details of the PMC model will be revised and augmented. The PMC model of minimal chemical life is chemically detailed and connected with contemporary protocell experiments. The process of constructing and defending provisional models helps us to improve the models (Wimsatt, 1987). Similarly, perhaps the PMC model can help improve our understanding of minimal chemical life.
The PMC model of minimal life entails that the program, metabolism, and container are not merely spatially contiguous. They must be chemically integrated and mutually support each other in such a way that the continual operation of each component functionality depends on support from each of the other two functionalities. The integrated triad is a kind of chemical cooperation.9
The key types of chemical cooperation involved in minimal living systems are depicted in Rasmussen diagrams (Rasmussen et al., 2009b). These diagrams distinguish 10 kinds of elementary chemical components: C—container P—stored information program E—energy harvester E*—energized form of E F—fuel M, L—material precursors of P, C pM, pE, pL—precursors of M, E, L
Rasmussen diagrams also distinguish five kinds of functional dependence with five kinds of arrows and enclosures: thin black arrows for chemical reactions; thick red double arrows for self-assembly processes; blue circle dotted arrows for chemical catalysis; green diamond dotted arrows for energy transfer; and yellow dotted enclosures for spatial concentration.
Arrows point to other arrows when they catalyze a chemical reaction. The number of different Rasmussen diagrams is vast. Rasmussen diagrams depict chemical substances and processes purely functionally. No particular chemical substances or processes are specified; only general functional types are required. Any number of different chemical substances and processes might fit the same Rasmussen diagram.
Figure 1 shows components of Rasmussen diagrams. A Rasmussen diagram represents a program, metabolism, or container with a small chemical network, in which precursor molecules are chemically transformed into components of larger structures (such as stored information programs, P, or containers, C) and in which chemical cycles of different types represent replication, reproduction, or energy flow.
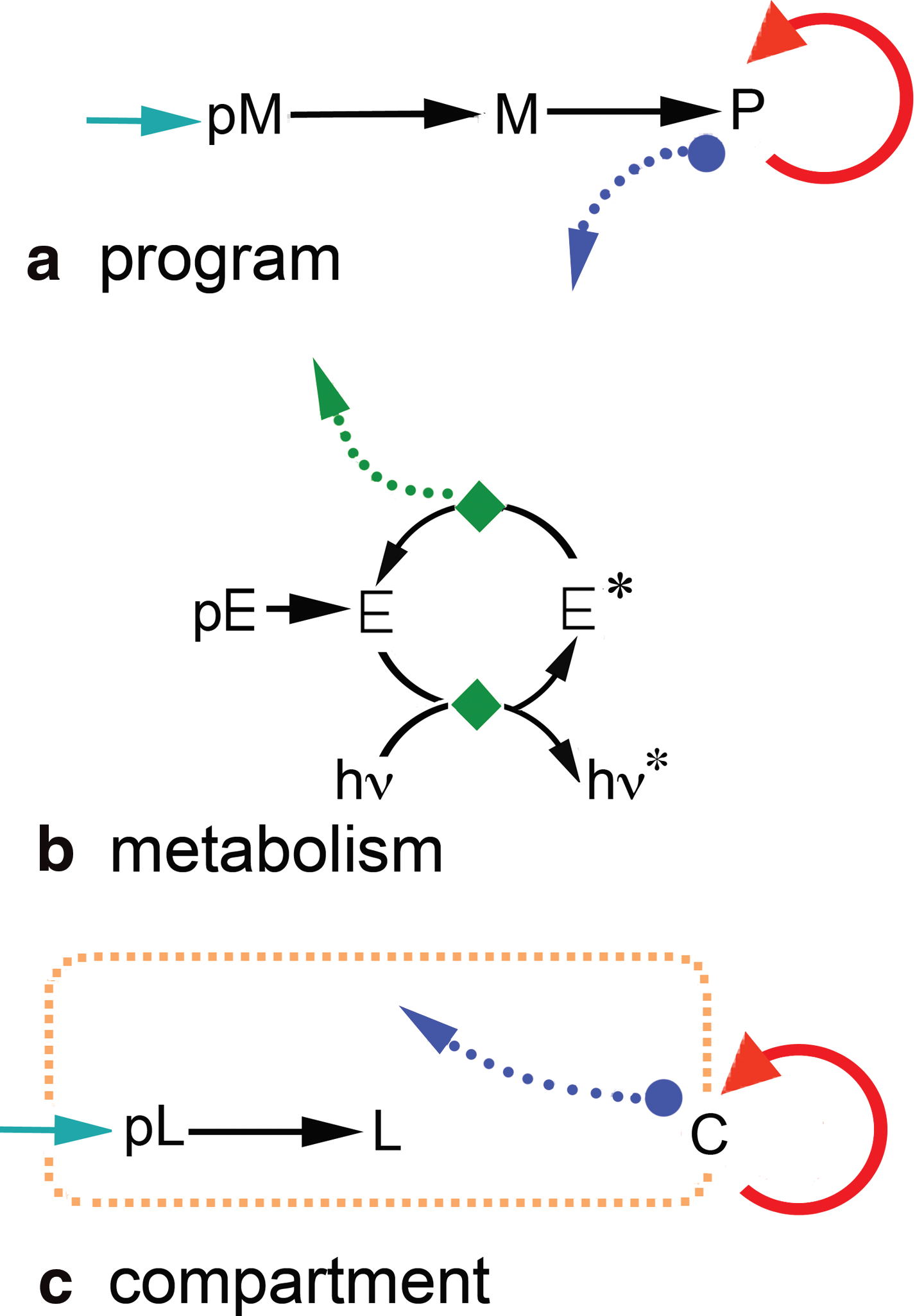
The three chemical subsystems in minimal chemical life, represented by Rasmussen diagrams (Rasmussen et al., 2009b). Rasmussen diagrams represent minimal chemical life as a chemically integrated system with three main parts. (
