
Abstract
Lipid biomarkers are widely used to study the earliest life on Earth and have been invoked as potential astrobiological markers, but few studies have assessed their survival and persistence in geothermal settings. Here, we investigate lipid preservation in active and inactive geothermal silica sinters, with ages of up to 900 years, from Champagne Pool, Waiotapu, New Zealand. Analyses revealed a wide range of bacterial biomarkers, including free and bound fatty acids, 1,2-di-O-alkylglycerols (diethers), and various hopanoids. Dominant archaeal lipids include archaeol and glycerol dialkyl glycerol tetraethers (GDGTs). The predominance of generally similar biomarker groups in all sinters suggests a stable microbial community throughout Champagne Pool's history and indicates that incorporated lipids can be well preserved. Moreover, subtle differences in lipid distributions suggest that past changes in environmental conditions can be elucidated. In this case, higher archaeol abundances relative to the bacterial diethers, a greater proportion of cyclic GDGTs, the high average chain length of the bacterial diethers, and greater concentrations of hopanoic acids in the older sinters all suggest hotter conditions at Champagne Pool in the past. Key Words: Extremophiles—Silica sinters—Lipid biomarkers—Archaeol—Bacterial diethers—Fatty acids. Astrobiology 11, 259–274.
1. Introduction
M
Current astrobiological and origin-of-life investigations focus on the detection of microbial remains preserved in rocks. Indeed, the ability to recognize the signature of life in rocks from Earth's fossil record as well as in extraterrestrial materials is one of the primary goals in the NASA Astrobiology Roadmap (Des Marais et al., 2008). Geothermal systems are commonly host to silica deposits, which form rapidly and often preserve a chemical signal of the spring inhabitants. As such, silica sinters and their associated microbiology have been studied in a range of hot springs in Yellowstone National Park, USA (e.g., Jahnke et al., 2001; Blank et al., 2002; Guidry and Chafetz, 2003), Krisuvik, Iceland (e.g., Schultzelam et al., 1995; Konhauser et al., 2001) and the Taupo Volcanic Zone, New Zealand (e.g., Jones et al., 2001; Mountain et al., 2003). DNA and RNA in these settings are typically poorly preserved, and progressive silicification can destroy morphological details, which makes identification of fossilized microorganisms in geothermal deposits difficult (Jones et al., 1997).
Lipid biomarkers, which are characterized by a wide variety of hydrocarbon structures, serve as powerful tools in the characterization of microbial community structure in diverse environments. Their structures have been well preserved throughout geological time, such that they have been extensively utilized as indictors of past biological activity on Earth (e.g., Summons et al., 1996; Freeman, 2001; Hayes, 2001; Simoneit, 2002) and in some cases used to characterize past microbial communities (e.g., Thiel et al., 2001; Peckmann and Thiel, 2004; Brocks et al., 2005; Birgel et al., 2006). Recently, it was shown that lipid biomarkers appear to be well preserved in geothermal sinters, with rapid silicification aiding geochemical preservation (Pancost et al., 2005, 2006; Talbot et al., 2005; Gibson et al., 2008; Kaur et al., 2008), and it was proposed that such compounds, once encased in the silica matrix, could persist for extended periods of time. Since the structure and distributions of the membrane lipids reflect the chemical and microbiological conditions present during the time of sinter formation, it follows that these compounds could be used to assess past spring chemistry and microbiology. For example, monomethyl alkanes record the presence of cyanobacteria (Shiea et al., 1990); archaeal lipids, specifically glycerol dialkyl glycerol tetraethers (GDGTs), are predominant at high temperatures and low pH (Robertson et al., 2005), and longer-chain fatty acids and saturated components are more abundant at higher temperatures (Weerkamp and Heinen, 1972; Russell, 1984; Zeng et al., 1992).
This paper focuses on the lipids preserved in both active and inactive silica sinters from Champagne Pool, a geothermal spring located in the Taupo Volcanic Zone (TVZ), New Zealand, and expands on results published in our previous paper, Kaur et al. (2008). Active sinters are defined as those precipitating at the air-water interface, whereas inactive sinters are those that are no longer precipitating in the spring. These inactive deposits would have initially formed at the pool edge, and, as a result of subsequent deposition of silica and contraction of the spring over time, they are now in non-geothermal settings and no longer precipitating. Thus, they have the potential to record past geothermal conditions. Accordingly, this work has three primary goals: (1) assess microbial lipid preservation in active geothermal silica sinters and their ability to record extant microbial populations; (2) utilize lipid profiles to assess the preservation of lipids in inactive sinter and investigate past variations in microbial community structure and environmental conditions; and (3) identify novel biomarker-based tools that may have particular chemotaxonomic potential or offer insight into survival strategies in such extremes.
2. Experimental Methods
2.1. Sample details
The TVZ is situated centrally on the North Island of New Zealand. Up to 60 km wide and extending approximately 300 km from the Tongariro and Ruapehu volcanic centers on the North Island to the active White Island volcano located 50 km offshore, the TVZ is the largest and most active rhyolitic magmatic system on Earth (Houghton et al., 1995). This extensive volcanism is coupled with several high-temperature (>250°C) geothermal systems, including the Waiotapu geothermal system in which Champagne Pool is located. The pool occupies a hydrothermal explosion crater formed approximately 900 years ago (Lloyd, 1959). It is approximately 60 m in diameter, 150 m in depth, and has a surface area of 3000 m2. The spring water is anoxic and of a mildly acid chloride type, with a pH of 5.5 and a constant temperature of approximately 75°C. This slight acidity is partly due to the upward flow of dissolved CO2 from the deep geothermal aquifer. The spring has a high gas flux, largely CO2 (82.1%) with relatively high H2S concentrations (7.2 %) (Phoenix et al., 2005). It is supersaturated with respect to amorphous silica (430 mg kg−1 SiO2; Mountain et al., 2003) and contains a wide array of trace elements, including Au, Ag, Sb, W, and As (Jones et al., 2001; Table 1). Above the air-water interface, spicular sinters surround the pool. These are predominantly composed of amorphous silica that occurs as porous and nonporous laminae, the former containing silicified filamentous and nonfilamentous microbes (Mountain et al., 2003).
