
Abstract
Volcano-ice interaction has been a widespread geological process on Earth that continues to occur to the present day. The interaction between volcanic activity and ice can generate substantial quantities of liquid water, together with steep thermal and geochemical gradients typical of hydrothermal systems. Environments available for microbial colonization within glaciovolcanic systems are wide-ranging and include the basaltic lava edifice, subglacial caldera meltwater lakes, glacier caves, and subsurface hydrothermal systems. There is widespread evidence of putative volcano-ice interaction on Mars throughout its history and at a range of latitudes. Therefore, it is possible that life on Mars may have exploited these habitats, much in the same way as has been observed on Earth. The sedimentary and mineralogical deposits resulting from volcano-ice interaction have the potential to preserve evidence of any indigenous microbial populations. These include jökulhlaup (subglacial outflow) sedimentary deposits, hydrothermal mineral deposits, basaltic lava flows, and subglacial lacustrine deposits. Here, we briefly review the evidence for volcano-ice interactions on Mars and discuss the geomicrobiology of volcano-ice habitats on Earth. In addition, we explore the potential for the detection of these environments on Mars and any biosignatures these deposits may contain. Key Words: Volcanism—Mars—Ice—Habitability—Hydrothermal systems. Astrobiology 11, 695–710.
1. Introduction
T
The martian crust is predominantly igneous in nature and ranges from basaltic to andesitic in composition (McSween et al., 2009). Therefore, it is imperative to understand martian volcanic environments in terms of their habitability and potential for microbial colonization. In particular, where volcanism interacts with liquid water, there is the potential to support life, as seen on Earth (e.g., Boston et al., 1992). Liquid water is unstable at the martian surface today and has been for a considerable part of its history. Water currently exists as a largely continuous global cryosphere within, or below, the regolith (Clifford, 1993; Kuzmin, 2005; Clifford et al., 2010), with the largest known reservoirs of water today frozen at the poles (Carr, 1987; Jakosky and Phillips, 2001; Hvidberg, 2005; Clifford et al., 2010) and within a latitude-dependent mantle (Levy et al., 2010a). Differences in localized lithospheric heat flow and crustal thermal properties are likely to result in spatial variation in the cryosphere thickness (Clifford, 2010). This cryosphere, coupled with volcanic activity, has the potential to produce several kinds of environments for life on Mars with a wide range of thermal and chemical conditions, particularly through the generation of hydrothermal systems (Chapman et al., 2000; Head and Wilson, 2002; Schulze-Makuch et al., 2007). It has previously been suggested that where regions of volcano-ice interactions are found, suitable sites may exist to search for evidence of martian life (Boston et al., 1992; Farmer, 1996; Gulick, 1998; Payne and Farmer, 2001; Hovius et al., 2008). Here, we review glaciovolcanism on both Earth and Mars within the context of assessing the range of microbial habitats that exist through volcano-ice interaction, as well as the potential for biosignature preservation within these environments.
2. Glaciovolcanism on Earth
The interaction between volcanism and ice on Earth is ongoing and widespread. Glaciovolcanism specifically describes any interaction between volcanism and ice, including glaciers, snow, firn (recrystallized snow), and ground ice (Smellie, 2006, 2007). Chapman et al. (2000) described three types of volcano-ice interaction: “Type 1” is an alpine interaction with volcano summit snow and valley glaciers; “Type 2” a continental ice sheet/glacier interaction; and “Type 3” involves the interaction with lava and surface ground ice. Type 2 includes subglacial volcanism in its true definition, which is specific to volcanic eruptions beneath thick glaciers and ice sheets (Smellie, 2006), and on Earth subglacial volcanism is a common feature of volcanically active, high-latitude terrains. Examples of widespread subglacial volcanism today include those found in Iceland (Gudmundsson et al., 1997; Bourgeois et al., 1998), British Columbia (Edwards et al., 2002), and Antarctica (Smellie and Skilling, 1994; Smellie et al., 2008). In Iceland in particular, many volcanoes are situated beneath the Vatnajökull ice cap (Chapman et al., 2000), some of which maintain subglacial meltwater lakes (see Section 4.1). Geomorphological products indicative of basaltic subglacial volcanism include tuyas (Fig. 1a) and moberg/hyaloclastite ridges. Tuyas form as a result of central vent eruptions into an overlying thick ice sheet (Bourgeois et al., 1998), while hyaloclastite ridges result from a series of fissure eruptions beneath ice, which form long ridges that follow the strike of the rift. These eruptive features display a distinctive elevated topography in contrast to the surrounding terrain due to the restrictive role of the ice into which the lava was erupted, preventing the lateral flow of lava away from the eruptive center. Subsequent retreat of ice reveals these distinctive volcanic landforms (Fig. 1a, 1b).

Icelandic examples of subglacial volcanic products and environments. (
During an eruption, conductive heat flow melts the surrounding ice, while the low temperatures of the ice begin to solidify the magma under high water pressure, typically forming effusive pillow lava formations (Jakobsson and Gudmundsson, 2008). Convection also plays a large role in the transfer of heat from the magma body to the overlying ice (Höskuldsson and Sparks, 1997), which produces a growing zone of meltwater. Over time, a subglacial edifice can grow within this meltwater “lens’”(Fig. 2a), broadly consisting initially of pillow basalts (Fig. 1b) and then hyaloclastite beds and palagonite tuffs as the confining pressure reduces and the eruption becomes more explosive (Smellie and Skilling, 1994; Jakobsson and Gudmundsson, 2008). If the edifice becomes large enough to break through the ice, a cap rock of horizontal subaerial lava may be deposited (Fig. 2b).
