
Abstract
The iron-sulfur world (ISW) theory is an intriguing prediction regarding the origin of life on early Earth. It hypothesizes that life arose as a geochemical process from inorganic starting materials on the surface of sulfide minerals in the vicinity of deep-sea hot springs. During the last two decades, many experimental studies have been carried out on this topic, and some interesting results have been achieved. Among them, however, the processes of carbon/nitrogen fixation and biomolecular assembly on the mineral surface have received an inordinate amount of attention. To the present, an abiotic model for the oxidation-reduction of intermediates participating in metabolic pathways has been ignored. We examined the oxidation-reduction effect of a prebiotic FeS/S/FeS2 redox system on the interconversion between several pairs of α-hydroxy acids and α-keto acids (i.e., lactate/pyruvate, malate/oxaloacetate, and glycolate/glyoxylate). We found that, in the absence of FeS, elemental sulfur (S) oxidized α-hydroxy acids to form corresponding keto acids only at a temperature higher than its melting point (113°C); in the presence of FeS, such reactions occurred more efficiently through a coupled reaction mechanism, even at a temperature below the phase transition point of S. On the other hand, FeS was shown to have the capacity to reversibly reduce the keto acids. Such an oxidoreductase-like chemistry of the FeS/S/FeS2 redox system suggests that it can determine the redox homeostasis of metabolic intermediates in the early evolutionary phase of life. The results provide a possible pathway for the development of primordial redox biochemistry in the iron-sulfur world. Key Words: Iron-sulfur world—FeS/S/FeS2 redox system—Oxidoreductase-like chemistry. Astrobiology 11, 471–476.
1. Introduction
T
However, previous studies have mainly focused on the reduction and fixation of carbon/nitrogen compounds and the resulting biomolecular assembly (Blöchl et al., 1992; Brandes et al., 1998; Wächtershäuser, 2007). A geochemical model for the origin of reversible oxidation-reduction metabolisms, especially the oxidation subprocesses, is still a critically important unanswered question about the emergence of life. Molecular oxygen is recognized as a desirable candidate for the electron acceptor in oxidation reactions. But clues from ancient rocks have demonstrated that there was little or no oxygen on primordial Earth throughout the Archean (Kump, 2008). On the other hand, highly reactive reduced gases (e.g., H2S and H2) probably rendered oxidative species such as Fe3+ unavailable in the ISW. Elemental sulfur (S0) is an essential element for organisms and is grouped in the same chemical family with oxygen (with the same outer electron configuration). Sulfur could be easily accessible near hot springs due to the photolysis and precipitation of volcanic gas SO2 (Butterworth, 2004; Bekker et al., 2009), the disproportionation of sulfides (Cody et al., 2000), or the electrochemical oxidation of S2− by submarine hydrothermal chimneys (Nakamura et al., 2010). Could elemental sulfur then have functioned as an electron sink for the prebiotic oxidation processes?
To assess this possibility, we experimentally examined the oxidation effect of elemental sulfur on three key biochemical intermediates (i.e., lactate, malate, and glycolate) under anaerobic conditions and in the presence or absence of FeS. In reverse, the reduction of pyruvate and oxaloacetate by FeS has also been investigated.
2. Materials and Methods
2.1. Materials
Keto acid and hydroxy acid sodium salts (≥99%, Aldrich) were used without further purification. All other chemicals were obtained in analytical grade (Sinopharm Chemical Reagent Co. Ltd, China), with the exception of methanol (chromatographic grade) and fine sulfur powder (100 mesh, 99.999%). Ultrapure water (Millipore) was deoxygenated with high-purity argon gas (Ar, 99.999%) by bubbling it (approximately 1 L/min) through 500 mL water for 1 h before used.
2.2. Experimental procedure
All experiments were conducted in 10 mL headspace vials fitted with metal caps and Teflon septa. Batch experiments were prepared anaerobically inside a vacuum glove box kept under an Ar atmosphere. In a typical run, a vial was charged with 1 mmol FeSO4·7H2O, 100 μmol keto acid salts (or 200 μmol hydroxy acid salts +5 mmol S), and 3 mL of deaerated water. Subsequently, a solution of 1 mmol Na2S·9H2O in 2 mL water was added for in situ precipitation of FeS. For the H2S-bearing experiments, additional 0.5 mmol Na2S·9H2O and 0.5 mmol H2SO4 were surcharged for in situ generating H2S. The sample pH values were adjusted as required with small volumes of 0.1 and 1 M NaOH. To ensure the anaerobic environment in the oxidation experiments, the vials were enclosed by using hydrothermal reaction vessels in the glove box. All reactions were carried out at 100°C or 120°C in an oven.
