
Abstract
The thermal springs Trollosen and Fisosen, located on the High Arctic archipelago Svalbard, discharge saline groundwaters rich in hydrogen sulfide and ammonium through a thick layer of permafrost. Large amounts of biomass that consist of filamentous microorganisms containing sulfur granules, as analyzed with energy dispersive X-ray analysis, were found in the outflow. Prokaryotic 16S rRNA gene libraries and quantitative polymerase chain reaction (qPCR) analyses reported bacteria of the γ- and ɛ-proteobacterial classes as the dominant organisms in the filaments and the planktonic fractions, closely related to known chemolithoautotrophic sulfur oxidizers (Thiotrix and Sulfurovum). Archaea comprised ∼1% of the microbial community, with the majority of sequences affiliated with the Thaumarchaeota. Archaeal and bacterial genes coding for a subunit of the enzyme ammonia monooxygenase (amoA) were detected, as well as 16S rRNA genes of Nitrospira, all of which is indicative of potential complete nitrification in both springs. 16S rRNA sequences related to methanogens and methanotrophs were detected as well. This study provides evidence that the microbial communities in Trollosen and Fisosen are sustained by chemolithotrophy, mainly through the oxidation of reduced sulfur compounds, and that ammonium and methane might be minor, additional sources of energy and carbon. Key Words: Chemolithotrophic microorganisms—Thermal springs—Svalbard—Arctic—Thaumarchaeota—Cave system. Astrobiology 11, 665–678.
1. Introduction
T
Svalbard has always been directly affected and influenced by the many tectonic phenomena that have taken place in the Norwegian Greenland Sea. For western Svalbard, the early continental fragmentations in the floor of the Norwegian-Greenland Sea led to a tectonic uplift in the late Cenozoic about 25 million years ago (Vågnes and Amundsen, 1993). The uplift (>1 km) led to the creation of several long north-south oriented fault zones close to the continental margin, which marks the transition between the thin oceanic crust and the thick continental crust of Svalbard (Martinsen and Nøttvedt, 2008; Bergh and Grogan, 2010). It is in this uplifted area, in parallel to the fault zones, that the Arctic springs are located; from the Troll, Jotun, and Gygre thermal springs in the Breibogen-Bockfjorden fault system in the northwest (Banks et al., 1998; Salvigsen and Hogvard, 1998) to the Fisosen and Trollosen thermal springs in the Sørkapp-Hornsund fault system in the southwest (Fig. 1) (Lauritzen and Botrell, 1994). It is along the northernmost part of this uplifted area that three extinct quaternary volcanoes are situated, in addition to areas with tertiary basaltic lava that indicate that this system has been active for a substantial amount of time (Vågnes and Amundsen, 1993). Though the details on the origin of the geothermal anomaly in western Svalbard are unclear, it has been suggested that it extends from the Yermak hot spot north of Svalbard (Fig. 1) (Feden et al., 1979; Vågnes and Amundsen, 1993). The elevated geothermal gradient is thought to be a regional phenomenon that broadly coincides with the uplifted area of western Svalbard, which thereby affects both the northwestern and southwestern fault systems mentioned (Vågnes and Amundsen, 1993).
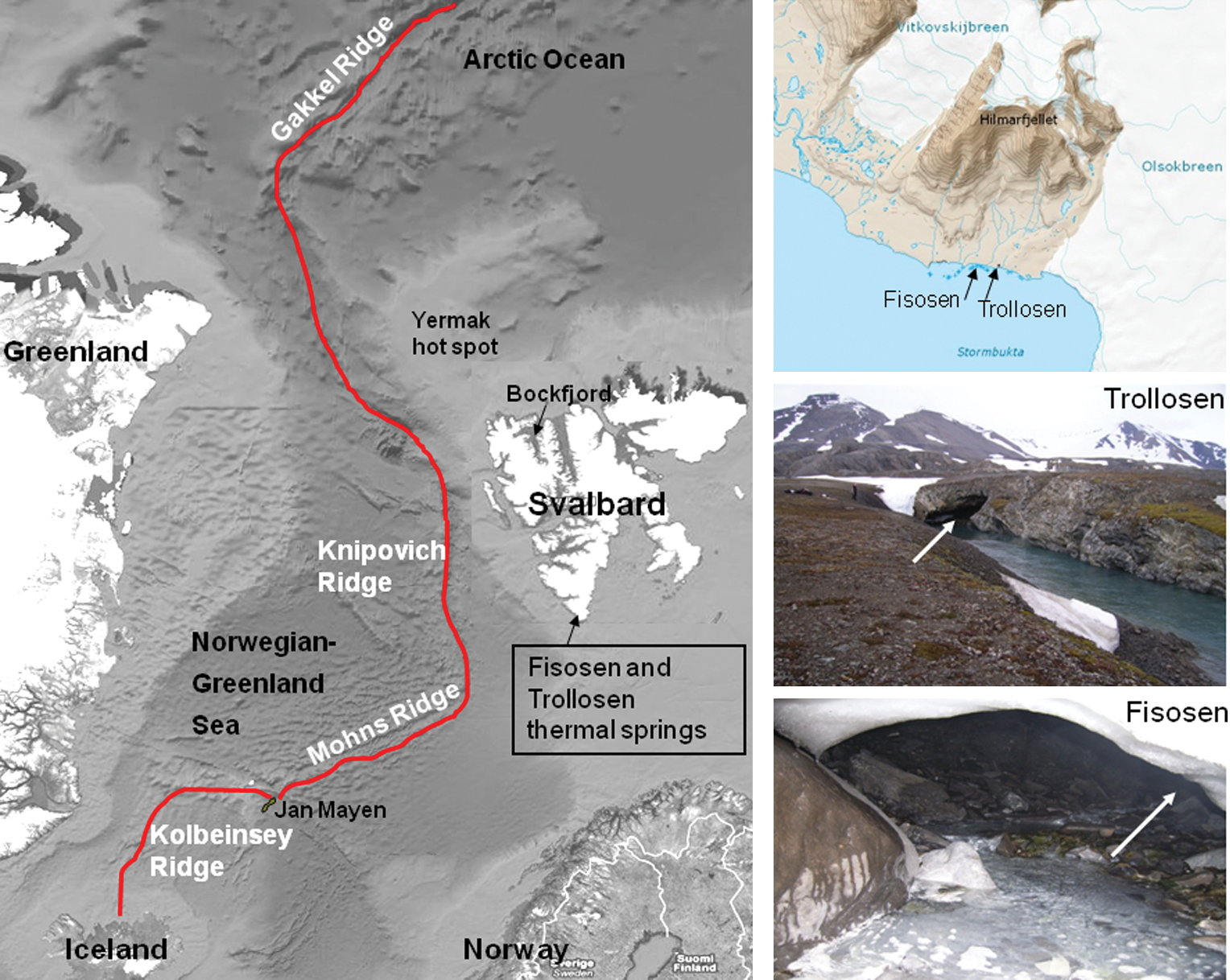
Map of the Norwegian-Greenland Sea highlighting the location of Svalbard (76–78°N) with the Fisosen and Trollosen thermal springs at Sørkapp. The North-Atlantic Spreading Ridge is marked with a line. Arrows in the site photos indicate location of spring openings. The basic satellite photo was obtained from Google Maps. Color images available online at
The presence of liquid water is considered a major prerequisite for the possible development and maintenance of life on Mars (Beaty et al., 2005, and references therein; Hoehler and Westall, 2010). Water at, or near, the martian surface is locked in permafrost, and calculations have indicated that water, if present, could be in liquid state at kilometers of depth under the permafrost layer (Clifford and Parker, 2001). There is also evidence for geologically recent groundwater seepage and surface runoff (Malin and Edgett, 2000; Malin et al., 2006). Since terrestrial permafrost environments like those in Svalbard might provide similar conditions, they could be valuable analogues of martian environments.