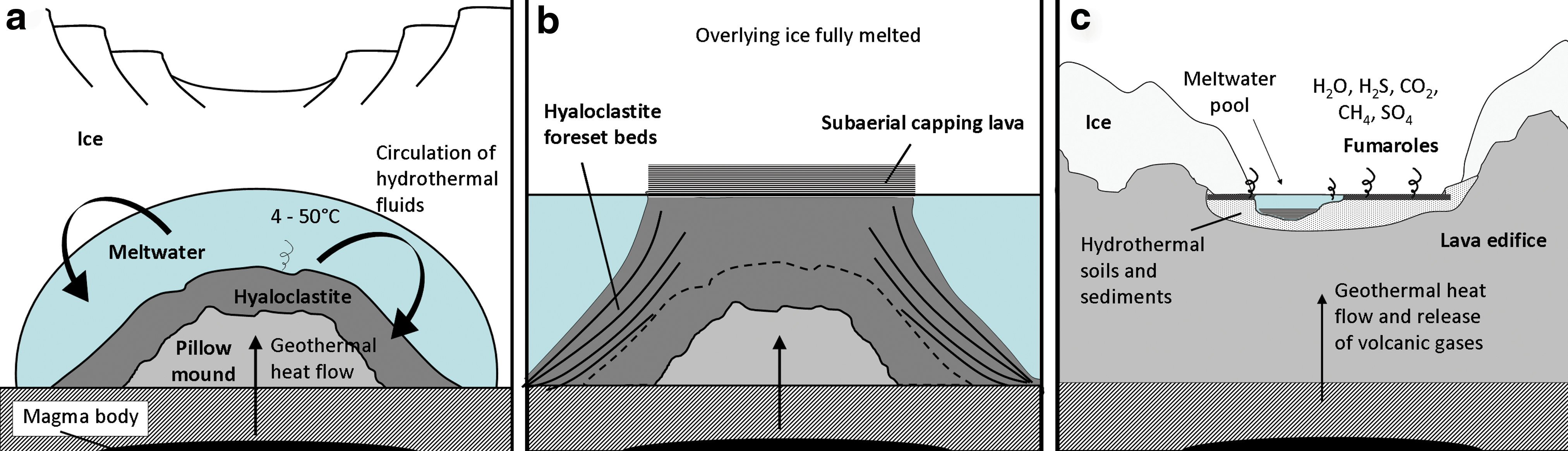
Simplified diagram showing the processes of volcano-ice interaction: (
When the eruption is smaller, perhaps the result of a fissure, entirely subglacial hyaloclastite ridges or pillow mounds (Fig. 1b) will form. These edifices will remain beneath the glacier until exposed and eroded. As magma flow diminishes, the growth of the lava edifice ceases, but the overlying ice continues to melt due to the convective transfer of heat through the liquid water interface between the magma and the ice (Head and Wilson, 2002). Figure 2 summarizes these processes and associated environments. Additionally, subglacial hydrothermal systems may continually melt the base of the glacier, which would sustain a subglacial caldera lake between eruptions (Björnsson, 2002). Such caldera lakes, and meltwater generated during an eruption, are typically catastrophically released as jökulhlaups (Roberts, 2005; Fig. 2c). In the case of Eyjafjallajökull—the Icelandic volcano that erupted in April 2010—the eruption was initially subglacial beneath the small ice cap, but after a few hours this changed to phreatomagmatic activity coupled with meltwater discharge, with the lava eventually emerging from the eruption site ∼1 week after the initial eruption, having melted through the ice (Gudmundsson et al., 2010).
3. Glaciovolcanism on Mars
3.1. Volcanism and the cryosphere
Volcanism on Mars has occurred throughout its history (Carr and Head, 2010); evidence of volcanic activity (e.g., lava flows) spans from the Noachian right up to the very recent Amazonian (Hartmann, 2005; Werner, 2009). Indeed, at specific localities such as Olympus Mons and Hectes Tholes, the ages of lava flows span ∼80% of martian history (Neukum et al., 2004). Evidence of past glaciation is also widespread, both spatially and temporally, with evidence of large polar ice caps in the Hesperian and low-latitude Amazonian glaciations (Kargel and Strom, 1992; Carr and Head, 2010). Likewise, the subsurface cryosphere has been a long-lived and widely distributed source of ice (Clifford et al., 2010). Therefore, it is highly probable that these major processes have interacted in the past (Chapman et al., 2000; Head and Wilson, 2002) and may even continue to do so today deep within the subsurface (Schulze-Makuch et al., 2007). As a result, volcano-ice interaction may represent an environment that has persisted over a significant part of martian history.
3.2. Glaciovolcanism through martian history
The processes and occurrences of volcano-ice interactions on Mars have been reviewed and discussed in depth by Chapman et al. (2000), Head and Wilson (2002, 2007), and Chapman (2003) and involve the emplacement of sills, dykes, lava flows, and large magma bodies into cryospheric permafrost or into an existing ice cap. It has been suggested that glaciovolcanic activity has occurred throughout the history of Mars (Chapman et al., 2000; Head and Wilson, 2007), and there are many topographic features on Mars that have been interpreted as products of volcano-ice interaction (Table 1, Fig. 3). Allen (1979) identified many putative subglacial volcanoes in both the northern plains and near the south polar cap of Mars. Since then, more candidate subglacial volcanoes and regions of volcano-ice interaction have been identified (examples summarized in Table 1 and their locations shown in Fig. 3). These include flat-topped tuyas/edifices (Fig. 5a; Ghatan and Head, 2002; Head and Wilson, 2007), lava ridges/dykes (Fig. 5b; Ghatan et al., 2003; Head and Wilson, 2007), pseudocraters (Fig. 5f; Lanagan et al., 2001; Fagents and Thordarson, 2007), major outflow channels typical of glacial outburst floods caused by geothermally melted ice (jökulhlaups) (Fig. 5c, 5d, Fig. 5b; Head and Wilson, 2002), and marginal drainage channels (Head and Wilson, 2007). Jökulhlaups in particular have been proposed as an explanation for some of the numerous outflow channels and valleys (Figs. 5d and 6b) apparently carved by liquid water (Rice and Edgett, 1997; Baker, 2001; Carr and Head, 2003; Fassett and Head, 2007), with the large flood deposits and catastrophic outwash plains identified on Mars as comparable to those generated by Icelandic jökulhlaups (Fishbaugh and Head, 2002; Hovius et al., 2008). As illustrated by Gulick (1998), much of the fluvial erosion on Mars is spatially and temporally related to volcanic activity. Baker (2001) and Burr et al. (2002a) also observed that catastrophic flood channels and volcanic lava flows are closely associated in the Cerberus Fossae and Marte Vallis region. This further demonstrates the potential importance of volcanism in the generation of liquid water available to life on Mars.
