
Abstract
Icelandic streams with mean annual temperatures of less than 5°C, which receive the cationic products of basaltic rock weathering, were found to host mats of iron-cycling microorganisms. We investigated two representative sites. Iron-oxidizing Gallionella and iron-reducing Geobacter species were present. The mats host a high bacterial diversity as determined by culture-independent methods. β-Proteobacteria, Actinobacteria, α-Proteobacteria, and Bacteroidetes were abundant microbial taxa. The mat contained a high number of phototroph sequences. The carbon compounds in the mat displayed broad G and D bands with Raman spectroscopy. This signature becomes incorporated into the weathered oxidized surface layer of the basaltic rocks and was observed on rocks that no longer host mats. The presence of iron-oxidizing taxa in the stream microbial mats, and the lack of them in previously studied volcanic rocks in Iceland that have intermittently been exposed to surface water flows, can be explained by the kinetic limitations to the extraction of reduced iron from rocks. This type of ecosystem illustrates key factors that control the distribution of chemolithotrophs in cold volcanic environments. The data show that one promising sample type for which the hypothesis of the existence of past life on Mars can be tested is the surface of volcanic rocks that, previously, were situated within channels carved by flowing water. Our results also show that the carbonaceous signatures of life, if life had occurred, could be found in or on these rocks. Key Words: Bioenergetics—Chemolithotrophic microorganisms—Iron-oxidizing bacteria—Iron-reducing bacteria—Volcanism. Astrobiology 11, 679–694.
1. Introduction
I
Aerobic iron oxidation
Anaerobic iron oxidation
Phototrophic iron oxidation
Iron-oxidizing bacteria have been described in a number of environments around the world; most of these are deep-sea, temperate, or tropical environments (Emerson and Revsbech, 1994a, 1994b; Straub et al., 2000; Emerson and Moyer, 2002; K.J. Edwards et al., 2003, 2004; Sobolev and Roden, 2004; Haaijer et al., 2008; Blöthe and Roden, 2009; Duckworth et al., 2009; Wang et al., 2009; Bruun et al., 2010). None have yet been described in cold land-based volcanic environments.
The study of the distribution of iron-oxidizing bacteria is of interest from two perspectives. First, we know very little about the factors that control the distribution of different metabolic groups of chemolithotrophs in terrestrial environments and particularly the limitations to energy acquisition that might influence their distribution. By investigating the habitats of iron-oxidizing microorganisms, we advance our knowledge of the physical and chemical limits to the chemosynthetic biosphere on Earth. Second, establishing the habitats of chemolithotrophs is of importance for assessing the habitability of volcanic, iron-rich, extraterrestrial environments, particularly on Mars, and for identifying promising locations within which to test whether life exists or existed there. The possibility of detecting volcanism on extrasolar planets (Kaltenegger et al., 2010) might even lead to the ability to assess these planets as places with volcanically generated redox couples and, thus, as abodes for their associated life, including metal-cycling microorganisms. To make these assessments would require that we link the temperature ranges and other physiological limits of microorganisms associated with volcanic environments on Earth to the inferred conditions on extrasolar planets, accepting the conservative interpretation of the potential of extraterrestrial life based on what we know about the limits of terrestrial life.
Iron-oxidizing microorganisms are of special interest in the context of life detection because the biological oxidation of reduced iron results in iron-oxide minerals that often have distinct biogenic properties, which are exhibited in, for example, their morphology or isotopic composition. Perhaps best characterized in this regard are iron-oxidizers of the genera Gallionella (phylum β-Proteobacteria) (e.g., Hallbeck and Pedersen, 1995; Verran et al., 1995; Hallberg and Ferris, 2004) and Mariprofundus (phylum ζ-Proteobacteria) (Emerson et al., 2007) that produce distinctive stalks. These structures are not only easily observed microscopically, but they can be preserved in the fossil record and, thus, result in long-lived records of chemolithotrophic life (e.g., Schelble et al., 2004; Hofmann et al., 2008). Other examples include the tube-like iron-oxide sheaths produced by members of the genus Leptothrix (Emerson and Ghiorse, 1992).
In the present study, we investigated iron-oxidizing bacteria-containing microbial mats associated with streams in cold basaltic environments in Iceland, and we discuss here the applications of this work to the search for life on Mars.
2. Methods
2.1. Sample sites
Fieldwork was conducted in Iceland (Fig. 1). Some small streams in Iceland display a bright orange-red color (Fig. 2), which can be attributed to iron oxidation. Where iron oxidation is observed, the stream types generally fall into three categories: (1) streams in watersheds or catchments, (2) cooled water from volcanic hydrothermal systems that collects into streams, (3) streams emanating from the subsurface, either from the water table, melting ice, or snowpack (Fig. 2A).
Map of Iceland showing location of iron-oxidizing mats studied at Sites 1 and 2.

Iron-oxidizing mats. (
Two streams of type (1) in the south of Iceland were sampled in June 2008, 2009, and 2010 to investigate the hypothesis that they are sites of biological iron oxidation and cycling. The locations of the two streams are 64°29.334′N, 21°31.191′W (Site 1) and 63°46.253′N, 18°10.453′W (Site 2) (Fig. 1). The stream beds are covered by a microbial mat of approximate thickness 3–5 mm, which grows on the surface of rocks and sediment (Fig. 2A, B) and exhibits a striated, corrugated texture (Fig. 2C). At Site 2, the stream contains luxuriant iron-oxidizing mats (Fig. 2D). Site 1 is a stream at the base of a hill composed of basaltic rock with intermittent vegetation cover of primarily moss and lichen. The water source is surficial runoff from the basaltic rocks into the stream bed. Site 2 is a stream that runs perpendicular to a gorge across a vegetated hill. The stream traces back to a spring outlet that has a subsurface source and runs year-round. Either the water is from a subsurface aquifer or it collects water draining from the surrounding region.
2.2. Temperature logging, oxygen and pH measurements
Temperatures in the streams were measured over the course of one year from 15 June 2009 to 27 June 2010 and from 4 June 2007 to 4 June 2008 at Sites 1 and 2, respectively. Temperatures were measured with a Hobo UA Pendant datalogger (Tempcon instrumentation, Arundel, UK), which was programmed to measure the temperature at 10 min intervals at Site 1 and 15 min intervals at Site 2. The loggers were submerged and fixed to the bottom of the streams within the iron-oxidizing microbial mats. At Site 1, temperatures were also measured on the surface of unobstructed exposed rocks near the stream by using a U12-008 datalogger (Tempcon instrumentation, Arundel, UK) that was linked to a TMC6 HB external thermister.