A geological study showed that the two sulfidic springs Fisosen and Trollosen in the Sørkapp-Hornsund fault system discharge from a hot subpermafrost karst aquifer (Fig. 2) (Lauritzen and Botrell, 1994). Although located only a few meters from the seashore, both springs have significantly different water chemistry than that of the neighboring seawater. When compared to the seawater, both springs have a large excess of Cl−, and Ca2+ over Mg2+, in addition to a significant depletion in sulfate. The high salinity was suggested to originate from a deep thermal brine component (Lauritzen and Botrell, 1994). Modeling of the heat transport under the springs has indicated that, in order to explain the ablation rate and the thermal flux of the springs, the temperature of the deepest parts of the aquifer must be 600–800°C (Jensen, 1998; Lauritzen, unpublished data). After passing through several hundred meters of permafrost, the resulting surface fluids hold temperatures of 15°C and 6°C for Fisosen and Trollosen, respectively. To date, the microbiota of the springs in Svalbard has not been characterized. The cold perennial sulfidic springs on Axel Heiberg Island in the Canadian High Arctic are among the few examples of Arctic springs where the microbial composition has been studied (Perreault et al., 2007, 2008). These springs were populated by a community dominated by sulfur-oxidizing and sulfur-reducing bacteria, similar to communities reported from other sulfidic environments, like deep-sea hydrothermal vents (Campbell et al., 2006), marine sediments (Teske and Nelson, 2006), terrestrial caves (Engel et al., 2001, 2003; Chen et al., 2009), and sulfidic springs (Moissl et al., 2002; Koch et al., 2006). Chemoautotrophic microorganisms that gain energy from the oxidation of chemicals such as reduced inorganic sulfur compounds or methane are the primary producers in these ecosystems.
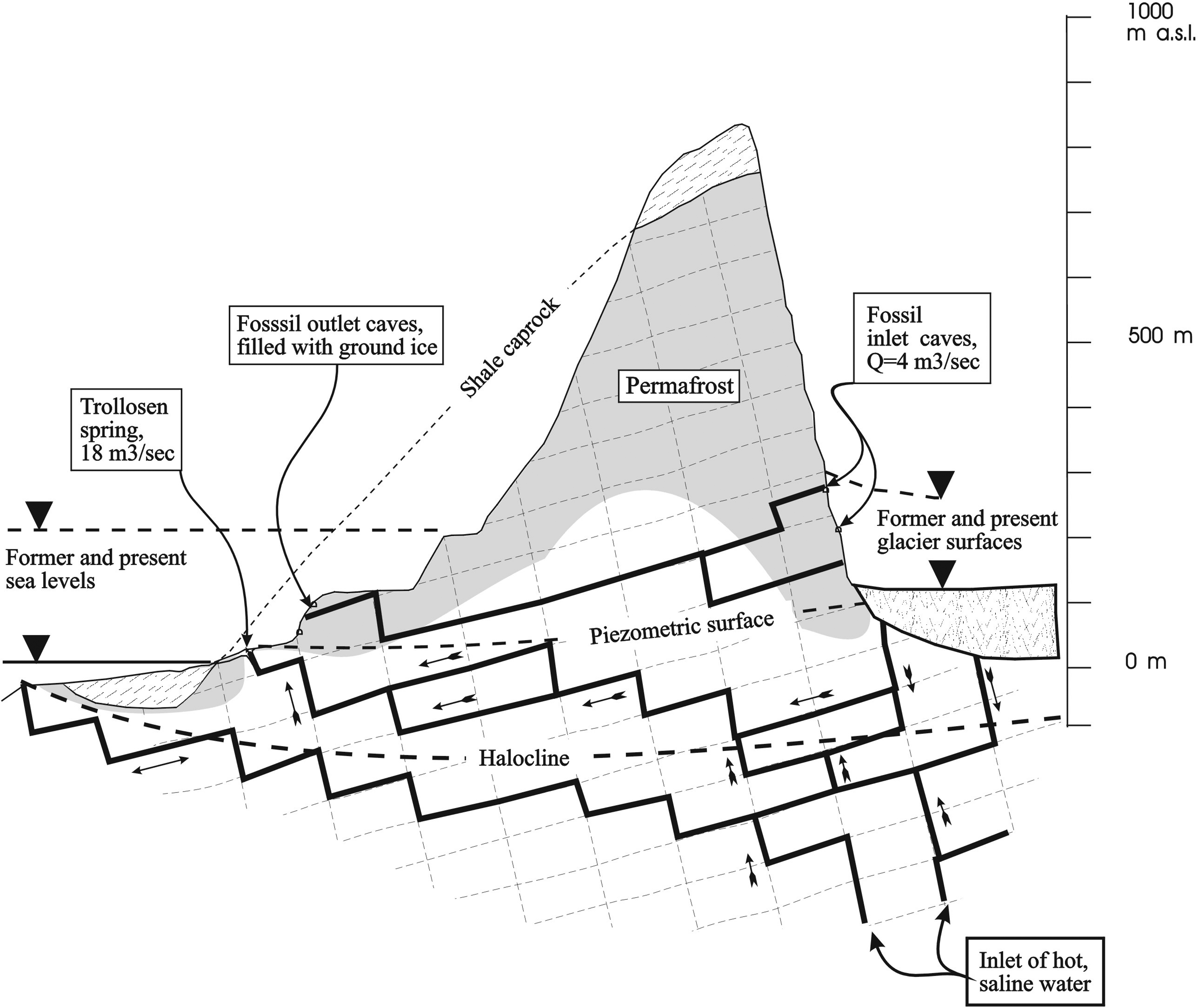
Cross section through Hilmarfjellet and the thermal Trollosen spring at Sørkapp. The spring conveys two components of water: meltwater mainly from the Vitkovskij glacier in the upland and deep hot brine. The two components are mixed in the cave conduits within the aquifer. Enthalpy calculations suggest that the rock/water interface in the deepest parts of the aquifer must approach 600–800°C (Jensen, 1998). From Lauritzen 1996.
We report here the first comprehensive microbiological characterization of the communities in the thermal springs Fisosen and Trollosen at Svalbard, using molecular microbial approaches that include 16S ribosomal RNA fingerprinting, qPCR, functional gene screens, microscopic techniques, and geochemical analyses.
2. Materials and Methods
2.1. Site description and sample collection
The Trollosen and Fisosen thermal springs close to the base of Hilmarfjellet at Sørkapp in the South-Spitsbergen National Park of Svalbard were sampled for microbiological and geochemical analyses (Figs. 1 and 2; Table 1). The Trollosen spring (76°42′N, 16°14′E) has a cave opening of 3×1 m and is dominated by glacial meltwater during the summer from the nearby Vitkovskij glacier, which makes the water highly turbid with a discharge of 18–20 m3/s in midsummer (Lauritzen and Botrell, 1994) and a winter base flow of 2–3 m3/s (Lauritzen, unpublished data). Therefore, the sampling time was set to early June (2006) just before the main melting season from the Vitkovskij glacier when the springwater was still clear with a low discharge of approximately 8 m3/s (Fig. 1). The underlying cave of Trollosen is always completely water-filled and inaccessible, so all material, both for chemical and biological analysis, was sampled from the spring pond. The white filamentous material was sampled by an ethanol/sterile water-cleaned fine-mesh net and either homogenized on site and directly fixated in RNALater (see below) or kept untreated on ice for 20 h until it was transferred to laboratory facilities in Longyearbyen. Part of the material was treated for inductively coupled plasma mass spectrometry and microscopy analyses (see below). Spring fluid was collected directly into plastic containers treated in advance with ethanol and sterile water. The Fisosen spring (76°42′N, 16°13′E) with an opening of about 20 cm in diameter discharged about 1 L/s of clear fluid and is apparently independent of the glacier meltwater inflow. At sampling time, the Fisosen spring was partly covered with snow. In contrast to Trollosen, the outflow of the Fisosen spring was covered with white filaments attached to the rocks. The attached filaments were harvested directly with sterile syringes. The filaments and fluid samples were treated as at Trollosen.