In total, 16 samples, CPa1–CPa16, were collected from this system (Table 2; Fig. 1). CPa1–CPa4 are active sinters (precipitating at the air-water interface) collected from different margins of the pool. CPa5–CPa16 represent inactive sinters sampled at increasing distances from the spring (Fig. 1). Distance from the pool correlates with sinter age, owing to radial deposition of silica and contraction of the pool over time. CPa16 represents the oldest sinter of those collected, which is located approximately 20 m from the pool edge and estimated to be around 900 years old, the approximate time that Champagne Pool was formed (Lloyd, 1959). Note that samples CPa15 and CPa16 were fragmented and partially buried in the surrounding soil, providing the opportunity to examine the persistence of the geothermal signal into weathered sinter.

Map of Champagne Pool's rim and sinter wall, marking locations of CPa1–CPa16. Inserts show (
2.2. Lipid analysis
Samples were pre-extracted with dichloromethane (DCM)/methanol (MeOH) (1:1 v/v) prior to workup, such that the compounds identified likely derive from microorganisms encased in the silica matrix (Pancost et al., 2005) rather than post-lithification endoliths or, in the case of CPa15 and CPa16, soil bacteria or archaea. Lipids are present in the pre-extracts but are of lower abundance than in the samples, which suggests that background contamination of the latter is minimal. Samples were dried, ground to fine powder, and sequentially extracted by sonication with DCM, DCM/MeOH (1:1 v/v), and MeOH. Activated copper turnings were then added to the lipid extracts and left for 24 h to remove elemental sulfur. An aliquot (50%) of the total lipid extract was fractionated by using aminopropyl solid phase extraction columns (Phenomenex; NH2, 500 mg, 6 mL). The fractions were eluted sequentially with 12 mL DCM/isopropanol (2:1 v/v; neutral fraction, containing, e.g., hydrocarbons, bacterial diethers, archaeol, GDGTs), 12 mL of 5% acetic acid in ether (acid fraction, containing, e.g., free fatty acids, hopanoic acids, and possibly glycolipids), and 24 mL of MeOH (polar and inferred glyco- and phospholipid fraction). Subsequently, 5α-androstane and hexadecan-2-ol (200 ng) were added to the neutral fraction as internal standards. The neutral fraction was then further separated into neutral apolar (containing hydrocarbons) and neutral polar (containing alcohols, bacterial diethers, archaeol, and GDGTs) fractions by elution through an activated alumina column with hexane/DCM (9:1 v/v) and DCM/MeOH (1:2 v/v), respectively.
The glycolipid and phospholipid fatty acid components were released by saponification. Due to the possibility of some glycolipids eluting in the acid fraction, both acid and polar fractions were saponified. Fractions were heated with 1 mL of fresh 0.5 M 95% methanolic NaOH at 70°C for 1 h. The hydrolyzed mixture was left to cool and acidified to pH 1–2 with 1 M HCl ( ∼ 1 mL), then extracted with hexane (3 × 2 mL), combined and evaporated under N2. The fatty acids were methylated by using 100 μL BF3/MeOH solution at 70°C for 1 h. After cooling, 1 mL of double-distilled water was added, and the methyl esters were extracted with DCM as above. The fatty acid methyl esters were dissolved in approximately 1 mL of DCM and eluted through a pre-washed anhydrous Na2SO4 column to remove residual water. An n-C19 standard was added, and the fractions were dried under N2.
2.2.1. Gas chromatography and gas chromatography–mass spectrometry
Before analysis, neutral polar and methyl esterified acid and polar fractions were derivatized with 25 μL pyridine and 25 μL BSTFA (70°C, 1 h) to convert hydroxyl functional groups into trimethylsilyl ethers; the latter two fractions were silylated in order to quench hydroxyl groups in hydroxy fatty acids. Samples were analyzed by a Carlo Erba Instrument HRGC 5300 Megaseries gas chromatograph equipped with a Chrompack CP SIL-5CB capillary column (50 m × 0.32 mm i.d.; 0.12 μm film, dimethylpolysiloxane equivalent) and a flame ionization detector. Hydrogen was used as the carrier gas, and samples were injected at 70°C with a temperature program of 20°C min−1 to 130°C, and 4°C min−1 to 300°C (held for 25 min). Gas chromatography–mass spectrometry analyses were performed by using a Thermo Finnigan Trace gas chromatograph interfaced to a Trace mass spectrometer. The gas chromatograph column and temperature program were the same as those described previously. Electron impact ionization (70 eV) was used, and full scan spectra were obtained by scanning the range m/z 50–800 at 1 scan s−1.