Rasmussen diagrams typically combine two or three components from the chemical triad of program, metabolism, and container. The Rasmussen diagram for the paradigm of a chemical system that has all the requirements for being a minimal form of chemical life consists of a program, metabolism, and container that functionally support each other (Fig. 2). The program is generated from monomers, and it replicates by a complex and well-known process involving self-assembly. The metabolism extracts energy and materials from the environment and delivers them to the system's program and container. And the container concentrates and protects the metabolism and program and also catalyzes the production of the building blocks of the program, metabolism, and container.

Rasmussen diagram of a protocell, that is, a minimal mutually self-supporting network of chemical interactions including a program, metabolism, and container. Rasmussen diagrams represent the lower-level chemistry that makes protocells possible. We see a self-replicating program, P, that catalytically controls other protocell chemical functioning; a metabolism, E*, that makes light energy from the environment available for protocell chemical functioning; and a reproducing container, C, that concentrates and protects protocell chemical functioning. These three chemical functionalities (P, M, and C) are chemically integrated by catalytic and energetic support (dotted arrows). Color images available online at
The rough consensus in the protocell community gives the functional triad view some credibility, but not enough to show that the PMC model is true. A full explanation and defense of the PMC model of minimal chemical life is beyond the scope of this paper, but we do briefly discuss a couple of further open questions. One concerns the PMC model's exclusive focus on containment, metabolism, and program. Are each of those three functionalities really necessary for life? Will any pair suffice to explain the other? Can something deeper explain the triad? Such questions naturally arise, especially because life does seem to involve various other functionalities, such as autonomous behavior, autonomous information processing, responsiveness to the environment, self-interested drives, and purposeful behavior. Why does the PMC model focus only on containment, metabolism, and program?
