
Abstract
The haloarchaea Natrialba magadii and Haloferax volcanii, as well as the radiation-resistant bacterium Deinococcus radiodurans, were exposed to vacuum UV (VUV) radiation at the Brazilian Synchrotron Light Laboratory. Cell monolayers (containing 105 to 106 cells per sample) were prepared over polycarbonate filters and irradiated under high vacuum (10−5 Pa) with polychromatic synchrotron radiation. N. magadii was remarkably resistant to high vacuum with a survival fraction of (3.77±0.76)×10−2, which was larger than that of D. radiodurans (1.13±0.23)×10−2. The survival fraction of the haloarchaea H. volcanii, of (3.60±1.80)×10−4, was much smaller. Radiation resistance profiles were similar between the haloarchaea and D. radiodurans for fluences up to 150 J m−2. For fluences larger than 150 J m−2, there was a significant decrease in the survival of haloarchaea, and in particular H. volcanii did not survive. Survival for D. radiodurans was 1% after exposure to the higher VUV fluence (1350 J m−2), while N. magadii had a survival lower than 0.1%. Such survival fractions are discussed regarding the possibility of interplanetary transfer of viable microorganisms and the possible existence of microbial life in extraterrestrial salty environments such as the planet Mars and Jupiter's moon Europa. This is the first work to report survival of haloarchaea under simulated interplanetary conditions. Key Words: Vacuum ultraviolet—Synchrotron—Radiation resistance—Planetary protection—Panspermia. Astrobiology 11, 1034–1040.
1. Introduction
T
The extraterrestrial environment is considered lethal for organisms due to the high levels of radiation, high-vacuum conditions, and extreme temperatures, all of which could have an impact on relevant macromolecules such as nucleic acids and lead to increased mutation rates, cell damage, and inactivation (Horneck, 1999; Paulino-Lima et al., 2010). In particular, UV radiation has been mostly studied because it causes extensive damage to cells and can be utilized as an indicator of habitability elsewhere (see, e.g., Buccino et al., 2006, 2007).
Survival of microorganisms and biomolecules under space conditions has been evaluated in many ways, either under direct exposure in real flight missions in Earth's orbit or under simulated space UV radiation and vacuum (Horneck et al., 2010; Olsson-Francis and Cockell, 2010). Different microorganisms have been tested, including bacteria, fungi, bacterial or fungal spores, and viruses, as well as biomolecules such as DNA, amino acids, and liposomes (Horneck et al., 2010; Olsson-Francis and Cockell, 2010).
Negligible UV radiation shorter than 290 nm reaches Earth's surface because it is strongly absorbed by the atmosphere. UV radiation should be considered, however, in studies that concern life in extraterrestrial environments. Unfortunately, the experimental techniques used to work with these wavelengths are complex due to the requirement of vacuum conditions and high radiation fluxes (Cefalas et al., 1993; Sarantopoulou et al., 1994). Nonetheless, a number of authors have reported effects of such wavelengths on microorganisms (Koike and Oshima, 1993; Cefalas et al., 2001; Saffary et al., 2002; Schuerger et al., 2003; Heise et al., 2004; Newcombe et al., 2005; Clauss, 2006; Sarantopoulou et al., 2006; Schuerger et al., 2006; Tauscher et al., 2006; Fajardo-Cavazos et al., 2010; Galletta et al., 2010, Wassmann et al., 2010; Sarantopoulou et al., 2011).
In the present study, we investigated for the first time in an extraterrestrial simulation facility the survival of two different non-sporulating halophilic archaea or haloarchaea (family Halobacteriaceae), Haloferax volcanii (Mullakhanbhai and Larsen, 1975) and Natrialba magadii (Tindall et al., 1984; Kamekura et al., 1997). It is well known that extreme halophiles such as haloarchaea are relevant to astrobiology not only due to their capacity to inhabit environments of high salinity, for example, Mars (Mancinelli et al., 2004) or Jupiter's moon Europa (Marion et al., 2003), but also for their ability to cope with extreme temperatures, pH, and radiation.
These strains, as well as the radiation-resistant non-sporulating bacterium Deinococcus radiodurans, were desiccated and subjected to UV and vacuum conditions similar to those found in interplanetary space near Earth orbit (Table 1). Our experiments took place at the Brazilian Synchrotron Light Laboratory located in Campinas, Brazil, where we implemented our investigation with use of the synchrotron beamline equipped with a toroidal grating monochromator (TGM) and an end station with a vacuum chamber (Cavasso Filho et al., 2007).
