
Abstract
The compelling evidence for an ocean beneath the ice shell of Europa makes it a high priority for astrobiological investigations. Future missions to the icy surface of this moon will query the plausibly sulfur-rich materials for potential indications of the presence of life carried to the surface by mobile ice or partial melt. However, the potential for generation and preservation of biosignatures under cold, sulfur-rich conditions has not previously been investigated, as there have not been suitable environments on Earth to study. Here, we describe the characterization of a range of biosignatures within potentially analogous sulfur deposits from the surface of an Arctic glacier at Borup Fiord Pass to evaluate whether evidence for microbial activities is produced and preserved within these deposits. Optical and electron microscopy revealed microorganisms and extracellular materials. Elemental sulfur (S0), the dominant mineralogy within field samples, is present as rhombic and needle-shaped mineral grains and spherical mineral aggregates, commonly observed in association with extracellular polymeric substances. Orthorhombic α-sulfur represents the stable form of S0, whereas the monoclinic (needle-shaped) γ-sulfur form rosickyite is metastable and has previously been associated with sulfide-oxidizing microbial communities. Scanning transmission electron microscopy showed mineral deposition on cellular and extracellular materials in the form of submicron-sized, needle-shaped crystals. X-ray diffraction measurements supply supporting evidence for the presence of a minor component of rosickyite. Infrared spectroscopy revealed parts-per-million level organics in the Borup sulfur deposits and organic functional groups diagnostic of biomolecules such as proteins and fatty acids. Organic components are below the detection limit for Raman spectra, which were dominated by sulfur peaks. These combined investigations indicate that sulfur mineral deposits may contain identifiable biosignatures that can be stabilized and preserved under low-temperature conditions. Borup Fiord Pass represents a useful testing ground for instruments and techniques relevant to future astrobiological exploration at Europa. Key Words: Europa—Biosignatures—Life detection—Analog—Biomineralization. Astrobiology 12, 135–150.
1. Introduction
T
Sulfur as an element is chemically very versatile in its ability to exist in multiple oxidation states and thus form the basis of a diverse suite of chemolithotrophic metabolisms on Earth today (Ehrlich, 2002). These metabolisms include the oxidation, reduction, or disproportionation of sulfur compounds by members of all three domains of life (Le Faou et al., 1990; Jorgensen and Bak, 1991; Doeller et al., 2001; Friedrich et al., 2001), and they encompass both autotrophic and heterotrophic carbon metabolisms (Pepper and Miller, 1978; Kelly et al., 1997). Phylogenetic evidence that shows sulfur-metabolizing organisms in multiple deeply rooted branches of the small subunit rRNA Tree of Life potentially indicates an early origin for metabolisms that include, but are not limited to, elemental sulfur reduction, sulfate reduction, and anoxygenic photosynthesis (Canfield and Raiswell, 1999). Moreover, isotopic evidence from 3.5 billion-year-old marine deposits indicates that elemental sulfur reduction may have formed an important part of early metabolisms (Philippot et al., 2007; Thamdrup, 2007).
Geological deposits of elemental sulfur (S0) can occur either abiotically or via microbial mediation, under a wide range of environmental conditions. The common α-sulfur phase, the high-temperature β-sulfur phases, and the metastable γ-sulfur phase (a mineral known as rosickyite) are each composed of eight sulfur atoms in a ring. The three solid allotropes differ in their molecular packing and crystal structure, which produce distinctive X-ray diffraction features. The “sheared penny roll” structure (4-molecule-unit cell) of γ-sulfur gives way to the “crankshaft” structure (16-molecule-unit cell) of α-sulfur, with an associated decrease in density from 2.19 g/cm3 to 2.069 g/cm3 (Meyer, 1976).
Studies into the nature of intracellular and extracellular S0 formation by bacteria date from the late 19th century (Winogradsky, 1887). Many sulfur-producing genera are known, and some have been extensively studied, including both lithotrophic and phototrophic S0 accumulators (Blankenship et al., 2004; Dworkin et al., 2006). Accumulation of extracellular sulfur in mats of sulfide-oxidizing bacteria has been described for numerous marine and terrestrial environments where steep gradients of both sulfide and oxygen are present (e.g., Taylor et al., 1999; Wirsen et al., 2002; Engel et al., 2007; Macalady et al., 2008). Determining the possible biogenicity of S0 deposits is challenging due to the lack of characterization of the microfabrics or morphologies associated with microbial S0 formation, as well as an understanding of the environmental and biological conditions under which S0 is stabilized and preserved (Gleeson et al., 2011).
The potential for the generation and preservation of evidence of microbial processes in sulfur-rich, icy conditions relevant to Europa has remained unexamined because of the rarity of such environments on Earth. Therefore, it is valuable to investigate whether evidence of these biological processes can be detected in Arctic analog samples to understand the potential for future biosignature detection at Europa.
2. Field Site: Borup Fiord Pass
Located at 81°019N, 81°359W, on northern Ellesmere Island in the Canadian High Arctic (Fig. 1), perennial subsurface saline springs rich in sulfide and sulfate discharge through and onto glacial ice and deposit elemental sulfur (S0), gypsum (CaSO4·2H2O), and bassanite (CaSO4·½H2O) on the glacial surface (Grasby et al., 2003). Carbonate minerals, including calcite (CaCO3) and its rare polymorph vaterite, have also been detected (Grasby, 2003).

(
This system represents a unique opportunity to investigate sulfur minerals in association with ice in a terrestrial context, which may be the best-known chemical analog to the arguably sulfur-rich surface of Europa (Grasby et al., 2003; Gleeson et al., 2010; Lorenz et al., 2011). A systematic scale-integrated approach was applied previously to query orbital (Hyperion), field, and laboratory spectra to identify sulfur-rich materials precipitated on the glacier (Gleeson et al., 2010). Sulfur as the main constituent of the deposits is well represented in the Hyperion data, although minor constituents such as calcite and gypsum are partially or entirely masked. Absorption features of sulfates, where present, are shifted in wavelength because of the effects of mixing or temperature. Autonomous detection methods were successfully applied to monitor the generation and extent of the deposits (Castano et al., 2008), which show similar spectral features to the low-temperature bound-water absorptions in Europa's non-ice materials (Gleeson et al., 2010). However, neither the analysis of remote-sensing data nor stable isotopic analysis of the sulfur deposits previously collected from the site can be used to infer whether the sulfur is biogenic. For example, Grasby et al. (2003) concluded that sulfur stable isotope data indicate extensive biological sulfate reduction of sedimentary anhydrate lying at depth beneath the glacier. This process represents the probable source of the sulfide in spring waters. However, large fractionations associated with the reoxidation of sulfide to sulfur by non-phototrophic bacteria have not been reported, so sulfur isotopic evidence cannot effectively distinguish between abiotic versus biological processes.