Single-point oxygen measurements were made at Site 1 and 2 with a handheld calibrated Extech Instruments Model 407510 O2 meter (Extech Instruments, Breda, Netherlands) on 27 June 2010. Measurements were made at the surface of the microbial mat and at 0.5 cm depth underneath the microbial mat. Measurements were made in at least four locations along the stream.
Water samples were collected and returned to the laboratory on 27 June 2010. The pH of the stream was measured with a calibrated Hydrus 300 pH meter (Fisher Scientific, Loughborough, UK). Measurements were repeated on three separate water samples from each site.
2.3. Water analysis
Water was collected from both sites on 27 June 2010 and examined for major cations, anions, and trace elements. From each site, 300 mL of water was collected by placing an acid-washed plastic bottle within the stream water that flows over the mats, while avoiding the collection of any biomass. Water was maintained at ambient temperatures. Water for cation analysis was acidified with nitric acid in the field. Analysis was undertaken by Anglian Water (Huntingdon, UK) with the use of standard procedures. Cations were measured by using inductively coupled plasma mass spectrometry (Elan 6100, PerkinElmer, Cambridge, UK). Ammonium, chloride, alkalinity, and nitrogen compounds were measured on an Aqua900 Autoanalyzer (Seal Analytical, Fareham, UK) and fluoride on a Burkard Autoanlyzer (Burkard Scientific, Uxbridge, UK). Total iron and reduced iron were also measured in the stream water by using the ferrozine assay according to the method of Viollier et al. (2000). Iron concentrations were determined with a Biochrom Biowave spectrometer (Biochrom, Cambridge, UK) in the field. Samples were examined within 5 min after collection from the stream.
2.4. Bright field and fluorescent microscopy
Small segments of microbial mat separated from field-collected samples with a sterile blade were distributed onto a microscope slide, and doubly distilled H2O was added to suspend the material. The material was examined with bright field microscopy on a Leica DMRP fluorescence microscope (Leica Microsystems, Wetzlar, Germany). To visualize prokaryotes, samples were stained by adding a small quantity of SYBR Green I DNA binding dye according to the manufacturer's instructions (Invitrogen, Paisley, UK). Stained cells were observed fluorescing green by using an excitation wave band of 450–490 nm (Leica filter cube I3) and an emission long-band cutoff filter of >515 nm.
To enumerate prokaryotes, three separate samples of the microbial mat were collected in the field from each site and immediately fixed in 2% formaldehyde in stream water. On return to the laboratory, the material was shaken by hand to disperse the mat into a homogeneous mixture. Half a milliliter of this mixture was vacuum filtered onto black 0.2 μm Nuclepore polycarbonate filters (Whatman, Maidstone, UK) and stained with SYBR Green I as described above, and microorganisms were enumerated under at least 50 fields of view for each sample. The numbers of prokaryotes were estimated per gram of wet weight of microbial mat. The weight of the mat was determined by placing a known volume of the formaldehyde-fixed mat onto absorbent paper until most of the water was removed. The mat was separated from the paper and weighed.
2.5. Scanning electron microscopy
Samples of microbial mat were examined with a scanning electron microscope (SEM). Samples of microbial mat collected from Sites 1 and 2 and dried overnight in open dishes in the laboratory were fixed to aluminum stubs with double-sided carbon tape. Samples were carbon coated (15–20 nm thickness) and examined in secondary mode at 20 kV accelerating voltage and 7–15 mm working distance with a Quanta 3D dual beam FIBSEM (FEI, Oregon, USA). To obtain high-resolution images, a field emission gun (FEG) SEM (ZEISS Supra 55-VP, Zeiss Microimaging, Göttingen, Germany) was used that operated at 2–15 kV accelerating voltage and a 7–10 mm working distance. Three different samples from each site were examined.
Scanning electron microscopy was also used to investigate the surface of the basaltic rocks in the stream on which iron-oxidizing microbial mats grew. The mats were gently removed from rocks with a sterile blade in the field, and the rocks were air-dried and returned to the laboratory for analysis. Seven rocks from Site 1 and three rocks from Site 2 were prepared in this way and examined.
Two other sample types were examined with a SEM: (1) The surfaces of rocks obtained from a small oxbow-like deviation in the stream at Site 1. The rocks had not been subjected to stream water flow during the 2 years of our field studies, and the vegetation that grew around them suggested that the period of inactivity was considerably longer. We investigated these rocks to determine whether any evidence of iron-oxidizing bacteria could be observed on their surfaces. (2) The surface of the underside of the rocks on which the iron-oxidizing mats grew. We examined these samples as controls to compare to the surface of rocks on which the iron-oxidizing mats grew. Three different rocks from these two categories were examined.
2.6. X-ray diffraction
To investigate the minerals in the iron-oxidizing microbial mat, X-ray diffraction (XRD) was employed. Three ground samples of dried microbial mat from Site 1 were mixed together for analysis. Powder XRD patterns were determined with a Siemens D5000 X-ray diffractometer (Siemens, Munich, Germany) operating with a Cu X-ray tube (K-alpha average wavelength=1.5408 Å). Data were obtained in Bragg-Brentano mode on the samples attached to the surface of a silicon wafer coated with a thin layer of silicone grease. Measurements were made in steps of 0.02 degrees 2-theta from 5–80 degrees 2-theta with counting time of 12 s per step. X-ray diffractograms were compared to the 2002 release of the JCPDS release of the Powder Diffraction File by using the Bruker AXS software Diffrac.
2.7. Raman spectroscopy
Raman spectroscopy was used to investigate the biological and mineral components of samples at each site. Raman was carried out with a Horiba Jobin Yvon LabRAM (Horiba Jobin Yvon, Stanmore, UK). Samples were excited with a 473 nm laser (MPC 6000, Quantum Laser, Stockport, UK). To avoid sample damage, a low laser power of 0.7 mW at source was used. The footprint area of analysis was a 2 μm diameter spot. Spectra were the mean of five separate spectra, each acquired over 20 s to increase the signal-to-noise ratio. Results are accurate to ±1 cm−1 as determined by using an Si standard. Three different sample sets were examined, as follows: (1) Iron-oxidizing microbial mat. Samples of iron-oxidizing microbial mat were dried onto the surface of plastic Petri dishes for analysis. (2) The surface of rocks in an oxbow-like deviation in the stream that once hosted microbial mats (see SEM description above). (3) Areas of a thin section through the altered surface of a rock on which iron-oxidizing mat was growing. Twenty different spots on each of these materials were examined. Data was gathered in the program LabSpec.
