
Abstract
Manganese oxide (Mn oxide) minerals from bacterial sources produce electron paramagnetic resonance (EPR) spectral signatures that are mostly distinct from those of synthetic simulants and abiogenic mineral Mn oxides. Biogenic Mn oxides exhibit only narrow EPR spectral linewidths (∼500 G), whereas abiogenic Mn oxides produce spectral linewidths that are 2–6 times broader and range from 1200 to 3000 G. This distinction is consistent with X-ray structural observations that biogenic Mn oxides have abundant layer site vacancies and edge terminations and are mostly of single ionic species [i.e., Mn(IV)], all of which favor narrow EPR linewidths. In contrast, abiogenic Mn oxides have fewer lattice vacancies, larger particle sizes, and mixed ionic species [Mn(III) and Mn(IV)], which lead to the broader linewidths. These properties could be utilized in the search for extraterrestrial physicochemical biosignatures, for example, on Mars missions that include a miniature version of an EPR spectrometer. Key Word: Spectroscopic biosignatures. Astrobiology 11, 775–786.
1. Introduction
Unlike iron, which is easily oxidized at neutral pH values, reduced manganese [Mn(II)] is stable as a redox-neutral dissolved salt at pH values up to 8.5; so Mn oxides are not common despite the fact that Mn is a relatively abundant metal on the Earth. In fact, abiotic homogeneous oxidation of Mn(II) is very slow up to pH values of 8.5 and requires years for completion at modest concentrations (Ehrlich, 2002; Tebo et al., 2005). For most of Earth's surface and for biological cytoplasm, which tends to occur at circumneutral pH and relatively low Eh, it might be expected that Mn(II) would be the dominant form and abundant Mn (IV) oxides would be rather rare. Given this paucity and the fact that many different types of microbes are known to catalyze the oxidation of Mn(II) to Mn(IV) oxides, it is widely believed that the primary Mn oxides in natural waters and oxic surface environments are often, if not usually, the result of biological catalysis (Tebo et al., 2005; Bargar et al., 2009). This argument, for example, leads to the inference that desert varnish, Mn nodules, and other such precipitates are likely to be biological in origin; however, this supposition requires proof with convincing data.
Results of our analyses from electron paramagnetic resonance (EPR) spectra (Fig. 1) suggest that it is in fact possible to distinguish biogenic from abiogenic MnO2, which leads to the exciting possibility that MnO2 could be used as a potent biosignature in the search for life. As far as we know, this is the first application of EPR to the search for Mn oxide biosignatures.
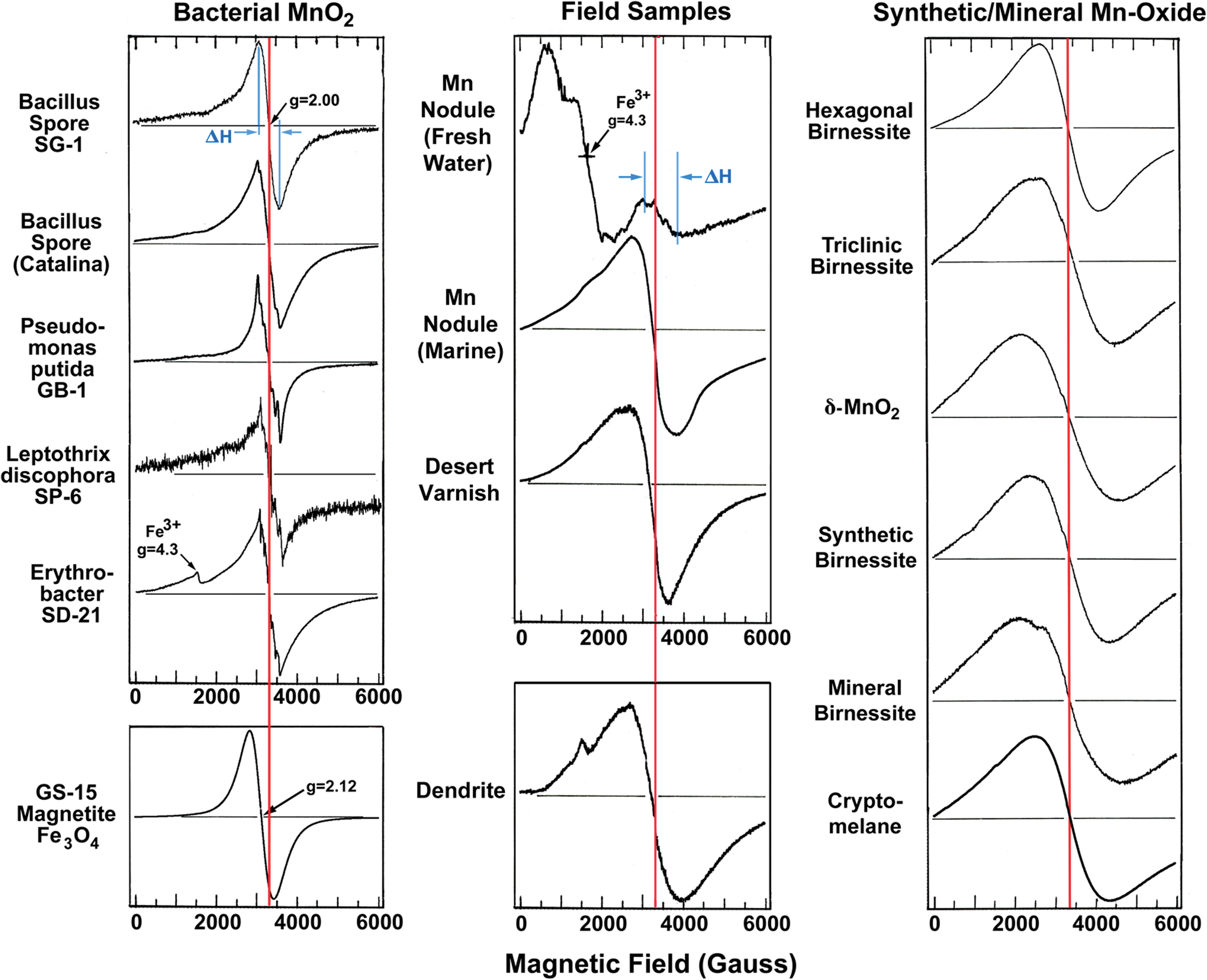
Left: EPR spectra of bacterial Mn oxide samples. Some of the bacterial Mn oxides show overlapped six-line hyperfine structure characteristic of highly diluted Mn(II); however, quantitatively, the Mn(II) contribution is very low when estimated by the area under the curve. The EPR spectrum of superparamagnetic biogenic magnetite induced by GS-15 (courtesy of P.R. Buseck) is shown for a comparison. Middle: EPR spectra of field samples. Right: EPR spectra of synthetic and mineral counterparts. The red lines mark the g=2 position; and EPR linewidth, ΔH, definition is shown in the top spectra. Color graphics available online at
1.1. Biologically formed Mn oxides
On Earth, a broad range of bacteria have the capability to oxidize Mn(II) to Mn(IV) and thereby precipitate Mn oxides. Microbes that can oxidize Mn(II) include Gram-positive and Gram-negative bacteria, a number of different filamentous fungi (Ehrlich, 2002; Tebo et al., 2005), and a few cyanobacteria and algae, which appear to oxidize the Mn(II) via local changes in Eh and pH (Richardson et al., 1988). To date, no reports of archaea that oxidize Mn(II) have appeared. Studies of the mechanism of Mn oxidation include genetic studies to identify the genes and proteins involved (Francis and Tebo, 2001; Francis et al., 2001), as well as studies that have utilized synchrotron radiation to monitor changes in the redox state of the metal during oxidation (Tebo et al., 2004). Genetic/physiological studies have revealed that many (perhaps all) of the bacteria known to oxidize Mn utilize a multi-copper oxidase enzyme system (Francis et al., 2002; Tebo et al., 2005), although only a few such organisms have been studied to date. The mechanistic studies have revealed that a transient state of Mn(III) appears to be involved in the oxidation but that it is not part of the final product; that is, Mn(III) is probably an enzyme-bound intermediate that is the result of a one-electron transfer mechanism, but the final product is Mn(IV). This could account for the low content of Mn(III) in the biogenic Mn oxide samples discussed in this report.