The recent geomicrobiological cultivation of microorganisms from Borup Fiord Pass has shown that members of the microbiological community present at the site have the potential to cause the precipitation of sulfur deposits (Gleeson et al., 2011). Those experiments targeted the isolation of microorganisms potentially responsible for the production of the S0 observed in the field. The cultivated S0 biomineralizing consortia were dominated by Marinobacter and also included Shewanella, Loktanella, Rubrobacter, Flavobacterium, and Sphingomonas spp.; these species were also relevant to environmental communities on the basis of 16S rRNA sequences amplified from DNA extracted from the deposits. In laboratory cultures provided with opposing gradients of sodium sulfide and oxygen, these consortia were shown to accumulate sulfur globules along filaments and sheath-like structures, which radiated out from a central mass of spherical mineral aggregates of sulfur. These structures were not observed in abiotic controls.
The potential influence of microbiology on the mineralogy of the site, coupled with the fact that these sulfur-rich deposits can be detected and monitored by hyperspectral satellite observations, offers an opportunity to link orbital measurements to a dynamic biogeochemical process. The mineral deposits of Borup Fiord Pass allow for investigation of how materials analogous to those of the non-ice materials at Europa can be queried for potential biosignatures, whether morphological, mineralogical, or organic. Here, we summarize our investigations into the evidence for microbiological activity within the sulfur deposits of Borup Fiord Pass by the following methods: (1) optical microscopy, environmental scanning electron microscopy (ESEM), scanning electron microscopy (SEM), and scanning transmission electron microscopy (STEM) imaging of mineral morphologies to seek evidence of organic constituents and biomineralization; (2) X-ray diffraction (XRD) measurements to investigate the presence of metastable sulfur phases potentially associated with microbial activity; and (3) infrared and Raman spectroscopy of the deposits to determine the nature of organic components. Organic materials could represent components of the microorganisms themselves or any extracellular polymeric substances (EPS) generated, which could be used as potential organic biomarkers in association with biologically mediated sulfur species. We discuss the factors pertinent to future investigations of analogous materials at Europa and the potential for these materials to contain biosignatures.
3. Methods and Materials
Samples of off-white, very fine-grained deposits from alongside channels on the surface of the glacier were collected aseptically with sterilized spatulas from spring deposits during a field visit by S.E. Grasby and K.E. Wright in July of 2009. These materials were returned from the field cold, and subsets of each were subsequently stored at 4°C and −80°C. The following analyses were carried out to evaluate whether these samples preserved evidence of microbial presence or activities.
3.1. Aqueous geochemistry
Water samples were collected from the actively flowing spring site. Unstable parameters (temperature, pH, dissolved O2) were measured in situ. Total dissolved sulfide was measured gravimetrically through precipitation of CdS. For chemical analyses, water samples were passed through 0.45 μm filters and stored in the dark at 4°C in high-density polyethylene bottles until analyzed. Samples for cation analyses were acidified with ultrapure nitric acid to pH<2.
Subsequent chemical analyses were carried out at the Geological Survey of Canada. Alkalinity was determined by a standard end-point titration. Anions were measured by ion liquid chromatography, and cations were measured by inductively coupled plasma emission spectrometry. Analytical error was estimated to be less than 2% (Table 1).
TDS, total dissolved solids.
3.2. Microscopy
3.2.1. Optical microscopy
Light (bright field and differential interference contrast) microscopy was carried out with a Zeiss Axio Imager Z1 on samples of the deposits returned from the field site in 2009 and maintained at 4°C. The samples for optical microscopy were diluted 1:10 in deionized H2O to increase dispersion. An individual sample from a deep area of the deposits (BF09_06) and a sample pooled from all sulfur deposits on the glacier (BF09_pooled) were examined and compared at varying magnifications. A total of 28 images from the two samples were collected from an estimated 60 fields of view.
3.2.2. Environmental scanning electron microscopy (ESEM)
Fully hydrated samples were examined with an FEI (Philips) XL30 environmental scanning electron microscope equipped with a field emission electron gun. The samples from two different locations on the glacier (BF09_05 and BF09_06) were mounted onto carbon tape and directly onto the stage without preparative treatment. A total of 51 images were collected from an estimated 100 fields of view.
3.2.3. Scanning electron microscopy (SEM)
Samples were fixed with glutaraldehyde and dehydrated by immersion in ethanol of 50%, 70%, 75%, 90%, 95%, and 100% concentration for 30 min at each concentration. The samples were then applied to carbon filter paper, and a 7 nm gold-palladium coating was applied to the samples to reduce charging during imaging. Dehydrated samples from BF09_pooled were examined with JSM-6480LV (low vacuum) and JSM-7401F (field emission) microscopes. A total of 20 images were collected from an estimated 50 fields of view. An energy dispersive X-ray spectrometer (EDS) installed on the field emission scanning electron microscope was used to conduct elemental analyses of mineral and organic structures present within the samples.
3.2.4. Scanning transmission electron microscopy (STEM)
To examine relationships among minerals and microbes at the nanometer scale, thin sections of the samples were prepared. This was done by suspending ∼0.1 g of the samples in narrow cylinders of agar, which were subsequently stained with OsO4 and uranyl acetate for 1 h each and then dehydrated as above. Uranyl acetate and osmic acid provide heavy metals to the sample for later visual contrast during STEM imaging. The samples were then immersed in 50:50 ethanol:acetone and then 100% acetone for 15 min each. They were then covered with a solution of 50:50 acetone:Epon-812 embedding resin overnight in a fume hood with the lids off to allow infiltration of the resin into the sample during acetone evaporation. The next morning, the agar cylinders were transferred to embedding moulds, covered with 100% Epon resin containing polymerization catalyst, and baked for 48 h at 60°C in a vented oven. The resulting blocks were used to cut 60 nm thin sections on an ultramicrotome with a diamond knife. The thin sections were picked up on Formvar-carbon coated 200 mesh copper grids and examined (Werlin et al., 2011). A total of 47 images were collected from an estimated 150 fields of view.