2.8. Molecular biological analysis of microbial mats: DNA extraction
Site 1 was chosen for a molecular biological analysis of the microbial mat community. Samples were collected from two sources, as follows: (1) Community DNA was extracted from 10 g of rock with intact microbial mat on its surface (yield ∼18 μg/mL), according to the procedure previously described by Herrera and Cockell (2007). This sample was designated as Sample A. As the rock was non-vesiculated and fine-grained, the contribution of DNA from organisms that were in the rock interior was assumed to be minimal. (2) DNA was extracted from another sample of the microbial mat, scraped from the rock surface into a Petri dish. This sample was designated Sample B. DNA was extracted using a FastDNA Spin kit for soil (Qbiogene, Cambridge, UK), in accordance with the manufacturer's instructions.
2.9. Construction of bacterial 16S rRNA gene clone libraries
Bacterial 16S rRNA genes were amplified with primer sets pA-com2 (Bruce et al., 1992; Schwieger and Tebbe, 1998) and pA-pH (Bruce et al., 1992) for Samples A and B, respectively. For both primer sets, the 50 μL amplification reactions contained 0.2 μM each primer, 200 μM each dNTP (New England Biolabs, Herts, UK), 2.5U Taq DNA polymerase, and 1×PCR buffer [200 mM Tris-HCl (pH 8.4), 500 mM KCl] (Invitrogen, Paisley, UK). Reactions also contained MgCl2 at concentrations of 2.0 mM and 1.5 mM, and template DNA at concentrations of approximately 40 and 100 ng, for primer sets pA-pH and pA-com2, respectively. Each amplification was performed in duplicate in a G-Storm GS1 thermal cycler under the following cycling regimen: 95°C for 5 min, followed by 35 cycles of 95°C for 1 min, 55°C for 40 s, and 72°C for 40 s, with a final extension for 10 min at 72°C. Duplicate products were combined and either gel purified (Sample A) or purified directly (Sample B) (Illustra GFX, GE Healthcare, Buckinghamshrire, UK). Purified products were cloned immediately into pCR4-TOPO vector and transformed into chemically competent One Shot TOP10 E. coli (Invitrogen, Paisley, UK). Vector inserts were sequenced with primer pA (Mclab, California, USA).
2.10. Phylogenetic analysis
Seventy-four Sample A and 44 Sample B clones (total 118) were sequenced. To check for the presence of chimeras, sequences were submitted to Bellerophon (Huber et al., 2004) and subjected to signature-shift analysis (Wang and Wang, 1996). Clone sequences and their closest phylogenetic matches, as retrieved from the Ribosomal Database Project (release 10) with SeqMatch, were aligned with BioEdit (version 7.0.9) (Hall, 1999) by ClustalW (Thompson et al., 1994) and exported to Mega4 (Tamura et al., 2007) for further phylogenetic analysis. A bootstrapped (Felsenstein, 1985) neighbor-joining (Saitou and Nei, 1987) phylogenetic tree was constructed, with evolutionary distances determined with the Jukes-Cantor algorithm (Jukes et al., 1969).
2.11. GenBank accession numbers
Clone sequences were deposited in GenBank under accession numbers HM572425–HM572521.
3. Results
3.1. Temperature, oxygen, and pH measurements in streams
Temperature profiles of the streams at Sites 1 and 2 were obtained over 1 year (Fig. 3). At Site 1 the mean stream temperature over the measurement year was 4.05°C; the maximum temperature was 22.23°C at 15:01 on 3 August 2009. The mean temperature of a rock outcrop near the stream over the same time period was 4.18°C, but the temperature extremes were greater (Fig. 3B). The rock surface experienced a temperature of −11.25°C at 7:39 on 5 January 2010 and a temperature of 29.16°C at 13:39 on 6 July 2009. At Site 1, the mats first reached 0°C on 4 October 2009 (03:00) (when they are presumed to have frozen, as suggested by the low cation concentrations in the water, which would not significantly modify the freezing temperature) and last froze on 27 April 2010 (08:01) (a total period of 199 days). Between these times, the streams intermittently rose above 0°C, and these periods corresponded to warm periods also measured on the surface of the exposed rocks near the stream, which shows that they were periods of warm air temperatures, not independent changes in the temperature of the water source. During this period, temperatures in the stream did not exceed 7.4°C. During the summer, the mats experienced diurnal temperature variations. An example of such excursions is shown in Fig. 3C. The minimum temperature during these excursions only exceeded 10°C at Site 1 in July. Diurnal temperature excursions were often greater than 12°C. At Site 2, the mean stream temperature over a year was 4.50°C; the maximum temperature was 16.14°C at 13:16 on 12 July 2007. The winter profile of the stream at Site 2 was significantly different from that at Site 1 (Fig. 3D). The mats rarely experienced prolonged freezing.

Temperature measurements associated with iron-oxidizing mats. (
The oxygen concentrations at the surface of the iron-oxidizing microbial mats varied between 9.6 and 10.1 mg O2/L. Measurements at a depth of 0.5 cm within or underneath the microbial mats were <1.1 mg/L in all cases.
The mean pH of the stream water was slightly alkaline: 7.67 at Site 1 and 7.54 at Site 2 with less than 5% standard deviation in measurements.
3.2. Water composition
The concentration of the major analytes from stream water collected in both sites is shown in Table 1. The water had a low conductivity typical of freshwater rivers. Cation and anion concentrations were within ranges reported for cold Icelandic streams that contain the products of basaltic rock weathering (Gíslason et al., 1996; Stefánsson and Gíslason, 2001; Louvat et al., 2008). Consistent with previous data from basaltic watersheds (Stefánsson and Gíslason, 2001), the cation concentrations were in the order Na+ > Ca2+ ≥ Mg2+ > K+ and anion concentrations
3.3. Bright field and fluorescent microscopy
Bright field microscopy showed the presence of iron oxides with abundant iron-oxide stalks and tubes (Fig. 4A, 4B). The total number of prokaryotes enumerated by staining was between 1.3×107 and 1×108 cells/g wet weight of mat. Cells were non-homogeneously distributed and clustered around, and/or attached to, organic material within the mat, and they were embedded in iron oxides.