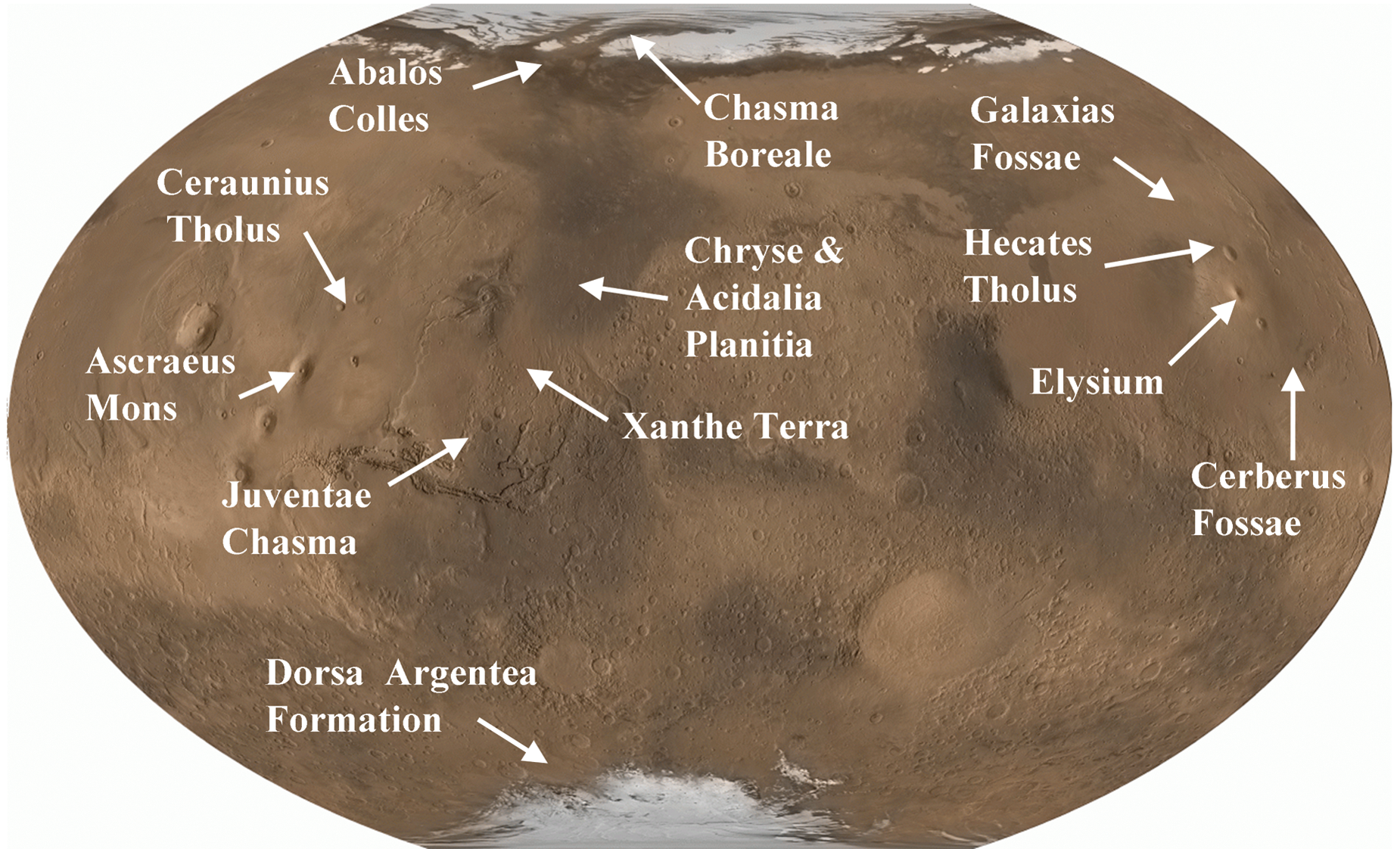
Map of localities given in Table 1 (Image credit: National Geographic Society, MOLA Science Team, MSS, JPL, NASA). Color images available online at
Alternative interpretations of these sites are also shown for comparison.
It has been widely suggested that Noachian Mars (Fig. 4) represented a warmer, and perhaps more clement, period of martian history (e.g., Craddock and Howard, 2002; Chevrier et al., 2007; McKeown et al., 2009) that was followed by a change to acidic, cold, and desiccating surface conditions at the beginning of the Hesperian (Bibring et al., 2006). If true, and if life did indeed evolve in the very early history of Mars, glaciovolcanic environments during the Hesperian and Amazonian may have provided a subsurface refuge as an alternative to the increasingly hostile surface conditions. Here, both Hesperian and Amazonian examples of glaciovolcanism are described (see Table 1 and Figs. 3, 5, and 6 for locations and images).