2. Materials and Methods
We studied sets of Mn oxide samples: bacterial samples, synthetic analogues, field-collected samples of suspected biominerals, and natural mineral samples from a commercial source (Minerals Unlimited). The chemical MnO and MnO2 were purchased from Alfa Aestar.
2.1. Natural mineral samples
The local structures around Mn atoms in the natural mineral samples were characterized by extended X-ray absorption fine structure (EXAFS). EXAFS data were collected for the following Mn oxide polymorphs: lithiophorite, hexagonal birnessite, mineral birnessite, chalcophanite, coronadite, cryptomelane, ramsdellite, and pyrolusite. EXAFS spectra, their Fourier transforms, and representative structures are shown in Fig. 2 in the order of increasing relative proportion of corner-sharing Mn-Mn pairs (with an interatomic distance of ca. 3.5 Å) and, at the same time, decreasing proportion of edge-sharing Mn (at ca. 2.85 Å). For example, hexagonal birnessite and lithiophorite are layered Mn oxides. Lithiophorite has no corner-sharing Mn-Mn pairs, and hexagonal birnessite has only a few layers created when Mn(II)/(III) atoms bond above layer vacancies (Webb et al., 2005a, 2005b; Villalobos et al., 2006). Chalcophanite is a layered Mn oxide that contains corner-sharing Mn-Zn neighbors (one zinc atom for every seven Mn atoms) (Post and Appleman, 1988). The tunnel-structure oxides (coronadite, cryptomelane, ramsdellite) have corner-sharing Mn-Mn pairs along the “folds” of the structure, with an overall higher proportion of these sites in structures with smaller tunnels (i.e., ramsdellite; Post and Heaney, 2004). Pyrolusite, with a rutile structure, contains the highest abundance of corner-sharing Mn-Mn pairs. Fourier transforms of EXAFS spectra identify these systematics. The corner-sharing Mn-Mn pair correlation is very weak in hexagonal birnessite, as expected. However, the amplitude of this corner-sharing pair correlation increases systematically through this mineral series, which results in a 5-fold higher amplitude in pyrolusite. Concurrently, the amplitude of the edge-sharing Mn-Mn pair correlation decreases as the size of the structural layer motif (including the layerlike sides of the tunnels in coronadite, cryptomelane, and ramsdellite) decreases through this series.
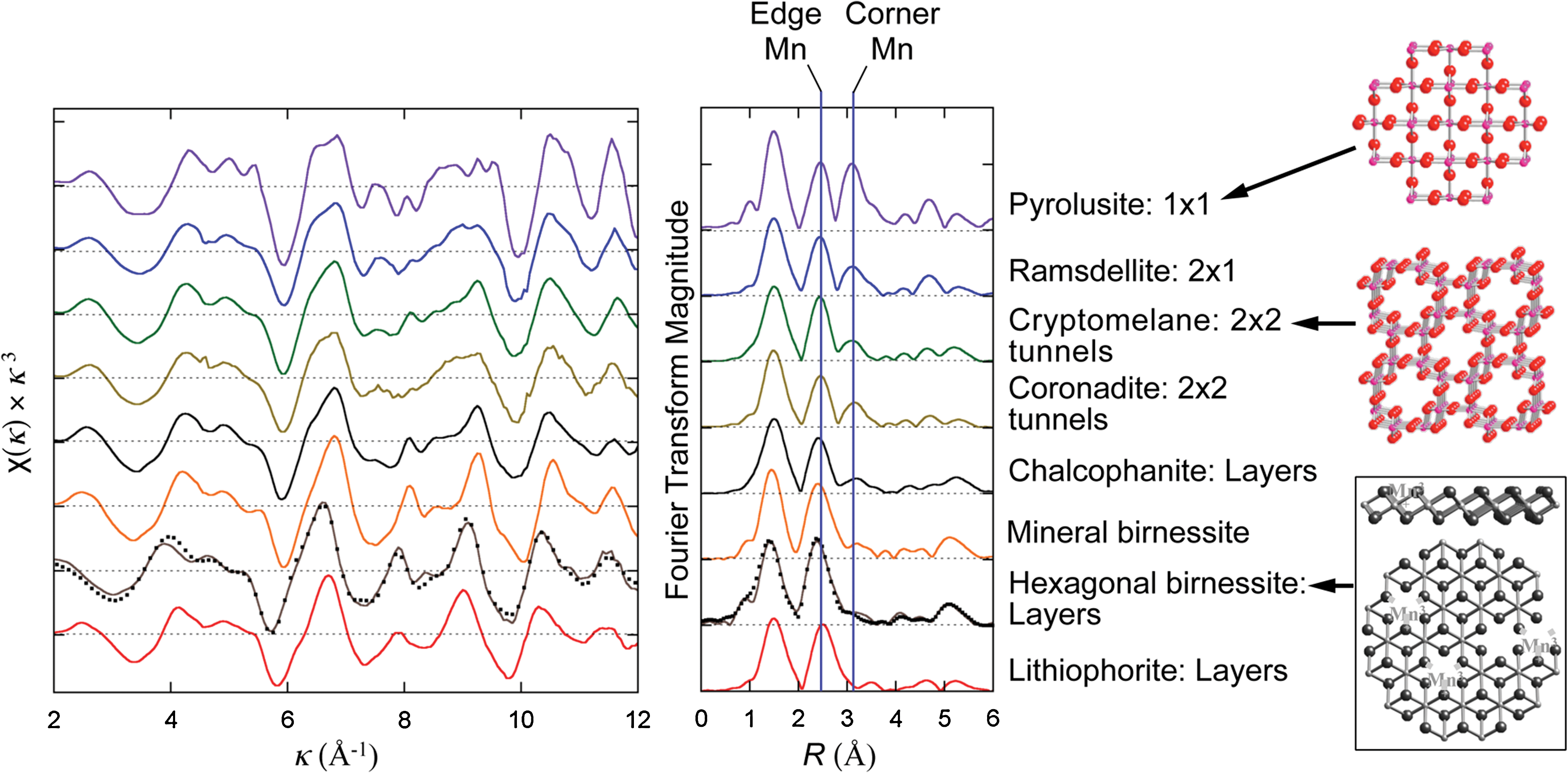
EXAFS spectra (left) and their Fourier transforms (middle) and representative structures (right) for a synthetic hexagonal birnessite and a series of natural Mn oxides obtained from a commercial source (Minerals Unlimited). Localities of the specimens are pyrolusite, Lake Valley, Sierra County, New Mexico; ramsdellite, Mohave Mining & Milling Co., manganite mine, near Wickenburg, Maricopa County, Arizona; cryptomelane, Reaphook Hill, Southeast of Blinman, South Australia, Australia; chalcophanite, in limonite (hydrous zinc iron manganese oxide) Tamera, Tunisia; birnessite, Clara Mine, Wolfach, Germany; lithiophorite, Postmaasburg, Griqualand West, South Africa. Color graphics available online at
2.2. Bacteriogenic Mn oxide
Bacteriogenic Mn oxide samples used in the present study were grown in laboratories under controlled conditions with pure cultures as listed in Table 1. The Mn oxide from Bacillus spore SG-1 was grown by the procedure described by Bargar et al. (2005). With the exception of the Bacillus strain isolated from rock fragments in an abandoned lead and zinc mine on Catalina Island, California, all strains are well-characterized Mn oxidizers. All strains were grown in batch culture on a shaker in an incubator at 25°C. The Bacillus strain from Catalina Island and Pseudomonas GB-1 were grown in K medium (van Waasbergen et al., 1993). Leptothrix SP-6 was grown in PYG medium (Adams and Ghiorse, 1985). Erythrobacter SD-21 was grown in commercially available Marine Broth 2216 (Difco) with an addition of 100 μM of MnSO4·4H2O prior to autoclaving. After the Mn oxides were formed, the cells were washed three times via centrifugation in a fresh salt solution. Leptothrix was washed in 1% NaCl. The other strains were washed in a marine-strength salt solution (per liter: 20 g NaCl, 150 mg CaCl2·2H2O, 500 mg KCl, 3 g MgCl2·6H2O). Then the cellular pellets were dried in a rotor-evaporator at 30°C and stored at room temperature before EPR characterization. Characterization by X-ray diffraction (XRD)/EXAFS of the bacterial Mn oxide samples has been reported, and references are listed in Table 1.