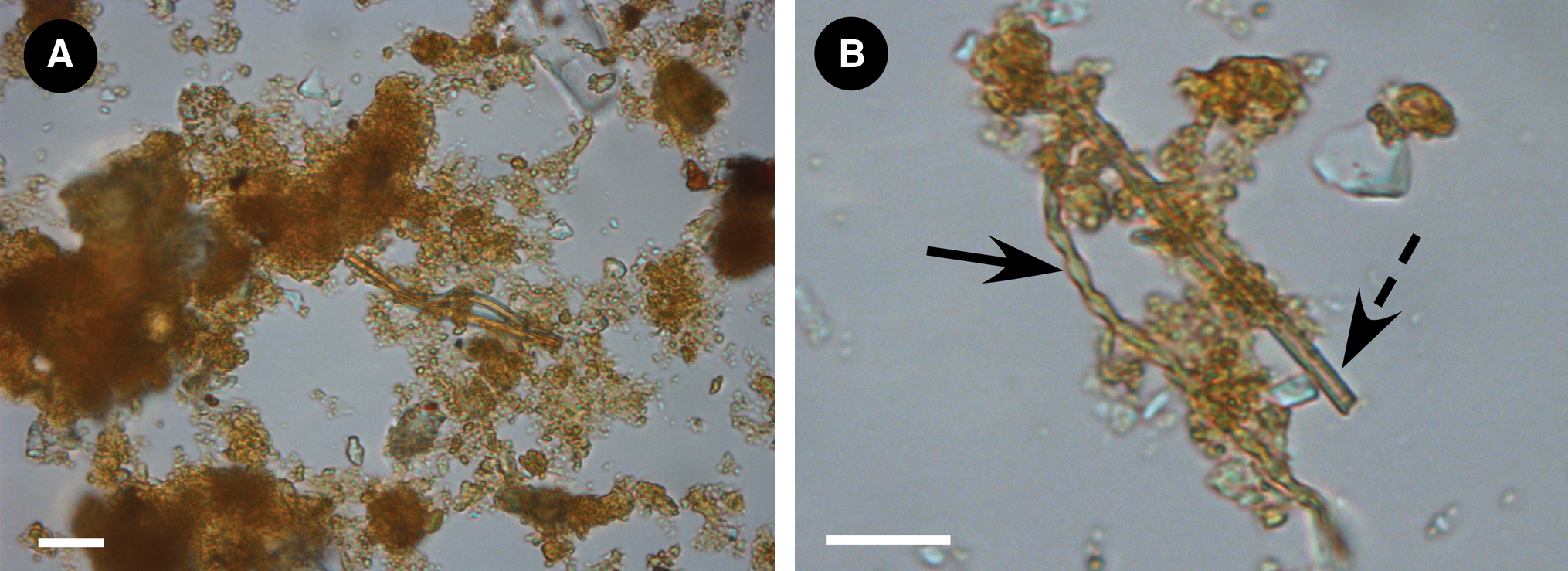
Bright field micrographs. (
3.4. Scanning electron microscopy
Scanning electron microscopy showed that the mat was comprised of a mass of amorphous iron oxides (Fig. 5A). Conspicuous within the mat were large numbers of ribbon-shaped iron-oxide spirals (Fig. 5A, 5B) characteristic of Gallionella spp. and tube-shaped iron-oxide sheath deposits characteristic of Leptothrix spp. (Fig. 5C). The presumptive Gallionella stalks had conspicuous spheroidal concretions on their surfaces (Fig. 5B). The mat hosted a large number of diatoms (Fig. 5D) that were often embedded within the iron-oxide deposits.

(
The surface of the basaltic rocks on which the iron-oxidizing bacteria grew was flat and fractured (Fig. 5E). Notable features were circular indentations or hollows in the rock surface (Fig. 5F). These features were not observed on the underside (control) surface of the rocks (example surface is shown in Fig. 5G). The rocks examined that were once part of the stream did not show hollows or indentations similar to the rocks on which active mats grew. Detritus was evident on the rock surface and appeared to be partially incorporated or strongly attached to the altered surface layer of the rock (Fig. 5H).
3.5. X-ray diffraction
The results of the XRD indicated the following precipitates and phases were present in the microbial mat: primarily amorphous iron oxides (FeOOH), feldspars, pyroxene with minor amounts of magnetite, wustite, goethite, and calcite.
3.6. Raman spectroscopy
Raman spectroscopy was performed on dried samples of the microbial mat. The microbial mat showed similar spectral features in different locations. An example is shown in Fig. 6A (Spectrum 1). The most prominent features are broad bands at ∼1350 and 1590 cm−1, which most likely correspond to carbonaceous D bands and G bands, respectively, but are less clearly defined than those reported in fossil carbon (Jehlička et al., 2003; Guedes et al., 2005; Marshall et al., 2010). It is likely to correspond to degraded, complex organic carbon. We note that here we used a laser at 473 nm, and the D band peak is subject to linear variation, depending upon excitation wavelength (Pocsik et al., 1998). A broad peak was obtained at ∼680 cm−1, which corresponds to disordered iron oxides (including ferrihydrite) (De Faria et al., 1997; Mazzetti and Thistlewaite, 2002). In some locations of the mat, well-defined peaks were obtained (Fig. 6A; Spectrum 2), which are indicative of minerals within the mat matrix. The peak at 1028 cm−1 is likely to correspond to pyroxene minerals (Huang et al., 2000; Muniz-Miranda et al., 2009), and the peaks below 600 cm−1 to Fe–O stretching bands from localized crystalline iron oxides (De Faria et al., 1997). Features above 1100 cm−1 may correspond to organic components of colonies of prokaryotes (Zeiri and Efrima, 2005; Smith-Palmer et al., 2010). The surface of rocks that once hosted microbial mats yielded a similar spectrum to Spectrum 1 (Spectrum 3). G and D band features are evident, although they are not as pronounced as those obtained from dried mat.
