
Abstract

“Membranes Came First!”
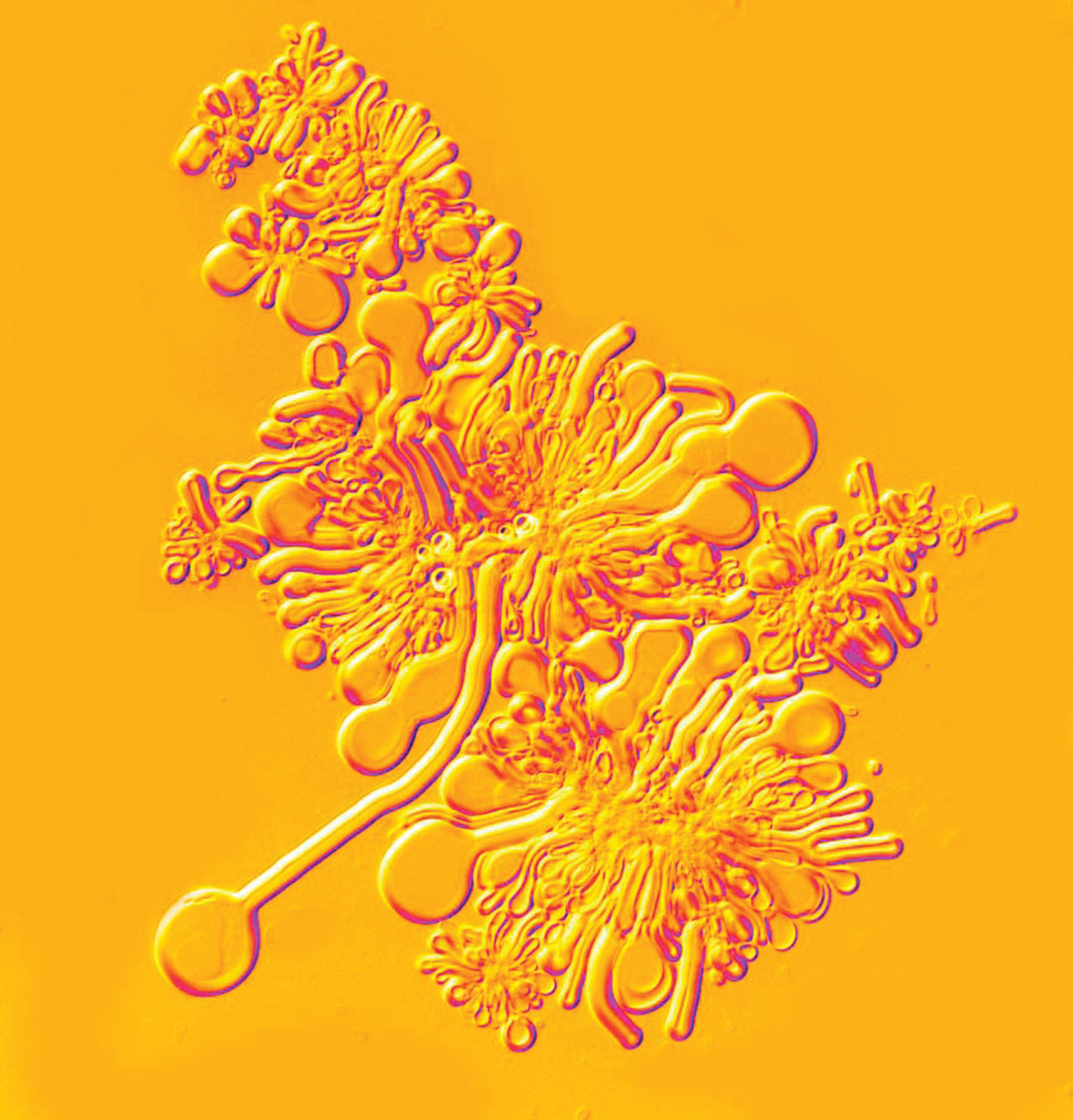
Dry phospholipid assembles into highly organized multilamellar structures upon exposure to aqueous solutions. Color images available online at
Alec also found that the boundary membranes of the vesicles were composed of lipid bilayers that could maintain concentration gradients of potassium or sodium ions. This was the first direct evidence that lipid bilayers were the primary barrier to the free diffusion of ions across cell membranes, a property now known to be essential for all living cells.
One summer day Alec and I were driving to London in his beloved Morris Mini and had pulled over to enjoy some cucumber sandwiches prepared by Rosalind Bangham for our journey. The conversation turned to a lecture he had given a few years earlier at Bristol University, with the title “Membranes Came First!” In his talk, Alec argued that the self-assembly properties of lipids were such that membranous vesicles must have been present on early Earth long before nucleic acids and proteins appeared. But then we were stumped. What was the prebiotic equivalent of lipids, and where did they come from? No one knew.