Values differ depending on the orientation and the distance to the Sun.
Values in Earth orbit depend on outgassing of the spacecraft.
2. Material and Methods
Haloferax volcanii (DS70) and N. magadii (ATCC 43099) strains were kindly provided by Dr. R.E. de Castro, Universidad Nacional de Mar del Plata, Argentina; and D. radiodurans (R1 wild type strain) was obtained at Instituto de Radioproteção e Dosimetria, Rio de Janeiro, Brazil.
Haloferax volcanii was grown aerobically at 30°C with shaking at 200 rpm and was cultivated in Hv-YPC broth (Kauri et al., 1990) that contained (g L−1): yeast extract (5); peptone (1); casaminoacids (1); NaCl (144); MgSO4·7H2O (21); MgCl·6H2O (18); KCl (4.2); CaCl2 (3 mM); and Tris-HCl (12 mM), with pH adjusted to 6.8. N. magadii was grown aerobically at 37°C with shaking at 200 rpm. Growth medium composition was (g L−1): yeast extract (5); NaCl (200); Na2CO3 (18.5); sodium citrate (3); KCl (2); MgSO4·7H2O (1); MnCl2·4H2O (3.6×10−4); FeSO4·7H2O (5×10−3), with pH adjusted to 10 (modified from Tindall et al., 1984). D. radiodurans was cultivated in TGY broth that contained (g L−1) tryptone (10); yeast extract (6); glucose (2) (Anderson et al., 1956) with shaking at 200 rpm, at 32°C for 10 h, or until the early stationary phase was reached.
The optical density of the cultures was spectrophotometrically measured at λ=600 nm, and aliquots of 10 μL were taken for direct cell counting under the microscope. Results were compared to colony-forming units (CFUs) grown on agar-solidified culture media to assess the viability of the culture. After the estimation of the cell concentration, 1 mL of haloarchaea cultures was washed twice in saline solutions with the same composition and pH of the culture medium but without organic compounds. Similarly, 1 mL of the D. radiodurans culture was washed twice in previously sterilized distilled water. At this stage, aliquots of these cultures were separated and kept outside the vacuum chamber to be used as controls. Other aliquots were taken to expose the cells to vacuum alone and to vacuum plus UV in the TGM workstation. All experiments were performed in triplicate.
The samples to be exposed were prepared by using a hexagonal copper sample-holder that can be introduced inside the vacuum chamber of the workstation. Several square pieces of polycarbonate filter (Millipore) with a surface area of approximately 25 mm2 were mounted on the surface of the sample-holder with double-sided carbon tape. The system was sterilized by exposure to germicidal UV lamp (254 nm, 1620 J m−2) in a laminar flow. After sterilization, a volume of 1 μL of the cell suspensions (containing 108 to 109 cells per milliliter) was loaded on the polycarbonate filters (monolayers containing 105 to 106 cells per sample). The system was kept inside the laminar flow for at least 30 min for dehydration of the samples. The sample-holder was then placed inside the vacuum chamber at the TGM beamline workstation.
Inside this chamber, cells were exposed to decreasing vacuum pressure for 3 h, due to the required pumping time needed to reach high vacuum (10−5 Pa) from atmospheric pressure, before irradiation. This pressure was maintained throughout the entire irradiation experiment, the duration of which was about 1 h. At the end of the experiment, venting the chamber took 15 min.
Samples were exposed at room temperature to UV plus vacuum UV (VUV) radiation. A gas filter (neon) was interposed between the beamline and the experimental chamber to attenuate the X-ray portion of the synchrotron radiation spectrum, below 57.6 nm. The synchrotron emission reached 400 nm, but the emitted energy decays exponentially beyond this, which results in an effective cutoff of longer wavelengths after 124 nm. Therefore, the complete wavelength ranged from 57.6 to 124 nm (Fig. 1). The exposure times were 0, 10, 30, 100, 300, and 900 s, which resulted in fluences of 0, 15, 45, 150, 450, and 1350 J m−2, respectively. The photon flux was measured by using photodiodes to check for electrical current variations in real time. The irradiance of the TGM beamline is compared with those at the orbits of Earth, Mars, and Europa in Table 2.