After the reaction, all samples containing sulfides were centrifuged and filtered by using 0.22 μm microvoid filter films. High-performance liquid chromatography (HPLC) was used to determine the concentrations of keto acids (Petrarulo et al., 1990) and hydroxy acids (Gao et al., 2004) with a C18 column (Phenomenex, Luna, 5 μm, 250×4.6 mm). The gas samples were analyzed by gas chromatography (detection limit 0.01 μmol) (Cao et al., 2010).
3. Results
In Table 1, it can be seen that small amounts of pyruvate, oxaloacetate, and glyoxylate were evolved from lactate, malate, and glycolate, respectively, within 5 h at 120°C by using elemental sulfur as the oxidizing agent (runs 2, 10, and 13), while at 100°C no traces of keto acids were detected (runs 1, 9, and 12). Blank tests at 120°C revealed that no keto acid formed in the absence of elemental sulfur.
In each run samples were run in triplicate; shown are mean values.
Numbers in parentheses are standard deviations of the mean.
Undetectable.
Not adjusted.
Such a temperature effect can be explained by a thermodynamic consideration. For example, the oxidation of lactate to pyruvate accompanying the elemental sulfur (S0) reduction is shown below:
where the positive ΔG 0 value, discerned by relying on the standard free energies of formation (Table 2), indicates that it favors pyruvate reduction over lactate oxidation. Above its melting point (113°C), however, the elemental sulfur will partly transform into several liquid forms with Gibbs free energies >25 kJ/mol. This change will reverse Eq. 1 from endergonic to exergonic and make it thermodynamically feasible, resulting in the formation of pyruvate. As circumstantial evidence, we found that the sulfur powder melted completely to form a saponaceous globule in the reaction at 120°C, and there was a faint H2S odor with a drastic pH decline in the solution. At 100°C no such changes were noted.
The physical state of each substance is indicated as crystalline solid (c), liquid (l), gaseous (g), and aqueous (aq).
Besides the above-mentioned phase transition strategy, a more moderate, alternative approach to fine-tuning the hydroxy acid oxidation can be applied by utilizing the waste heat from the “first energy source” (Wächtershäuser, 1988; Kalapos, 2002):
If Eq. 2 is coupled with Reaction 1, then the following exergonic process occurs:
The coupled Reaction 3 is in tactical agreement with the knowledge of extant biochemistry (Ould-Moulaye et al., 1999). A coupled reaction system is two reactions that occur together, in which one is necessary for the other to take place. Part of the chemical energy unleashed in the exergonic half-reaction will be collected by the endergonic one and can be consumed or conserved within the system. By stoichiometrically consuming the H2S produced in Eq. 1, the coupled Reaction 3 will go forward to pyruvate formation, indirectly overcoming the thermodynamic barrier.
This scheme was demonstrated in our experiments. With regard to the results presented in Table 1, elemental sulfur or FeS alone had no influence on the lactate oxidation at 100°C (runs 1 and 3); while, in conjunction with each other, solid FeS and S could heterogeneously oxidize lactate under the same temperature, with no need for the high-temperature phase transition of elemental sulfur (run 4). The coupled reaction not only lowered the threshold temperature for the oxidation process but also enhanced the yields of keto acids by several times. It was also observed that in the coupled reaction the formation of pyruvate at 100°C (run 4) was more than twice as efficient as that at 120°C (run 6) and was slightly improved by prolonging reaction time (run 5). The yield decline at higher temperature may be ascribed to the high-temperature thermolysis of the product. Thus, although the two temperatures are not totally representative in the ISW, it can be concluded that the oxidation process could take place in a moderate local environment (≤100°C), which would alleviate the instability of organic molecules at high temperatures near certain hydrothermal vents.
Meanwhile, we found that the formation of pyruvate was significantly enhanced in the presence of additional H2S (run 4) but seriously inhibited under basic conditions (run 8). A thorough explanation for the differences remains an open question. We presume the reaction is H2S dependent, since the excess H2S can participate in the exothermic process (2) and spurs Reaction 1 by releasing waste energy. Another possible speculation is that the reaction is pH related, that is, the influence of pH on the ionization state of lactate and the particular details of molecular scaffolding on the surface. At low pH, the protonated lactate (pK a=3.86) is more accessible to the hydrophobic surfaces of FeS and S, and more easily oxidized. Under basic conditions, besides the hydrophobic interactions, the surface modification of FeS by OH− (Butler and Hayes, 1998) also makes the lactate oxidation inefficient. A pH-dependent equilibrium between the hydroxylated and dehydroxylated forms of FeS surface, with the dehydroxylated form having greater reactivity, is partly responsible for the low yields of pyruvate at high pH.