H2S detection limit is 10 μM.
bd, below detection limit; nd, not determined.
2.2. Filtration of spring fluids
Spring fluid from Trollosen and from Fisosen was filtered on arrival at the laboratory less than 20 h after sampling. Ten liters were first filtered through a 10 μm (Nalgene) filter to remove the large filamentous material, then through a 0.2 μm (Nalgene) filter to capture the microbial cells. The filters were either cut into smaller pieces and frozen for direct transfer to bead beating tubes, or the cells were scraped off and immediately immobilized into agarose plugs for subsequent cell lysis (see DNA extractions below).
2.3. Chemical analysis of spring fluids
A few hours after sampling, liquid from Trollosen and from Fisosen was centrifuged (10,000g for 10 min) to remove cells, and HNO3 was added to the supernatant (2% final concentration). The geochemical composition of liquids was analyzed with a Finnigan Element2 inductively coupled plasma mass spectrometer (ICP-MS) and a Metrohm ion chromatograph at the Centre for Element and Isotope Analysis at the University of Bergen. The hydrogen sulfide concentration of the springwater was determined with the methylene blue method described by King and Morris (1967). Ammonium concentrations were determined according to Kandeler and Gerber (1988).
2.4. DNA extractions
DNA was extracted from the free-floating macroscopic filamentous material in Trollosen, the attached filaments in Fisosen, and from the fluid (10 L) of both springs. For the DNA isolation from the filaments of Fisosen and Trollosen, two different lysis and DNA extraction procedures were carried out. First, the filaments were homogenized and the cell suspension immobilized into agarose plugs for in-gel lysis by lysozyme and proteinase K [for details see Reigstad et al. (2011)], followed by DNA isolation from the plug with Gelase (Epicentre Biotechnologies). This DNA is later referred to as plugDNA. In the second approach, the filamentous cells were lysed by a standard bead beating approach with the FastPrep DNA Extraction Kit for soil (Q-BIOgene) and the manufacturer's recommendations. In this bead beating approach, the analyses were performed by transferring 0.5 g filament to the bead vial directly from the sample with sterile forceps. The DNA extracted by this approach is denoted bbDNA (bead beating DNA).
2.5. RNA extraction and cDNA synthesis
Filamentous material (5 g wet weight) from both springs was homogenized in sterile mortars immediately after sampling and transferred to sterile tubes containing 25 mL RNALater solution (Invitrogen). This RNALater solution was later removed from the sample by centrifugation (5 min at 10,000g, 4°C). The remnants of the RNALater solution were washed out with 2 mL 1× TE buffer pH 8 with repeated centrifugation as above. The cell pellet was then resuspended in 1× TE buffer pH 8 (100 μL), and from this the RNA was subsequently isolated with the RNeasy kit following manufacturer's protocol using lysozyme and mercaptoethanol (Qiagen). The resulting RNA pool was subjected to RQ1 RNase-free DNase (Promega) to remove any DNA before cDNA synthesis. The digestion was carried out as recommended by the manufacturer. First-strand cDNA synthesis was performed by Superscript-III reverse transcriptase (Invitrogen) following producers' recommendations.
2.6. Amplification of ribosomal 16S rRNA genes and clone library construction
Ribosomal 16S RNA genes were amplified from both filamentous material and spring fluid filtered on 0.2 μm filters by using the archaea-specific primers 20F (5′-TCCGGTTG ATCCTGCCGGA-3′) and 958R (5′-YCCGGCGTTGAMTCC AATT-3′) (DeLong, 1992) and the prokaryotic primers 515F (5′-GTGYCAGCMGCCGCGGTAA-3′) (Moyer et al., 1998) and 1408R (5′-GACGGGCGGTGTGTRCAA-3′) (Amann et al., 1995). The prokaryotic primer pair 515F/1408R covers 93% and 94% of bacterial and archaeal 16S rRNA sequences from the Ribosomal Database Project II (RDPII) without mismatch. Both PCRs were performed at 95°C/5 min followed by 25 cycles at 95°C/30 s, 55°C/30 s, and 72°C/60 s, followed by a single 72°C step for 10 min and cooling to 4°C. The PCRs (20 μL) contained 0.17 mM of each dNTP, 1.75 mM MgCl2, 0.7 mM of each primer, 1× PCR buffer, and 0.5 U of Taq polymerase (Fermentas). The PCR-amplified 16S rRNA gene fragments were cloned (TOPO TA system, Invitrogen), and inserts were amplified from the clones by using PCR (program as above) with M13 primers supplied with the kit (Invitrogen). A total of 11 different 16S rDNA clone libraries were made from Trollosen and Fisosen (Table 2). Randomly chosen M13 PCR products (n=320) of correct length from these 11 clone libraries were sequenced with BigDye v3.1 chemistry and run on an ABI 3730xl sequencer (Applied Biosystems). All the sequenced clones were checked for chimeras with RDPII (Ribosomal Database Project II; v8.1) Chimera Check (Cole et al., 2007) and the Bellerophon Chimera Detection Program (Huber et al., 2004).
na, not analyzed.
Prokaryotic and archaeal refer to the primer sets used for the 16S rRNA genes amplified.
2.7. Taxonomic assignment of 16S rRNA gene fragments
The 320 sequenced ribosomal 16S rRNA gene fragments were taxonomically assigned with use of a reference database (Urich et al., 2008, Lanzén et al., 2011) and the program MEGAN (Huson et al., 2007). Briefly, a blastn search was performed against a local SILVA database (Release 100) (Pruesse et al., 2007), and the output was subsequently analyzed with MEGAN. The following LCA parameters were used for taxonomic assignments on the 50 best BLAST hits: minimum support 1, minimum bit score 150, top percent 10. The SILVA taxonomy was used for most bacterial groups, with the exception of the ɛ-proteobacteria, the Acidobacteria, and Chloroflexi, where a hand-curated taxonomy was implemented (Lanzén et al., 2011). The archaeal taxonomy was similarly hand-curated, according to recent literature (Lanzén et al., 2011). Blastn searches were performed against the Genbank nonredundant nucleotide database to identify best BLAST hits of selected sequences.