2.2.2 Liquid chromatography–mass spectrometry
Samples were analyzed by high performance liquid chromatography/atmospheric pressure chemical ionization–mass spectrometry based on a procedure modified from Hopmans et al. (2000) and by using an Agilent 1100 series/Hewlett-Packard 1100 MSD series instrument equipped with an auto-injector and Chemstation software. Separation was achieved on a Prevail Cyano column (2.1 i.d. × 150 mm, 3 μm; Alltech), maintained at 30°C. Typical injection volume was 10 μL. GDGTs were eluted isocratically with 99% hexane and 1% isopropanol for 5 min, followed by a linear gradient to 1.6% isopropanol for 40 min. Flow rate was set at 0.2 mL min−1. After each analysis, the column was cleaned by back-flushing hexane/propanol (95:5, v/v) at 0.2 mL min−1 for 10 min. Detection was achieved by using positive-ion atmospheric pressure chemical ionization of the eluent. Conditions for the Agilent 1100 atmospheric pressure chemical ionization–mass spectrometer were as follows: nebulizer pressure 60 psi, vaporizer temperature 400°C, drying gas (N2) flow 6 L min−1 and temperature 200°C, capillary voltage − 3.5 kV, corona current 5 μA. Positive ion spectra were generated by scanning from m/z 900 to 1400.
3. Results
Biomarker concentrations and distributions are highly variable among the sinters from Champagne Pool. Bacterial biomarkers include free fatty acids (Appendix
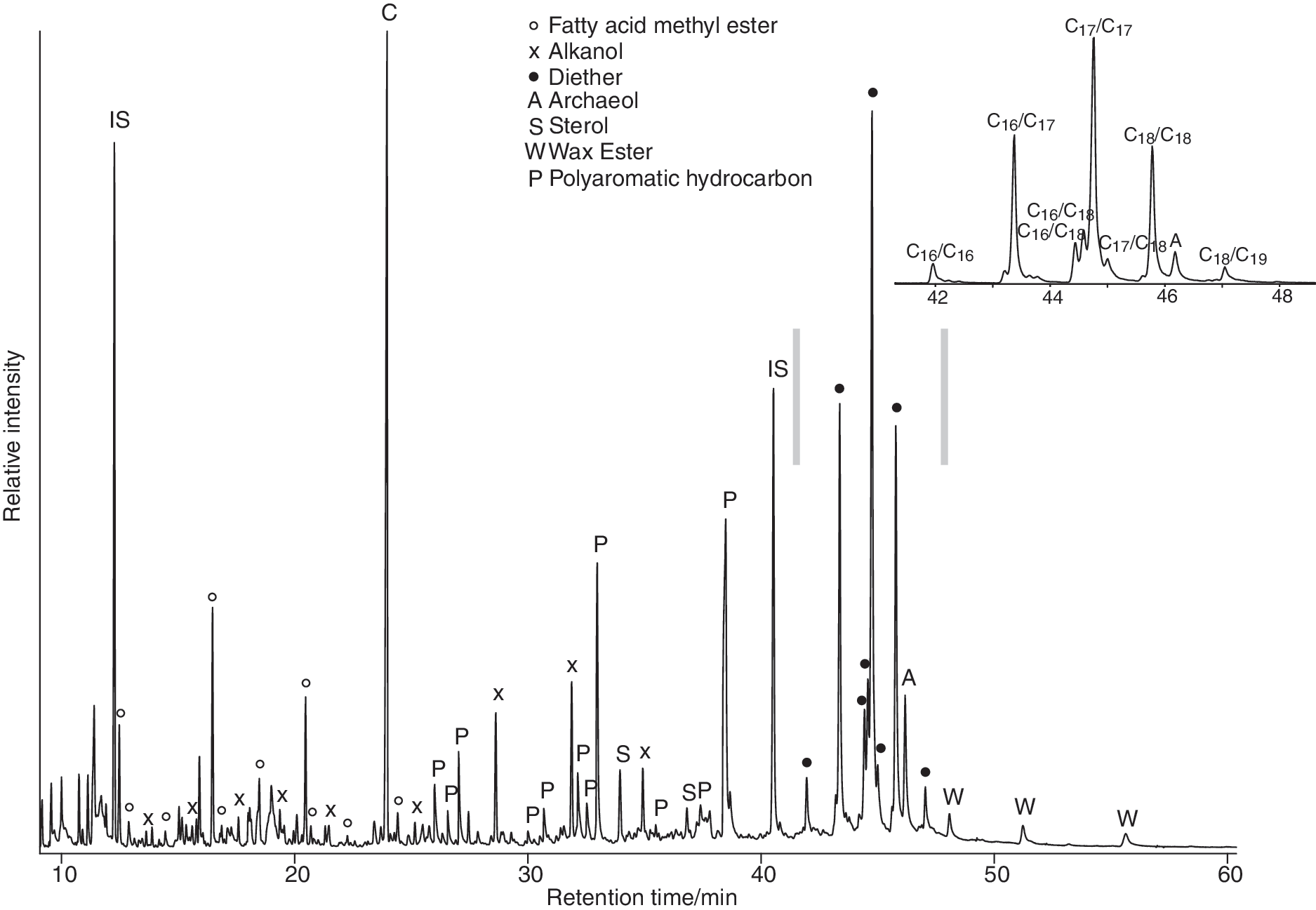
Partial gas chromatogram showing the neutral polar fraction of an inactive sinter (CPa5); inset shows the partial m/z 133 mass chromatogram and distribution of bacterial diethers. Note IS denotes internal standard and C denotes contamination by phthalate.