Supporters of the PMC model can acknowledge that other functionalities like autonomous information processing and sensitivity to the environment are essential to living systems if those functionalities can be explained by the PMC model. The PMC model can at least partly explain why minimal living systems have properties like autonomy, autopoiesis, and self-reproduction. For example, each of the three systems in the PMC triad (Fig. 2) is continually recreated and sustained by its own operation and by that of the two other parts of the triad, so the triad continually recreates and sustains its own operation and structure. The triad has an autonomous drive to survive and the means of self-regulation required to achieve this goal. Furthermore, autopoiesis (Maturana and Varela, 1980) involves a somewhat similar kind of autonomy that comes from self-maintenance, and the PMC diagram depicts the main kinds of functional support that bring about this autonomous self-maintenance. Some of the simplest autopoietic systems involve only metabolism and containment and have no genetic program whatsoever; their PMC diagrams have functional support only between C and M. A second kind of autonomy is present in minimal chemical life (Fig. 2), from the genetic program, P, gives the system control over its own behavior. No doubt there are further kinds of autonomy that can be identified in PMC diagrams. The natural way in which Rasmussen diagrams represent several different kinds of autonomy in minimal chemical life further supports the PMC model.
6. The PMC Model Explains the Phenomena of Life
The Aristotelian methodology evaluates accounts of life by seeing how well they explain the phenomena of life (i.e., the eight hallmarks of life, borderline cases, and five puzzles about life). The PMC model is a concrete, chemically detailed, scientifically illuminating model of minimal chemical life. This section shows how the PMC model explains or fits with explanations of many of the hallmarks, borderline cases, and puzzles about life. The examples have two purposes. The first is to illustrate how an account of life could explain the phenomena of life and, thereby, help clarify the Aristotelian approach. The second purpose is to illustrate the scientific support for the PMC model.
First, notice that the PMC model has a natural and plausible explanation for life's eight hallmarks: (1) Holism. The PMC model depicts minimal chemical life as a specific kind of autonomous, self-sustaining holistic system of chemical interactions (depicted in Fig. 2). This holistic system will not sustain itself if its parts cannot interact (if the catalytic and energetic arrows are dropped). (2) Mortality. PMC systems are self-supporting functional systems, so the whole system will fall apart and “die” unless the parts remain correctly functionally connected. (3) Active information. The P in PMC is precisely an active information system that programs and supports many of the living cell's important chemical processes. This information can mutate, so natural selection can shape a population of PMC systems. (4) Metabolism. The M in PMC is metabolism, signifying its central role in minimal chemical life. (5) Inherent stability. A properly functioning PMC system preserves its overall structure and organization in a changing environment by adapting to environmental fluctuations. It is an open question exactly which PMC systems achieve this stability. (6) Flexible control. The programs in PMC systems control many of the system's important holistic emergent properties, including those that enable the system to continue to exist and flourish, though it is an open question exactly which programs accomplish flexible control. (7) Growth and reproduction. Individual PMC systems can grow until they are so large that they spontaneously divide and thereby reproduce, although it is an open question exactly which PMC systems have these capacities. A population of PMC systems could survive and flourish if enough of them grow and divide quickly enough. (8) Evolvability. A population of PMC systems could evolve, because the individual systems can reproduce, and pass on traits that affect the ability of the daughter systems to survive and reproduce. Exactly which PMC systems could form populations that could undergo open-ended evolution is an important open question.
So the PMC model has a natural and unified, though sometimes tentative, explanation for many of life's heterogeneous hallmarks—one part of the phenomena of life. This is one piece of support for the PMC model.