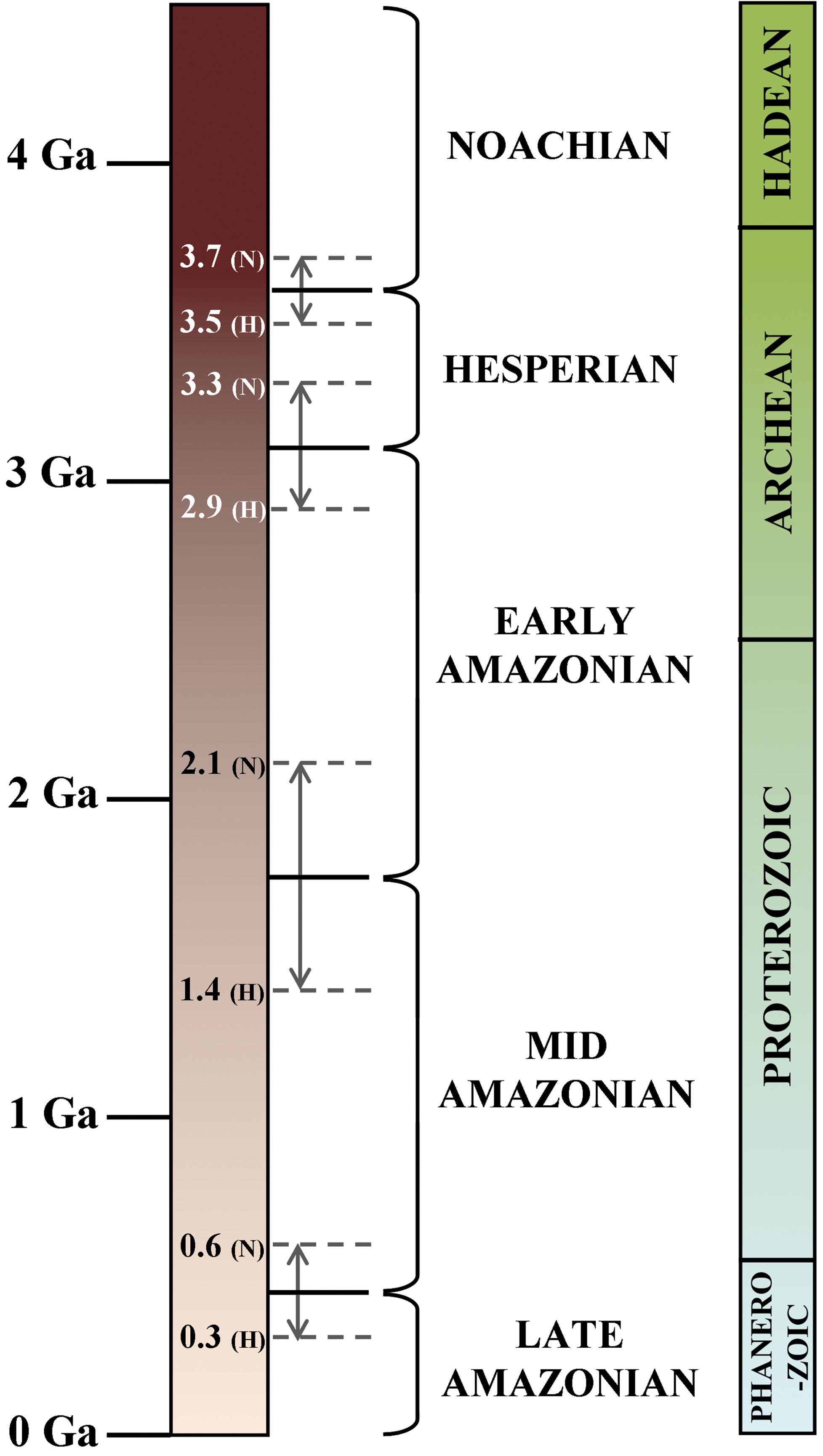
Geological timescales for Mars (left) and the corresponding divisions for Earth (right). Boundary variations for the Neukum (N) or Hartmann (H) chronology models (Hartmann and Neukum, 2001) are shown. Color images available online at

Examples of putative glaciovolcanic features on Mars as described in the text and given in Table 1. (
More examples of putative glaciovolcanic features on Mars as described in the text and given in Table 1. (
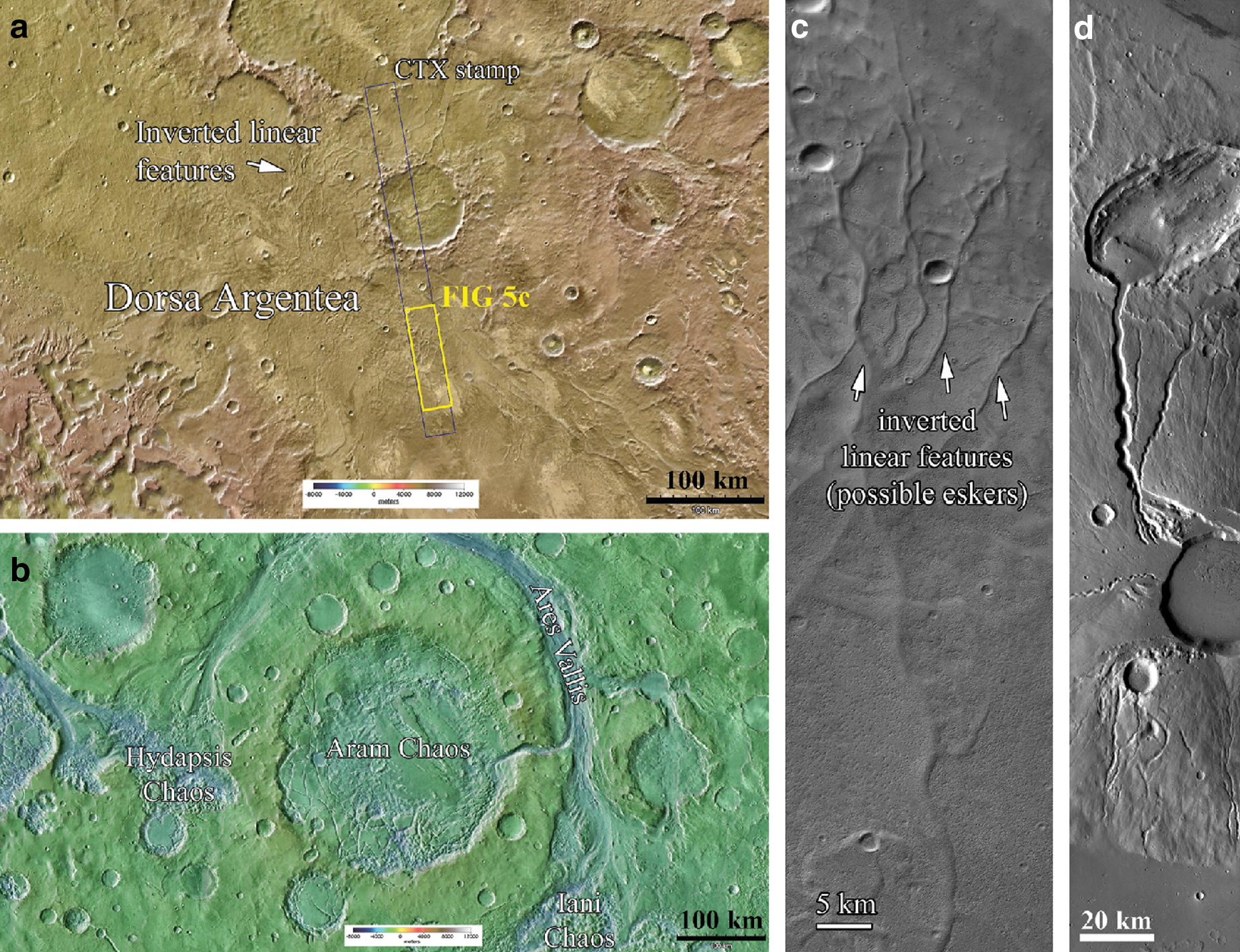
At the Hesperian volcano Ceraunius Tholus (see map in Fig. 3), there is clear evidence for drainage valleys and a depositional fan originating from the caldera rim (Fig. 6d). The geometry of this rim is such that it would favor the accumulation of meltwater sourced from the geothermal melting of snowpack at the summit of the volcano (Fassett and Head, 2007). Likewise, the south polar Dorsa Argentea Formation (Fig. 6a) has been interpreted several times to be an area of multiple subglacial volcanic eruptions with associated meltwater accumulation and drainage (Ghatan and Head, 2002, 2003; Milkovich et al., 2002; Dickson and Head, 2006). This Hesperian-aged, volatile-rich deposit displays evidence for significant melting (e.g., channels, eskers, Fig. 6c), with valleys interpreted to have been outflow regions that drained significant quantities of meltwater from a thinning southern circumpolar ice sheet, induced by volcanic activity (Head and Pratt, 2001; Milkovich et al., 2002; Ghatan and Head, 2004). Finally, interior layered deposits (Fig. 5e) within the Late Hesperian Juventae Chasma have been interpreted by some workers to be the result of sub-ice volcanism (Chapman and Tanaka, 2001; Chapman, 2003). These, and other nearby “light-toned layered deposits” have been found to contain a number of hydrated minerals, including monohydrated sulfates, opal, and ferric sulfates, along with mafic minerals that include pyroxenes and olivine (Bishop et al., 2009). It has been hypothesized that hydrothermal processes may have been involved in the deposition of opal, with sub-ice volcanism providing the necessary heat source (Bishop et al., 2009).
More recently, possible subglacially emplaced dyke swarms (Fig. 5b), and potentially also moberg ridges, have been identified between the Elysium Rise and Utopia Basin (Pedersen et al., 2010), while Levy et al. (2010b) identified features at Galaxias Fossae that bear a striking similarity to volcanogenic glacial cauldrons on Earth (Fig. 5g). The martian cryosphere has of course changed significantly over time, due to a combination of local and global climate change (Baker et al., 1991; Clifford et al., 2010) and effects of obliquity variations (Forget et al., 2006). Evidence for Amazonian glaciation at mid- to low latitudes due to high martian obliquity is now well recognized (Head et al., 2003a; Neukum et al., 2004; Schorghofer, 2007; Fassett et al., 2010). As such, glaciovolcanic products have been identified in the equatorial regions of these terrains (Chapman, 2003; Leask et al., 2006; Kadish et al., 2008) as well as in more polar latitudes. In