Hexagonal birnessite different from Manceau and Combes (1988) in its turbostratic stack layering and very high Mn(IV) contents.
Close to hexagonal H+-birnessite.
Collected from 10 m depth in a Mn nodule zone in Oneida Lake, New York.
Mid-Pacific deep-sea nodule field.
Desert varnish from Four Buttes, Mojave Desert, San Bernardino County, California (Potter and Rossman, 1979). FT-IR, Fourier transform infrared.
Collected from Mount Dunfee Summit, Nevada.
Villalobos et al., 2006, for Pseudomonas MnB-1, which is essentially identical to GB-1.
Jürgensen et al., 2004.
Villalobos et al., 2003.
This paper.
2.3. Synthetic Mn oxides
Triclinic birnessite, δ-MnO2, and hexagonal birnessite (acid birnessite) were synthesized in the laboratory by chemical reduction of KMnO4 solutions, the first two with suspensions of Mn(OH)2 and the third with concentrated, boiling HCl. Details of the procedures have been reported previously (Villalobos et al., 2003). The main difference between the synthesis procedures for the triclinic birnessite and δ-MnO2 was the ratio of Mn(VII)/Mn(II) used to yield Mn(IV); a stoichiometric ratio of 2/3 was used for δ-MnO2, whereas a 0.39 ratio was used for the triclinic birnessite. For the latter oxide, the resulting suspension was aged for one day at 55°C. After extensive washing with NaCl solution to obtain the corresponding Na-saturated Mn oxides and subsequent washings with nanopure water (18.2 MΩ-cm), all Mn oxide suspensions were dried at 60–65°C, and the powders were stored under dry conditions before use. X-ray characterization (XRD/EXAFS) of the synthetic Mn oxides has been reported (Villalobos et al., 2003, 2006), as listed in Table 2.
2.4. EPR experiment
The amount of sample used was typically 5∼50 mg, and the EPR data were obtained at room temperature (290 K) and at 77 K. A commercial EPR spectrometer operating at 9.3 GHz (Bruker 500E Elexys system) was used.
3. Results
3.1. EPR characterization
Electron paramagnetic resonance spectroscopy has been extensively employed to study transition metal ions with unpaired electrons in crystals and glasses for the characterization of oxidation states and symmetry of paramagnetic sites, as well as interactions with matrix environments (Abragam and Bleaney, 1970; Al'tshuler and Kozyrev, 1974). Without an applied magnetic field, the unpaired electrons with spin up or down states have the same energy. However, with an applied field, they split in energy (Zeeman interaction); and by microwave irradiation, transitions between the electronic spin states can be induced. When the magnetic field is scanned to the point that the energy difference (ΔE) between the spin states of the sample matches the microwave frequency quantum (hν), a resonance occurs, and the absorption of microwave quanta by the sample is detected as a signal. Each atom or ion with an unpaired spin(s) exhibits a characteristic magnetic resonance spectrum, and from the measurement of magnetic field strength (H 0) and microwave frequency (ν) at resonance, the spectroscopic splitting factor, g-value (g=hν/βH 0, in which h=Plank's constant, β=Bohr magneton), can be determined, and the electronic structure of the paramagnetic species can be characterized. For transition metal ions, the g-values deviate from the free electron value (2.0023) through spin-orbit interactions with the matrix environment.