Life's characteristic borderline cases are a second important part of the phenomena of life. Elsewhere, Bedau (2010) detailed the case that the PMC model provides a natural explanation for the virus borderline case. In a nutshell, the argument is that Rasmussen diagrams clearly depict the important differences between an inert virus particle and a virus that is infecting a host. The inert virus particle has the Rasmussen diagram of an isolated program, P, which is not replicating and is not being replenished or repaired. Most people would agree that this complex molecule by itself (or even housed within a protein coat) is clearly not alive, though it is a central functional component (the program) in the Rasmussen diagram for minimal chemical life. The Rasmussen diagram for the infected host, on the other hand, would be a more complex version of the Rasmussen diagram for minimal chemical life (Fig. 2). In addition to the program, P, this more complex diagram would depict a second program, P* (and perhaps a second container, C*, for the virus's newly constructed coat). The second program, P*, receives functional support from the original PMC system, but it does not provide any support in return; that is, it is a parasite. (The same holds for the second container, if any.) This Rasmussen diagram for the infected host would contain the original Rasmussen diagram for the PMC host, as a proper part of the whole chemical system. But the whole chemical system itself could be considered as one larger living system with various living parts. So the PMC model has a natural explanation of why, and in what respect, viruses inhabit the space between nonlife and life, once the inert and infectious states of a virus are appropriately mapped. The way that the PMC model naturally locates viruses on the border between life and nonlife indirectly supports the PMC model. This gives a flavor of how the PMC model can explain some of life's other borderline cases.
To complete its explanation of the phenomena of life, the PMC model must still explain life's characteristic puzzles, listed above, which we now consider in turn: (1) Origin of life. The PMC model provides a natural conceptual framework for describing the stages by which a nonliving chemical system gradually becomes more and more alive. Essentially, the process involves adding more and more of the chemical details in the Rasmussen diagram for minimal chemical life (Fig. 2), until the entire picture is present. Life differs from nonlife in the specific chemical complexity depicted in Fig. 2. (2) Emergence of life. Minimal chemical life is the paradigm case of a system with weakly emergent properties, which are caused by especially complex molecular mechanisms. See Bedau (1997, 2003) for further discussion. (3) Degrees of life. The PMC model gives a natural explanation for how minimal chemical life is a matter of degree (Bedau, 2010). Briefly, the space of all possible Rasmussen diagrams can be ordered by counting the number of components and arrows in the diagram. A local neighborhood can be defined, so that Rasumssen diagrams of immediate neighbors differ by only one component or arrow. Then a distance metric is defined for the whole space of Rasmussen diagrams. The Rasmussen diagram for minimal chemical life (Fig. 2) has about 29 components (depending on exactly how you count). If we count all the Rasmussen diagrams that do or do not contain each component, we have 229 ≈ 5 × 108 different diagrams (a tiny subset of these Rasmussen diagrams are shown in Fig. 3). It seems arbitrary to single out any subset of those diagrams and surround them by a bright red line, saying that those inside are “alive” and those outside are not. Instead, it makes more sense to admit that minimal chemical life comes in degrees. See Bedau (2010) for further discussion. (4) Matter, form, and function. Each individual life-form is a complex chemical object. And being alive requires certain kinds of materials; it sets various constraints on the material components of life-forms. But, in a way, the materials seem inessential. For one thing, they are transitory; the molecules in your body are continually cycling and recycling with material in the environment. What matters, in general, is the functional organization that is realized by combinations of those molecules. In other words, the defining feature of life is not the material out of which it is constituted but the form in which that material is arranged and organized. The form of a living system is the functional network in which those materials interact. This formal, functional perspective on life supports all functional approaches to life. The PMC model is one such functional approach. (5) Why is life so puzzling? The PMC model itself can somewhat help explain why life is so puzzling, because the PMC model construes life as a certain kind of complex chemical system, and this complexity in itself will generate some puzzles. More important, the model is consistent with an independent explanation of why life is so puzzling: because people give the question a Cartesian rather than Aristotelian interpretation. The Cartesian seeks necessary and sufficient conditions for being an individual living organism, independent of any particular environment. The Aristotelian seeks the most plausible, comprehensive, and unified underlying explanation for the characteristic phenomena associated with life; this is a complex task that involves explaining a number of hallmarks, borderline cases, and puzzles.