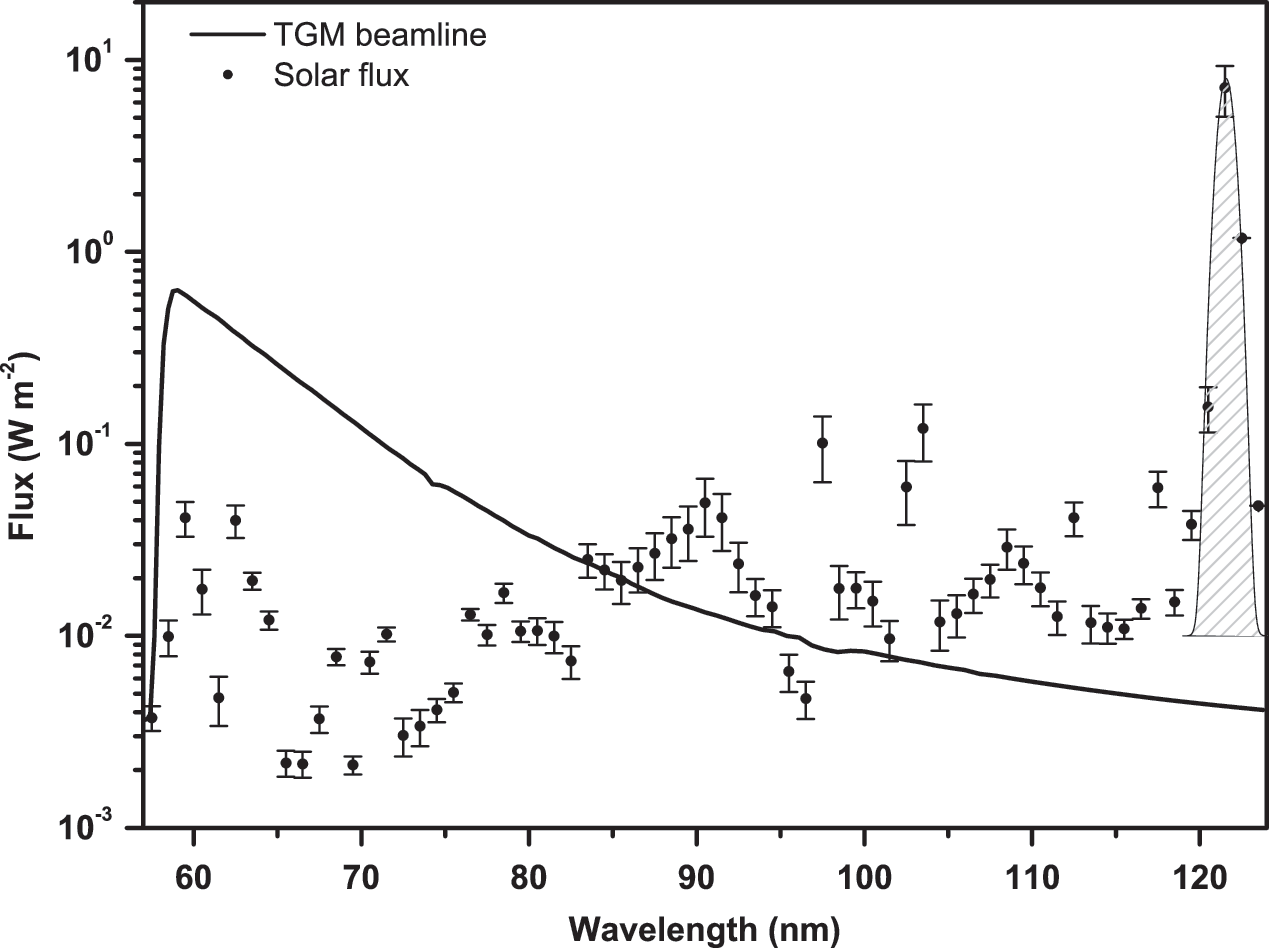
TGM beamline spectrum in comparison to the solar flux as obtained with the software Solar 2000 v2.36 with database S2K+ASTM490, with daily solar spectra from 1999–2010 (covering a full 11-year solar cycle). The error bars represent the dispersion of the data (its variation throughout the years). The curve at 121.6 nm is the Lyman-alpha line.