Meanwhile, back in Davis…
I took that question home with me when I returned to the University of California, Davis. Will Hargreaves had just joined my lab group, and I posed the question to him as a possible topic for his doctoral research. Will was virtually unique in his ability to be inspired by a challenging question. Over the course of the next few years, Will showed that almost any amphiphilic molecule could form membranes, either by itself or in mixtures with other amphiphiles. Will was able to produce vesicles from molecules as simple as fatty acids, which are little more than hydrocarbon chains with a carboxyl group at one end. Hydrocarbon chains with phosphate or sulfate end groups also worked. Mixtures were often better at self-assembly than pure compounds. For instance, neither dodecanol nor dodecyl sulfate form membranes as pure compounds, but together they produced amazingly stable vesicles. One day I was in the lab when Will burst out laughing while looking at a slide with our microscope. I came over to see what was funny, and he invited me to have a look. The slide was filled with countless numbers of beautiful vesicles, as nice as any we had seen. “What is this stuff?” I asked. Will replied, “Shampoo!” Puzzled, I checked the fine print on the bottle, and the mystery was solved. The main ingredient was ammonium dodecyl sulfate mixed in with some long-chain alcohols and amines.
We didn't publish the shampoo images, but Will was first author on papers in Nature (Hargreaves et al., 1977) and Biochemistry (Hargreaves and Deamer, 1978) in which we showed how membranous vesicles could form under simulated prebiotic conditions, fulfilling Alec Bangham's prophetic claim that membranes came first. Those papers apparently caught the attention of someone on the panel that reviewed grant proposals for the NASA Exobiology program, and a couple of years later I was invited to become a member of the panel. This is a bit of advice I have for young investigators: When you are offered membership on a peer review panel, take it! For one thing, you will meet some very smart and talented colleagues. My first panel was chaired by Harold “Chuck” Klein, who was the Biology Team Leader for the Viking Mars landers. A young rambunctious guy named Steve Squyres was also on the panel. Thirty years later, Steve is the principal investigator of the current Mars rover program, and it's fun to see an older rambunctious guy on television, enthusiastically explaining the latest news from Mars. Another reason to be on a peer review panel is that nothing is better preparation for writing competitive grant proposals than seeing firsthand how proposals are reviewed. It can be disconcerting to discover that a certain amount of luck is involved in getting a proposal funded, but you also learn how to present your ideas clearly, how to argue that they are significant and, even more important, how to avoid offending potential reviewers. From my service on the panel, I realized that NASA was a source of funding for just the kind of research I wanted to do, so I wrote my first grant in 1981, it was approved in 1982, and my lab has been funded ever since by the NASA Exobiology and Astrobiology programs.
Membranes and Meteorites
As I became more involved in origins research, I discovered that quite a few scientists at NASA Ames shared my interests. In particular, Sherwood Chang was an expert on the organic chemistry of meteorites. Sherwood invited me to give a talk at Ames, so I made the 2-hour drive from Davis to Moffett Field in Mountain View where NASA had established enormous wind tunnels to test future space shuttles. They also constructed Building 239, covered with concrete decoratively “cratered” to mimic the lunar surface, where scientists working in the exotic field of exobiology had their laboratories. In my talk I described the self-assembly properties of lipids and how membranous vesicles were likely to have been around on early Earth. By that time we had also solved another problem. Membranes are virtually impermeable to large molecules, so how could the precursors of proteins and nucleic acids get into vesicles? The answer was surprisingly simple and years later led to the research I am doing right now. We needed to go beyond thinking that early Earth was just a giant test tube filled with seawater in which life was generated by chemical reactions in solution. Instead, there were multiple interfaces between volcanic landmasses resembling Hawaii and Iceland today, in contact with the atmosphere and liquid water. Furthermore, the interfaces were not at equilibrium but were undergoing cycles of wetting and drying imposed by tides or precipitation. We found that if lipid vesicles were put through a wet-dry cycle, the dry lipid fused into multilamellar structures. Furthermore, anything that was present in the original solution was trapped between the lipid layers, including molecules as large as enzymes and nucleic acids. If we added water back to the dry lipid, vesicles formed again but now up to half the material originally outside was encapsulated in vesicles. The result was protocells, trillions of microscopic compartments per milligram of lipid, each different from all the rest, and each representing a kind of natural experiment. I speculated that if one of the protocells happened to contain a system of molecules that could capture energy and nutrients to make polymers, that happenstance would be the first step toward the origin of life.