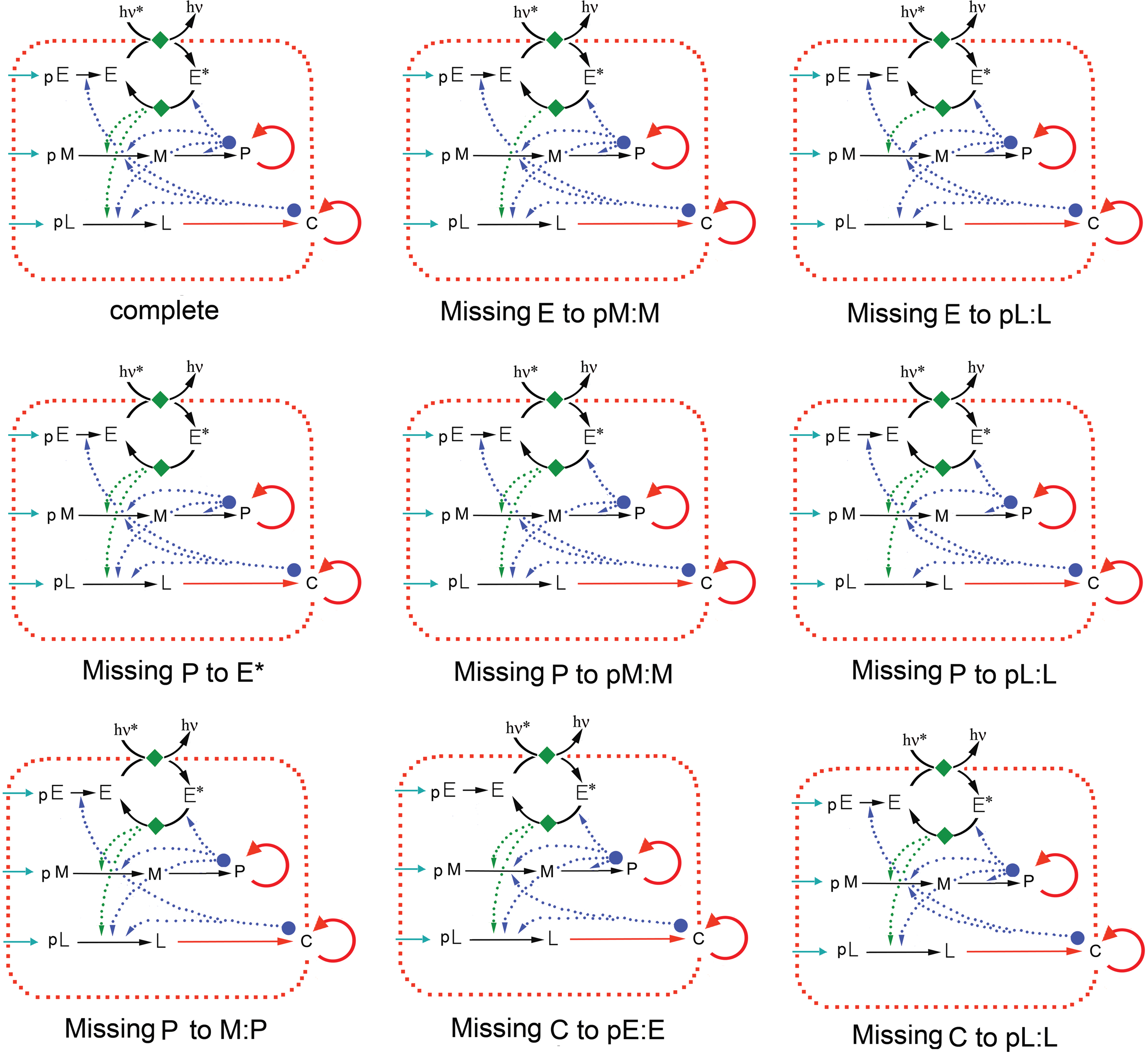
Eight minimally different Rasmussen diagrams, each of which differs from the representation of minimal chemical life (top, left; see also Fig. 2) by omitting exactly one catalytic or energetic support (i.e., one dotted arrow). It takes close scrutiny to detect the differences between these diagrams. If the diagram for minimal chemical life represents a system that is fully alive, then each of these kinds of systems is at least almost fully alive. Color images available online at
7. Conclusion
The overall argument of the paper can now be summarized as follows: The phenomena of life consist of life's hallmarks, borderline cases, and puzzles, and the Aristotelian approach identifies life with the underlying explanation of these phenomena. Furthermore, it turns out that the PMC model helps provide, or is at least consistent with, a natural and plausible explanation of the phenomena of life. To show this involves examining each of the eight hallmarks, explaining a classic borderline case, and resolving five puzzles. So the PMC model has some significant plausibility among the contenders for general accounts for minimal chemical life.10 An alternative to the PMC model is a viable contender only if it provides an equally adequate account of the phenomena of life. Furthermore, the PMC model provides a detailed, concrete example of how an account of life could meet the Aristotelian standard. As part of a virtuous circle, this exercise demonstrates the plausibility of the Aristotelian approach to explaining what life is.