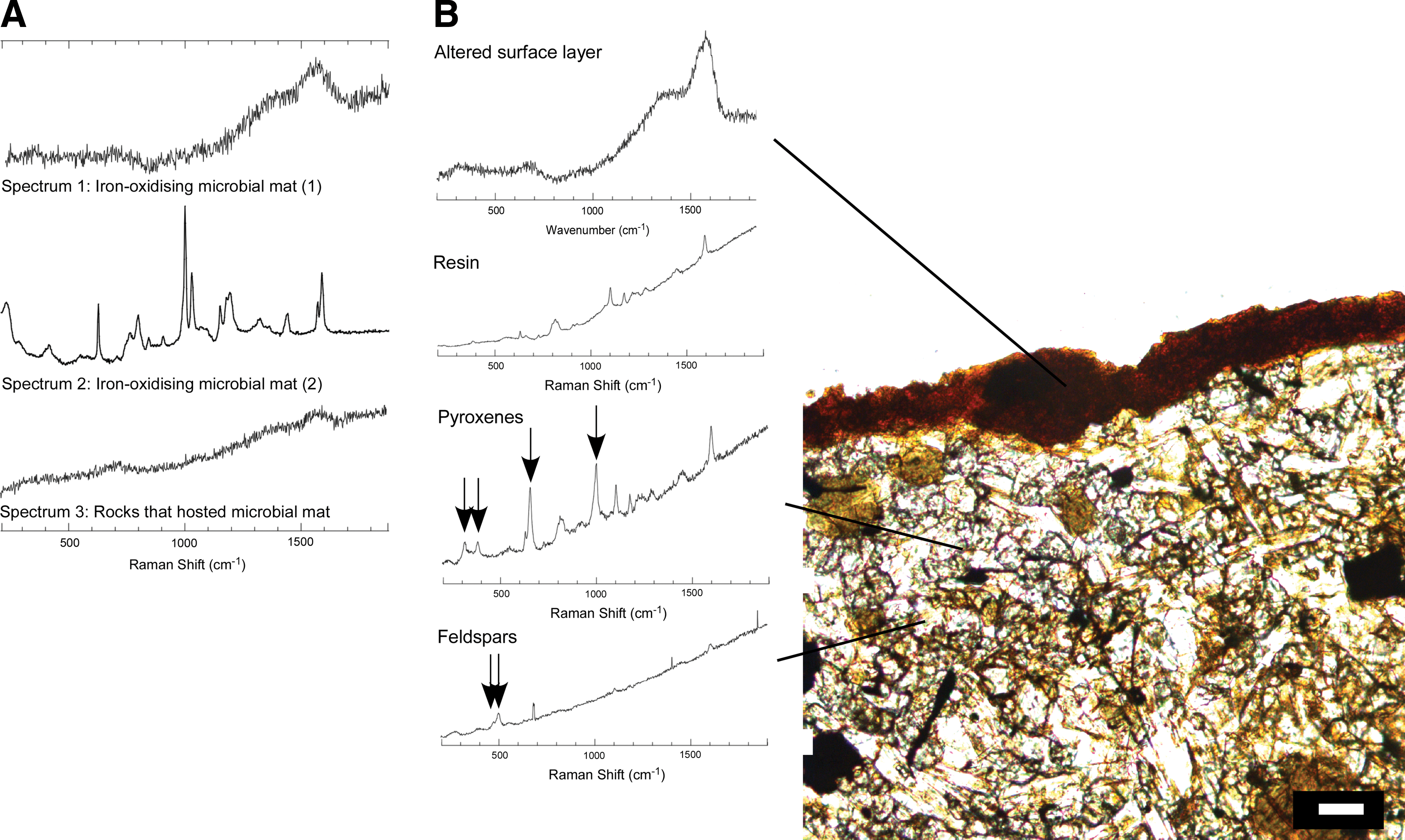
Raman spectroscopic spectra (excitation at 473 nm). (
The spectra obtained from the thin section (cross section of rock on which microbial mats were growing) are consistent with these observations (Fig. 6B). The surface-altered layer of the rock exhibited spectral features identical to Spectrum 1. Underneath the surface layer, spectral features typical of the major components of crystalline basalt were acquired; pyroxenes (four major peaks attributable to pyroxene minerals shown with arrows) and feldspars (double peak with a major peak at ∼500 cm−1 shown with arrows) (Mernagh, 1991; H.G.M. Edwards et al., 2004) but no carbonaceous signatures were observed apart from those attributable to the resin in some regions of the thin section (the pyroxene spectrum shown here exhibits resin signatures). The surface of the rock that once hosted iron-oxidizing mats showed a similar spectrum to the iron-oxidizing mats (Fig. 6A; Spectrum 3): a broad peak of disordered iron oxides ∼680 cm−1 and broad peaks at 1350 and 1590 cm−1, which correspond to carbonaceous material.
3.7. Culture-independent analysis
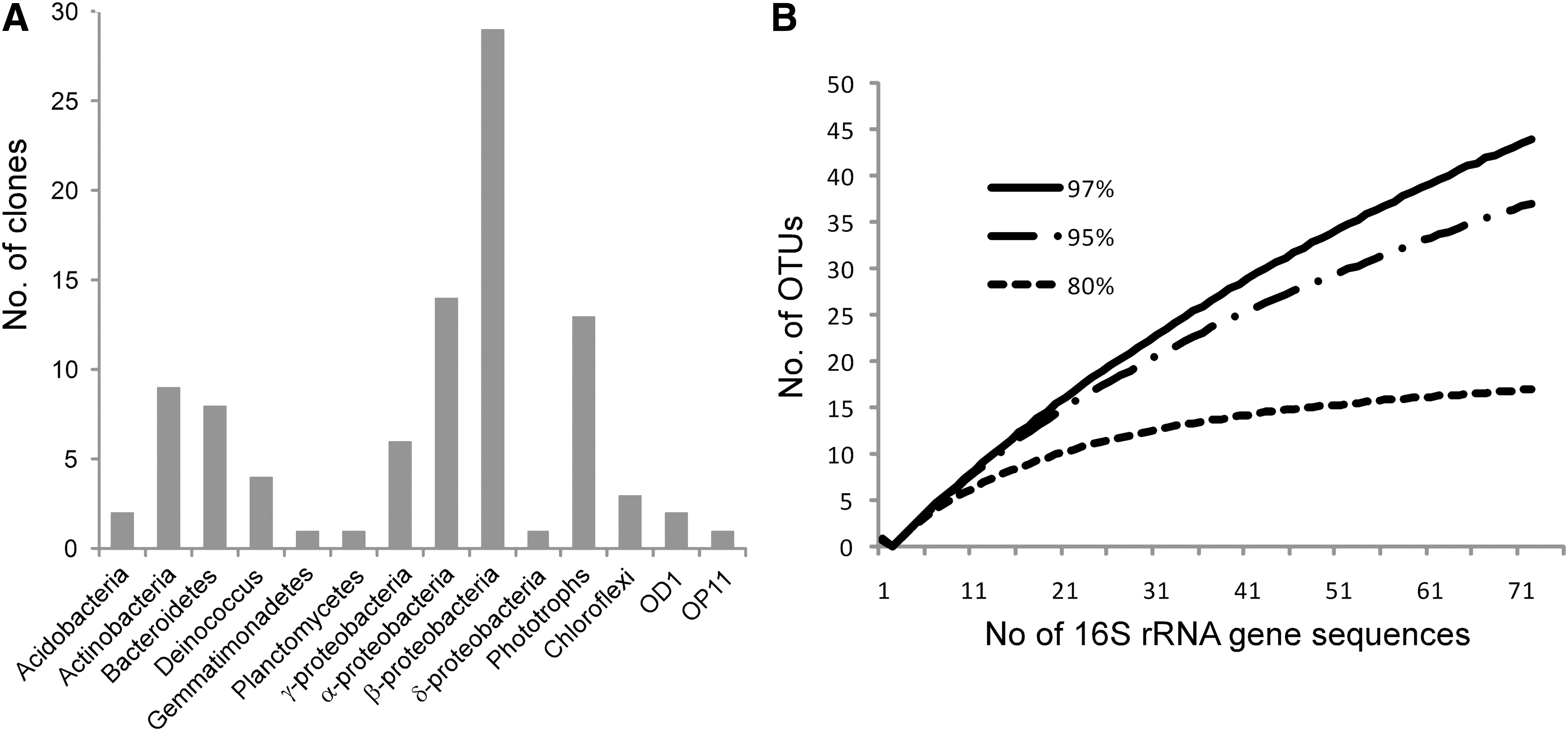
Culture-independent 16S rRNA gene analysis by clone libraries showed a high phylogenetic diversity in the microbial mat (Figs. 7A, 8, 9), which was not sampled to saturation (Fig. 7B).