2.8. Real-time quantitative PCR
Archaeal 16S rDNA genes were quantified by using primers 915F (5′-AGGAATTGGCGGGGGAGCA-3′) (Blank et al., 2002) and 1059R (5′-GCCATGCACCWCCTCT-3′) (Stahl and Amann, 1991). Each reaction (20 μL) contained 1 × QuantiTech Sybr Green PCR master mixture (Qiagen), 0.5 mM of each primer, and 1 μL template DNA (approximately 10 ng DNA). The thermal cycling program was 15 min at 95°C, 40 cycles of 94°C for 15 s, 61°C for 30 s, 72°C for 30 s. Quantification standard consisted of a dilution series of purified PCR amplicon from a conventional PCR by using primer pair 20F (DeLong, 1992) and 1392R (5′-ACGGGCG GTGTGTRC-3′) (Lane et al., 1985) with a pure culture of Archeoglobus fulgidus as template. PCR conditions were as previously described. The standard ranged between 17.8×10 and 1.78×107 16S rDNA copies/μL. Genomic DNA from E. coli was used as negative control. The R 2 value for the standard curve was 0.997 and slope value was −3.157, giving an estimated amplification efficiency of 107%.
Bacterial 16S rRNA genes were quantified by using primers 338F (5′-ACTCCTACGGGAGGCAGC-3′) and 519R (5′-TTACCGCGGCKGCTG-3′) (Ovreas and Torsvik, 1998) and following the protocol described by Einen and colleagues (2008). Quantification standard consisted of a dilution series (between 1.38×10 and 1.38×107 16S rDNA copies/μL) of a known amount of purified PCR product obtained by using the 16S rDNA-specific primers 8F (5′-AGA GTTTGATCCTGGCTCAG-3′) (Edwards et al., 1989) and 1392R (Lane et al., 1985). Genomic DNA from Sulfolobus sulfataricus was used as negative control. The R 2 value for the standard curve was 0.999 and slope value −3.403, giving an estimated amplification efficiency of 97%. All qPCR experiments were conducted in a Step-OnePlus real-time PCR system (Applied Biosystems) with SYBRGreenI as fluorescent dye. To confirm product specificity, melting curve analyses were performed after each run for all experiments, and each qPCR setup contained samples, standard series, negative control, and blank, all in duplicates. All DNA concentrations were measured with a Nanodrop ND-1000 (Nanodrop Technologies). In the conversion from numbers of bacterial 16S rDNA copies per gram to numbers of bacterial cells per gram (Table 3), we divided the copies per gram number by 2.09 for Fisosen, as the dominating sulfur oxidizers detected in this spring belong to the Helicobacteriales (Sulfurovum genus), which have been shown to have an average content of 2.09 copies of the 16S rDNA in their genome (Lee et al., 2009). The cells per gram in the Trollosen spring was calculated by dividing the number of 16S rDNA copies per gram by 3, as the spring was shown to be dominated by organisms of the Thiotrichales, which have an average of three 16S rDNA copies in their genome (Lee et al., 2009). The archaeal population being dominated by Thaumarchaeota representatives is known to have only one 16S rDNA gene per genome, so for the archaea the number of 16S rDNA copies per gram is assumed to correspond to archaeal cells per gram.
16S rDNA copies are given per gram of sample material wet weight, exception is Trollosen 0.2 μm filter where the copy numbers are given per milliliter sample material. For the Fisosen sample the number of bacterial cells per gram was calculated assuming an average of 2.09 16S rDNA copies per cell, as the dominating species belonged to the Sulfurovum genus (Lee et al., 2009). For the Trollosen samples, the number of bacterial cells per gram was divided by 3 as the spring was dominated by members of the Thiotrichales family with an average of three 16S rDNA copies per genome (Fig. 4) (Lee et al., 2009).
2.9. Screening for sulfur oxygenase reductase genes
The DNA from Fisosen and from Trollosen was analyzed for presence of genes encoding sulfur oxygenase reductase (SOR), a key-enzyme of sulfur-oxidizing archaea, by PCR with newly designed primers SOR-1F (5′-GTBGGBCCN AARGTNTG-3′) and SOR-2R (5′-TGCATNTCYTCRTGRTC-3′), which resulted in a DNA fragment of approximately 200 bp. Gradient PCRs were performed at 95°C/5 min followed by 30 cycles at 95°C/30 s, gradient 40–50°C/30 s, and 72°C/30 s, followed by a single 72°C step for 10 min and cooling to 4°C. The PCRs (20 μL) contained 0.17 mM of each dNTP, 1.75 mM MgCl2, 0.7 mM of each primer, 1 M Betain (Sigma), 1× PCR buffer, and 0.5 U of Taq polymerase (Fermentas).
2.10. Screening for archaeal and bacterial ammonia monooxygenase (amoA) genes
The DNA from filaments, the 0.2 μm filter, plus cDNA of Fisosen and Trollosen were further analyzed for the presence of genes encoding the archaeal and bacterial ammonia-oxidizing protein, amoA. The archaeal amoA gene was PCR amplified by using the amoA-specific primers CrenAmoA19F (5′-ATGGTCTGGCTWAGACG-3′) (Leininger et al., 2006) and CrenAmoA616R (5′-GCCATCCATCTGTATGTCCA-3′) (Tourna et al., 2008). For the bacterial amoA gene, the following primers were used in the PCR: 1F (5′-GGGGTT TCTACTGGTGGT-3′) and 2R (5′-CCCCTCKGSAAAGCCTT CTTC-3′) (Rotthauwe et al., 1997). The PCR mixtures and conditions were the same as those for the prokaryotic 16S rDNA clone library above, with the use of 25 cycles for archaeal amoA and 35 cycles for bacterial amoA. PCR-amplified archaeal amoA genes were cloned (TOPO TA system, Invitrogen), and inserts were amplified from the clones by using PCR (program as for the prokaryotic 16S rDNA library) with M13 primers supplied with the kit (Invitrogen). Archaeal amoA clone inserts of correct length, obtained from 0.2 μm filter, were sequenced with BigDye v3.1 chemistry and run on an ABI 3730xl sequencer (Applied Biosystems). The bacterial amoA genes were not cloned or sequenced.
2.11. Screening for 16S rRNA genes of Nitrospira
DNA from Fisosen and Trollosen were analyzed for the presence of ribosomal 16S rRNA genes from chemolithoautotrophic nitrite oxidizers belonging to the Nitrospira genus. The screening was done by PCR with the general bacterial primer 616F (also termed Eub8F or 27F: 5′-AGAGTTTGATCCTGGCTCAG-3′) together with the Nitrospira-specific primer 1158R (5′-CCCGTTMTCCTGGGCAGT-3′) (Maixner et al., 2006). The PCR mixture and conditions were the same as those for the prokaryotic 16S rDNA clone library above but with an annealing temperature of 54°C. PCR-amplified 16S rRNA genes (10 from each spring) were cloned, M13-amplified, and sequenced as for the amoA gene.
2.12. Elemental analysis of bacterial cells in electron microscope
Frozen biomass specimens from the Fisosen spring, which were characteristic of the starlike filaments, were thawed and applied to aluminum grids (100 mesh, Agar Scientific) supported with a carbon-coated Formvar film and air-dried. The filaments were viewed and analyzed for light elements in a Philips CM200 electron microscope with energy dispersive x-ray analysis (EDAX) (Fig. 3, right panels). The microscope was operated at an acceleration voltage of 80 kV, a spot size of 14 nm (spot size 3), and an accumulation time (live time) of 30 s. The light element detection system consisted of an EDAX detector DX-4 combined with SIS Soft Imaging Software.

The filamentous organisms dominating Fisosen and Trollosen thermal springs. Macroscopic pictures of the featherlike filaments of Fisosen (upper left panel) and Trollosen (lower left panel). Light microscopy picture (120×) of the long filaments reveals organisms arranged in a star- or rosetta-like fashion (upper middle panel). Increased magnitude (300×, lower middle panel) highlights the many refractive particles present. EDAX revealed the predominant presence of intracellular sulfur granules (upper right panel) and phosphorous-filled vacuoles (lower right panel) in the long filaments. Color images available online at
2.13. Nucleotide sequence accession numbers
The bacterial 16S rRNA gene sequences reported in this study have been deposited in GenBank with the following accession numbers JF837612–JF837809, the archaeal 16S rRNA sequences as JN083865–JN083985 and the archaeal amoA sequences as JF286602–JF286607.