Constructing minimal chemical forms of life in the laboratory requires one to target some particular kind of chemical systems. The PMC model captures the current research target in the protocell community: systems that chemically integrate containment, metabolism, and internal informational control and thereby are able to maintain their existence, grow, reproduce, and in the long run evolve. As far as anyone knows, a population of reproducing and chemically mutating PMC systems could start to evolve by natural selection. In fact, PMC systems are the simplest chemical systems that can sustain themselves and grow and reproduce. It is one more feature in favor of the PMC model that the model naturally reflects how life and evolution are deeply connected.
Footnotes
Acknowledgments
Author Disclosure Statement
No competing financial interests exist.
Abbreviation
PMC, Program-Metabolism-Container.
1
The creation of a “synthetic cell” by Venter's team (Gibson et al., 2010) does not raise any special questions about the nature of life, nor does it particularly shed any light on what it means to be alive. The cell is only 1% synthetic; only its genome is synthetic. A natural living bacterium internalized and expressed the synthetic genome. But the synthetic cell is fully alive; it is just as alive as the natural living cell from which it derives. The synthetic cell's living state is simply inherited from the natural living state of the bacteria that Venter's team manipulated. Things would be different for a 100% synthetic cell or protocell (Rasmussen et al., ![]() ). Making 100% synthetic life would involve making a living chemical system out of components that are not themselves alive and did not come from something that ever was alive. Pursuing synthetic life does require guidance from a working hypothesis about life, and actually creating new forms of life in the laboratory would significantly illuminate what life is by increasing the sample size of forms of life.
). Making 100% synthetic life would involve making a living chemical system out of components that are not themselves alive and did not come from something that ever was alive. Pursuing synthetic life does require guidance from a working hypothesis about life, and actually creating new forms of life in the laboratory would significantly illuminate what life is by increasing the sample size of forms of life.
2
Another motivation for reconsidering the definition of life comes from artificial life software systems that attempt to synthesize or simulate systems with life's essential properties (Bedau, 2003). Artificial life has created remarkably lifelike software systems, which seem genuinely alive to some (Langton, 1989). Recent artificial life achievements in hardware include the domestic vacuuming robot, Roomba (Brooks, ![]() ). Such robots inevitably raise the question as to whether a device fashioned out of plastic, silicon, and steel could ever literally be alive.
). Such robots inevitably raise the question as to whether a device fashioned out of plastic, silicon, and steel could ever literally be alive.
3
There are additional interesting explanations for why it is so hard to say what life is. For example, Shields (![]() ) proposes a complex, Aristotelian “focal” meaning account of life, which could be argued to predict difficulty with defining life. Dennett thinks that we should expect minimal chemical life to be a matter of degree because life arose from nonlife by a succession of small, incremental steps of increasing chemical complexity. This approach explains life by appeal to “a finite regress, in which the sought-for marvelous property (life, in this case) was acquired by slight, perhaps even imperceptible, amendments or increments” (1996, p 200). The perspective on degrees of life defended in this paper is like Dennett's perspective, except that this paper gives a concrete and chemically detailed explanation for how to measure degrees of life, and my degrees of life are not mere epistemological “stances” but reflect the real, objective, chemical structure of the systems in question.