Typical EPR spectra from Mn oxide samples are shown in Fig. 1 and plotted as the derivative of microwave absorption (at 9.3 GHz) versus applied magnetic field, H 0. The EPR measured parameters are summarized in Table 1. All the EPR data (at room temperature) are plotted in Fig. 3 as g-values versus spectral linewidths. Biogenic Mn oxides cluster around g=2.0 and linewidths, ΔH<560 G, whereas abiogenic Mn oxides have widely scattered g-values and much greater linewidths, ΔH>1200 G. Suspected biomineral samples of desert varnish and Mn nodules show moderate linewidths, 600 G<ΔH<1200 G. Also note that in Fig. 3 even the synthetic analogue to a bacterial counterpart, δ-MnO2 (Webb et al., 2005a) shows 4–5 times the linewidths of biogenic Mn oxides.
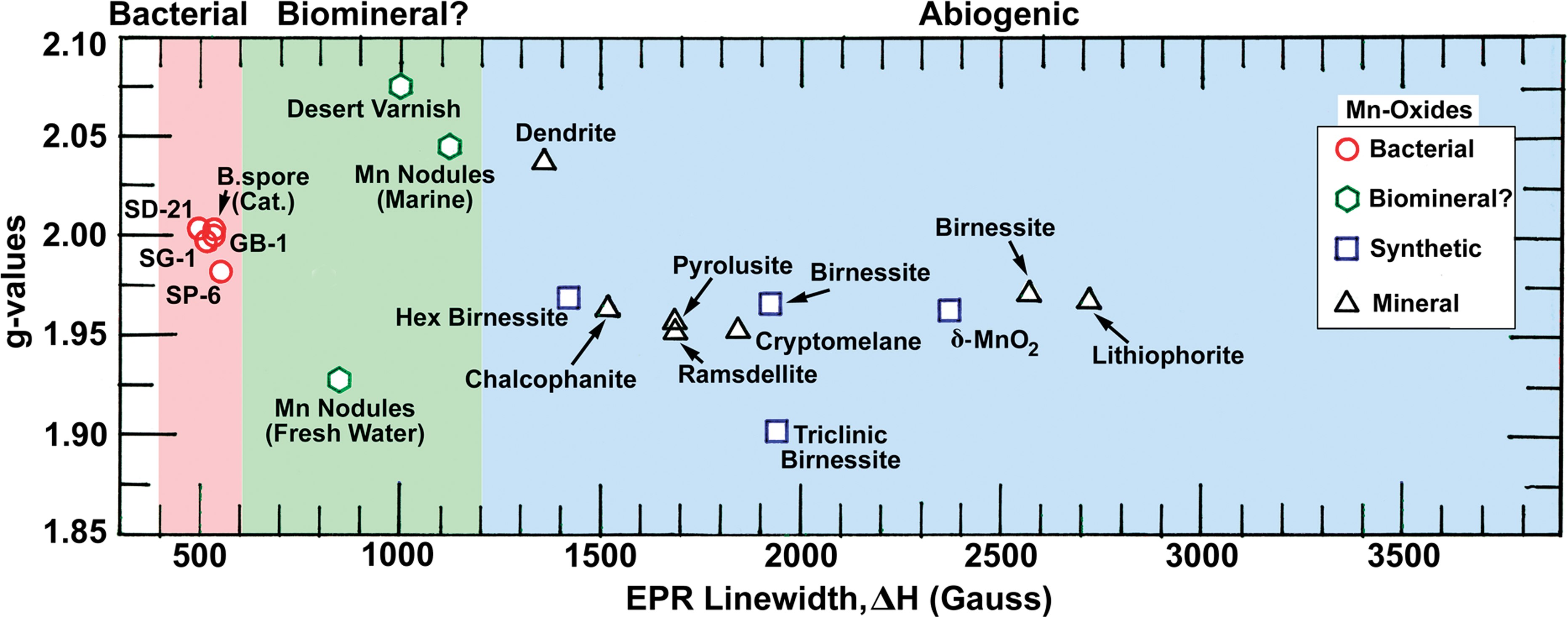
The EPR parameters (at 290 K) of all the Mn oxide samples studied are plotted as g-value versus spectral linewidth. The biogenic samples show a narrow distribution in g-values and linewidths, ΔH<560 G, whereas the synthetic as well as abiogenic mineral samples show a very wide distribution. Note the samples regarded as Mn oxide biominerals, Mn nodules, and desert varnish show narrower linewidths than the rest of the abiogenic mineral samples. Non-birnessite Mn oxides (chalcophanite, ramsdellite, pyrolusite, cryptomelane, lithiophorite) are included to demonstrate the range of ΔH values observed. Color graphics available online at
The narrow linewidths observed from bacterial MnO2 are very unique for a pure Mn oxide since comparable linewidths are only observed from highly diluted samples, for example, 0.5 wt % Mn ions supported on alumina (Kijlstra et al., 1997). Thus, among the pure Mn oxides, the linewidth criterion can be used to distinguish biogenic sources (red and green background in Fig. 3) from abiogenic ones (blue background in Fig. 3). Manganese oxides in the terrestrial weathering environment have been studied in the form of desert varnish (Potter and Rossman, 1977, 1979), a dark coating on rock in arid regions consisting of layer silicates, iron oxide, and Mn oxide. The manganese in desert varnish is present as a layered structure oxide identified as birnessite with an average Mn oxidation state that ranges from 3.8 to 3.9. Significant discussion has occurred about the probability that this manganese is a biomineral, but definitive proof has been lacking. Here again, EPR results suggest that desert varnish is of biomineral origin. Abiotic transformation of layered biogenic Mn oxides to todorokite (Tebo et al., 2005), as is thought to happen upon aging in Mn nodules, may occur with relatively minor modification to the original EPR biosignature. This may explain the relative proximity of EPR ΔH values for biogenic oxides and Mn nodules.
4. Discussion
4.1. EXAFS characterization
Bacteriogenic Mn oxides produced by Bacillus sp., Pseudomonas putida, and Leptothrix discophora have recently been shown to be nanolayered Mn oxides, which generally exhibit a defect-rich hexagonal birnessite structure (Jürgensen et al., 2004; Bargar et al., 2005; Webb et al., 2005a, 2005b; Villalobos et al., 2006) (Fig. 4). Key aspects of their structures that distinguish them from abiogenic counterparts include relatively low Mn(III) content (0–5%), high cation vacancies (15–50%), and poor or absent stacking.