3. Results
3.1. Spring descriptions and physicochemical parameters
The Trollosen and Fisosen thermal springs are located 400 m apart close to the seashore Fisneset at Sørkapp in the protected South-Spitsbergen National Park of Svalbard (Fig. 1) and are recognized by a strong odor of H2S (Lauritzen and Botrell, 1994). The two sulfidic springs differ from one another both in appearance and geochemical composition. The Trollosen spring has a cave opening of approximately 3×1 m and discharged approximately 8 m3/s of 6°C fluid during our sampling in June 2006. The outflow of this spring generates a small river that reaches the seashore approximately 60 m downstream of the cave opening (Fig. 1), which is an entry to a yet to be described cave system below. Large amounts of whitish filamentous material were floating in the water flowing out from the underlying cave system. In contrast, the Fisosen spring has a small opening of 20 cm in diameter and a water temperature of ∼13°C (Fig. 1). The spring discharged about 1 L/s of clear fluid. At sampling time, the spring was partly covered with snow. In contrast to Trollosen, the outflow of Fisosen spring was covered with white filaments attached to the rocks. Abiotic analyses of both spring fluids are given in Table 1. The spring fluids of Fisosen and Trollosen were different in H2S content; whereas Fisosen contains high amounts of H2S (115 μM), it was chemically not detectable in Trollosen, though the water smelled of H2S (Table 1). Both springs contained a high concentration of ammonia; 104 and 188 μM in Fisosen and Trollosen, respectively (Table 1). Nitrate and phosphate concentrations were below detection limits in both springs. The dominating anion was chloride; 957 and 1997 mg/L in Fisosen and Trollosen, respectively (Table 1).
3.2. Microscopic analyses and enumeration of Bacteria and Archaea by quantitative PCR
The filamentous material from both springs was of a featherlike appearance (Fig. 3, left panels). Under the microscope, this was predominantly composed of long filamentous organisms arranged in a star- or rosetta-like fashion (Fig. 3, middle panels). Many refractive particles were present within the filaments, as observed by light microscopy, that resembled vacuoles and intracellular granules. They were distinguishable as electron-dense particles by electron microscopy. EDAX revealed the predominant presence of sulfur granules, in addition to vacuoles containing calcium phosphate or sodium phosphate (Fig. 3, right panels).
Enumeration of Bacteria and Archaea was carried out by qPCR of 16S rRNA genes (Table 3). The enumeration clearly showed a dominance of Bacteria over Archaea, in which Archaea constituted less than 1% of the total communities in both, except for the Trollosen spring fluid where Archaea constituted about 4%. The numbers of cells per gram (wet weight) were 3.3×108 and 1.2×109 in the filaments of Fisosen and Trollosen, respectively. In the spring fluid of Trollosen, the estimated number of cells was 2.2×105 per milliliter.
Additionally, microscopy revealed the presence of nematodes in the Trollosen spring, but they were not detected in Fisosen (data not shown).
3.3. Detection and cloning of ribosomal 16S rRNA genes of Bacteria and Archaea
We investigated the prokaryotic community of the thermal springs for ribosomal 16S rRNA genes by using both prokaryotic- and archaea-specific primers (see Materials and Methods for details). The community-profiling analyses were performed on filamentous samples and spring fluids. The organisms in the spring fluid were harvested by a two-step filtration process (10 μm and subsequently 0.2 μm) in order to separate particles and the featherlike filaments from the smaller organisms. Two different DNA extraction procedures were applied on the filament fraction of Fisosen to investigate methodological influence on the analysis; either a combined mechanical and chemical cell lysis involving bead beating or immobilization of the cells into a matrix before chemical cell lysis (see Materials and Methods). Whereas a 16S rRNA gene library of the latter reported only Sulfurovum, a library from the combined mechanical and chemical lysis reported 96% Sulfurovum and 4% Thiotrix, which indicates that filamentous Thiotrix might be difficult to lyse when applying only chemical methods.
Concerning the filamentous samples, both DNA and RNA were isolated and analyzed to focus on both the composition of the community (DNA) as well as its active fraction at the time of sample collection (RNA).
A total of eleven 16S rRNA clone libraries were made from the two springs, which resulted in 172 analyzed clones from Fisosen and 148 clones from Trollosen (Table 2). The taxonomic assignments of the 320 sequenced ribosomal genes obtained in the 16S rDNA clone libraries were done with the program MEGAN and the SILVA SSU Ref 100 database as reference (Pruesse et al., 2007) according to Urich and colleagues (2008) and Lanzén and colleagues (2011).
The data from the 16S rDNA gene amplifications for which the prokaryotic primers (515F/1408R) were used show that both Fisosen and Trollosen are dominated by Bacteria, as no archaeal 16S rRNA gene fragments were detected among the 199 sequenced ribosomal genes (Fig. 4). The filamentous fractions of both Fisosen and Trollosen are mainly composed of sulfur oxidizers; Fisosen is dominated by members of the Sulfurovum genus, (ɛ-proteobacteria) (Fig. 4, columns 2, 3, and 4), while the Trollosen filaments are dominated by organisms from the Thiotrichaceae family (Thiotrix genus) (γ-proteobacteria) (Fig. 4, columns 6 and 7). For Fisosen, there is a notable difference between the community analyses done on the RNA level compared to the DNA level, with a higher proportion of Thiotrichaceae assigned on the RNA level. It has been proposed that ribosomal RNA can be used as a proxy of the active population at the sampling point due to their rapid turnover rates, whereas the ribosomal DNA reflects the abundance of cells regardless of their activity state (Kramer and Singleton, 1993; Roussel et al., 2009). The cDNA library indicates that, even though Sulfurovum and Thiotrix are present in different amounts, they are equally active (Fig. 4, column 3). However, it is noteworthy that the number of ribosomes per cell might vary a lot between different taxa (especially when comparing giant sulfur bacteria with normally sized bacteria) and between different physiological states (Kramer and Singleton, 1993).