3.3. X-ray diffraction (XRD) spectroscopy
Investigations of the bulk mineral phases present in field materials from 2009 were carried out by powder XRD. Samples of BF09_06 and BF09_pooled, which had been stored at −80°C since their return from the field, and those that had been stored at 4°C were investigated. Laboratory sulfur (orthorhombic S8) was also analyzed to provide a standard for comparison.
Samples were allowed to air-dry and then ground in an agate mortar and pestle, and ∼1 g of each of the powdered samples BF09_06 and BF09_pooled was fixed on ground quartz plates with acetone. The powders were analyzed in a Scintag XDS 2000 X-ray diffractometer by using Cu-Kα radiation. Minerals were identified on the basis of their dominant peaks and referenced to the Mineral Powder Diffraction File (JCPSDS, 1980).
3.4. Infrared and Raman spectroscopy
Fourier transform infrared (FTIR) spectroscopy was carried out with material returned from the field in 2009, which had been stored in a −80°C freezer to prevent degradation, in order to look specifically for organics, including materials associated with cellular and extracellular components. A 25 mg sample was dried under nitrogen at 25°C (∼1 h) and mixed with IR-grade potassium bromide (Aldrich). The infrared spectra were acquired by using a diffuse reflectance attachment (Pike Technologies) with a Bio-Rad FTS 6000 FTIR spectrometer. The sample was then extracted with dichloromethane, and the extract was deposited onto KBr to concentrate the organic components for further diffuse reflectance analysis.
Raman spectroscopy was carried out for this work with use of a Bruker Senterra system equipped with a 785 nm excitation laser with 9 nm resolution. The sample was placed on a stainless steel mirror and air-dried under dry nitrogen at 25°C. The microprobe beam was focused on the surface with approximately 2 min acquisition time.
4. Results
4.1. Aqueous geochemistry
Measured temperature of the spring waters was −0.2°C, and pH was 7.8. High levels of sulfide were measured at the spring outlet at 152.35 mg/L, the highest recorded for any sulfur spring in Canada. Table 1 contains the major ion geochemistry for spring waters. pH within the deposits varied between 6 and 7. The total dissolved solids of 5900 mg/L is significantly higher than previously reported by Grasby et al. (2003). This probably reflects less dilution by glacial melt water for the 2009 samples; therefore, these results provide a more accurate spring water composition.
4.2. Microscopy
Initial optical microscopic evaluation of the samples returned from the field (Fig. 2) revealed spheres ranging in size from 1 to 20 μm and assorted mineral grains in a similar size range. Semi-opaque material was observed between and around mineral constituents. Elongated crystals and materials whose morphologies were consistent with those of microbial sheaths produced by Borup cultivars (e.g., Gleeson et al., 2011) were also observed.

Optical microscopy images of the structure of material in the deposits. (
Higher-resolution ESEM investigations of wet unaltered samples revealed a mineral matrix dominated by spherical mineral aggregates and rhombic crystals frequently enveloped in EPS material (Fig. 3). Figure 3a provides a representative wide-angle view of one of the samples, which demonstrates the ubiquitous distribution of spherical and rhombic morphologies. Smaller needle-shaped crystals were also observed (e.g., Fig. 3c), as well as partially dissolved sulfur aggregates (Fig. 3c).

ESEM images of unaltered samples. (