3.1. Bacterial diether lipids
Dialkyl glycerol diethers (
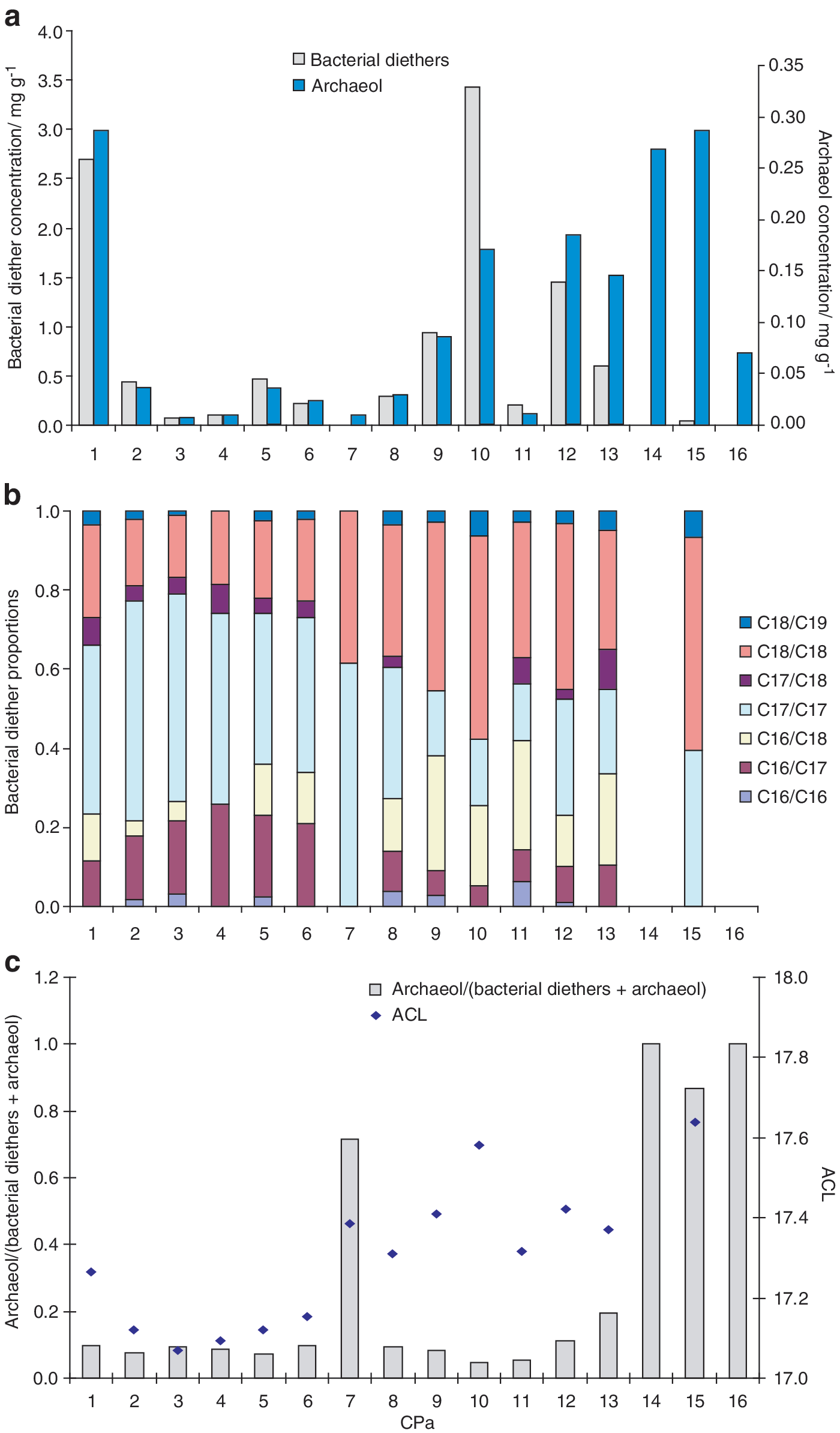
(
Note samples have been renamed since Kaur et al. (2008) to include additional samples and to maintain the chronological ordering.
3.2. Archaeal lipids
Archaeol (
Isoprenoid GDGTs (
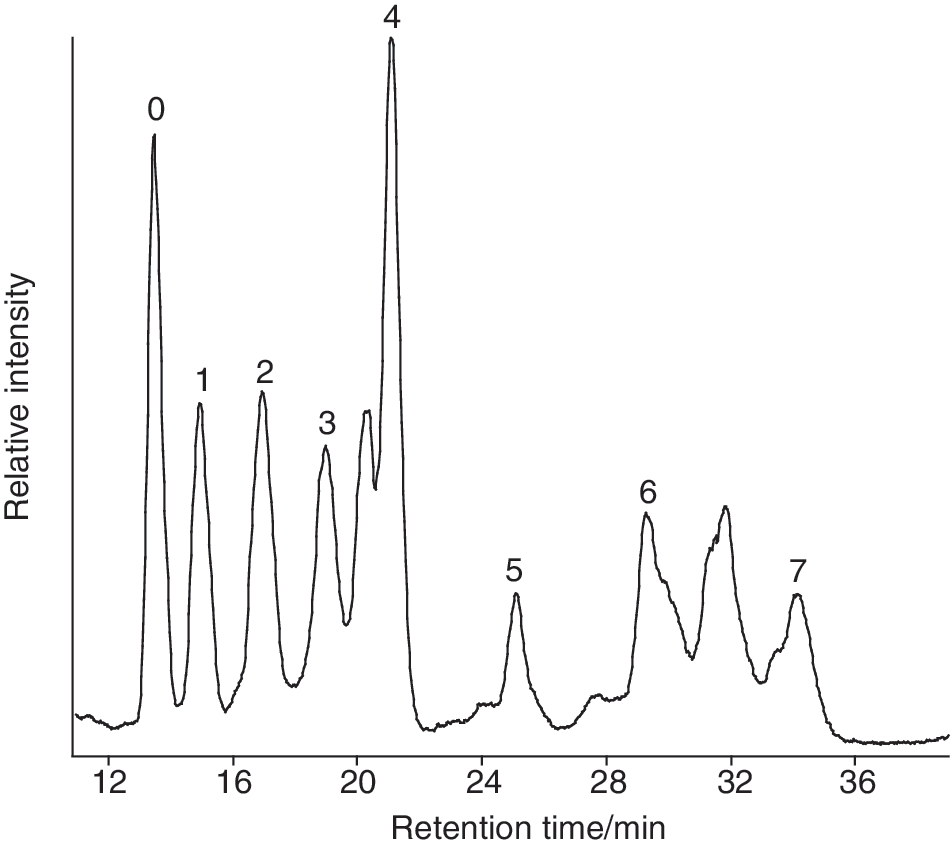
Partial liquid chromatography/atmospheric pressure chemical ionization–mass spectrometry total ion current chromatogram showing the GDGTs identified in the inactive sinter CPa5. Numbers denote the total number of cyclopentyl rings in the two biphytanyl chains.