(

Phylogenetic tree of bacterial taxa within microbial mat and putative phylogenetic relatives. A maximum likelihood tree was generated by using a neighbor-joining tree as the input tree. Aquifex sp. (accession number AB304892) was used as an outgroup. Bootstrap values for nodes with >50% support, based on 1000 replicates, are displayed as percentages. The scale bar represents the number of changes per nucleotide position. Mat sequences are shown in bold. β-proteobacteria shown in Fig. 9.
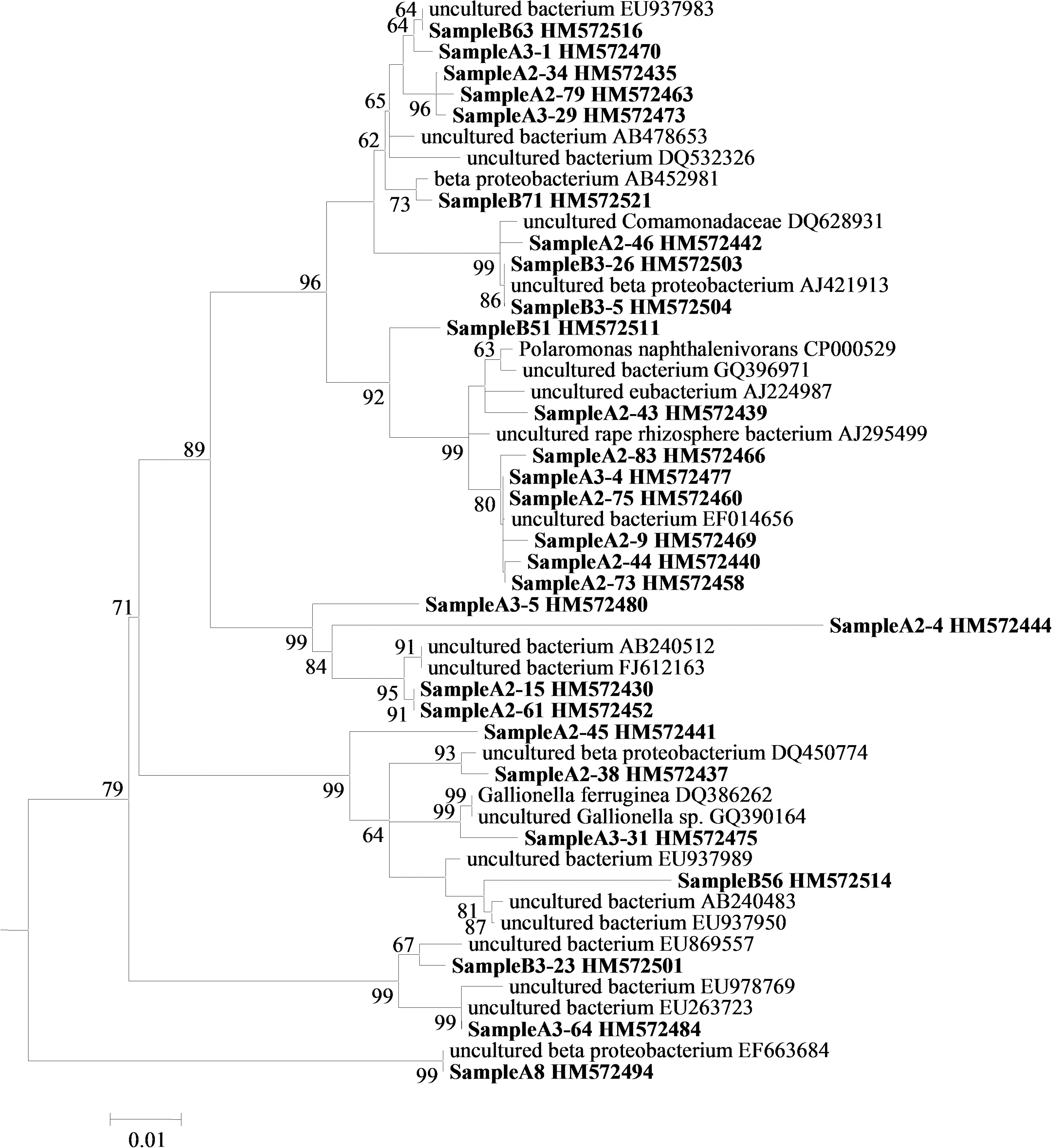
Phylogenetic tree of β-proteobacterial taxa within microbial mat and putative phylogenetic relatives (wedge in Fig. 8). The scale bar represents the number of changes per nucleotide position. Mat sequences are shown in bold.
Proteobacteria constituted 53% of the clones. Representatives of the α-Proteobacteria closely matched uncultured organisms previously reported from moraine meltwaters and glacial streambeds near Mount Everest (DQ675504) and bacteria associated with cyanobacterial blooms (AM989062). Some of the organisms closely associated with sequences obtained from iron-oxidizing biofilms in Shibayama lagoon, Japan, (AB252934) and streams affected by acid mine drainage (FJ203920). Representatives of the β-Proteobacteria matched organisms previously associated with cold freshwater environments. One group closely matched phylotypes previously isolated from a site exhibiting circumneutral iron oxidation (EU937983) and submerged Alpine river biofilms (AB478653) and biofilms of polluted rivers (AJ421913). A 16S rRNA gene sequence most closely related (97%) to Gallionella was identified in the libraries, and it closely matched Gallionella previously described from circumneutral environments (DQ386262; Gallionella capsiferriformans and GQ390164 from the oxic-anoxic interface of Lake Pavin, a volcanic crater lake in the French Massif Central at 45°N). We did not obtain sequences matching Leptothrix spp. Other closely related sequences in this group were obtained from a glacial metagenome (EU978769) and a cold, water-saturated tundra soil (DQ450774).