The same phenomenon has also been observed to malate and glycolate (runs 9–14), which favors the suggested coupled reaction mechanism (see Supplementary Figs. S1–S3; Supplementary data are available online at
In order to further understand the above observations, we investigated the linkage between lactate oxidation and hydrogen evolution (Eq. 3). Typical results are shown in Table 3. Only in the simultaneous presence of FeS, S, and lactate could molecular hydrogen be formed. A complete set of control experiments unequivocally established that the heterogeneous coupled Reaction 3 was the mechanism for the improved hydroxy acid oxidation. The observed yield discrepancies between pyruvate (Table 1) and hydrogen (Table 3) may have been caused by the aqueous solubility of the newly evolved hydrogen, which was about several μmol in 5 mL sealed water at 100°C (Pray et al., 1952).
The averages and the standard deviations of the H2 yields of three repeats.
Undetectable.
Not adjusted.
On the other hand, FeS has been demonstrated to have the capacity to act as a fine reducing agent in the prebiotic synthesis (Wächtershäuser, 2007). It was calculated that the FeS/FeS2 redox pair had a pH-dependent reduction potential in the range from −200 to −760 mV (Kaschke et al., 1994). In comparison, the reduction potentials of
4. Discussion and Conclusions
The chemical feasibility of the FeS/S/FeS2 redox system in promoting the interconversion of α-hydroxy acids and α-keto acids was experimentally confirmed. The results suggest a possible redox pathway for the primordial metabolism before the embryonic emergence of primal life. The experimental data show that basic conditions are not conducive to the reversible redox reactions. However, the submarine seepage in the ISW was indeed suggested to be ultramafic and the resulting exhalations thereby alkaline (Russell and Hall, 1997; Martin and Russell, 2007). Thus, at first sight the application of the FeS/S/H2S redox system would seem to strongly conflict with this model of prebiotic chemical evolution. However, according to Russell and his coworkers (Russell and Hall, 1997; Nitschke and Russell, 2009), the requirement of neutral to acidic conditions could have been reached in an inorganic FeS membrane since protons from the ocean could be drawn to the alkaline interior of the iron sulfur vesicles by a chemiosmotic potential or a proton motive force between the two sides of the membrane.
It has been frequently argued that life's emergence can be postulated as a continuum from Earth's inorganic geochemical processes to chemoautotrophic biochemical processes (Russell and Hall, 1997; Russell and Martin, 2004; Wächtershäuser, 2007). However, how prebiotic metabolic systems could have evolved from sulfide surface remains a challenge. The reverse Krebs cycle (RKC, see Fig. 1), which is employed by a number of extant chemoautotrophs, has been suggested to be the center pathway for the primordial carbon fixation (Wächtershäuser, 1990; Morowitz et al., 2000; Zhang and Martin, 2006; Guzman and Martin, 2009). Although the reduction of CO2 and CO by FeS was successfully demonstrated (Heinen and Lauwers, 1996; Wächtershäuser, 2007), the abiotic synthesis of key metabolic intermediates in the RKC has so far failed. In a pioneering effort, Cody et al. (2000) developed a heuristic process for pyruvate synthesis. However, their reaction conditions, that is, 250°C and 200 MPa, are thought not to be the real case in the authentic ISW (Wächtershäuser, 2000). The excellent semiconductor photocatalysis hypothesis proposed by Martin and his colleagues (Zhang et al., 2004; Zhang and Martin, 2006) is also confronted with the same predicament, since there is no UV light available in the submarine ISW. It has been argued that formaldehyde was an important precursor available for the prebiotic syntheses (Pinto et al., 1980; Shapiro, 1988; Kasting and Catling, 2003). Thus, following up the synthesis of lactate (Bhattacharyya et al., 1970; Kalapos, 1997) and glycolate (Bhattacharyya and Vir, 1957; Dicosimo et al., 2008) from formaldehyde, the oxidation reaction model suggested in this study could have served as an anaplerotic pathway for the antique RKC by generating pyruvate and glyoxylate (Fig. 1) (Owen et al., 2002).