particular, both glaciation and volcanism are thought to have occurred as recently as the Late Amazonian (Dickson et al., 2008, Mangold et al., 2010), and Head et al. (2003b) identified deposits consistent with a possible martian ice age 2.1–0.4 million years ago. At Olympus Mons, Neukum et al. (2004) found the youngest lava flows to be <30 million years in age and identified multiple episodes of volcanic and glacial activity, with associated hydrothermal water release caused by the melting of ground ice by magma intrusion.
4. Glaciovolcanic Microbial Habitats
The importance of subglacial volcanism for martian exobiology lies in the observation that basaltic subglacial eruptions on Earth generate large volumes of liquid water that can be stored and transported beneath the overlying glacier (Wilson and Head, 2002), and that many of the environments that result from such volcanism exist within the subsurface. In particular, the interaction between geothermal heat flow and an overlying cryosphere or ice cap is highly conducive to the generation of hydrothermal systems (Schulze-Makuch et al., 2007), both during and between eruptions (Björnsson, 2002; Wilson and Head, 2007). Subglacial volcanic habitats range from the overlying cryosphere to deep within the lava edifice, and these are discussed here individually. Examples of the microbiota and physicochemical characteristics of selected environments are also provided in Table 2.
4.1. Subglacial caldera lakes
During and between subglacial eruptions, meltwater can be confined as a subglacial caldera lake (Gudmundsson et al., 1997). Such caldera lakes exist in Iceland (Björnsson, 2002; Johannesson et al., 2007) and have been inferred to have existed on Mars (Fassett and Head, 2007). The lakes in Iceland are inhabited by a specialized population of psychrotolerant and chemotrophic bacteria (Table 2) in the lake water and volcanic sediments that lie at the bottom of the lake (Gaidos et al., 2004, 2008). One of these caldera lakes is characterized by a largely anoxic mixture of glacial meltwater and sulfidic geothermal fluid (Gaidos et al., 2008). A bacterial community based on acetogenesis, sulfate reduction, sulfide oxidation, and potentially methanogenesis is tentatively inferred, with acetogenesis in particular hypothesized to be an important input of carbon into this ecosystem (Gaidos et al., 2008). These caldera lakes can exist as a habitable environment until catastrophically drained as a jökulhlaup and can be highly dependant upon the underlying geometry of the volcanic edifice and overlying ice (Gudmundsson et al., 1997). At Grimsvotn, the topography is such that meltwater can accumulate and form a relatively stable lake until either there is an eruption event or the ice damming the lake is breached (Björnsson, 2002). Conversely at Gjálp, continual drainage of ∼20°C temperature meltwater away from the eruption site has been observed, with no subsequent ponding of water (Gudmundsson et al., 2004; Jakobsson and Gudmundsson, 2008). These subglacial caldera lakes represent one of the most potentially exciting environments within the volcano-ice system.