MIR, mid-infrared; NIR, near infrared; VIS, visible.
Based on Schuerger et al. (2003).
Calculated in this study.
Based on direct measurements at the TGM beamline using photodiodes.
After exposure, 10 μL of culture medium was dropped on the polycarbonate filters and mixed with a micropipette for 10 s to recover the cells. Afterward, samples were diluted and aliquots seeded on nutrient plates, which were incubated. The efficiency of cell recovery was determined to be between 40% and 50% by comparison with cell counts scored in nondeposited samples.
Survival fractions were calculated as N/N 0, where N is the number of CFUs after treatment and N 0 is the number of CFUs that correspond to counts of the non-irradiated sample. Survival fractions for the exposure to vacuum alone and for the exposure to vacuum plus irradiation at several doses are shown in Figs. 2 and 3, respectively.

Survival fractions (N/N 0) to high vacuum (10−5 Pa; non-irradiated), where N is the number of CFUs after treatment and N 0 is the number of CFUs corresponding to counts of the non-irradiated sample. For each species, the mean values of three replicates are shown, and the error bars represent the standard deviations. Survival rates were calculated considering controls without any treatment (20±1.41)×108 UFC/mL for N. magadii, (6.80±3.11)×108 UFC/mL for H. volcanii, and (2.8±1.13)×108 UFC/mL for D. radiodurans. Significant differences were observed between strains in all comparisons (Fisher's least-difference mean separation test, p<0.05).
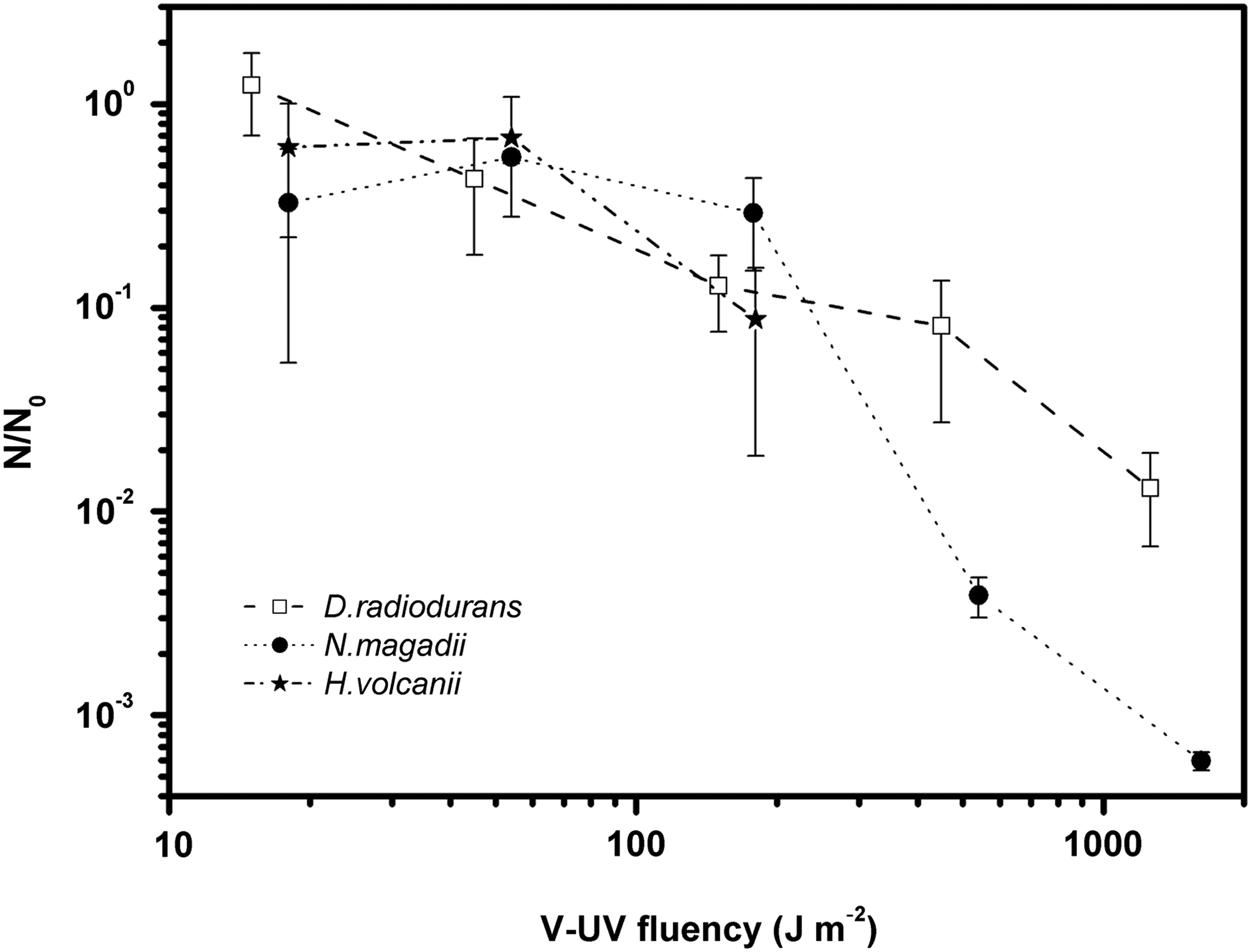
Survival curves to VUV radiation at the TGM beamline, where N/N 0 is the survival fraction (N is the number of CFUs after treatment, and N 0 is the number of CFUs corresponding to counts of the non-irradiated sample). For each species, the mean values of three replicates are shown, and the error bars represent the standard deviations. No survivors were detected for H. volcanii at fluence rates above 150 J m−2. Statistical tests (Fisher's least-difference mean separation test) showed no significant differences between strains for fluences up to 150 J m−2 (p>0.05). For fluences above 150 J m−2, on the other hand, the differences observed were significant (p<0.05).
To assess the statistical significance of the results, analysis of variance tests were performed, followed by a Fisher's least-difference mean separation test (p=0.05) with the OriginPro8 software (OriginLab Corporation, USA). Since the difference between our data is close to, or higher than, 1 order of magnitude, data were log-transformed to achieve homogeneity of variances.
3. Results