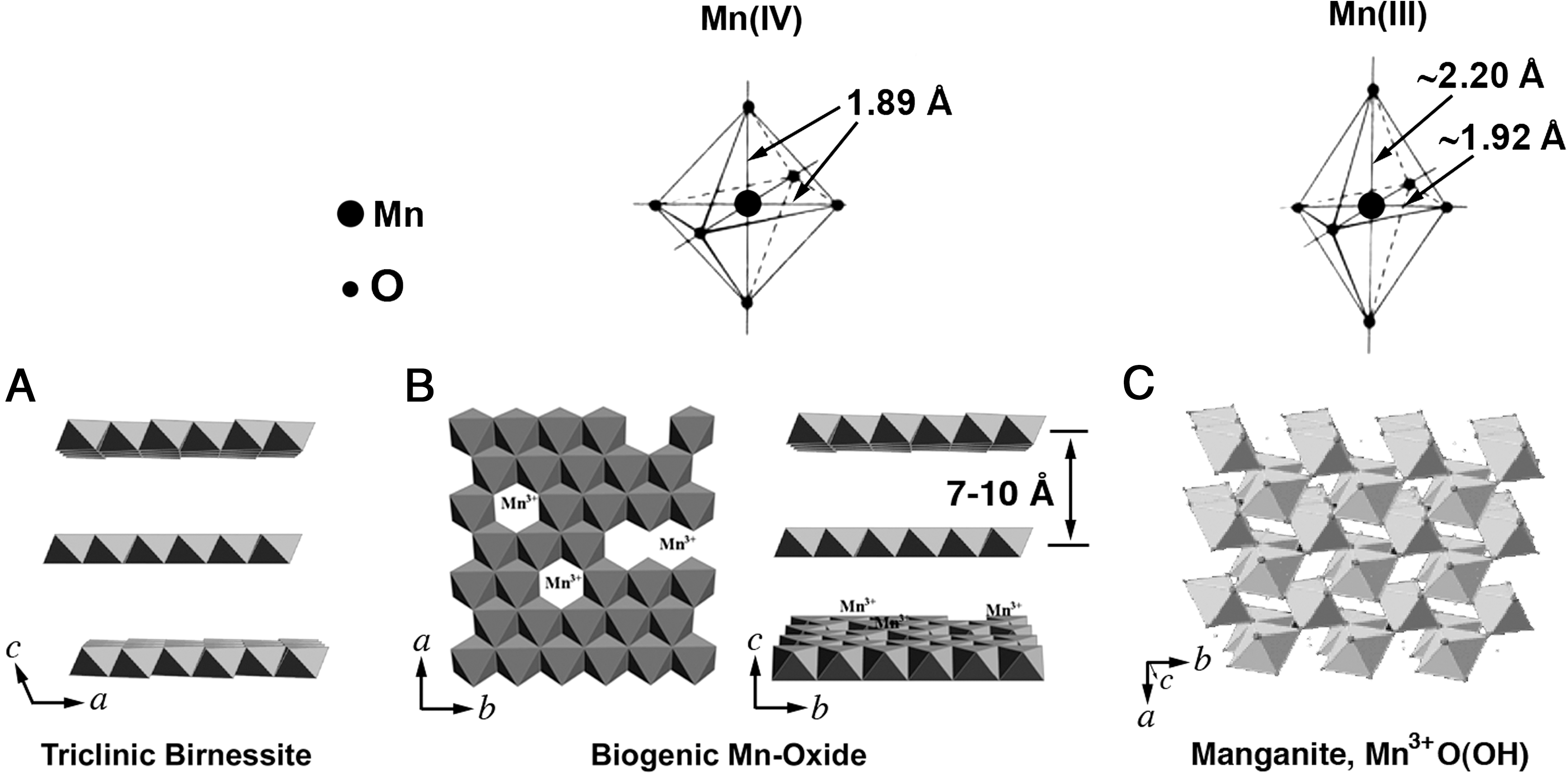
Polyhedral representation of key Mn oxides. (
In general, Mn(III) is a ubiquitous structural substituent in natural abiogenic birnessite (nominally defined as layered MnO2), where it can occupy up to one-third of all octahedral Mn sites (Lanson et al., 2002). At such high concentrations, Mn(III) is believed to order into rows parallel to the b crystallographic axis and thereby induce a loss of hexagonal symmetry of the Mn oxide layer, which results in a triclinic structure. A summary comparison of the X-ray structural data is provided in Table 2.
4.2. Structural basis of the EPR biosignature
Our EPR results with biogenic and abiogenic Mn oxide minerals match nicely with structural information obtained by X-ray absorption techniques. Paramagnetic ions act as magnetic dipoles, and the more of them that occur in proximity, the stronger they will interact, which results in magnetic dipolar broadened spectra (ΔH d). The importance of the dipolar contribution to observed EPR linewidth through interactions between neighboring Mn ions has been demonstrated in the case of alumina-supported Mn oxides (Mn/γ-Al2O3) used as catalysts, in which the Mn ions are dispersed on the surface of alumina as reported by Kijlstra et al. (1997). They observed an EPR linewidth of 580 G similar to the biogenic MnO2 at a Mn loading of 0.5 wt %, with gradual broadening as the concentration of surface Mn ions was increased. The uniqueness of bacterial Mn oxides resides in the fact that even though they are pure Mn oxides (100%), they can still show narrow linewidths comparable to a diluted counterpart. Highly dispersed Mn(II) will produce a distinct six-line pattern (Fig. 1) that can easily be recognized, and Mn(III) is most often not directly detectable by EPR at ambient temperature (Veridine, 1984). However, in mixed samples [i.e., Mn(III) and Mn(IV)], Mn(III) will influence the Mn(IV) spectra through exchange broadening.
All the Mn oxide samples we have studied thus far display a Lorentzian lineshape (Fig. 1), which is characteristic of contributions from exchange-coupled paramagnetic moments (Anderson and Weiss, 1953). In general, interactions between neighboring identical ionic species, for example, Mn(IV)-Mn(IV), result in exchange narrowing, ΔH e, whereas interactions between neighboring different species, for example, Mn(III)-Mn(IV), show exchange broadening, ΔH eb (Glaunsinger et al., 1979, 1980). The observed EPR linewidths are results of such contributions, that is, the static dipolar linewidths, ΔH d, are modified through the exchange interactions, ΔH e (Pake, 1962).
The relative contributions to the observed linewidth can be estimated by comparing the spectra taken at two temperatures, 290 and 77 K, of the same sample and using Eqs. 1 –5, as described below. The biogenic SG-1 MnO2 data are used as an example for the analyses, and results are summarized in Table 3.
Alfa Aestar, MnO, 99%.
Alfa Aestar, Puratonic, Mn(IV), 99.997%.
Samples contained freeze-dried bacterial cells, and the net Mn ion contents are estimated.
Lidiard, 1954.
Kakazey et al., 2001.
Kittel, 1951.