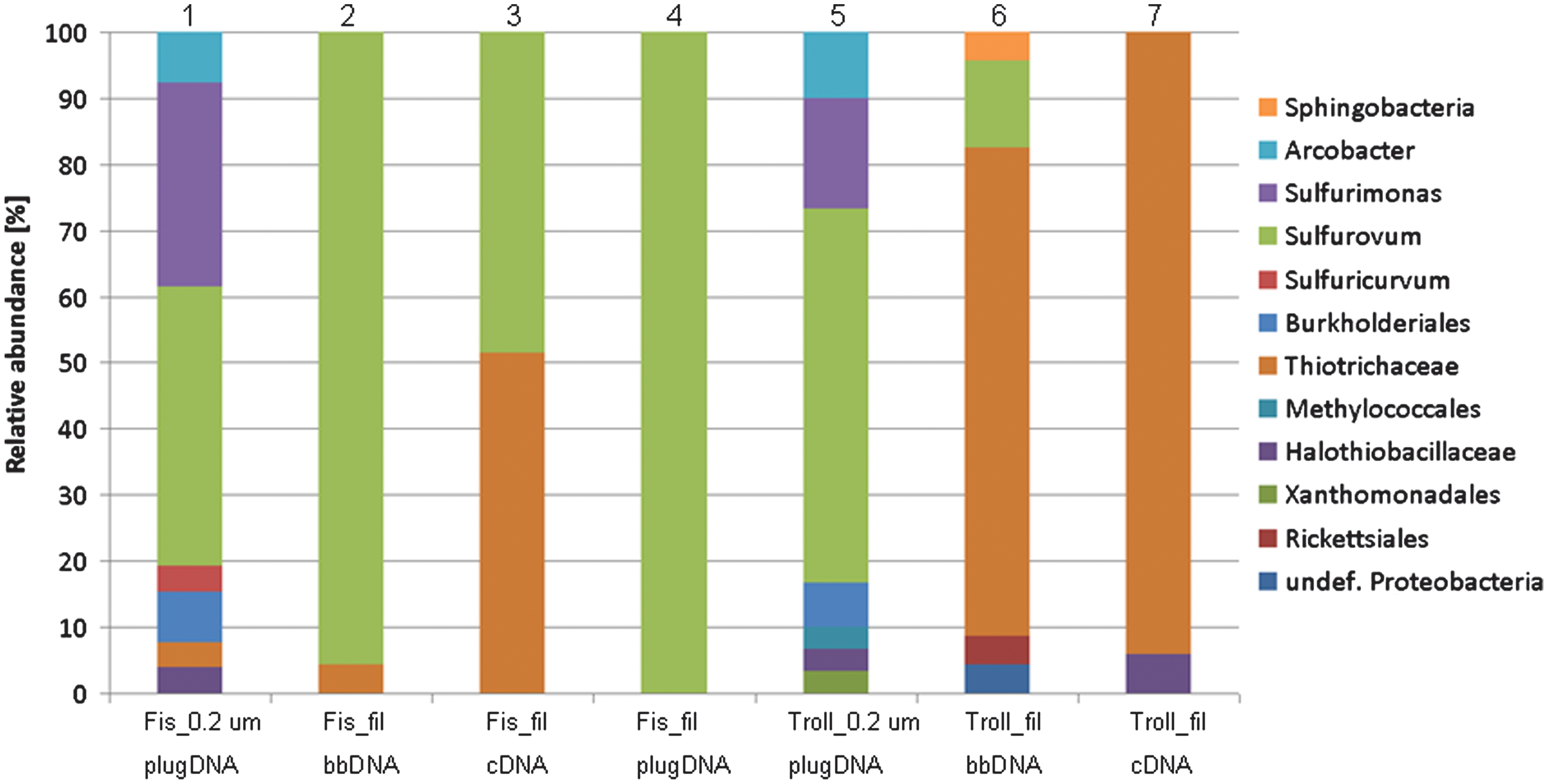
16S rRNA community profiles of Trollosen and Fisosen obtained with prokaryotic primers (515F/1408R). Different taxonomic resolutions are displayed; where possible and necessary, genera are shown, whereas in other cases lower taxonomic resolution is displayed. Different sample material and different DNA/RNA isolation procedures were carried out. Fis, Fisosen; Troll, Trollosen; 0.2 μm_plugDNA, DNA from spring liquid obtained by filtration onto a 0.2 μm filter where the filter scrape-off was immobilized into agarose plugs before cell lysis and DNA extractions; fil_bbDNA, DNA from filamentous material where bead beating was used to lyse cells and extract DNA; fil_plugDNA, DNA from filamentous material that was homogenized and immobilized into agarose plugs before cell lysis and DNA extraction; fil_cDNA, cDNA made from RNA extracted from the filaments. Color graphics can be found at
Most of the Sulfurovum 16S rRNA gene sequences were closely related to each other (98–100% 16S rRNA sequence identity) and belonged to a single operational taxonomic unit that populated both springs. The identity to the type strain Sulfurovum lithotrophicum, a mesophilic sulfur- and thiosulfate-oxidizing bacterium isolated from sediments in a hydrothermal field in the mid-Okinawa Trough (Inagaki et al., 2004), was 94–95%. The closest environmental sequence was 99% identical and was obtained from the Frasassi cave in Italy (accession No. EF467592). Similarly, most of the Thiotrix sequences were closely related to each other (98–100% sequence ID) and were closely related (98–100% sequence ID) to Thiotrix lacustris, Thiotrix caldifontis, and Thiotrix fructivorans. The T. lacustris and T. caldifontis have recently been isolated from sulfidic springs in the Caucasus (Chernousova et al., 2009). Only in the Trollosen filaments, a few sequences could not be assigned to sulfur oxidizers but were affiliated with potentially heterotrophic bacteria that belong to Sphingobacteria and Rickettsiales (Fig. 4).
The diversity of the microbial community was higher in the springwaters than in the filaments (Fig. 4, columns 1 and 5 versus 2, 3, 4, 6, and 7). In the water of both springs, about 85% of the 16S rRNA sequences belonged to the Campylobacteriales order of ɛ-proteobacteria (Sulfurovum, Sulfurimonas, Arcobacter, and Sulfuricurvum), while the remaining ones belonged to Thiotrichaceae (γ-proteobacteria), Halothiobacillaceae (γ-proteobacteria), in addition to Xanthomonadales (γ-proteobacteria) and Burkholderiales (β-proteobacteria) (Fig. 4, columns 1 and 5). One sequence from Trollosen belonged to the genus Methylobacter within the order Methylococcales (γ-proteobacteria).
Two archaeal 16S rRNA clone libraries were made from each spring (Table 2; Fig. 5), one based on DNA from filaments, the other made from spring fluid DNA (see Materials and Methods). For both the filaments and the spring fluid in Trollosen, as many as 85–90% of the 16S rRNA genes belonged to organisms of the recently suggested Thaumarchaeota kingdom (Brochier-Armanet et al., 2008; Spang et al., 2010) (Fig. 5, columns 3 and 4), namely, the Groups I.1a and I.1b. For the filaments and fluid of Fisosen, approximately 50% of the organisms belonged to the Thaumarchaeota kingdom (Fig. 5, columns 1 and 2). Furthermore, in the Fisosen fluid 20% were affiliated with the Deep Sea Hydrothermal Vent Euryarchaeotic Group 3 (DHVE-3), 10% to the Miscellaneous Crenarchaeotic Group (MCG), while the remaining 17% were classified as anaerobic methane-oxidizing Euryarchaeotes of Group 1a (98% identity to sequence AM745235), undefined Methanomicrobia and Crenarchaeota of the Marine Benthic Group B. The Fisosen filament fraction (Fig. 5, column 2) contained, in addition to the before-mentioned Thaumarchaeota, 17% Marine Benthic Group D, 13% MCG; and the remaining organisms belonged to Marine Benthic Group B, Terrestrial Miscellaneous Euryarchaeota Group (TMEG), and undefined Euryarchaeota.

16S rRNA community profiles of Trollosen and Fisosen obtained with archaeal primers (20F/958R). Different sample material and different DNA isolation procedures were carried out. Fis, Fisosen; Troll, Trollosen; fil, filamentous material; 0.2 μm, the DNA analyzed was isolated from spring liquid filtered down on a 0.2 μm filter; plugDNA, the harvested cells were immobilized into agarose plugs before performing cell lysis and DNA extractions; bbDNA, Bead beating was used to lyse cells and extract DNA. ANME-1a, anaerobic methane-oxidizing Euryarchaeotes of group 1a. Color graphics can be found at
In the spring fluid of Trollosen, the remaining sequences belonged to DHVE-3, MCG, and the Marine Hydrothermal Vent Group, whereas in the filaments they were affiliated with Marine Benthic Groups D and E, and two families of methanogens: the Methanosarcinaceae and the Methanomicrobiaceae (Fig. 5, column 4). The latter two were closely related to the genera Methanolobus (98% sequence ID) and Methanoplanus (95% sequence ID). It should be noted that we detected no sequences related to taxa with known sulfate reducers in any clone library.