Figure 4 shows evidence of microbial presence within the samples. These include an example of a eukaryote in Fig. 4a, which was identified as a pinnate diatom on the basis of parallel lineations visible on the frustule at higher magnification. Clusters of microbial cells enveloped in EPS are shown in Fig. 4b, with arrows indicating points of beam damage sustained by the organic material during the course of our analyses. An example of a possible microbial sheath is shown in Fig. 4c, including a view of the open-ended cylindrical structure. Figure 4d shows a combination of both cellular and mineral materials enveloped in EPS, with at least one cell captured in the process of subdividing (indicated by the shortest arrow). More angular grains could be reliably identified as mineral rather than cellular material, but mineralization of organic constituents complicated identifications as microbial materials became progressively obscured. EPS was observed to be variable in distribution but common within the samples, and unmineralized cells and sheath-like materials were observed less frequently.
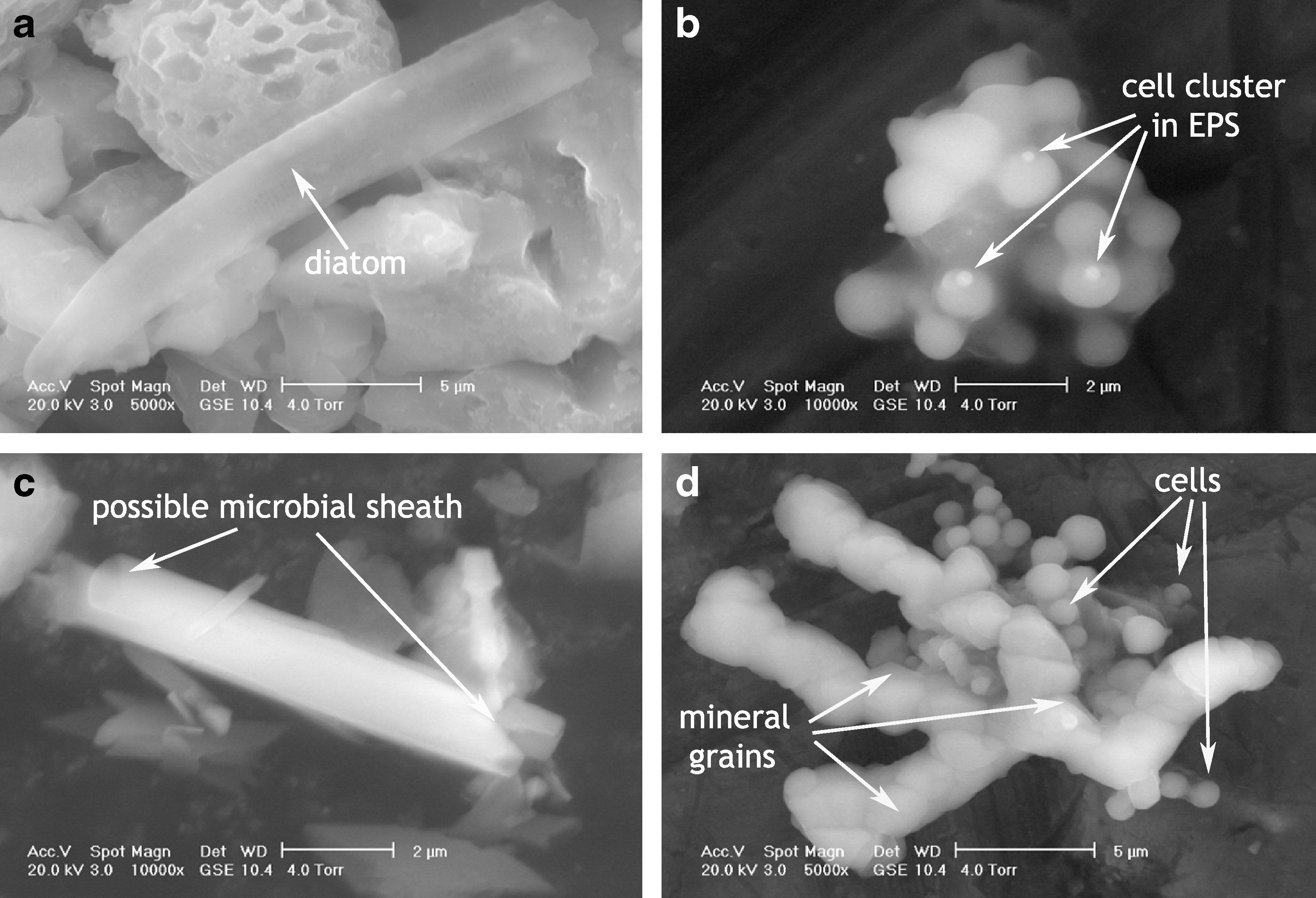
ESEM images of biological constituents encountered within the deposits. (
Scanning electron microscopy imaging and energy dispersive X-ray spectroscopy of dehydrated samples provided additional detail on the morphologies and composition of mineral constituents within the deposits (Figs. 5, 6, and 7). The rough aggregate texture of the large spheres of sulfur is clearly visible in Fig. 5a. Associated mineral grains are identified as mainly sulfur, present both as rhombohedral grains of α-sulfur and characteristically prismatic or tabular crystals of γ-sulfur, the mineral known as rosickyite. Rosickyite was distinguished from sulfate on the basis of Ca content. Calcium and sulfur were observed in point spectra shown in Fig. 7, which correspond to sulfate minerals. Additional grains rich in Si and Ca were observed in the samples (Fig. 6) and likely represent silica and carbonate minerals, which were observed to comprise a minor constituent of the mineral deposits. Calcite grains were distinguished from gypsum on the basis of sulfur content. Gold and palladium are present in the EDS spectra, as these are used to coat materials to prevent charging during imaging. Carbon is present in all spectra, as samples were loaded on carbon tape.

SEM images of dehydrated samples. (

EDS maps of carbon, sulfur, and silicon showing the composition of mineral aggregates and other grains from Fig. 5a. Background carbon tape on which samples are mounted dominates the carbon maps except where sulfur (S), silicate (Si), or carbonate (Ca) grains are present. (A Ca map was not acquired for this image, but a point spectrum confirmed the grain in the center right of the image had a large calcium peak and no sulfur peak). Color images available online at
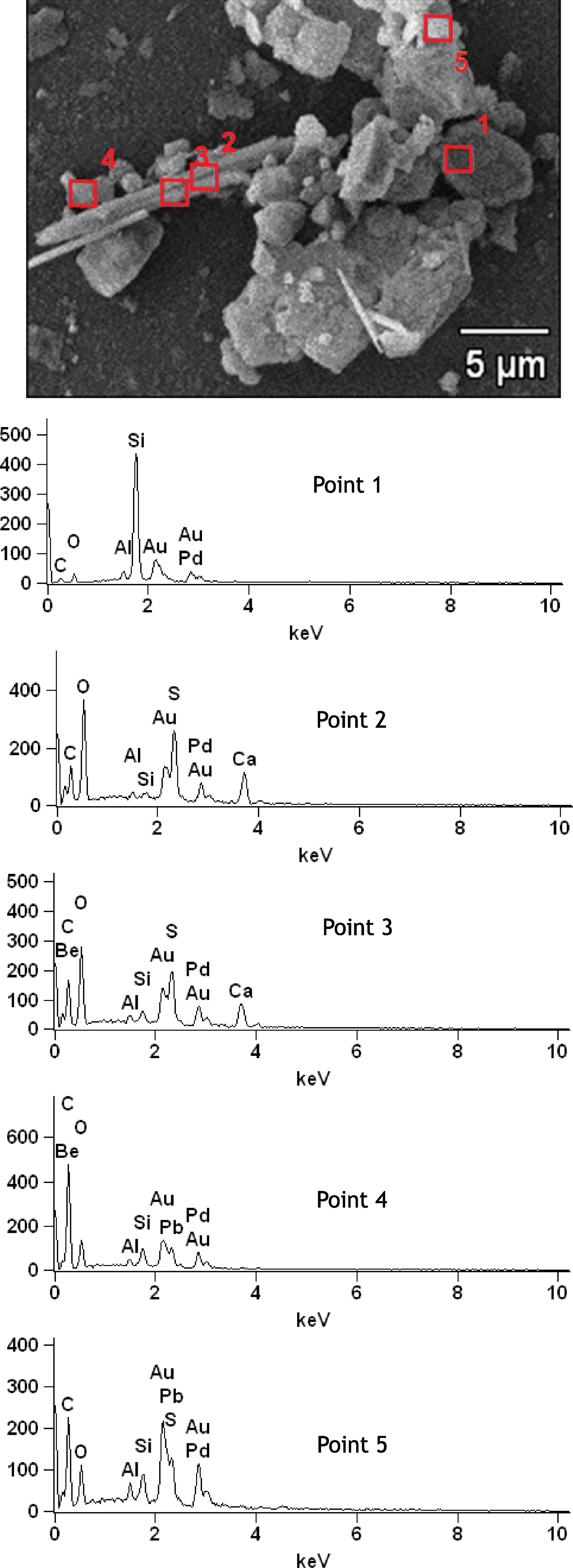
EDS point spectra showing the composition of materials from locations 1–5 shown above. Point 1 is an Si-rich grain of silicate mineral. Points 2 and 3 are rich in S, O, and Ca corresponding to sulfate minerals. Point 4 may represent C-rich EPS or cellular material although background C is present in all spectra from carbon mounting tape. Point 5 contains S-rich mineral aggregate. Color images available online at
The use of STEM imaging of resin-fixed thin sections of samples provided a cross-section view into the interior of mineral and organic constituents of the deposits. In this technique, mineral components appear bright, while stained organic components appear darker. Black areas are empty voids in the thin section where larger, heavier grains fell out of the resin matrix. Microbial cells, sheath-like materials, and EPS are surrounded by brighter mineral components in the form of submicron needle-shaped mineral grains (Fig. 8).