In addition to GDGTs comprising isoprenoidal biphytanyl chains, non-isoprenoidal branched GDGTs (
3.3. Free and bound fatty acids
The free fatty acid fraction consists of a variety of alkanoic acids (
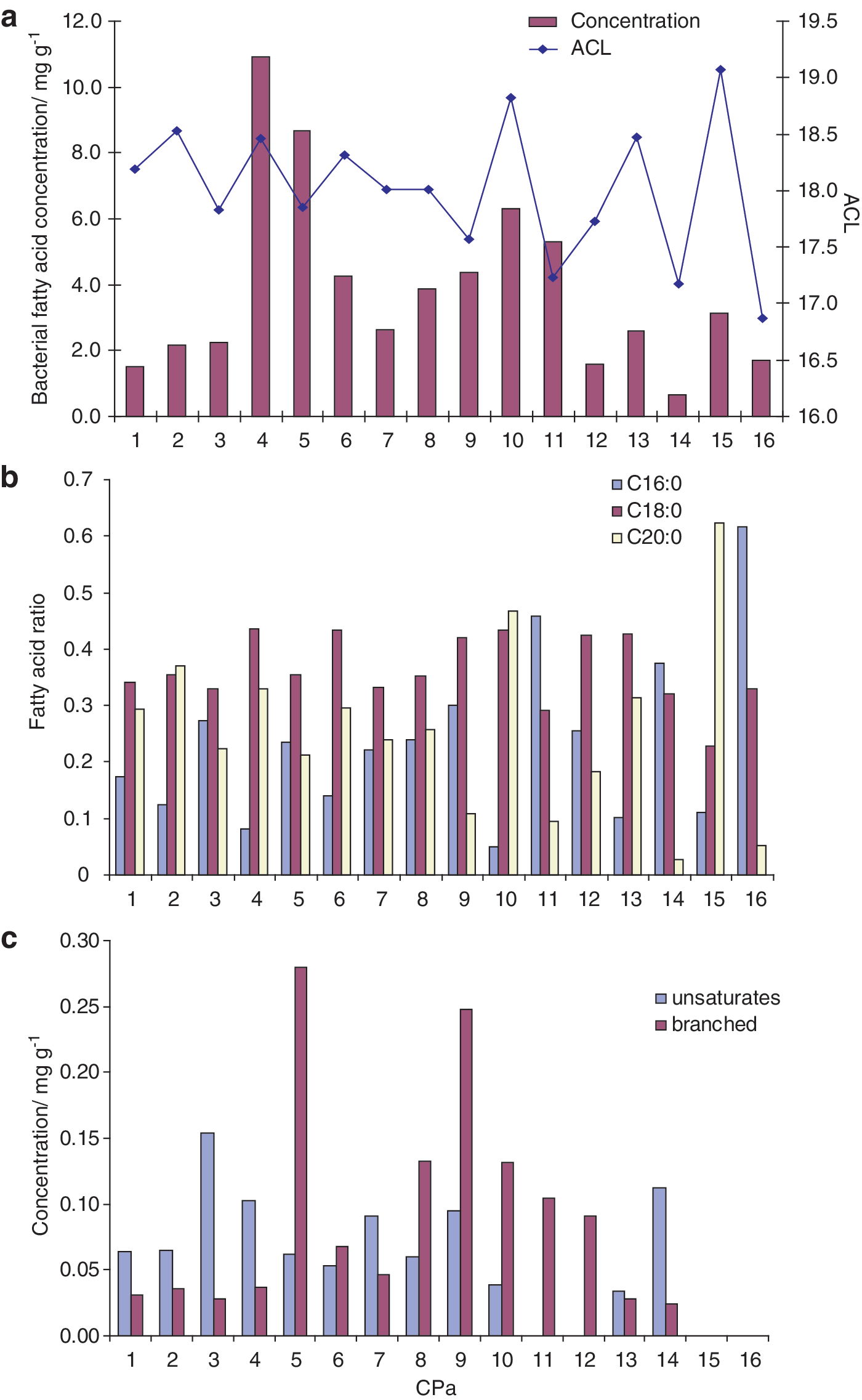
(
Calculated from C14–C20 fatty acids.
In the active and inactive Champagne Pool sinters, β-OH alkanoic acids are widespread and are present in all but one sample, CPa11 (Table 6; Fig. 6). As with the nonhydroxylated fatty acids, a predominance of n-C16:0, n-C18:0, and n-C20:0 components was observed, although total abundances are typically an order of magnitude lower than those observed for the nonhydroxylated counterparts (Table 6). The ACL of these hydroxylated fatty acids shows little variation in the younger sinters, CPa1–CPa10; but in the three oldest, CPa14–CPa16, the value is significantly lower (Fig. 6).

β-OH fatty acid concentrations (mg g−1 TOC) and ACL in Champagne Pool sinters.