But the question remained: what were the first lipids? About 10 years before my talk at Ames, Keith Kvenvolden was working with a team of Ames scientists (Kvenvolden et al., 1971) and showed that there was a suite of amino acids in a carbonaceous meteorite that had exploded in the sky over the little town of Murchison, Australia, in September 1969. A shower of meteoritic fragments scattered over perhaps 5 square miles, and within a week hundreds of stones were recovered, many of which reside in museums around the world. The discovery of amino acids in the Murchison meteorite was as revolutionary as Stanley Miller's synthesis of amino acids in a simulated prebiotic atmosphere. There was now no doubt that biologically relevant organic compounds were synthesized in the early Solar System. It was even possible to think that life began from a suite of such compounds delivered to Earth by comets and meteorites, an idea that had been put forward by John Oro in 1960 (Oro, 1961).
It occurred to me that if carbonaceous meteorites contained amino acids, lipid-like compounds might also be present. I asked Sherwood Chang if he had a meteorite sample I could use, and he was happy to provide a gram of powder produced from the interior of the Murchison meteorite by grinding under ultraclean conditions. Lipids are defined by the fact that they are soluble in organic solvents. I had often used a mixture of chloroform and methanol to extract lipids from liver tissue, blood cells, egg yolks, and spinach leaves, so I added a small amount of the standard chloroform-methanol solvent to the Murchison powder, stirred, and then centrifuged it to remove the mineral particles that compose most of the meteorite. I placed a drop of the faintly yellow solution onto a microscope slide and let it dry. From the obvious spot that remained on the slide, it was apparent that the solvent had dissolved something in the powder. I also noticed a strange odor coming from the slide. It was simultaneously earthy, dusty, slightly sour, and I realized that I was smelling organic compounds that were at least 4.5 billion years old, probably older than Earth itself. I added a drop of dilute buffer, put a cover slip over the wet spot, and placed the slide on the microscope stage.
Figure 2 shows what I saw that day. It was a stunning image. All around the edges of the spot I could see vesicles forming, often at the end of threadlike strings, and then being released into the buffer solution. We could now add membrane-forming molecules to the growing list of biologically relevant organic compounds that were present in the parent body of the Murchison meteorite.
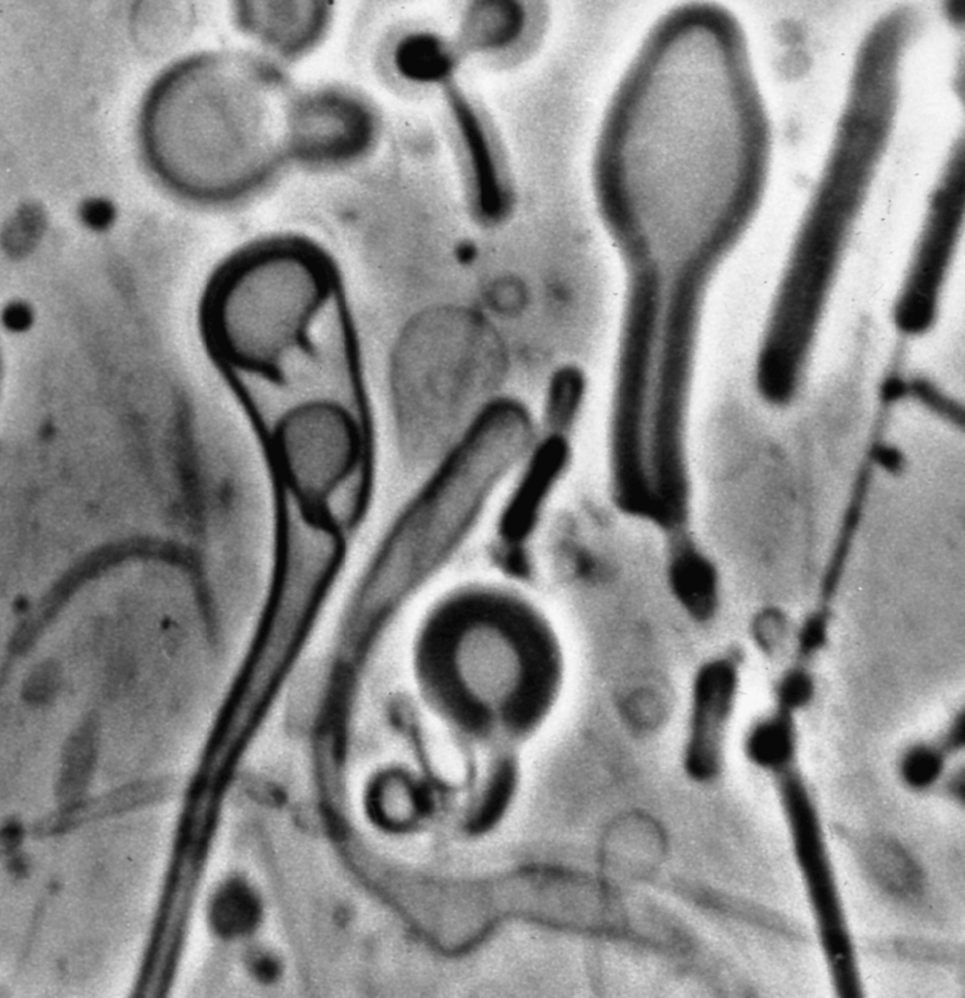
Amphiphilic substances extracted from the Murchison meteorite self-assemble into membranous vesicles.