STEM images of fixed samples. (
4.3. X-ray diffraction
Figure 9a shows an XRD scan from 5° to 70° of a sample of material collected from the deposits, which had been maintained at 4°C since its collection (although the technique required the sample to be dried out at room temperature prior to analysis). The results of the scan were compared with mineral cards from the Mineral Powder Diffraction File (JCPSDS, 1980) for sulfur (both orthorhombic α-sulfur and monoclinic γ-sulfur), gypsum, and calcite, which were the materials detected in investigations carried out in previous years (Grasby et al., 2003; Gleeson et al., 2010). Figure 9a reveals α-sulfur as the dominant crystalline constituent of the deposits, which accounts for 39 of the 40 labeled peaks. Comparison of this spectrum with a laboratory sulfur standard reveals a few differences, principally the presence of a peak at 20.8°, which is not accounted for by α-sulfur. Gypsum has a major peak at 20.8°, and minor amounts of gypsum in the sample could be contributing to this spectrum.
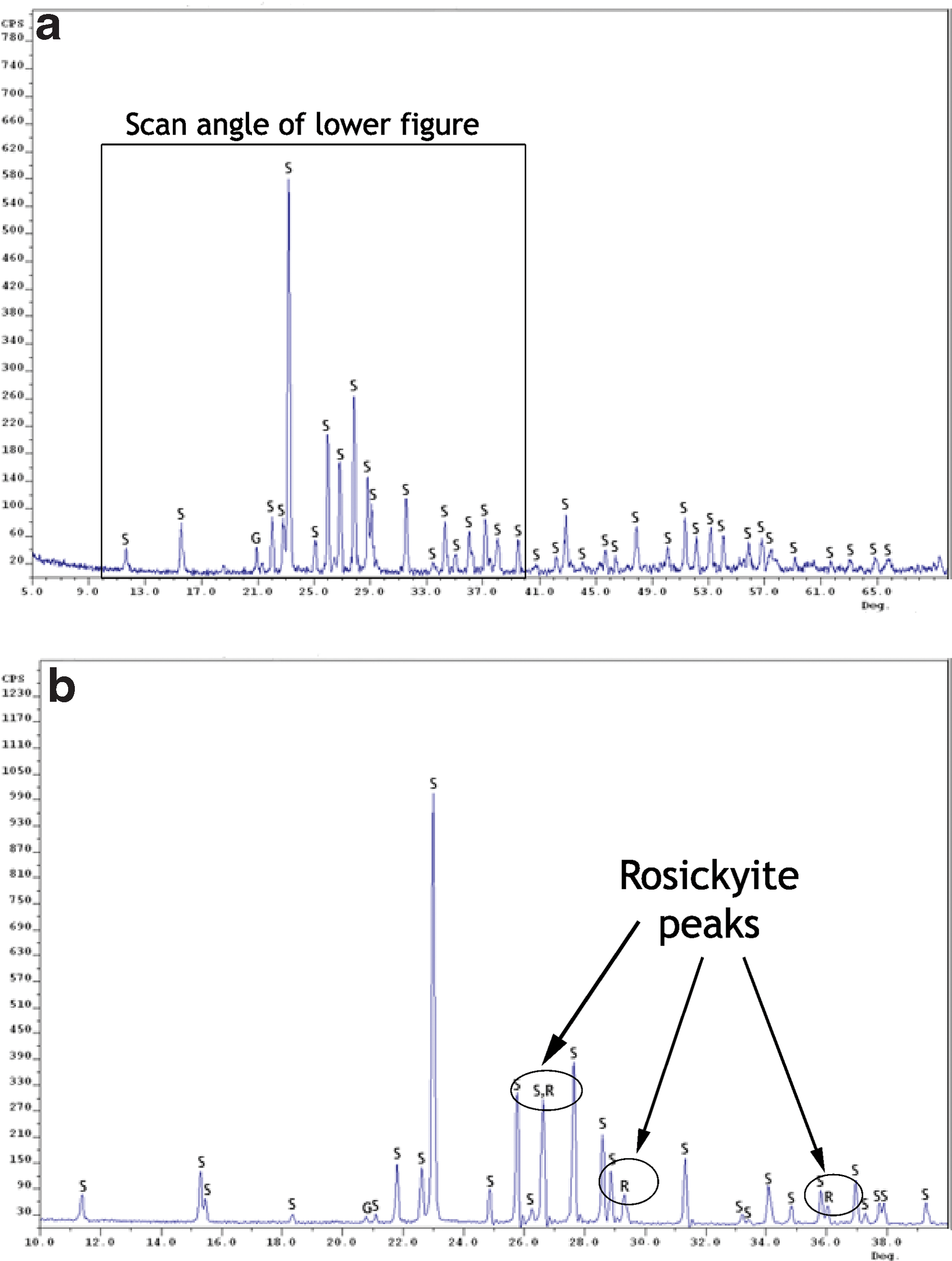
X-ray diffraction patterns from sample BF09_06 of material from the deposits. (
Figure 9b shows a spectrum of material from the same sample site, which had been stored at −80°C since its return from the field. For this sample, the scan time was increased to improve the signal-to-noise ratio (SNR), and the scan angle was reduced to an area between 10° and 40°, which included the location of the major rosickyite peaks. Although the sample is similarly dominated by the presence of α-sulfur (accounting for 26 of 29 labeled peaks), some evidence exists for the presence of rosickyite in the sample. The peak close to 27° has a higher intensity in comparison to that of the α-sulfur mineral card, while peaks are present at 29.5° and 36° that cannot be accounted for by α-sulfur and are not recorded in the sample stored at 4°C. Rosickyite has diagnostic peaks at 29.5° and 36°, and may be contributing to the spectrum in this sample maintained at −80°C. The potential gypsum peak is considerably lower than that of the sample stored at 4°C. Similar results were obtained in spectra collected of material pooled from multiple sites within the sulfur deposits.