4.2. Subglacial lava edifices
Basalt, combined with localized areas of hydrothermal activity, has the potential to be colonized by a chemosynthetic-based ecosystem on Mars (Boston et al., 1992; Fisk and Giovannoni, 1999). Mild hydrothermal activity within the volcanic edifice is thought to occur in the several years following an eruption, based on observations of modern subglacial eruptions in Iceland, such as Gjálp (Jakobsson and Gudmundsson, 2008). Basalt is the most abundant geological substrate on Earth and Mars, and as such a significant amount of work has focused on exploring life that inhabits this environment on Earth. Terrestrial basaltic habitats exist predominantly at, and below, the seafloor, within the continental subsurface environments (e.g., aquifers), and as subaerial substrates (e.g., lava flows). Oceanic basaltic lava flows in particular have been the subject of much investigation regarding their microbiota over the past few decades. Fresh basalt erupted from mid–ocean ridge systems is widely found to be colonized and altered by a range of bacterial and archaeal chemosynthetic microbial communities (Santelli et al., 2008). These can exploit the redox gradients between reduced species and oxygenated seawater, such as for Fe oxidation (Edwards et al., 2003), as well as employ anaerobic pathways such as methanogenesis, S0 reduction, sulfate reduction, and Fe reduction (Martin et al., 2008). Additionally, basaltic habitats within the terrestrial deep subsurface have been of interest in terms of understanding subsurface ecosystems on Earth and potentially on other planets, such as Mars (Stevens and McKinley, 1995; McKinley and Stevens, 2000).
Volcanic edifices that currently exist beneath glaciers on Earth are directly analogous to those that may have existed on Mars, but these environments are yet to be explored regarding their microbiota. Those edifices that have been exposed by glacial retreat have been found to host surprisingly diverse bacterial communities. Recent work by Cockell et al. (2009a, 2009b) and Herrera et al. (2009) demonstrated the exploitation of subglacially erupted basaltic hyaloclastites as a favorable volcanic habitat for crypto- and chasmoendolithic life (see Table 2). This widespread utilization of basaltic environments on Earth suggests that any potential biological colonization of subglacial volcanic systems on Mars is likely to exploit the basaltic volcanic edifice as both a physical substrate on which to attach and as a source of energy.
4.3. Cryospheric hydrothermal environments
Glaciers and permafrost on Earth are known to contain a diverse array of psychrophilic and mesophilic life, particularly in basal ice (Table 2) or at the ice-rock boundary (Priscu and Christner, 2004). Such communities could be incorporated into temporary hydrothermal systems within the cryosphere. Martian permafrost also has the potential to provide a habitable environment through the interaction with elevated geothermal heat and the subsequent production of meltwater. This is especially true where magma intrusions have a large surface area/volume ratio, such as dykes and sills (Head and Wilson, 2002). Although no present-day geothermal anomalies have been detected (Christensen et al., 2003), the widespread evidence of significant volcanism and endogenic hydrothermal activity suggests higher heat flow in the past (Schulze-Makuch et al., 2007). McKenzie and Nimmo (1999) calculated that a 16 km wide dyke intrusion into a 5 km thick permafrost layer (ice fraction 0.2 by volume) would produce a subsurface lens of meltwater with a volume of 6.5 km3 for each kilometer length of the dyke, and that such a meltwater zone would not start to refreeze until ∼8 million years after the dyke intrusion. Similarly, Travis et al. (2003) showed that hydrothermal circulation can occur on Mars with sufficient geothermal heat interacting with the overlying permafrost and also suggested that these upwelling hydrothermal plumes could provide a suitable environment for chemosynthetic life (Travis, 2004). Such permafrost hydrothermal systems would remain within the subsurface, except for directly above magma intrusions or where springs breach the surface (e.g., along fractures) (Chapman et al., 2000).
In addition, where a volcanic eruption has taken place beneath a glacier, there is the potential for glacier caves to form within the ice itself, carved by the drainage of hydrothermal fluids and meltwater. Little is known about the processes that occur at the glacier base in volcano-ice settings, including the formation of these glacial caves (Tuffen et al., 2002). Some of the best described caves are those at Mount Rainier, where fumarole interaction with overlying firn and snow produced caves over 1.5 km in length (Kiver and Mumma, 1971; Zimbelman et al., 2000). Some of these caves were observed to be steam-filled through fumarolic activity (Zimbelman et al., 2000), and meltwater was seen to drip continuously from cave walls and ceilings (Kiver and Mumma, 1971). A small crater lake was also observed within part of the cave system (Kiver and Steele, 1972). Glacier caves associated with subglacial volcanism also exist in Iceland (Fig. 1d), and similar “ice towers” have been identified at Mt. Erebus, in Antarctica (Hoffman and Kyle, 2003). These caves provide an ice- and water-rich subsurface environment, potentially coupled with fumarolic input. Such environments would be highly favorable for microbial colonization on Mars, and exploration into the microbiology of those on Earth would shed significant light on this issue.
Finally, high localized geothermal heat flow can also melt the overlying glacial ice or permafrost in isolated areas at the surface and form glacial springs (Fig. 1f) and intraglacial meltwater lakes (Fig. 1e) that interact with surface fumaroles. Such volcanically driven environments exist in the Atacama (Costello et al., 2009), Antarctica (Soo et al., 2009), and Iceland (Ólafsson et al., 2000), and often produce “islands” of biodiversity within an otherwise highly hostile environment (Costello et al., 2009). Martian hydrothermal systems have been suggested many times as an environment suitable for microbial life (e.g., Rathbun and Squyres, 2002; Varnes et al., 2003; Pope et al., 2006), and those generated through volcano-ice interaction are no exception.
5. Biosignature Preservation
The generation of widely varying environments through volcano-ice interaction results in a diverse range of deposits within the geological record. Evidence for putative glaciovolcanism appears to span almost the entirety of martian geological history (Table 1, Fig. 4), although the majority of examples are from Hesperian and Amazonian terrains where geomorphological features are best preserved. Those features that are consistent with the generation and ponding of meltwater are perhaps the most optimum targets, regardless of their age. In particular, deposits representative of environments analogous to the subglacial caldera lakes seen in Iceland (such as jökulhlaup deposits) could be primary targets. These and other products of volcano-ice interaction, including basaltic lavas and hydrothermal mineral deposits, are discussed below regarding their biosignature preservation potential.