In his theoretical study of magnetic resonance lineshape, Van Vleck (1948) showed that the second moment contains only the static dipolar contribution, ΔH
d, and the fourth moment contains contribution from the exchange integral, J, or exchange magnetic field, ΔH
e. The observed EPR linewidths, ΔH, can be summarized for the exchange-narrowed case (Pake, 1962; Kittel, 1968; Kakazey et al., 2001) by
and the exchange-broadened case in the presence of Mn3+ ions (Kakazey et al., 2001) by
in which x=Mn3+/Mn4+ and k is a coefficient of broadening that can be determined through variation of Mn3+ concentrations in samples. Elucidation of the dependence of ΔH eb on the value of x of abiogenic Mn oxide samples will be deferred to a future effort.
From the fundamentals of EPR, the absorption of microwave power,
in which ω is the microwave frequency, H
1 is the microwave magnetic field, χ″ is the absorptive part of complex susceptibility, χ0 is the static susceptibility, and H
0 is the magnetic field at resonance. The static susceptibility for antiferromagnets can be estimated as follows:
in which N is number of ions per unit volume, β is the Bohr magneton, S is a spin quantum number, k is the Boltzmann constant, T
N is Neel temperature, and θ is Curie-Weiss temperature. As shown in Fig. 5 and Table 3, SG-1 MnO2 is still paramagnetic at 77 K (T
N<77 K), so one can apply Eqs. 3 and 4 for the ratio of EPR signal intensity,
in which ΔH(290)=520 G and ΔH(77)=538 G. The derived value of θ is 49 K for the SG-1 MnO2. The value of θ is in turn connected to the exchange integral J by the relation (Anderson and Weiss, 1953)
in which z is the number of nearest neighbors in the lattice. Once the values of θ are known, and assuming the z values are the same, J values (ΔH e) can be estimated by proportionality from Eq. 6. For example, the chemical MnO2 [θ=316 K, Lidiard (1954); ΔH=3200 G; ΔH d=4300 G, Kakazey et al. (2001); then ΔH e=5780 G from Eq. 1] and SG-1 MnO2, the ratio of θ is 6.44 (316/49). This applies to the J ratio; thus, for SG-1, the room temperature linewidth from exchange interaction can be obtained as ΔH e=896 G. The value of dipolar linewidth, ΔH d=683 G, was calculated from Eq. 1 and is listed in Table 3. As shown, SG-1 MnO2 has smaller linewidths; both dipolar as well as exchange, at the level of 16% of the chemical MnO2. This is consistent with the X-ray observations described below.
In the layered Mn oxides, including biogenic ones, the distance between nearest neighbor Mn ions (Mn-O-Mn distance) is r≈2.8 Å in the same layer. As shown in Fig. 4, the layers are separated by 7–10 Å, with the distance depending on the extent of hydration. The dipolar and exchange interactions are functions of distances between paramagnetic ions, with the former varying as r −3, and the latter requires overlap of wave functions to fall off more rapidly with distance (Pake, 1962). Thus, for the layered Mn oxides, the effects of dipolar and exchange interactions on the observed EPR linewidths are mostly limited to the same layer, with little influence from the neighboring layers. However, Mn oxides without such layer structures (Fig. 4), for example, manganite (monoclinic) or CaMnO3 (perovskite), would exhibit large exchange interactions throughout the solid matrices (Huber, 1998; Huber et al., 1999).
In general, biogenic Mn oxides are believed to exhibit hexagonal layer symmetry and nanolayered particle morphologies, with particle diameters as small as ca. 30 nm, and thicknesses ranging from 1 to 11 nm (Webb et al., 2005a, 2005b; Villalobos et al., 2006; Bargar et al., 2009) (Table 2). They are relatively poor in Mn(II) and Mn(III) (Villalobos et al., 2003) and exhibit remarkably high Mn(IV) layer site vacancies, approaching 20%. From their nickel sorption experiment, Peña et al. (2010) showed that 4% of the total Mn was present as Mn(II) in the biogenic hexagonal birnessite produced by Pseudomonas putida GB-1. Generally, Mn(II) is not detected in Mn K-edge X-ray absorption near-edge spectroscopy (XANES) spectra for the biogenic Mn oxides in similar studies (Bargar et al., 2005; Webb et al., 2005b; Villalobos et al., 2006). The detection limit for Mn K-edge XANES is ca. 2.5% of total Mn. Below this level, Mn(II) may be present but not detected. It is therefore reasonable to conclude that the Mn(II) content in layered biogenic Mn oxides is often in the range of a few percent of the total Mn present (Bargar et al., 2005). Edge terminations, Mn(IV) site vacancies, and edge sites are sufficiently abundant in biogenic Mn oxides that the majority of occupied Mn(IV) sites will reside in the immediate vicinity of a vacancy (or edge). This structural property should push EPR linewidths to narrower values, whereas synthetic Mn oxides with lower Mn vacancies, large particle diameters should exhibit broader linewidths. Thus, a significant EPR linewidth variation is expected from Mn oxides, depending on their underlying structures.
This behavior is observed in Fig. 3. The samples that cluster in the low-ΔH end of the plot are exclusively pure-culture biogenic Mn oxides. Hexagonal birnessite, a structural analogue for layered biogenic Mn oxides but with lower vacancy content (Villalobos et al., 2006), exhibits a 3-fold higher ΔH value. The δ-MnO2, which exhibits lower vacancy content (6%) than the biogenic Mn oxides or acid birnessite (ca. 17% and 12%, respectively), exhibits a yet-higher ΔH value, which suggests that the linewidth is particularly sensitive to vacancy content in layered MnO2. Triclinic birnessite exhibits a ΔH value about 4-fold higher than that of the pure-culture layered biogenic Mn oxides, as expected based on its relatively high Mn(III) content and low vacancy content. Mn oxide minerals containing significant structural Mn(II) (chalcophanite, ramsdellite, and cryptomelane) exhibit ΔH in the middle of the range, which is approximately 3 times higher than those of the pure-culture biogenic Mn oxides. Other samples (mineral birnessite, lithiophorite) also exhibit relatively large ΔH values, which demonstrates the sensitivity of the technique to local ionic environment and underscores its potential for use in terrestrial and extraterrestrial exploration as a biosignature.
In summary, Mn-oxide samples with few cation vacancies and mixed ionic states, that is, Mn(III) and Mn(IV), will show broader linewidths through larger dipolar interactions, ΔH d, and exchange broadening, ΔH eb, whereas biogenic MnO2 samples with holes in the layers and little Mn(III) content will have smaller dipolar interactions and moderate exchange narrowing, the combination leading to narrower observed linewidth. Therefore, because of the sensitivity of EPR to magnetic interactions between Mn neighbors, EPR ΔH values should depend on the presence of Mn(III)-Mn(IV), Mn(III) content, and cation vacancies of the solid phase.