4.4. Spectroscopy
Fourier transform infrared spectroscopy is a rapid laboratory measurement technique applicable to the study of biomolecules (Kacuráková and Wilson, 2001). FTIR spectroscopy can additionally provide quantitative information about the analyte (Fuller and Griffiths, 2002). Figure 10 shows the diffuse reflectance infrared spectra of the bulk sample and extracted organic components. Although C–H stretch peaks are visible in the bulk spectrum from 3000 to 2800 cm−1 (3.3–3.6 μm), the mineral component is obscuring the organic signal. Silicate and carbonate contributions dominate the spectrum. Extraction with dichloromethane concentrated organic components; thus aromatic carbon is revealed at 3050 cm−1 (3.28 μm) and carbonyl peaks from 1800 to 1600 cm−1 (5.5–6.3 μm), although mineral constituents are still detected.

(
The diffuse reflectance of the samples is shown in reflectance units in Fig. 10. When these data are converted to Kubelka-Munk units (Fuller and Griffiths, 2002), the amount of hydrocarbon functional groups can be estimated. For the organics measured directly in the sample, the estimated content was ∼5 μg in 25 mg. For the dichloromethane extract, this was ∼1 μg hydrocarbon extracted from 76 mg of sample.
In application to solids, Raman scattering provides similar but complementary information to the infrared absorption spectrum, the energies of molecular normal modes of vibration. Although SNR of Raman measurements are generally lower than those of FTIR, they can be carried out on aqueous or hydrated systems, and the fingerprints of complex biomolecules can have greater spectral simplicity in Raman spectra than do the overlapping bands of infrared absorptions (Thomas, 1999). Figure 11 shows the Raman spectrum of the bulk sample, which was dominated by the spectrum of sulfur, with peaks at 471, 219, and 153 cm−1 (21.2, 45.6, and 65.3 μm). The peak at 275 cm−1 was not matched by sulfur and may relate to carbonate or silicate components, as this area does not fall inside the fingerprint region of organic molecules (500–2000 cm−1). Organic content, as estimated by infrared spectroscopy (5 μg in 25 mg=100 ppm), was below the detection limit for this method, which typically ranges from 100 to 1000 ppm (Mann et al., 1987).
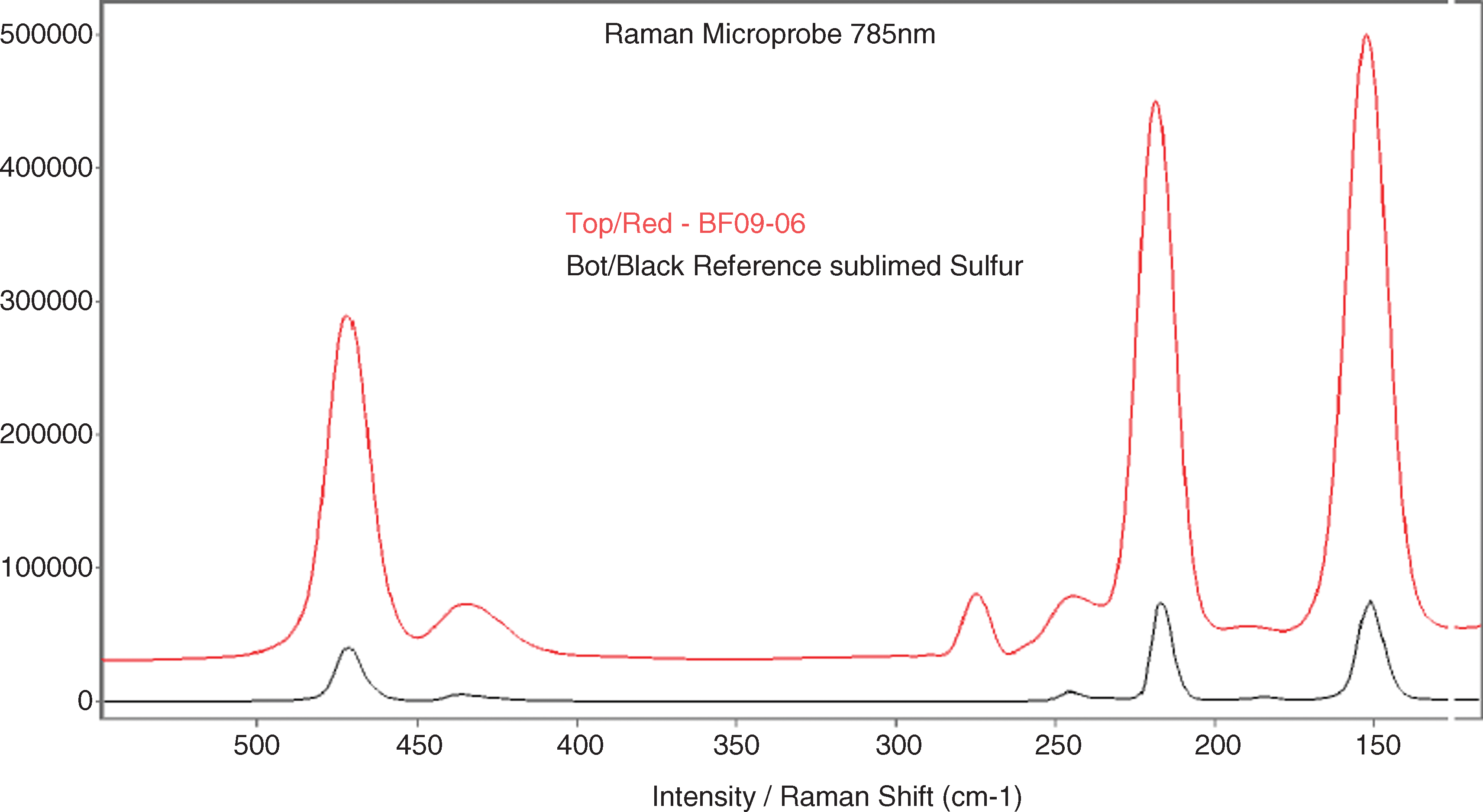
Raman spectrum of the bulk sample. This technique did not detect measurable quantities of organics. Sulfur dominated the subset of the spectrum shown above, with peaks at 471, 219, and 153 cm−1 (21.2, 45.6, and 65.3 μm). The peak at 275 cm−1 likely relates to carbonate or silicate components. Color images available online at
5. Discussion
The data collected from optical and electron microscopy, XRD, and FTIR and Raman spectroscopy, respectively, are discussed here in the context of what is currently known about the characterization and detection of morphological, mineralogical, and organic biosignatures.