3.4. Compounds released by saponification of the putative polar lipid fraction
The fatty acids released upon saponification of the putative polar lipid fraction are inferred to derive from the hydrolysis of 1,2-diacylglycoglycerolipids and 1,2-diacylglycerophospholipids. Caution is required when interpreting this data, particularly absolute abundances, due to the potential loss of phospholipids during analytical workup. Nonetheless, the distribution of the bound fatty acids is similar to that of the free fatty acids, with a general predominance of the n-C16:0, n-C18:0, and n-C20:0 components. Total abundances are variable in the 16 Champagne Pool sinters, ranging from 0.08 to 12.3 mg g−1, but branched and unsaturated components are generally more abundant in the younger sinters CPa1–CPa10. ACLs of the bound fatty acids exhibit little variation, ranging from 17.0 to 18.6.
Previously, archaeol and bacterial diether lipids were detected in the saponified polar fraction of a single Champagne Pool sinter (Pancost et al., 2006). In this study, such compounds were also detected in the saponified polar fraction of some sinters, albeit in lower concentrations. Because the methods employed here are not ideal for quantitative preparation of intact polar lipids, abundances should be interpreted with caution; however, as with our previous work there is evidence that intact archaeal and bacterial polar lipids do persist during sinter formation, and examining their structures is the focus of ongoing work.
3.5. Hopanoic acids
Hopanoids (
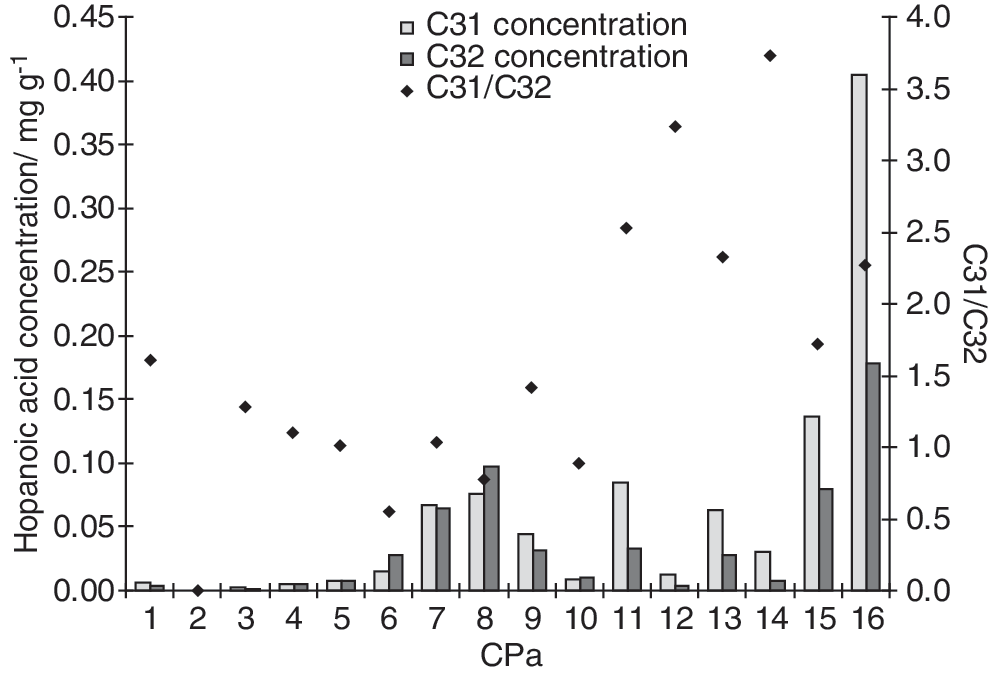
Concentrations of the 17β,21β(H) homohopanoic acid (C31) and 17β,21β(H) bishomohopanoic acid (C32), and C31/C32 ratios in Champagne Pool sinters.
4. Discussion
Central to our evaluation of the fidelity of lipid biomarker signatures is an assessment of both lipid incorporation during sinter precipitation and their subsequent preservation. In our companion paper (Kaur et al., 2011), we evaluated the likely sources of the microbial lipids identified in the Champagne Pool active sinters by comparison with previous molecular analyses of the spring as well as known sources of different lipid classes; that work complements earlier papers that were also focused on profiling lipid signatures in a variety of New Zealand geothermal systems (Pancost et al., 2005, 2006). Here, we focus on the temporal variations in lipid concentrations and distributions and interpret these in terms of preservation as well as spring chemistry and microbiology.
4.1. Sources of lipids
The lipid biomarkers observed in the inactive Champagne Pool sinters are generally similar to those reported in the active ones and likely derive from the same bacterial, archaeal, and in some cases allochthonous sources. In our study of the active spicular sinters (Kaur et al., 2011), bacterial non-isoprenoidal diethers were ascribed to a Thermodesulfobacteriales and Aquificales source, which is consistent with DNA analyses of Champagne Pool sinters and waters (Hetzer et al., 2007; Childs et al., 2008); archaeol and isoprenoid-GDGTs were ascribed to Sulfolobales or Thermofilum-like populations, or both (Hetzer et al., 2007); lower-molecular-weight fatty acids were attributed to a range of potential sources; branched fatty acids were attributed to Thermodesulfobacteriales (Langworthy et al., 1983); and hopanoic acids, degradation products of bacteriohopanoids, were ascribed to an unknown group of bacteria distinct from Aquificales and Thermodesulfobacteriales, as these organisms are not known to synthesize hopanoids. Note that other biomarkers for Aquificales, such as monoethers (Jahnke et al., 2001), were not detected, which perhaps suggests that Thermodesulfobacteriales are the primary source of the bacterial diethers or, alternatively, that the Aquificales species in this setting do not synthesize monoether lipids. The source of the branched GDGTs remains unresolved; possibly they derive from thermophilic bacteria indigenous to the hot spring (Sinninghe Damsté et al., 2007) or an allochthonous source, including bacteria living in surrounding geothermally heated soils (Schouten et al., 2007) or mesophilic anaerobic bacteria (Weijers et al., 2006). Since these assignments are partially based on phylogenetic analyses of active Champagne Pool facies (Hetzer et al., 2007; Childs et al., 2008), caution is required when interpreting the sources of lipid biomarkers in inactive materials. However, many of the main characteristics of the active sinter biomarker distributions—including the presence of C16, C18, and C20 fatty acids; non-isoprenoidal diethers, hopanoids, archaeal diethers and GDGTs; and the absence of monoethers and cyanobacterial or Chloroflexus biomarkers (e.g., Shiea et al., 1991; van der Meer et al., 2002)—are also characteristic of the inactive sinters.