5.1. Jökulhlaup deposits
Evidence for life in volcano-ice systems could be recorded via the presence of biomolecules within subglacially erupted basalt and jökulhlaup deposits. Data from the orbiting hyperspectrometers CRISM and OMEGA show the presence of phyllosilicate minerals at the martian surface, with smectite clay minerals such as montmorillonite and nontronite having been identified (e.g., Poulet et al., 2009). It has been proposed that clay-rich deposits may be suitable sites of organic preservation on Mars (Ehlmann et al., 2008). Such minerals are ubiquitous among subglacially erupted basaltic lavas, due to the widespread breakdown of volcanic glass to palagonite and smectite clays through contact with liquid water (Stroncik and Schmincke, 2002). Phyllosilicate detection on Mars has largely been restricted to Noachian terrains such as Nili Fossae (Mustard et al., 2009) and Mawrth Valles (Michalski and Noe Dobrea, 2007). It has yet been found to coincide with putative volcano-ice geomorphological features, although a recent study by Martinez-Alonso et al. (2011) tentatively indicates magnesium-smectite clays to be associated with mesas interpreted to be subglacial tuyas. Volcano-ice landforms therefore could be considered suitable spectroscopic targets for future investigation. If such deposits coincide with volcano-ice interaction terrains on Mars, these could be prime geological formations to search for evidence of life. Indeed, Warner and Farmer (2010) used visible–near infrared and shortwave infrared remote sensing to spectrally identify low-temperature hydrothermal mineralogical assemblages within jökulhlaup deposits in south Iceland. As suggested by these authors, such “mineralogical fingerprints” can be used to identify potentially past habitable conditions within a subglacial volcanic system and are therefore ideal astrobiological targets. An example of such a target includes the drainage valleys and deposits at the edge of the Dorsa Argentea Formation (Fig. 6a). Here, sinuous channels lead away from the bases and margins of candidate subglacial volcanoes (Ghatan and Head, 2002; Head and Wilson, 2007). This terrain is thought to be formed much in the same way as Icelandic jökulhlaup deposits, where drainage channels leading away from the Dorsa Argentea Formation are interpreted to represent volcanism-induced subglacial meltwater release (Ghatan and Head, 2004).
5.2. Hydrothermal deposits
Hydrothermal systems on Earth are noted for their ability to preserve detailed microbial fossils, particularly within silica (Preston et al., 2008) and carbonate (Allen et al., 2000) systems. Indeed, silica deposits of possible fumarolic or hydrothermal origin have been identified by the Mars Exploration Rover Spirit landing site (Squyres et al., 2008). However, such preservation is dependant upon the deposition of mineralized or solute-rich fluids and the subsequent precipitation of the mineral phases and preservation through fossilization of the in situ microbial community. There are examples where concentrated mineral deposits form within, or as a direct result of, volcano-ice interaction. At the Bockfjord volcanic complex in northwest Spitsbergen (Norway), the subglacially erupted volcanoes Sigurdfjell and Sverrefjell contain basaltic lavas with hydrothermal carbonate cement deposits (Blake et al., 2010). These carbonates demonstrate a potential mechanism for the preservation of microfossils and organic biosignatures within a volcano-ice system on Mars. Additionally, subglacially erupted pillow lavas in central Iceland (Fig. 1b) have been found to contain gypsum deposits within the lava vesicles, most likely precipitated during hydrothermal circulation within the subglacial edifice following eruption (Storrie-Lombardi et al., 2009). Such deposits, if found on Mars, would suggest a once-habitable subsurface hydrothermal environment that may have preserved signatures of life.
Alternatively, where subsurface silica-charged hot-spring fluids are frozen through eruption into a subzero environment, cryogenic opal-a is precipitated between ice crystals, which produces distinctive cryogenic particle morphologies (Channing and Butler, 2007). As suggested by Channing and Butler (2007), this precipitation may fossilize any microorganisms present within the hot-spring fluid, which are partitioned out of the growing ice crystals and into the surrounding liquid vein network along with the silica (Mader et al., 2006; Channing and Butler, 2007).
Finally, the subglacial volcano Kverkfjöll in Iceland is associated with several hydrothermal systems (Cousins, 2010). One of these—the hot spring Hveragil—has thick deposits of calcite along the floor of the gully that the hot spring flows along (Ólafsson et al., 2000) and, as with many hot-spring mineral deposits, is likely to contain biosignatures such as microfossils, organics, or both. Little is known regarding the preservation of biosignatures within such systems generated by volcano-ice interaction, and this represents a significant area of research with direct implications for the search for life on Mars. One significant drawback, however, is the often small-scale and highly localized nature of such mineral deposits (e.g., to an isolated spring), which could potentially hinder their discovery.
5.3. Subglacially erupted lavas
Glassy basaltic lavas on Earth often contain intricate tubular and pitted structures, which have been widely interpreted to be formed by the activities of euendolithic microbes at the glass-palagonite interface (Furnes et al., 2007, and references therein; McLoughlin et al., 2009). Typically, 80–90% of hyaloclastite is glass (Jakobsson and Gudmundsson, 2008), which leads to the possibility for the significant production of microbial bioalteration textures so commonly seen in submarine glassy lavas (McLoughlin et al., 2009). It has been previously suggested that these bioalteration textures would make suitable biosignatures when looking for life on Mars (Banerjee et al., 2006; McLoughlin et al., 2007), particularly when they are preserved by minerals such as zeolites and titanite infilling tubular textures (Furnes et al., 2004; Izawa et al., 2010). However, while these putative biosignatures appear to be ubiquitous in lavas within an oceanic setting, an abundance of such textures is yet to be found in basalt of a subglacial origin, despite sharing the same glassy lithologies (pillow lavas and hyaloclastites). A recent study by Cousins et al. (2009) demonstrated a possible environmental control on the formation of bioalteration textures and in particular showed that the subglacial environment was not as conducive to their formation as environments that are oceanic. Likewise, while Cockell et al. (2009a) described biogenic pitting in subglacially erupted hyaloclastites in Iceland, they also noted an absence of the characteristic tubular textures seen in oceanic lavas. Bioalteration textures therefore may not necessarily be the most suitable biosignature for identifying past life within subglacial basaltic lavas, and other alternative options, such as geochemical biosignatures, should also be explored. For example, distinctive trace element (Zr, Sc, and Mn) signatures have been found to result from the utilization of organic acids to dissolve basaltic substrates (Hausrath et al., 2007, 2009). Likewise, sulfur isotope (32S, 33S, 34S) compositions can provide evidence of microbial sulfate reduction within altered oceanic basalts (Rouxel et al., 2008).