The effects of vacuum alone on survival fractions of N. magadii, H. volcanii, and D. radiodurans were measured. As shown in Fig. 2, the haloarchaea N. magadii showed the maximum survival fraction to high vacuum (10−5 Pa), which corresponds to (3.77±0.76)×10−2. This fraction was almost 3-fold higher than that of D. radiodurans, which had a survival fraction of (1.13±0.23)×10−2 under the same conditions. Surprisingly, the survival fraction of the haloarchaea H. volcanii was (3.60±1.80)×10−4, which is remarkably low (2 orders of magnitude below values obtained for the other strains) compared with survival fractions for N. magadii and D. radiodurans. The statistical analysis showed that the survival fractions are significantly different at p<0.05 in all cases.
The samples that were subjected to vacuum only were kept in the same chamber where the irradiations were performed. Therefore, the values obtained for the vacuum exposure alone were taken as controls in order to calculate the survival fractions obtained for VUV experiments.
While exposed to high vacuum, both haloarchaea and D. radiodurans were exposed to synchrotron VUV radiation at different doses (0, 15, 45, 150, 450, and 1350 J m−2). As shown in Fig. 3, the three strains show similar survival fractions for fluences up to 150 J m−2. In fact, the differences in the survival fractions up to 150 J m−2 are not significant according to the statistical tests (p>0.05). At higher fluences, above 150 J m−2, the survival of the haloarchaeal strains diminishes much more rapidly in comparison to survival of D. radiodurans. In particular, we detected no survivors for H. volcanii. Statistical analysis showed that these differences are significant (p<0.05). Fluencies that resulted in 10% of survivors in the cell population (LD90) were similar between haloarchaea and D. radiodurans, between 200 and 300 J m−2.
4. Discussion
It is known that D. radiodurans is very resistant to desiccation under the present simulation setup (Paulino-Lima et al., 2010). For example, in an experiment in which simulated martian soil was used under low pressure, 30% of the initial community of D. radiodurans survived for 10 days, while E. coli did not survive (Diaz and Schulze-Makuch, 2006). However, our results show that N. magadii has a survival fraction 3-fold higher than that for D. radiodurans (Fig. 2). This could be explained by a more efficient mechanism against desiccation present in haloarchaea in general, due to their adaptation to environmental stresses caused by the high temperatures and salt concentrations in their natural habitat. A particular osmoadaptation mechanism in haloarchaea is based on the presence of high levels of intracellular concentrations of K+ ions inside the cell (Oren, 1999). For example, Kottemann et al. (2005) found that 25% of Halobacterium cells survived after 20 days under high vacuum (10−6 Pa).