5.1. Morphological biosignatures
The use of morphologies of cellular and extracellular components has been proposed as one means by which to focus the search for evidence of past life on Mars, although unambiguous identification of biosignatures in any context requires substantiating evidence from a multitude of techniques (Pullan et al., 2008). Cady et al. (2003) stated that morphological evidence is most useful as a precursor to chemical or structural analyses that can distinguish biogenic from nonbiogenic attributes. The direct morphological evidence for microbial presence within the icy sulfur deposits at Borup Fiord Pass included microbial cells and extracellular components such as sheath-like materials and EPS. Extracellular sheath materials can increase the rates of mineral precipitation by providing organic templates for crystal growth or attachment surfaces for colloidal aggregates (Banfield et al., 2000). We have also observed that both cells and extracellular components may act as nucleation surfaces for sulfur mineralization.
The larger spheroidal mineral aggregates observed both in Borup Fiord cultures and presented here in field samples show morphological similarities with framboids observed in pyrite (FeS2), which is generated in iron-sulfur microbial systems (Popa et al., 2004); biogenic spheroidal aggregates of zinc sulfide (ZnS) produced by sulfate-reducing bacteria (Labrenz et al., 2000; Moreau et al., 2004); and selinium nanospheres produced by Se-respiring bacteria (Oremland et al., 2004). The spheroids in these cases are typically coated by organic polymers and associated with microbial cellular surfaces (Moreau et al., 2004). The sulfur spheres observed in our samples were frequently enveloped in EPS (Fig. 4), which caused aggregation of multiple spheres by cohesion. This cohesive effect probably operates at multiple scales. Previous reports of sulfur spheroids are rare but not unknown. Dutrizac and Chen (1995) observed the formation of spheroids and subhedral crystals of elemental sulfur similar in size and morphology to those reported here during the course of galena (PbS) leaching experiments operating at 55–95°C. They concluded that dissolved H2S was undergoing oxidation to elemental sulfur and that this was occurring preferentially at active sites, such as pre-existing sulfur surfaces. Cellular surfaces could be providing additional active sites for sulfide oxidation in the Borup Fiord deposits.
5.2. Mineralogical biosignatures
The nature of sulfur produced by microorganisms has been the subject of considerable debate. To account for the hydrophilic nature of some of the sulfur generated by microorganisms, Steudel (1989) described the extracellular or intracellular storage product of sulfur bacteria as consisting of long-chain polythionates, which form globules of up to 1 μm diameter. Prange et al. (1999, 2002), Engel et al. (2007), and Franz et al. (2009) used X-ray absorption near-edge spectroscopy (XANES) to investigate the nature of sulfur globules produced by lithotrophic and phototrophic sulfur bacteria and found large species-dependent variations in terms of the production of one or more of the following: cyclooctasulfur, polymeric sulfur, polythionates, sulfur chains terminated by organics. Pasteris et al. (2001) used Raman spectroscopy to determine that sulfur harvested from sulfur-precipitating marine bacteria—including the large, filamentous Thioploca and Beggiatoa—was in the form of the common, stable, S8 ring configuration. Furthermore, they found this sulfur to be solid and microcrystalline, in contrast with previous reports of liquid sulfur (Hageage et al., 1970).
S8, also known as S0, is thermodynamically stable as cyclic S8 orthorhombic α-sulfur (Roy and Trudinger, 1970). Elemental sulfur converts to monoclinic β-sulfur at temperatures above 95.3°C and can be artificially prepared as monoclinic γ-sulfur (Meyer, 1976). Rosickyite (γ-sulfur) is a rare allotrope of sulfur and unstable under Earth surface conditions. The mineral has only been found in a few localities, typically as an oxidation product in pyrite-bearing environments rich in organic matter (Meisser et al., 2000). Its formation has been linked to the slow crystallization of sulfur in the presence of organic material (Anthony et al., 1990). Douglas and Yang (2002) and Douglas (2004) investigated the production and stabilization of γ-sulfur by sulfide-oxidizing microbes and proposed the presence of this metastable sulfur phase as a potential biosignature.
The abundant EPS that exists within the Borup sulfur deposits and the activities of microorganisms that are potentially oxidizing sulfide both point toward the necessary conditions for the formation of rosickyite at the field site. The link between biology and the presence of rosickyite may be directly related to the role of organic ligands provided by polysaccharides and lipids, and possibly polythionates such as detected by Steudel (1989). These act to stabilize liquid sulfur droplets or “globules” (Steudel, 1996), which allows time for the slow crystallization of rosickyite (Anthony et al., 1990). Such organic components would be amorphous to XRD analyses such as reported here.
The presence of rosickyite peaks in the XRD patterns of the Borup sulfur samples (Fig. 9b) are corroborated by (E)SEM and STEM observations (Figs. 3 –8), which show needle-shaped crystals of sulfur, the characteristic habit of this mineral. Although gypsum is also known to form needle-shaped crystals, the presence of soluble organic compounds alters this habit to a more lenticular morphology (Van Rosmalen et al., 1976; Cody, 1979). STEM observations in particular revealed small needle-shaped crystals nucleating directly on organic constituents. The dominance of α-sulfur peaks in XRD measurements in comparison to the limited evidence for rosickyite does not provide conclusive evidence of the biogenic or abiotic origin of the sulfur. The characteristics of biogenic sulfur described by Hageage et al. (1970) are such that biogenic sulfur globules are initially produced in a liquid (amorphous) form of S8, which transitions through unstable mineral phases before stabilizing as α-sulfur over a period of days. These measurements are based on wet samples of the globules extracted from cells. The inconsistencies in structural characterization of elemental sulfur in different studies of this material may relate to the different approaches taken to prepare the material for analysis, as well as the fact that recent XANES results suggest that phylogenetically and metabolically different bacteria may produce and accumulate different forms of sulfur (Prange et al., 2002).
Our sample collection and preparation protocols may not have fully preserved rosickyite within the Borup deposits, and it is also possible that rosickyite is continuously converted to the considerable quantities of orthorhombic sulfur that dominate the deposits, such as the mineral aggregates and rhombohedral grains characterized by SEM. Although it is tempting to infer that the material stored at 4°C became more oxidized than the material stored at −80°C, as suggested by the higher gypsum peaks within the sample stored at the warmer temperature, and that rosickyite stabilized as α-sulfur at the higher temperature, heterogeneity in the samples before storage may have been high.