In a natural environment, the vacancies of the bacterial Mn oxides, once formed, will gradually fill with various cations such as Fe3+ and/or may attain more layers with time. In the case of Mn nodules, sampling was done from different parts of the nodule, and a portion was used that gives the narrower linewidth, away from the broadening effect of contaminated Fe phases. However, most natural samples will also contain iron oxides to varying degrees, and quantification of the Fe influence by rock magnetic techniques will be elaborated elsewhere (Raub et al., unpublished data). In addition, biogenic layered Mn oxides may partially transform to tunnel structural motifs and/or acquire cations such as Ca2+ with aging in natural sediments (Bargar et al., 2009), which are expected to escalate EPR ΔH values. However, as desert varnish and Mn nodules show, they still maintain narrower EPR linewidths than their abiogenic counterparts (Fig. 3). This observation supports a biogenic origin for these Mn oxides.
4.3. Mn(II) contents in bacterial MnO2
Bargar et al. (2005) monitored the oxidation of Mn (II) by Bacillus spore SG-1 in situ with X-ray spectroscopic techniques that included K-edge XANES, X-ray emission spectroscopy, and in situ synchrotron radiation-based X-ray diffraction. They examined time-dependent changes in Mn oxidation states, local- and long-range structure in amorphous, crystalline, cell-bound, and solute Mn species. As shown in Fig. 5, the results displayed a decrease of intracellular Mn(II) from 100% at the start of incubation to below 2.5% of the total Mn content as the oxidation proceeded to produce MnO2.
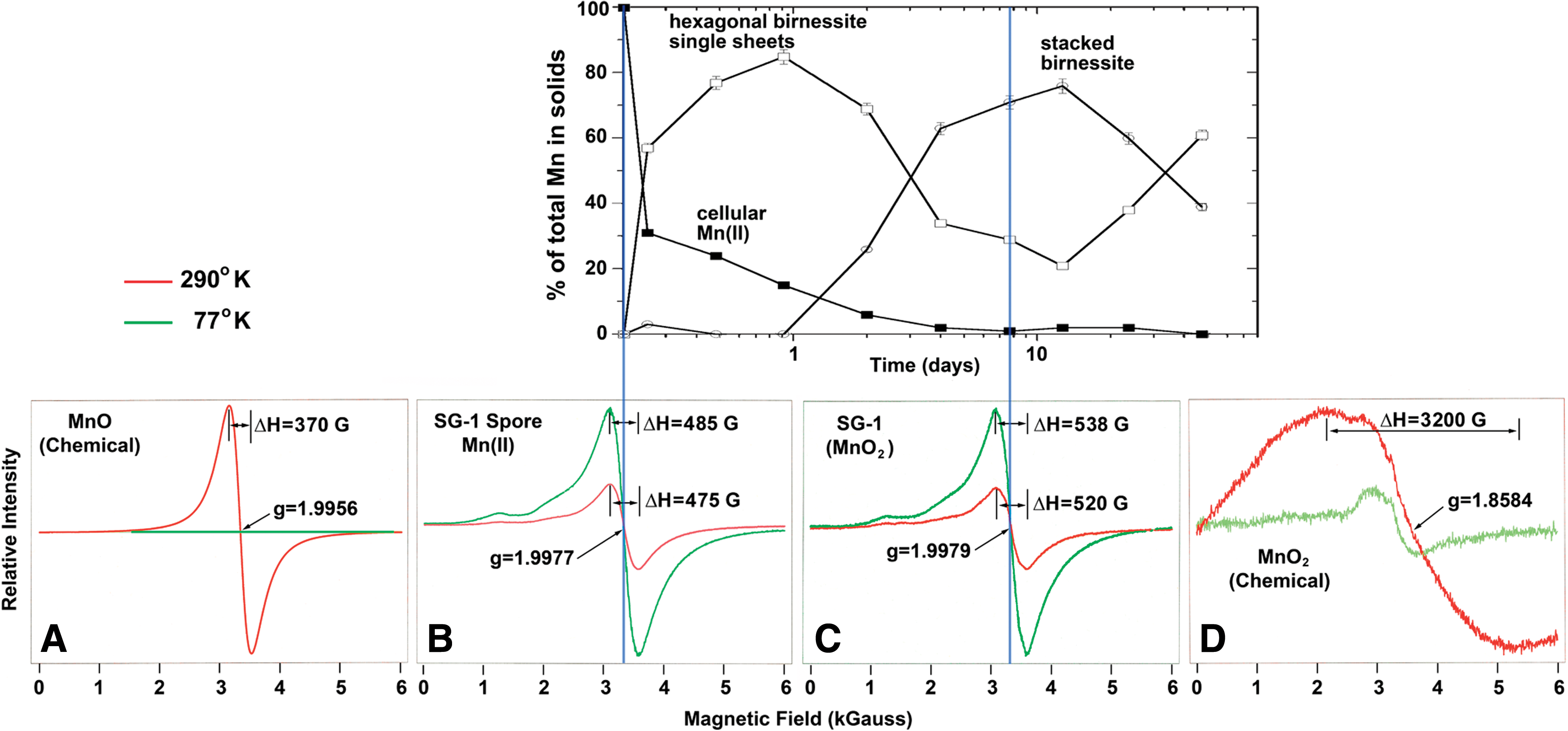
Top: the graph shows the result of in situ monitoring by XANES of the decrease of intracellular Mn(II) ions followed by the formation of phyllomanganates (Bargar et al., 2005). Bottom: EPR spectra of (
Figure 5 shows solid oxidation products obtained when 10 μM Mn(II) was oxidized by spores of Bacillus species (strain SG-1) in an aqueous medium containing 50 mM NaCl and 10 mM HEPES buffer (final pH=7.7–7.8). The solid fraction of these samples, initially just spores and subsequently spores encrusted in Mn oxides, were centrifuged and analyzed by XANES. Mn(II) is present in the initial time points because it occurs in the spores. As time progresses, Mn(IV) oxidation products accumulate in the solids, effectively swamping the Mn(II) initially present in the spores. The initial product is a nondiffracting hexagonal birnessite (single sheets). This phase reacts with Mn(II) in solution to produce a diffracting stacked hexagonal birnessite with a 10 Å basal plane spacing. After 4 days of reaction, the total Mn(II) content fell below the detection limit of 2.5% of total Mn.
In step with the XANES results, Fig. 5 also shows EPR spectra of freeze-dried SG-1 spores [starting point of the plot with 100% intracellular Mn(II), ΔH=475 G] and the harvested SG-1 MnO2 after incubation (ΔH=520 G), as well as chemical MnO (ΔH=370 G) and MnO2 (ΔH=3200 G) for a comparison.