6. Discussion and Conclusions
An active volcano-ice system can potentially provide all the necessary ingredients for life. The continual release of geothermal heat into an overlying glacier can sustain a subsurface meltwater environment, while the release of volcanic gases such as H2S, CO, CO2, and H2 could support a variety of chemosynthetic metabolisms. The presence of this heat flow will also mean that a continual convective system will create a cycling of material through the different environments, which will remove waste products from some niches and deliver nutrients to others.
It is clear that the presence of liquid meltwater is key to glaciovolcanic systems being suitable for life, but there are significant differences between terrestrial and martian systems that need consideration. On Mars, the melting efficiency of water ice is much reduced due to the low initial temperature of the ice (Hovius et al., 2008), which perhaps suggests volcano-ice systems on Mars were not as viable as those on Earth. Indeed, there are locations on Mars interpreted to be the result of subglacial volcanism that display a distinct lack of evidence for meltwater. Such places include the proposed subglacial lava flows at Ascraeus Mons, where rapid refreezing of a cold-based glacier would prevent any significant basal melting (Kadish et al., 2008). However, it is thought that the temperature of the meltwater is highly influential on the formation of jökulhlaups, whereby higher heat flow enables the enlargement of subglacial drainage tunnels (Gudmundsson et al., 1997). The occurrence of jökulhlaup-like flows and deposits on Mars therefore suggests that subglacial eruptions can lead to significant subglacial melting, even with cold-based glaciers (Head and Wilson, 2007).
While habitable environments potentially may exist in this subglacial volcanic setting on Mars, they are most likely to be transient and isolated. On Earth, any new body of liquid water will be rapidly colonized due to the widespread and globally connected biosphere (Cockell and Lim, 2005). While it remains possible that pockets of martian life could exist, as yet there is no evidence for a martian biosphere. As a result, the delivery of martian life, should it exist, to newly formed habitable environments remains a problem. It can be seen that, in Iceland, features indicative of subglacial volcanic activity often occur in clusters (e.g., Alfaro et al., 2007), which suggests that localized habitable regions may exist within a close enough proximity to allow transport of microorganisms between individual niches. On Earth, regions of high heat flow are rarely isolated to just one volcano. Indeed, in the case of Iceland, Vatnajökull (glacier) overlies seven individual volcanic centers. Additionally, it has been observed that rapid vertical transport of hydrothermal fluid occurs beneath Mýrdalsjökull via faults within the ice (Björnsson, 2002). Therefore, it is possible to envisage such subglacial systems to be connected via fractures and channels within the ice, where meltwater (and any microbial life it may carry) may circulate, distributing microorganisms from one system to another.
The vast majority of the terrestrial biosphere is dependent upon photosynthesis, either directly or indirectly (Varnes et al., 2003). Photosynthesis on Mars, however, would be hindered by the exposure to UV radiation and by the increased distance to the Sun, which reduces the flux of photosynthetically active radiation (PAR) to ∼55% of that typically experienced on Earth (Cockell and Raven, 2004). If photosynthetic communities were to exist within a subglacial volcanic system, they would be limited to the near-surface ice and specifically use blue and green wavelengths due to the high absorbance of red light within ice (Hawes and Schwarz, 2000). Cockell and Raven (2004) showed experimentally that the maximum depth within snowpack at which the minimum level of PAR can penetrate is ∼24 cm. Additionally, work on ice-covered lakes in Antarctica has shown there to be benthic photosynthetic communities residing at ∼16 m water depth beneath 3.5–5 m of ice cover (Vopel and Hawes, 2006), which is much shallower than the depths of many subglacial volcanic systems, which are typically beneath several hundred meters of overlying ice (Wilson and Head, 2002). At depths of 100 m within glacial ice, PAR is entirely absent (Warren et al., 2002). Subglacial volcanic environments, therefore, are not suited to a photosynthesis-based community. This limits the primary producers of this environmental setting to chemosynthetic pathways.
On Earth, chemoautotrophs are major contributors for communities residing within dark, extreme environments, such as deep sea vents (McCollom and Shock, 1997; Van Dover, 2000). Specifically to Mars, anaerobic chemolithoautotrophs can potentially inhabit subglacial volcanic environments through the oxidation of inorganic compounds and fixation of carbon dioxide as the carbon source (Boston et al., 1992). Numerous chemosynthetic pathways could be exploited due to the chemical disequilibrium that results from the mixing of high- and low-temperature fluids (Gaidos and Marion, 2003). On Earth, the majority of the chemosynthetic microbial communities residing in present-day hydrothermal systems are indirectly dependent on photosynthetically produced O2 (Varnes et al., 2003). However, an estimated 1–2% of these communities obtain chemical energy from redox reactions that are completely independent of photosynthesis (Varnes et al., 2003), and it is these microorganisms and their metabolic pathways that are potentially suitable for survivability on Mars, particularly within subglacial hydrothermal systems.
In conclusion, the conditions that exist as a result of volcano-ice interaction provide a wide range of habitats for life on Earth and may have provided a possible subsurface haven for life on Mars during past epochs. While there is still much work to be done with regard to understanding the thermal and geochemical conditions of such environments on Mars, the combination of basaltic lava, liquid water, and hydrothermal activity provides a possible subsurface haven for life. The wide range of geological deposits—be they jökulhlaup sediments, hydrothermal minerals, or subglacial basalt—provides several mechanisms for the preservation of any biosignatures for future discovery.
Footnotes
Acknowledgments
This work is funded by the Leverhulme Trust. Field observations made in Iceland in July 2007 were additionally supported by a Birkbeck College Faculty Research Grant; we thank Dr. Katie Joy and Dr. Oliver White for field assistance whilst in Iceland. Finally we especially thank Dr. Nick Warner for assistance with Mars images and detailed comments and two other anonymous reviewers for their suggested revisions that significantly improved the manuscript.
Abbreviation
PAR, photosynthetically active radiation.