3.4. Screening for nitrifiers and archaeal sulfur oxidizers
We screened for archaeal and bacterial ammonia oxidizers by using primers specific for genes encoding the alpha subunit of the key-enzyme ammonia monooxygenase (amoA) of both groups (see Materials and Methods). Archaeal amoA genes were detected in the planktonic fraction (fluid) of both Trollosen and Fisosen, but neither in the filaments nor cDNA from the two springs. Subsequent cloning and sequencing of seven archaeal amoA genes revealed two sequence types with highest similarity to amoA sequences from the marine cluster (96% identity to accession No. EU023046 and 98% identity to EU852663). Bacterial amoA genes were only detected in the planktonic fraction of Trollosen.
In addition, we screened for members from the bacterial genus Nitrospira, known to perform the oxidation of nitrite to nitrate, using PCR with specific 16S rRNA primers (see Materials and Methods). 16S rRNA genes were detected and were 94–96% identical to the chemolithoautotroph Nitrospira moscoviensis, one of the few isolates from this poorly understood group of organisms (Lucker et al., 2010).
We screened for the presence of genes encoding the enzyme SOR, the key-enzyme in the sulfur oxidation pathway of some archaea (Kletzin et al., 2004; Urich et al., 2006). However, no positive PCR product was obtained with our newly designed degenerated primers (see Materials and Methods).
4. Discussion
The archipelago of Svalbard is situated in the High Arctic harbors around 50 springs, some of which are thermal and some not (Liestøl, 1976). This first microbiological description of terrestrial thermal springs on Svalbard, which focuses on the Trollosen and Fisosen springs at Sørkapp in the South-Spitsbergen National Park (76°N), revealed two sulfidic environments in which substantial amounts of biomass were generated. The two springs are located along the shoreline (Figs. 1 and 2); Trollosen (6°C, pH 7.0) discharged 8 m3/s of clear water at sampling time from a completely undescribed underlying cave system with enormous amounts of biomass from filamentous microorganisms (Thiotrix) being washed out. Fisosen (13°C, pH 7.5) is a tiny spring that discharged about 1 L/s of clear water, with filamentous material attached to the rocky stream bed and the spring edges and present in the outflow (Fig. 3, left panels). The water in Trollosen is subglacial meltwater from the nearby Vitkovskij glacier mixed with hot brine from the underlying karst aquifer, which penetrates the permafrost (Lauritzen and Botrell, 1994). Of the two springs, Fisosen is believed to better represent the deep subpermafrost aquifer (Lauritzen and Botrell, 1994).
Ours and previous chemical analyses of the spring fluids showed both Fisosen and Trollosen to have a fluid composition significantly different from what is generally found in Arctic seawater (Table 1). The level of chloride, in particular, greatly exceeds that of seawater by 2 orders of magnitude. The concentrations measured in this study are highly similar to the previous measured values by Lauritzen (1996) and Lauritzen and Botrell (1994), which indicates a stable fluid composition (Table 1). The origin of the salts in the springs is unclear but might either be evaporate strata deep in the aquifer system, formation water expelled from sandstones, or formation water mixed with recent seawater within the underlying karst aquifer (Lauritzen, 1996).
Both springs are sulfidic, although the concentration of H2S differs greatly between Fisosen and Trollosen. The lower concentration in Trollosen is likely caused by oxidation of the H2S through mixing with larger volumes of oxygenated subglacial meltwater, which is likely not the case for Fisosen. Low oxygen concentration in Fisosen is likewise supported by the finding of anaerobic methanotrophic archaea in the clone libraries. The origin of the H2S is elusive, although it has been speculated, based on isotopic fractionation data, that it might be of biological origin (Lauritzen and Botrell, 1994). Interestingly, we did not find any sequence related to known sulfate reducers in the filaments; but in the planktonic fractions of both springs, 18–30% of the bacterial communities were related to Sulfurimonas. The Sulfurimonas are known mainly for their ability to oxidize sulfur, but under anoxic conditions these organisms seem to be able to produce H2S from hydrogen and elemental sulfur (Sievert et al., 2008). Likewise, sulfate reduction could be performed by some of the uncharacterized groups identified in the clone libraries, such as the Marine Benthic Group B, and thus a biological origin is not unlikely.
The Thiotrix filaments had the typical star- or rosetta-like morphology and contained refractive intracellular granules and vacuoles (Fig. 3, middle panels). EDAX showed the granules to be composed of sulfur (Fig. 3, right panels). In some filaments, the vacuoles contained phosphorus, likely in the form of phosphate. As the phosphate concentrations in the spring waters are low (not detectable by anion chromatography), intracellular storage can be considered an efficient strategy to prevent phosphorus limitations. Intravacuolar polyphospate inclusions have been reported from various giant sulfur bacteria (Schulz and Schulz, 2005, and references therein).
Consistent with the microscopic observations, the prokaryotic 16S rDNA and rRNA clone libraries identified Thiotrix as the dominant genus in Trollosen filaments. Interestingly, the situation was not as clear in Fisosen, where a large proportion of the 16S rDNA clone library was composed of Sulfurovum. Members of this genus are not typically found as filaments but rather as small rods, in contrast to Thiotrix. This incongruence might be explained by large numbers of Sulfurovum cells that grow attached to the Thiotrix filaments in Fisosen, something that is more unlikely in Trollosen as the velocity of the water flow is unfavorable for settling.
This different distribution of the dominating sulfur oxidizing groups among the two springs, as reported by the clone libraries, might be related to variations in the sulfide/oxygen ratios and the physical water flow, which is similar to what was described for sulfur-oxidizing communities in the Frasassi cave system in Italy (Macalady et al., 2008). There, it was suggested that high sulfide/oxygen ratios are preferred by ɛ-proteobacteria, such as Sulfurovum, while lower sulfide/oxygen ratios are preferred by Thiotrix (γ-proteobacteria), as an example of niche separation between the two groups.
Members of Thiotrix and Sulfurovum are both autotrophic organisms that fix CO2 via the Calvin-Benson-Bassham cycle and the reverse tricarboxylic acid cycle (Campbell et al., 2006; Teske and Nelson, 2006); therefore, they likely generate the majority of the biomass in the spring ecosystems through chemolithoautrophy, as is the case with other sulfidic caves and springs (Chen et al., 2009; Niederberger et al., 2009a, 2009b) and hydrothermal vent ecosystems (Campbell et al., 2006).
Interestingly, the planktonic fractions from both springs were rather similar in composition, dominated with different groups of sulfur oxidizers Sulfurovum, Sulfurimonas, and Arcobacter (ɛ-proteobacteria) (Fig. 4). In addition to the above-mentioned groups, sequences related to sulfur oxidizers Sulfuricurvum (ɛ-proteobacteria) and Halothiobacillus (γ-proteobacteria) were present in the clone libraries. In summary, the vast majority of the prokaryotic 16S rDNA clone library represented sulfur-oxidizing bacteria belonging to the γ- and ɛ-classes of the proteobacteria (Fig. 4).