The δ-Proteobacterial sequence obtained (Geobacter) most closely matched Geobacter psychrophilus (AY653549) (Sitte et al., 2010).
Representatives of the γ-Proteobacteria associated closely with Arctic bacterioplankton (FJ694605), but they also showed 94–95% similarity with Crenothrix and Methylobacter spp. associated with methane oxidation, including methylobacteria sequences previously obtained from volcanic Lake Pavin (GQ390223).
Acidobacteria and Bacteroidetes sequences most closely matched sequences previously obtained in soil and rhizosphere environments.
Phototroph sequences displayed strong affiliation to sequences previously reported from cold and volcanic environments, for example from Arctic streams (FJ849289), seasonal snowmelt, and meltwater in Svalbard, Norway, (FJ946582) and fumarole-fed microbial ecosystems in Chile (FJ592883).
The Actinobacteria sequences obtained matched sequences previously obtained from hot and cold desert streams (EU869760), deglaciated soils (GQ397040), and Antarctic sandy intertidal sediments (GQ496083). Close affiliation was also found to sequences previously retrieved in the phyllosphere (FN421505).
The clone library revealed a number of Deinococcus sequences that closely aligned to those previously observed in mountain endoliths (EF522236) and psychrophilic alpine Deinococcus isolates (e.g., EF635406).
4. Discussion
Iron-oxidizing microbial communities are not well documented in cold environments. Emerson and Revsbech (1994a) reported the presence of iron-oxidizing mats in a stream near Aarhus, Denmark, in which they recorded temperatures of 7°C in winter and 12.5°C in summer. In this study, we describe iron-oxidizing microbial mats in Iceland that use reduced iron derived from volcanic rocks as an electron donor and grow in streams with mean annual temperatures below 5°C.
The streams we studied, which contain products of basaltic rock weathering typical of Icelandic streams (Stefánsson and Gíslason, 2001; Louvat et al., 2008), are inhabited by iron-oxidizing mats, which form large accumulations of amorphous iron oxides. Bright field microscopy and scanning electron microscopy analysis suggested that Gallionella sp. are an abundant iron-oxidizing species in the mat. The ribbon-like polysaccharide extrusions produced by these bacteria are well documented (Ghiorse, 1984; Emerson and Ghiorse, 1992; Hallbeck and Pedersen, 1995; Verran et al., 1995; Hallberg and Ferris, 2004), and, similarly to Hallberg and Ferris (2004), we observed spheroid iron-oxide deposits to be formed on the surfaces of the stalks in the stream environment. The 16S rRNA sequence we obtained that most closely affiliated with Gallionella was most similar to Gallionella previously described from circumneutral environments, including a temperate volcanic crater lake.
Leptothrix was inferred to be an abundant component by the presence of the iron-oxide sheaths observed by bright field microscopy and scanning electron microscopy, although we did not detect them by 16S rRNA gene analysis. One explanation is that sampling was incomplete, consistent with the rarefaction curve, which does not reach an asymptote. However, the paucity of Gallionella sequences might suggest that PCR preferentially selects for other members of the mat community, or the clone library data might reflect the high abundance of other organisms in the mat, which are not morphologically distinct by bright field microscopy.
The presence of Geobacter, a genus of iron-reducing bacteria, detected by 16S rRNA gene analysis, shows that the mats are host to a biological iron cycle. Geobacter might either grow within anaerobic micro-sites within the mat and/or under the mat where we measured low concentrations of oxygen.
There are a variety of potential sources of the reduced iron to sustain the iron-oxidizing microorganisms in the mats. The surfaces of the rocks on which the mats grew were oxidized. Thin section and scanning electron microscopy analyses showed that the oxidized layer is a coherent rind on the rock surface, which is pitted and fractured. Microorganisms are known to be able to access reduced elements from rocks under favorable conditions (Adams et al., 1992; Bailey et al., 2009), and it is plausible that minerals such as pyroxenes and olivines in the crystalline basalt rocks provide a source of reduced iron for the iron-oxidizing bacteria, which would result in the surface rind. However, the oxidized layer, eventually, is likely to physically inhibit the interaction of the microorganisms with the reduced iron-containing minerals in the basalt.
There are other sources of reduced iron to sustain the iron-oxidizing component of the mats. The presence of pyroxenes within the mat observed by Raman spectroscopy and XRD shows that the mats trap particles of reduced iron-containing minerals that are entrained in the stream from weathered rocks, meltwater, sediment load from stream erosion, and other sources. The floccular and corrugated structure of the mat surface has a high surface area in which mineral entrapment could occur.
Another potential source of iron is aqueous reduced iron within the stream itself. Reduced iron is rapidly oxidized in neutral and high pH conditions (Santelli et al., 2001), such as those measured in our study, and it was below detection when using the ferrozine assay. However, we cannot rule out low concentrations of reduced iron, which would be continuously provided by the stream flow. Reduced iron might also be produced by the reduction of oxidized iron either present in the water or in the basaltic rocks on which the mats grow. Iron reduction would be carried out in the anaerobic zone of the mat by iron-reducing bacteria such as Geobacter. It is plausible that all of the above mechanisms contribute to the flow and cycling of reduced iron through the microbial mat to differing degrees.
Temperatures at Site 1 can exceed 20°C; so we cannot rule out the possibility that the organisms are mesophilic and grow only during these brief temperature excursions. Temperatures at Site 2 only once exceeded 16°C during the time period of measurement. These data suggest that the iron-cycling microorganisms are either psychrotolerant, psychrophilic, or both, and the data from Site 1 show that they are freeze-tolerant.
The microbial mats host high bacterial species richness with phylotypes similar to bacteria reported in other stream environments, including representatives of the Actinobacteria, Proteobacteria, and Bacteroidetes. Most of these sequences are most closely related to heterotrophs. The sources of carbon to sustain them are likely to include organics produced from the carbon autotrophically fixed by the iron-oxidizing members of the mats; phototrophs including cyanobacteria, as observed by 16S rRNA gene analysis; and diatoms, as observed by scanning electron microscopy and organic carbon associated with the microbial mat necromass.
Other functions of the mat are difficult to infer from phylogenetic data alone, but the close affiliation of α-Proteobacteria with sequences previously observed in iron-oxidizing biofilms might suggest that they have a role to play in iron oxidation or they are associated with the iron-oxidizing bacterial community in the mat. Other biogeochemical cycles might occur in the mat. For example, sequences in the γ-Proteobacteria had similarity with taxa (Crenothrix and Methylobacter), which have been previously associated with methane oxidation.