4.1.1. Interpretation of microbial community structure: fidelity of lipid biomarkers
In all four active sinters, the same principal biomarker classes are present, which represent a range of sources largely consistent with recent DNA analyses (Hetzer et al., 2007; Childs et al., 2008). Biomarker concentrations do vary among the active sinters; these are dependent on a wide range of factors but most likely reflect the rates and mechanisms of sinter formation and consequential impacts on the amount of biomass encapsulated. However, biomarker distributions are largely conserved within a given class: bacterial diether distributions, the bacterial diether to archaeol ratio, diether ACL, fatty acid distributions (hydroxylated and nonhydroxylated), fatty acid ACL (hydroxylated and nonhydroxylated), and hopanoic acid distributions are all relatively invariant in the active sinters. Thus, for a given compound class (reflecting a narrow range of organisms), little variation is observed. This suggests that these compound classes derive from similar microbial populations, and the fact that this is recorded by lipid biomarkers suggests that the biomarker composition of precipitated sinters does indeed record microbial populations with some fidelity (Kaur et al., 2011).
4.2. Lipid preservation
Concentrations of lipid biomarkers are typically highly variable; more importantly, concentrations mostly show no correlation with sinter age. In fact, of the compounds investigated, only unsaturated fatty acids and β-OH fatty acids are significantly less abundant in the oldest sinters than in the active and young inactive sinters. For all other compounds, concentration variations between young and old sinters are typically comparable to those that occur among the four active sinters themselves. This suggests that lipids are preserved once the sinter is formed, and biomarker concentrations are largely governed by the size of the microbial population and how well it is preserved during sinter precipitation.
The presence of highly functionalized compounds, for example, putative glycolipids and phospholipids (with distributions similar to their free fatty acid counterparts), in the inactive sinters also suggests that silicification facilitates geochemical preservation. Further evidence comes from the perseverance of lipid distribution patterns. Geothermal lipids are preserved even in highly weathered sinters, which suggests that these signatures can persist for extended periods of time.
4.2.1. Past changes in biomarker distributions
Although the predominant biomarker classes occur in all Champagne Pool sinters, there are clear differences in the lipid distributions from which past changes in microbiology can be inferred. Some of these could reflect a homeoviscous adaptation of the same microbial assemblage to differences in pH or temperature. Environmental conditions can exert a direct influence on lipid biosynthesis, with some organisms able to adjust membrane lipid composition to maintain membrane integrity at extremes of temperature and pH (e.g., Gliozzi et al., 1983; Zeng et al., 1992; Rothschild and Mancinelli, 2001; Schouten et al., 2007). At high temperatures, membrane lipids that are more stable or yield more thermally stable membranes become more abundant (e.g., Zeng et al., 1992; Beney and Gervais, 2001; Schouten et al., 2007) in order to maintain optimal membrane fluidity (i.e., homeoviscous adaptation; Sinensky, 1974; Hazel, 1995) or proton permeability (Albers et al., 2000), or both. Similarly, at low pH levels, acidophiles modify their lipid composition to maintain a high pH gradient across the cell membrane (Albers et al., 2000; Arakawa et al., 2001; Macalady et al., 2004). Alternatively or additionally, differences in lipid distributions can reflect changes in the microbial community structure.
Changes in microbial assemblage are likely to exert the dominant control on changes in the relative amounts of different compound classes. For example, the high abundance of archaeol, particularly relative to the bacterial diethers, in the oldest sinters (Fig. 3) suggests a different microbial assemblage in the past. This is unlikely to reflect differential preservation of microbial lipids, since bacterial and archaeal diether lipids of similar chemical structure and presumably similar preservation potential are being compared. Instead, the distribution trends here appear to reflect different microbial assemblages and, by extension, a different geothermal environment in the past. Since archaea tend to predominate at higher temperatures and lower pH (e.g., Robertson et al., 2005), high archaeol abundances suggest hotter geothermal temperatures or lower pH conditions at Champagne Pool in the past. This is consistent with previous studies that demonstrate a decrease in spring temperature on spring demise (Brock, 1978).