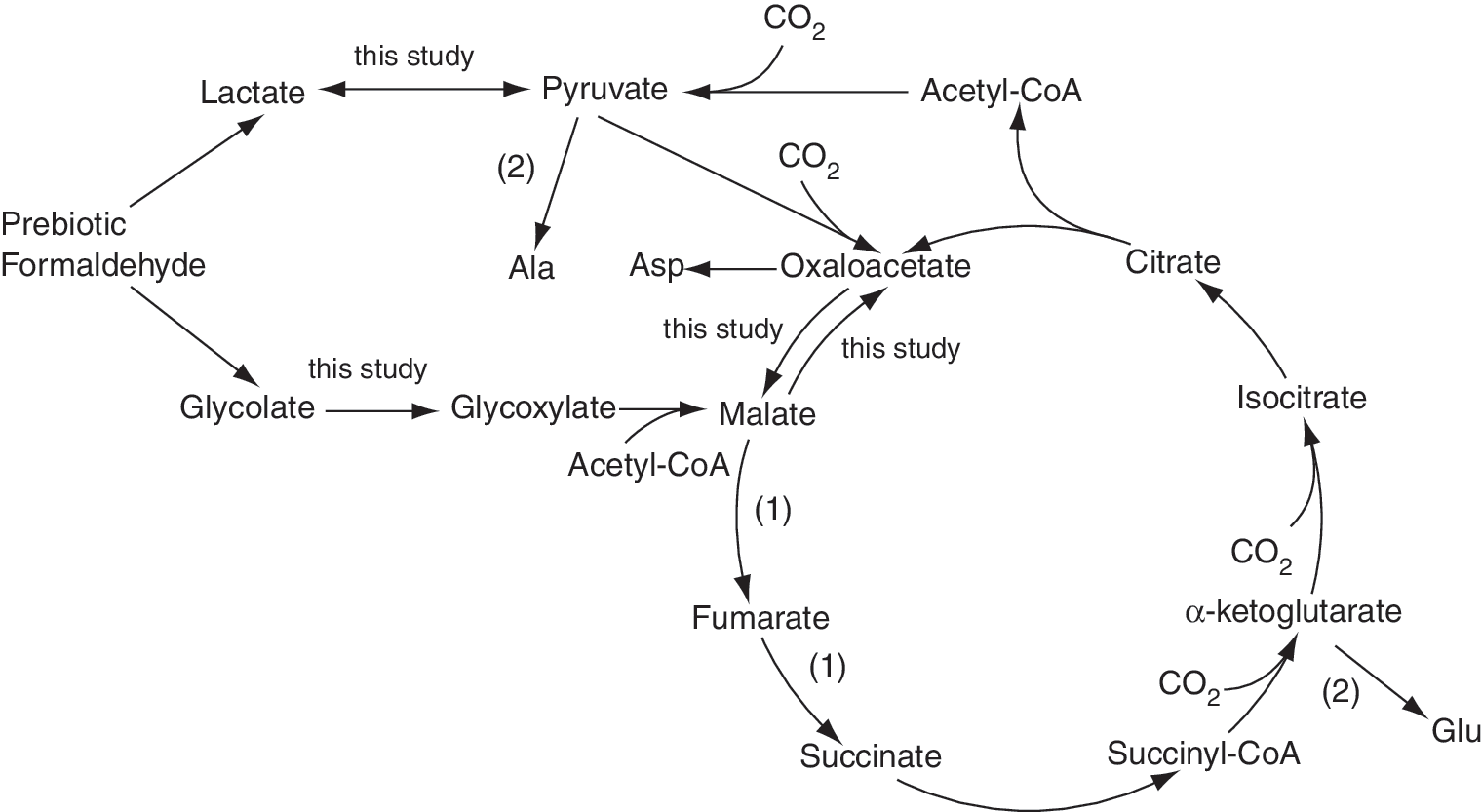
A cartoon of the reverse Krebs cycle. The FeS/S/FeS2 redox system could enhance anaplerosis and balance the entry and exit to the antique cycle via its reversible oxidoreductase-like chemistry. For the simulated prebiotic processes (1) and (2), please refer to Blöchl et al. (1992) and Huber and Wächtershäuser (2003), respectively.
As is well known, two fundamental properties of life are anaplerosis and redox homeostasis. The results presented in this study show that the FeS/S/FeS2 redox system possesses such characteristics. In view of its oxidoreductase-like chemistry, we suggest that the archaic geochemical FeS/S/FeS2 redox unit could have served as a redox catalyst for the primordial metabolism. Later, in the evolutionary context, the FeS/S/FeS2 redox system would have reacted with prebiotic peptides and finally evolved into extant Fe-S proteins, whose Fe-S cluster active centers, acting as a coenzyme, are strikingly similar to the iron sulfide minerals in tiny structure (Eck and Dayhoff, 1966; Russell and Hall, 1997; Milner-White and Russell, 2008). However, both the cofactors of extant lactate dehydrogenase and malate dehydrogenase, which catalyze the interconversion of
Footnotes
Acknowledgments
We greatly appreciate the constructive suggestions made by the anonymous referees. This work was funded by NSFC (40902014), CPSF (20080430918), and HIT.NSRIF (200811).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
HPLC, high-performance liquid chromatography; ISW, iron-sulfur world; NAD(H), nicotinamide adenine dinucleotide; RKC, reverse Krebs cycle.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.