Although we have not screened for key-enzymes of bacterial sulfur oxidation pathways like SOX or reverse dissimilatory sulfite reductase (Friedrich et al., 2005), the close phylogenetic relationship of the 16S rRNA sequences to known sulfur-oxidizing taxa, the presence of intracellular sulfur globules in the filaments, and the qPCR results that indicate a Bacteria-dominated system provide strong evidence that sulfur oxidation is the major energy source in these Arctic springs.
To test whether Archaea (although with approximately 1% only a minor part of the community) contributed to the sulfur oxidation, we screened for the presence of genes encoding the enzyme SOR. This is the key-enzyme in the sulfur oxidation pathway of some archaea (Kletzin et al., 2004; Urich et al., 2006). However, no positive PCR product was obtained.
In the case of Fisosen and Trollosen, the sulfur oxidation apparently involves the oxidation of H2S, which forms sulfuric acid (H2SO4). The area is founded on limestone bedrock (CaCO3), and though unknown for the sulfidic springs investigated here, it has been shown in other cave systems that the produced sulfuric acid can react with the carbonate in the limestone to form gypsum and carbonic acids and thereby be partly responsible for the architecture of the caves by creating porosity in the rocks (Hill, 1995; Macalady et al., 2006).
High concentrations of ammonia were measured in both springs (104 and 188 μM in Fisosen and Trollosen, respectively; see Table 1), whereas no nitrate was detected. The ammonia concentration is, as for chloride, almost 2 orders of magnitude higher than what is normally observed in seawater. An explanation for these high values could be the production of ammonia by nitrate reduction through the γ- and ɛ-proteobacterial sulfur oxidizers; members of both classes are reported to utilize this type of anaerobic respiration under suboxic conditions (Campbell et al., 2006; Teske and Nelson, 2006). Similarly, high ammonia concentrations were reported from the sulfidic waters of Movile cave (Chen et al., 2009) and Frasassi cave (Macalady et al., 2006) and from the water above hydrothermal vent sediments (Campbell et al., 2006), where γ- and/or ɛ-proteobacterial sulfur oxidizers were dominant.
It seems reasonable that the ammonia in the springs is being utilized by ammonia-oxidizing microorganisms. In that respect, it is interesting that the archaeal communities in both Fisosen and Trollosen were dominated by members of Groups I.1a and I.1b of the recently suggested Thaumarchaeota kingdom (previously classified to the Crenarchaeota) (Brochier-Armanet et al., 2008). Isolates and enrichments of both groups are known to gain their energy by the oxidation of ammonia to nitrite (Könneke et al., 2005; Hatzenpichler et al., 2008; Reigstad et al., 2008; Urich et al., 2008), thereby catalyzing the first step in nitrification in many environments. However, it has recently been shown that not all members of the Thaumarchaea are nitrifiers and harbor the amo genes (Muller et al., 2010). Accordingly, a PCR screening for the subunit A of the archaeal ammonia monooxygenase gene revealed its presence in the filtered fractions of both springs; all sequences clustered within the marine group, along with the amoA genes of Nitrosopumilus maritimus and Cenarchaeum symbiosum. Likewise, bacterial amoA genes were detected in the Trollosen fluid sample. Additionally, we detected 16S rRNA sequences related to Nitrospira, a widespread genus shown to include nitrite oxidizers (Lucker et al., 2010). Although there are additional groups that might be involved in nitrification in the springs for which we have not screened [e.g., the recently described genus Nitrotoga (Alawi et al., 2007)], our data provide evidence for the potential of complete nitrification performed by a similar consortium of microbial groups as found in other cave systems. One example is the Movile Cave (Romania), with comparably high ammonia concentrations, where nitrification was also detected. There, ammonia oxidation was related to Nitrosomonas and nitrite oxidation to Nitrotoga and Nitrospira (Chen et al., 2009); no archaeal ammonia oxidizers were found.
In contrast, Koch et al. (2006) identified archaea affiliated with the Group I.1a of Thaumarchaeota as a minor, but stable, component of Thiotrix-dominated filaments in a sulfidic spring in Germany. Perreault and colleagues detected Groups I.1a and I.1b in two High Arctic sulfidic springs in Canada (Perreault et al., 2007). However, none of the studies screened for the presence of the archaeal amo genes. Most recently, giant, filamentous Thaumarchaeota was shown to be living in mangrove swamps rich in H2S (Muller et al., 2010). One of the phylotypes was found in close association with sulfur-oxidizing bacteria, which were densely covered with bacterial rods. Interestingly, no archaeal amoA gene could be detected (Muller et al., 2010). It will be interesting to investigate whether similar giant archaea are present in Fisosen and Trollosen and whether these giant archaea are actively involved in nitrification.
Nevertheless, our data provide the basis for some speculation regarding a possible link between the sulfur and nitrogen cycles in the springs. Through anaerobic nitrate respiration by the sulfur-oxidizing bacteria, ammonia might be formed, which can be the substrate for the nitrifying prokaryotes that recycle the nitrate needed for the sulfur oxidizers and thereby generate a complete nitrogen cycle within the community.
In both springs, we detected 16S rRNA sequences related to methanotrophs. In Trollosen, a sequence was closely related to Methylobacter, an aerobic methanotrophic bacterium, whereas in Fisosen anaerobic methanotrophic archaea of the ANME-1 group comprised a small fraction of the planktonic population. Sequences closely related to the methanogenic archaeal genera Methanolobus and Methanoplanus were detected in Trollosen filaments. Although the methane concentrations in the spring water have not been determined, the molecular data are indicative of an internal methane cycle and methane as a likely additional, though minor, source of energy and carbon for the microbial community.
5. Conclusions
The presence of mainly lithoautotrophic organisms in the Arctic springs of Fisosen and Trollosen indicates that the community in the cave system in, and below, both springs is largely independent of the impact of organic matter from the surface. Both springs are dominated by autotrophic sulfur-oxidizing bacteria, which produce the majority of biomass and sustain the small fraction of heterotrophic prokaryotes as well as higher eukaryotes such as nematodes. Ammonia- and nitrite-oxidizing archaea and bacteria are also likely to be primary producers, although they are numerically less important. The sulfur and nitrogen cycles in these thermal springs might be tightly linked through microbial action. Besides reduced sulfur and nitrogen species, methane might be a third source of energy and carbon for the community.
These springs might provide valuable model systems with which to study subsurface ecosystems on Earth as well as analogues for putative fossil or extant martian subsurface ecosystems based on chemolithoautotrophy.
Footnotes
Acknowledgments
We are thankful to Sysselmannen at Svalbard for the possibility to sample in the protected Arctic area of Sørkapp in South-Spitsbergen National Park. Logistical support was provided by the Norwegian Polar Institute through the vessel MS Lance and the helicopter pickup at Sørkapp. Mikal Heldal [University of Bergen (UoB)] is acknowledged for performing the EDAX, as is Lill Knudsen (UoB) for excellent technical assistance. We are grateful to the Bergen Geoanalytical Facility (UoB) for the ICP-MS and anion-chromatography data. The work was supported by the Norwegian Research Council, Grant 172206 to C.S. and L.J.R., and by a European Molecular Biology Organisation (EMBO) long-term postdoctoral fellowship (Grant ALTF 468-2006) to T.U.
Abbreviations
DHVE-3, Deep Sea Hydrothermal Vent Euryarchaeotic Group 3; EDAX, energy dispersive X-ray analysis; ICP-MS, inductively coupled plasma mass spectrometer; MCG, Miscellaneous Crenarchaeotic Group; qPCR, quantitative polymerase chain reaction; RDPII, the Ribosomal Database Project II; SOR, sulfur oxygenase reductase; TMEG, Terrestrial Miscellaneous Euryarchaeota Group.