The observations reported here yield insights into the distribution of iron-oxidizing bacteria in cold volcanic environments. In no previous studies of exposed surface volcanic rocks of a variety of compositions and mineralogies in Iceland have we observed known iron-oxidizing bacteria by culture-independent methods (e.g., Herrera et al., 2008, 2009; Cockell et al., 2009a, 2009b, Kelly et al., 2010), nor have they been reported in surface volcanic rocks and ash in other regions of the world (Gomez-Alvarez et al., 2007; Ibekwe et al., 2007). They are present, however, in rocks in environments with more rapid weathering, for example, in tropical diorite saprolites (Buss et al., 2005). One factor that may explain the apparent lack of abundant iron-oxidizing organisms in surface rocks in Iceland is the kinetic limitation to the acquisition of reduced iron from the rocks in most environments under neutral conditions, even during periods of rainfall in Iceland when rock weathering can occur (Gíslason and Eugster, 1987; Stefánsson et al., 2001; Wolff-Boenisch et al., 2004). We cannot discount a limitation in other bioessential elements for iron-oxidizing bacteria in surface rock environments, but the presence of a large diversity of heterotrophic bacteria in the rocks (Kelly et al., 2010) suggests that other required elements are available. The observations here show that in these cold environments the weathering of large numbers of rocks in a watershed and the weathering of rocks in streams is capable of achieving sufficient iron concentrations to sustain iron-oxidizing mats. We note that we have not observed iron-oxidizing mats in large rivers or fast-running streams in Iceland. One hypothesis to explain this observation might be that fast flow disrupts and prevents the formation of stable microbial mat communities.
The mats exhibited Raman G and D bands associated with disordered carbonaceous material (Marshall et al., 2010). Apart from localized areas, we did not observe well-defined peaks associated with specific macromolecules that are observed in individual species of microorganisms (Zeiri and Efrima, 2005; Smith-Palmer et al., 2010).
The rocks we examined that were once part of the stream also preserved the G and D band carbonaceous signatures within their altered surface. The surface layer could not be easily removed with a scalpel blade. Either it represents dried biomass and iron oxides from the mats that have become attached to the rock surface, or it is the oxidized surface of the basaltic rock itself that was in contact with the mat. The observations suggest that the carbonaceous material of the rock-dwelling mats has become preserved within the altered surface of the rocks and could be used as one diagnostic indicator of past epilithic communities in aqueous environments, if abiotic sources of carbonaceous deposition on the rocks can be discounted (Pasteris and Wopenka, 2003).
4.1. Astrobiological implications
These data have implications for two areas of astrobiology: the interpretation of the fossil record and testing the hypothesis of past life on Mars.
The presence of iron-oxidizing bacteria in cold volcanic terrains raises questions about the provenance of fossil biogenic iron oxides. For example, Hofmann et al. (2008) reported the presence of Gallionella-like stalks in microcrystalline quartz-chalcedony infillings in basalt from Breiddalur, Iceland. Although the Gallionella-like features that these authors observed might have grown in warm hydrothermal environments, the incorporation of fossilizable iron oxides from iron-oxidizing microorganisms growing in low-temperature environments into hydrothermal deposits might be possible in dynamic hydrothermal settings. We have observed iron-oxidizing microbial mats in perennially cold habitats adjacent (within tens of centimeters) to silica-precipitating hot springs in Iceland (unpublished data). The data here show that the detection of Gallionella stalks in ancient volcanic rocks cannot be taken as prima facie evidence of growth in warm- or high-temperature hydrothermal environments.
The observations have applications to the search for life on other planetary bodies, particularly Mars. In this study, we investigated aerobic iron oxidizers. On Mars, the rocks are not in contact with oxygen in the atmosphere, but other plausible pathways for iron oxidation are iron phototrophy or anaerobic iron oxidation, both of which could use reduced Fe derived from weathered basaltic rocks in a similar way to aerobic iron oxidation. Anaerobic iron oxidation uses nitrate as an electron acceptor. Without a biological nitrogen cycle, fixed nitrogen would be produced in low abundance on Mars. However, fixed nitrogen could have been produced by volcanic or impact processing in the early history of the planet (Segura and Navarro-González, 2005; Summers and Khare, 2007; Manning et al., 2009).
The data show that promising locations where the hypothesis that Mars sustained past life can be tested are not necessarily any environments that theoretically have supplies of energy and water, such as isolated outcrops of volcanic rocks. The volcanic rocks in Iceland contain abundant reduced iron (between ∼3% and 14% depending on the type of rock) in contact with atmospheric oxygen, which thereby offers a thermodynamic disequilibrium that can be exploited by iron-oxidizing chemolithotrophs. The surface rocks are intermittently exposed to water in the form of snowmelt and rain, yet there is apparently a kinetic limitation to iron oxidation in most Icelandic rocks. The data are a demonstration of a “follow-the-kinetics” approach to life detection. In the analogy of Hoehler et al. (2007), the kinetic limitation to reduced iron availability from the rocks is a restriction in power availability to organisms. The approach shows that good locations to search for life may be in ancient rivers, streams, subglacial environments and other locations where sustained water movement could mitigate the kinetic limitations to the acquisition of mineral electron donors and acceptors.
Finally, the Raman spectroscopic analysis of the carbonaceous material within the rock surface showed that Raman spectroscopy could be used to search for the carbonaceous signatures of biota incorporated into the surface of rocks in once-cold basaltic water-rich environments on Mars. G and D bands are associated with non-biological carbonaceous materials (Pasteris and Wopenka, 2003; Marshall et al., 2010); so they are not unequivocal signatures of biological activity. However, given the known geological context of the materials analyzed in this study (i.e., the lack of exposure to hydrothermal sources of potential carbonaceous material), the presence of G and D bands could provide one line of evidence indicative of biotic-rock interactions.
The lack of evidence of chemolithotrophy in apparently favorable martian environments would not definitively prove the lack of life on Mars, but it would suggest important constraints on either the origin, or potential sustenance, of life on Mars.
Footnotes
Acknowledgments
This work was made possible by funding from the Leverhulme Trust (Project No. F/00 269/N). We thank William Schopf and Anatoliy Kudryavtsev for carrying out corroborative Raman spectroscopic investigations on the mat material and for discussions on its interpretation.
Abbreviations
FEG, field emission gun; SEM, scanning electron microscope; XRD, X-ray diffraction.