) proposes a complex, Aristotelian “focal” meaning account of life, which could be argued to predict difficulty with defining life. Dennett thinks that we should expect minimal chemical life to be a matter of degree because life arose from nonlife by a succession of small, incremental steps of increasing chemical complexity. This approach explains life by appeal to “a finite regress, in which the sought-for marvelous property (life, in this case) was acquired by slight, perhaps even imperceptible, amendments or increments” (1996, p 200). The perspective on degrees of life defended in this paper is like Dennett's perspective, except that this paper gives a concrete and chemically detailed explanation for how to measure degrees of life, and my degrees of life are not mere epistemological “stances” but reflect the real, objective, chemical structure of the systems in question.
4
Some people recommend that we should account for life only after we have amassed a much larger sample of different forms of life upon which to base our generalizations. But an epistemological paradox afflicts this recommendation; to follow this recommendation you need to use some prior conception of life when you judge whether some new system increases the sample size of life. So you cannot ask us to generalize about life only after we find many samples of life, because our preconceptions about life are generalizations that we use to identify the samples of life. A concrete example of this paradox arises for astrobiologists who are trying to recognize “weird” and unfamiliar forms of life (National Research Council of the National Academies, ![]() ). How can astrobiologists identify “weird” life-forms unless they employ at least an implicit preconception about what life is in general?
). How can astrobiologists identify “weird” life-forms unless they employ at least an implicit preconception about what life is in general?
5
Accounts of life include theories or definitions of life.
6
Various hallmarks of life have been identified in the literature (see, e.g., Mayr, 1982, 1997; Koshland, 2002; Gánti, 2003), and any adequate theory of life should explain why this heterogeneous list of interesting properties characterizes all and only things that are alive. Elsewhere (Bedau, ![]() ), I argued that the theory of life as supple adaptation explains one of Mayr's lists of life's hallmarks.
), I argued that the theory of life as supple adaptation explains one of Mayr's lists of life's hallmarks.
7
There are further borderline cases. Super-organisms, such as eusocial insect colonies, function like a single organism, and some biologists think of them as alive. Artificial life software systems are another kind of borderline case. For example, Tierra software creates a spontaneously evolving population of computer programs that reproduce, mutate, and evolve in computer memory, and Tierra's inventor thinks it is alive (Ray, 1992), though this conflicts with the preconceptions most of us have about life. Another borderline case consists of complex adaptive systems found in the social and economic sphere, such as financial markets or the social networks on the World Wide Web. These exhibit many of the hallmarks of life (Bedau, 1996, ![]() ), but most people would deny that they are literally alive.
), but most people would deny that they are literally alive.
8
There is a vigorous literature about functionalism with regard to artificial life software systems, like Ray's Tierra (Ray, 1992). Langton (1989) famously argued that computer instantiations of life are still genuine examples of life, and Sterelny disagreed (![]() ). The chemical functionalism that I advocate in this paper is orthogonal to this earlier debate about artificial life software, because the chemical systems that instantiate PMC diagrams are real, wet, chemical systems in the laboratory, not merely computer programs.
). The chemical functionalism that I advocate in this paper is orthogonal to this earlier debate about artificial life software, because the chemical systems that instantiate PMC diagrams are real, wet, chemical systems in the laboratory, not merely computer programs.
9
The containers can act as catalysts in various ways; for example, the thermodynamic conditions within a lipid aggregate or at the water-aggregate interface, which differ from those within bulk water, can be catalytic. Also, the program in a PMC system can support the triad in multiple ways; two typical examples are when biopolymer sequences or structures have catalytic properties or are used as structural material in a container. A third example of chemical cooperation is when the production of the building blocks of containers and programs feeds on an external energy source by means of a metabolism. In each of these examples, one of the three components of a PMC system is supporting the operation of other components.