5.3. Organic biosignatures
The extraction of organics from the bulk sample greatly increased SNR in the spectrum shown in Fig. 10b over that of Fig. 10a. This process did not extract the full content of organic material from the sample as quantified by FTIR spectroscopy, although this is not unexpected in mineral samples. A mineral component was still present in the spectrum of the extract. This may be due to the close association of organic and mineral components as characterized by microscopy, which can result in incomplete separation of organic and mineral components.
Proteins in general show amide I (O=C–N) at 1650 cm−1 (6 μm), and proteins containing tyrosine have strong absorptions at 1514 cm−1 (6.6 μm). The phosphodiester bonds of DNA, RNA, and ATP occur at 1240 and 1078 cm−1 (8.06 and 9.28 μm), but in our spectrum, this portion was obscured by mineral components. Amides, esters, and organic acids could represent components of proteins, lipids, and polysaccharides, which have been shown to comprise the bulk of extracellular sheath materials in cyanobacteria (Tease and Walker, 1987) and in iron- and manganese-oxidizing bacteria (Emerson and Ghiorse, 1993).
Although Raman spectroscopy has been successfully applied to biosignature investigations of fossilized sedimentary structures (Edwards et al., 2007) and communities relatively low in biomass content such as those of endoliths in Antarctica (Wynn-Williams et al., 1999), the detection limit for organics with this method of ∼100 to 1000 ppm (Mann et al., 1987) proved too high for the Borup Fiord samples, and alternative methods to concentrate organics would be required to increase SNR to a detectable level.
6. Conclusions
We found several overlapping lines of evidence for microbial activity within the icy, sulfur-rich supraglacial deposits at Borup Fiord Pass. The potential links between microbial organisms and elemental sulfur production include observations of microbial cells and sheath-like structures throughout the deposits, as well as the visualization of cellular materials mineralized with elemental sulfur; the presence of the potentially biogenic γ-sulfur phase rosickyite; and the presence of parts-per-million levels of organics diagnostic of cellular components. Both SEM and XRD investigations provided evidence for the presence of rosickyite within the deposits, especially in those samples maintained at −80°C since their return from the field, although the unstable nature of this phase makes measurement challenging. Infrared spectroscopy revealed the presence of functional groups diagnostic of proteins, lipids, and polysaccharides potentially associated with the microbial organisms and associated organic structures observed by microscopy. While DNA from the sulfur deposits can be successfully extracted and amplified as a short-term sign of microbial presence (Gleeson et al., 2011), biomineralized structures formed around cells and microbial sheaths have the potential to act as long-term indicators of microbiological activity, in association with the organic components, which may have been involved in promoting their formation. Benison et al. (2008) described clumps of organic bodies and sulfate crystals in modern acid lakes and analogs of these materials that were found preserved in Permian halite. The potential for preservation of the Borup Fiord microbial structures and biominerals is currently unknown and is worthy of further investigation. Regardless, the case for sulfur minerals as astrobiological targets is strengthened by their potential to host multiple types of biosignatures as elucidated by this work.
The evidence for sulfur minerals on Europa, coupled with sulfur's redox versatility and the suite of oxidative, reductive, and disproportionating metabolisms known on Earth, allows for the possibility that microbial sulfur cycling could provide a viable metabolic avenue for potential life there. As an intermediate of sulfur cycling, S0 has the potential to contain preserved morphological, mineralogical, and organic biosignatures, and as such provides a valuable target for astrobiology investigations on Europa. Analogous to the way in which potential stromatolites on Mars are frequently invoked as an appropriate starting place to search for small-scale microfabrics that would provide evidence of a biogenic origin (Farmer and Des Marais, 1999), the presence of large-scale, sulfur-rich deposits on Europa is an appropriate location to begin the search for small-scale morphological, mineralogical, and organic biosignatures. Only a combination of corroborating lines of evidence would have the potential to constitute a positive detection of life at Europa or at any location beyond Earth.
While abiotically produced organic molecules have been detected in the interstellar medium, comets, and meteorites (Ehrenfreund and Charnley, 2000), only small, relatively simple organic molecules have been detected in these environments. The detection of larger, more complex organic molecules in the deposits on the surface of Europa would suggest that they were formed on Europa itself, either within the deposits or below the surface, and that they were the result of relatively recent activity, as such molecules would be quickly degraded by the high radiation on Europa's surface. This radiation environment necessitates a sampling depth of greater than tens of centimeters for materials at the near surface of the satellite so as to reach below the radiation processing depth (Cooper et al., 2001). Although it would not constitute firm proof of life, the presence of large complex organic molecules (similar to those we have detected within the Borup sulfur deposits) on Europa would be a strong signal that life was potentially present, as these types of molecules are almost exclusively associated with biological processes on Earth. Low temperatures and lack of liquid water at the near surface could increase the preservation of organic or metastable structural components within Europa's non-ice materials.
Future astrobiological missions to the surface of Europa should carry a microscopic imager with the ability to spatially resolve mineral and organic morphologies on the order of hundreds of nanometers. A suite of instruments capable of fully characterizing potentially unstable organic and mineral components across scales relevant to the imager would be the most effective means of determining whether biosignatures that would provide evidence of life exist on the surface of Europa. These might include XRD and X-ray fluorescence for mineralogy and complementary chemical composition, and an infrared spectrometer to search for biologically produced organic molecules. While melting ice has been suggested as a means of concentrating material, and will likely be required for the analysis of organic materials by techniques that include Raman spectroscopy, these measurements should be carried out in concert with direct analysis of pristine samples, which could contain unstable materials.
Footnotes
Acknowledgments
We acknowledge the assistance of Benoît Beauchamp at the Arctic Institute of North America, Calgary, Canada, and Kennda Lynch at the Colorado School of Mines. Portions of this work were carried out at the Jet Propulsion Laboratory, California Institute of Technology, under contract to NASA, and were also funded in part by the NASA Astrobiology Institute Director's Discretionary Fund. We also acknowledge financial support from the Lewis and Clark Fund of the American Philosophical Society, the David and Lucile Packard Foundation, the Caltech postdoctoral program, and the Jupiter Europa Orbiter Project. We also wish to thank our anonymous reviewers, with whose help this manuscript has been considerably streamlined and improved overall.
Abbreviations
EDS, energy dispersive X-ray spectrometer; EPS, extracellular polymeric substances; ESEM, environmental scanning electron microscopy; FTIR, Fourier transform infrared; SEM, scanning electron microscopy; SNR, signal-to-noise ratio; STEM, scanning transmission electron microscopy; XANES, X-ray absorption near-edge spectroscopy; XRD, X-ray diffraction.