4.4. Possible source of isolated Mn(II)
In Fig. 1, some of the bacterial Mn oxides show overlapped six-line hyperfine structures characteristic of highly diluted Mn(II), which persisted through additional washings. Quantitatively, the Mn(II) contribution is low, well within the range of 2.5% when compared on the basis of areas under the derivative spectra. Bacteria have a number of different enzymes that utilize manganese as a cofactor, and since bacterial cells were not removed from the MnO2, the observed Mn(II) signal could be a reflection of the presence of mononucleated enzymes, that is, isolated Mn(II) in protein manifolds of the enzymes. These include enzymes such as Mn-superoxide dismutase and a number of other metabolic enzymes that use Mn at their active sites. To this end, most bacteria that have been tested utilize a Mn transport system that transports Mn(II) into the cell, and it may well be that a low level of Mn(II) is a common feature of the cytoplasm of such bacteria (Nealson, 1983; Nealson et al., 1989). However, the low-temperature EPR signals show no indication of fine structure (Fig. 6) that excludes Mn(II)-Mn(II) pairs such as cofactors in enzymes like catalase or the sulfur oxidizing enzyme SoxB (Wu et al., 2004). In Fig. 1, the Erythrobacter SD-21 EPR spectra indicate the presence of Fe3+ at g=4.3. Erythrobacter SD-21 was grown in Marine Broth 2216, which has visible Fe-oxide granules. These granules are inseparable from the washed bacterial pellets and are most likely the source of the ferric signal.
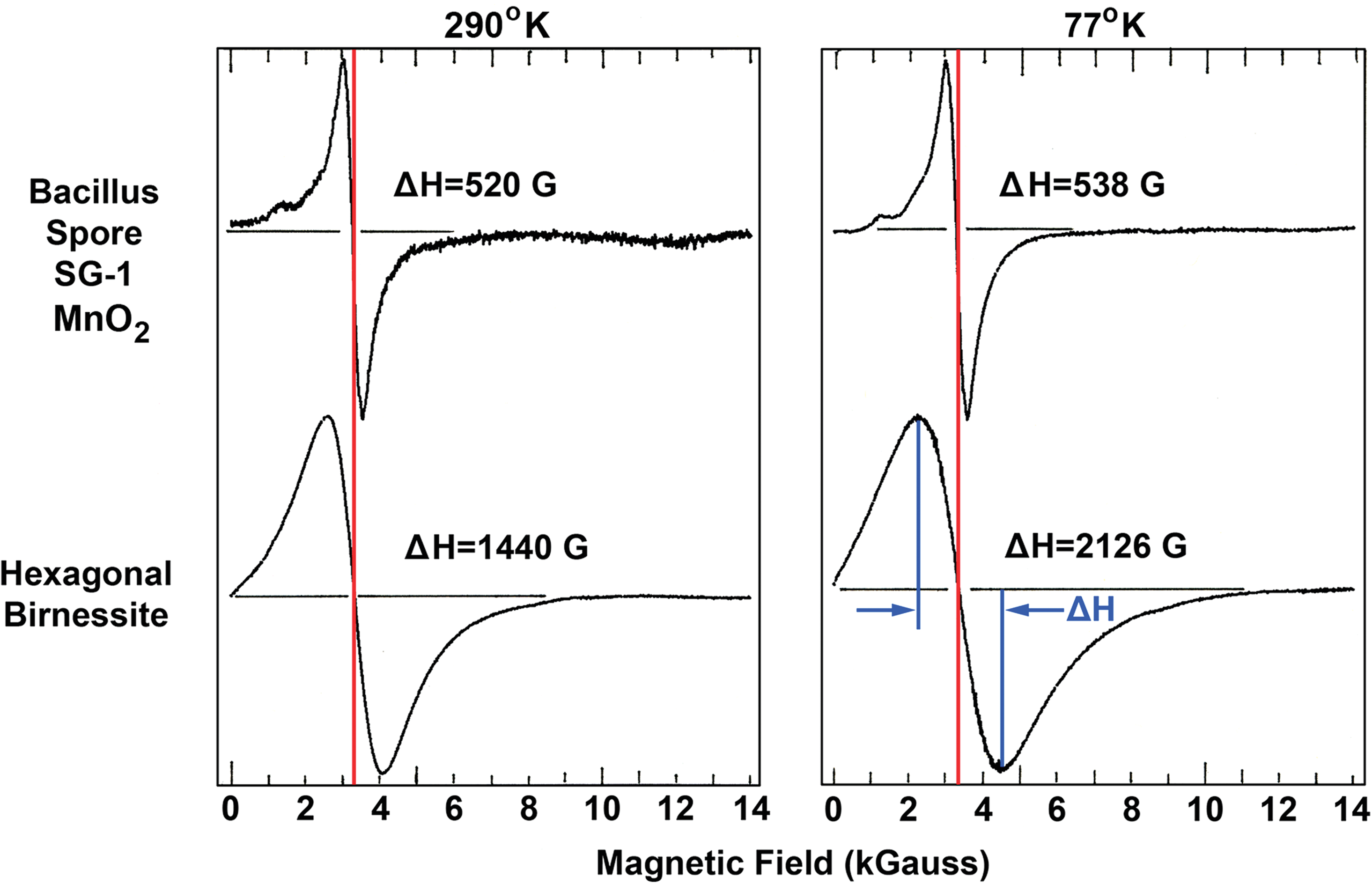
EPR spectra of Bacillus spore SG-1 MnO2 and synthetic analogue, hexagonal birnessite. Notice the increase of linewidths from room temperature to 77 K. Color graphics available online at
4.5. Case against Mn(II)-Mn(II) dimer
Harris (1972) summarized EPR observations of exchange-coupled Mn(II)-Mn(II) pairs in dilutely substituted Mn ions (1 atomic % Mn) in MgO and CaO crystalline matrices. The coupled Mn pairs can be described as a system with total spin of
5. Conclusion
Our EPR results are in good agreement with those from X-ray–based structural characterizations. Biogenic Mn oxides have mostly single ionic species, for example, Mn(IV), and more site vacancies than abiotic Mn oxides. These structural characteristics yield distinct EPR signatures, which can be used as a convenient means of prescreening biogenic Mn minerals against abiogenic counterparts.
Footnotes
Acknowledgments
S.S.K. is deeply grateful to George Rossman (Caltech) for the samples of desert varnish. M.V. is grateful to the IFS, project no. W/3912, for providing funds to synthesize and characterize synthetic Mn oxides. The research conducted by the Jet Propulsion Laboratory, California Institute of Technology was done under a contract with the National Aeronautics and Space Administration. Copyright 2011. All rights reserved.
Abbreviations
EPR, electron paramagnetic resonance; EXAFS, extended X-ray absorption fine structure; XANES, X-ray absorption near-edge spectroscopy; XRD, X-ray diffraction.