In contrast, changes in hopanoid distributions could either reflect homeoviscous adaptation or differences in microbial community structure. Homohopanoic acid (C31) and bishomohopanoic acid (C32) derive from the oxidative cleavage of vicinal diols in penta- and tetrafunctionalized bacteriohopanoids, respectively (Rohmer et al., 1984; Farrimond et al., 2000), and their distributions vary among different bacterial groups (Talbot and Farrimond, 2007; Talbot et al., 2008). In the Champagne Pool sinters, the abundance of C31 homohopanoic acid relative to C32 bishomohopanoic acid is much higher in the oldest sinters (Fig. 7). This is consistent with intact bacteriohopanoid analyses (Gibson, 2009), which revealed several novel pentafunctionalized bacteriohopanpolyols in the oldest sinters. Although the source of these hopanoids is unclear, this could reflect a change in community composition in the past. Alternatively, several studies have demonstrated changes in hopanoid content and distributions in bacteria under different growth conditions. Total hopanoid content increases with increasing growth temperature in the thermoacidophilic bacterium Alicyclobacillus acidocaldarius (Poralla et al., 1984), the ethanologenic Zymomonas mobilis (Schmidt et al., 1986), and an acetic acid bacterium Frateuria aurantia (Joyeux et al., 2004). Since hopanoids are thought to regulate membrane fluidity and induce order in the phospholipid membrane (Kannenberg and Poralla, 1999), an increase in hopanoid abundances at higher temperatures is not surprising. Joyeux et al. (2004) also reported the biosynthesis of pentafunctionalized hopanoids in response to heat stress in Frateuria aurantia. Here, higher hopanoic acid concentrations and elevated C31/C32 hopanoic acid ratios in the older sinters could therefore reflect either homeoviscous adaptations or a different microbial assemblage in response to higher geothermal temperatures in the past.
Other differences between young and old biomarker distributions could arise solely from homeoviscous adaptation. The proportions of GDGTs comprising multiple cyclopentyl rings are much higher in the oldest samples (Table 4). At high temperatures and low pH, thermophilic archaea tend to biosynthesize more GDGTs relative to diethers and a greater proportion of GDGTs with cyclopentyl moieties (Gliozzi et al., 1983; DeRosa et al., 1986), since these components stabilize the cell membrane, maintaining optimal membrane fluidity and a viable proton motive force (Albers et al., 2000). Thus, the higher proportion of GDGTs bearing cyclopentyl moieties in the oldest samples could reflect an archaeal adaptation to higher temperatures, lower pH levels, or a combination of both.
Similarly, the distributions of fatty acids are also affected by environmental conditions. Thermophilic bacteria respond to high temperatures by increasing their average fatty acid chain length (Weerkamp and Heinen, 1972; Oshima and Miyagawa, 1974; Russell, 1984) and decreasing the degree of unsaturation and degree of branching (Ray et al., 1971; Kaneda, 1991; Zeng et al., 1992). The fatty acid ACLs in the Champagne Pool sinters are higher than those typically observed in mesophilic environments (e.g., Zelles, 1999), which is consistent with a high-temperature environment. Within the sinters, fatty acid distributions are highly variable (Fig. 5). Nonetheless, the lack of branched and unsaturated fatty acids in the oldest sinters is consistent with a higher temperature at Champagne Pool in the past. In contrast, the ACL of the free and bound fatty acids shows no trend with sinter age, with older sinters exhibiting both the highest and lowest ACLs of the entire sample suite. This possibly reflects the complex and multiple sources of these compounds, including a range of microbial but also allochthonous sources; if so, the low ACLs could arise from soil bacterial inputs, and the higher ACLs (due to high abundances of the C20 fatty acid) may reflect elevated past temperatures.
Analogous to the behavior of acyl membrane lipids, it is possible that the average alkyl chain length of bacterial diether lipids will also increase with higher temperatures (Jahnke et al., 2001; Pancost et al., 2005), although this has not yet been directly studied. Here, we observed an increase in bacterial diether ACL with sinter age (Fig. 3c), highlighted by the dominance of the C18/C18 component in the older sinters. If these changes are indeed a result of homeoviscous adaptation, they are further evidence of hotter temperatures at Champagne Pool in the past. Such adaptive responses have not been previously reported and potentially represent new insight into survival strategies in extremes.
5. Conclusions
Biomarker analyses of the active and inactive sinters from Champagne Pool revealed excellent preservation of lipid biomarkers once the sinter has formed. Indeed, variations in lipid concentration between active and inactive sinters are comparable to those that occur among the active sinters themselves. Furthermore, the presence of highly functionalized compounds in the inactive sinters suggests excellent preservation. These findings suggest that silicification facilitates geochemical preservation, and once encased in the mineral matrix, these lipids can persist for extended periods of time, even after significant weathering. Consequently, they can be used to profile past microbiological and environmental conditions. Here, the predominance of generally similar biomarker groups in all sinters suggests a similar microbial community in Champagne Pool throughout its history. However, subtle differences in the lipid distributions between the younger and older sinters provide evidence for different environmental and microbiological conditions in the past. The increase in archaeol concentrations relative to the bacterial diethers in the older sinters suggests a somewhat different past microbial assemblage, perhaps due to higher temperatures or lower pH (Robertson et al., 2005), shortly after the formation of Champagne Pool. Further evidence of higher spring temperatures in the past includes changes in lipid distributions that could reflect homeoviscous adaptations: the high proportion of cyclic GDGTs, the high ACL of the bacterial diethers, and the high concentrations of the hopanoic acids, particularly the C31 component. Clearly, further work is necessary, but this study demonstrates that lipid biomarkers are well preserved in geothermal settings, which reinforces their potential as tracers for past life in such environments.
Footnotes
Acknowledgments
We thank R. Berstan and I. Bull of the Organic Geochemistry Unit and the Bristol Node of the NERC Life Sciences Mass Spectrometry Facility for analytical support, M. Hall for sample preparation, and R. Gibson and H. Talbot of Newcastle University for very useful feedback on this ongoing collaboration. We also acknowledge The Leverhulme Trust for funding assistance, the Geological Society for funding fieldwork, the EPSRC for supporting G. Kaur's Ph.D. studentship, and the Royal Society of New Zealand for providing an ISAT Grant to B.M., facilitating the collaboration.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
ACL, average chain length; DCM, dichloromethane; GDGTs, glycerol dialkyl glycerol tetraethers; MeOH, methanol; TOC, total organic carbon; TVZ, Taupo Volcanic Zone.