On the other hand, exposure of H. volcanii to vacuum reduced its survival 100-fold compared to that of N. magadii. It should be noted that the protection mechanisms against desiccation should be stronger for N. magadii, which is an extreme halophile that lives in salt concentrations between 3.5 and 4.0 M NaCl, than it is for H. volcanii, a moderate halophile that lives in the 1.7–2.5 M NaCl range. Furthermore, in special conditions, such as nutrient starvation, haloalkaliphilic strains like N. magadii are also capable of producing an organic solute (2-sulfotrehalose) that, in part, replaces intracellular KCl (Desmarais et al., 1997), a mechanism probably not present in H. volcanii, which lives in neutral pH.
Regarding UV resistance, McCready et al. (2005) compared the resistance profiles to UVC between the haloarchaea Halobacterium NRC-1 and D. radiodurans at different fluences (between 0 and 200 J m−2) and showed that both microorganisms are highly resistant to UVC radiation.
In this paper, the VUV range from 57.6 to 124 nm was focused as astrobiologically relevant in terms of microbial life spreading within the Solar System. Interestingly, unlike UVC photons, which penetrate deep into the cell and damage DNA, VUV photons can be mostly absorbed by the cell membrane (Cefalas, 2005; Sarantopoulou et al., 2006). In particular, Yagi et al. (2009) found that at 62 nm the absorption depth is around 1 nm, which is considerably less than the width of a typical haloarchaea membrane, which measures in the tens of nanometers, depending on the composition of the membrane (Steensland and Larsen, 1969). All the microorganisms studied in this work have similar cell walls, given that D. radiodurans has an atypical wall of similar composition to haloarchaea. Consistently, our radiation resistance profiles are similar between both haloarchaea and D. radiodurans, at least for fluences up to 150 J m−2 (Fig. 3).
For fluences greater than 150 J m−2, survival fractions are higher for D. radiodurans than for haloarchaea. In particular, H. volcanii did not survive beyond that fluence, probably due to synergistic effects between VUV and high vacuum. It should be noted that the D. radiodurans survival fraction is 2-fold higher than that of N. magadii, and therefore the radiation pathway to internal components of the cell as genetic material is twice as large. We point out that, although the composition of the cell wall is similar between D. radiodurans and haloarchaea, the cell wall structure of N. magadii is not known but should account for the observed VUV resistance.
5. Conclusions
This is the first work to report survival of haloarchaea under simulated interplanetary conditions. We measured the survival of haloarchaeal cells Natrialba magadii and Haloferax volcanii under irradiation with VUV photons in the range 57.5–124 nm. We used a VUV flux similar to the solar one at 1 AU and pressures similar to those found in low Earth orbit or on the surface of Europa (which is below 10−4 Pa). The survival curves were compared with the response of D. radiodurans, a microorganism considered a good candidate to endure the extreme conditions found in space and on the surface of other planets and moons (Dose et al., 1996; Saffary et al., 2002; Clauss, 2006; Diaz and Schulze-Makuch, 2006; de La Vega et al., 2007; Paulino-Lima et al., 2010).
Our results indicate that unprotected D. radiodurans cells drop to 1% survival at 1350 J m−2, while N. magadii survives some 0.1% to the same exposure. H. volcanii, on the other hand, did not survive beyond 150 J m−2 fluences. These survival fractions show that cells of H. volcanii and N. magadii fully exposed to solar VUV irradiation on a planetary surface or on meteorites would be quickly depleted by at least 3 orders of magnitude. However, several cells did survive, and much longer exposure times need to be tested to discern whether at least a small number of cells of N. magadii and D. radiodurans could survive, without protection, the VUV and vacuum damages present in space.
Footnotes
Acknowledgments
We would like to thank Laboratorio Nacional de Luz Síncrotron (LNLS), and Centro de Ciência e Tecnologia do Bioetanol, Campinas, Brazil, which partially supported this work and also provided the facility and the staff supervision to conduct part of the experiments. We acknowledge the comments by the referees, which helped to improve the paper.
Abbreviations
CFUs, colony-forming units; TGM, toroidal grating monochromator; VUV, vacuum UV.