I published this finding in Nature (Deamer, 1985) and spent the next few years working out what was in Murchison that could produce self-assembled membranous vesicles. We knew from Will Hargreaves' results that even simple amphiphilic molecules like fatty acids could make vesicles, and we also knew from a later analysis of Murchison organics by Jim Lawless that monocarboxylic acids up to 8 carbons in length were present (Lawless and Yue, 1979). We were pretty sure that 8-carbon chains were not long enough to form stable membranes, but Jim had worked with a total extract rather than using a solvent system that would specifically extract longer-chain molecules. I took a sample of our chloroform extract over to Dick Zare's lab at Stanford, where it was derivatized and run on his mass spectrometer. We had guessed right. Peaks were there for monocarboxylic acids up to 11 carbons long, and these can assemble into membranous vesicles.
To sum up, the first forms of life did not have phospholipids available to make their membranes, because phospholipids are products of complex metabolic pathways in living cells today. Instead, it seems reasonable to think that a variety of simpler amphiphilic molecules could assemble into vesicles on early Earth, and that replicating, catalytic polymers would have a membranous home waiting for them to form the first cells. While this story was developing in my lab in the early 1990s, Luigi Luisi and his research group were doing similar work at the ETH in Zurich. I met Luigi in 1993 at a meeting he organized in Maritea, Italy, and we have enjoyed a collegial friendship ever since, despite the distance separating us. Jack Szostak has begun to work on fatty acid vesicles and encapsulated polymerization reactions, and his research group has published a superb series of papers that are clearly on track toward artificial life.
There are major gaps in our knowledge, of course, and the question of how stable membrane vesicles first formed remains unanswered, despite the progress we, and others, have made. Membranes produced from pure fatty acids have a fairly narrow range of stability in terms of pH, because the carboxylic acid head groups need to be partially ionized, which requires a weakly alkaline pH range. Furthermore, the negative charge makes them susceptible to interacting with divalent cations such as calcium and magnesium, which precipitate the fatty acids into an amorphous curd rather than membranous vesicles. We still have a lot to learn.
What We Learned in Kamchatka
Fast forward to the year 2000 and beyond. In order to face up to the problems that self-assembling molecules might need to overcome on the way to the origin of life 4 billion years ago, I have been doing fieldwork beyond the lab, testing ideas in what we call prebiotic analog environments. My guess is that Earth was still pretty hot when life began. There was an ocean, probably salty, and volcanic islands like Hawaii and Iceland today. Water would evaporate from the sea and fall as rain, so there would be freshwater ponds and hot springs on the flanks of the volcanoes. With Sherry Cady, the editor in chief of Astrobiology, I organized the first such fieldwork in Kamchatka, Russia, in 2002. Figure 3 shows what it's like to collect samples from a large fumarole in the crater of Mount Mutnovski. What impressed me were the obvious heat, the fluctuations of wet and dry conditions around the ponds, and the acid pH range due to the sulfur in the water. The earliest forms of life would be considered extremophiles in current terminology—living in hot, acidic conditions.

Collecting samples from a fumarole in the crater of Mount Mutnovski, Kamchatka, Russia. Inset: the author. Color images available online at
What properties of analogue sites can guide laboratory simulations? It comes down to five Cs and an E, a mnemonic like CHONPS, but having to do with conditions rather than elements:
Lipids, Ester Bonds, and the First Nucleic Acids
The last part of this story requires a little knowledge of biochemistry. First off, recall that virtually all the polymeric substances of life are produced by one of the simplest chemical reactions, called condensation. The basic reaction of condensation is that the equivalent of water molecules are extracted from between monomers, which are then linked by ester bonds to produce nucleic acids or the peptide bonds of proteins. In life today, synthesis of the linking bonds of polymers is catalyzed by polymerases and ribosomes using activated monomers like ATP. However, nothing like this was available on prebiotic Earth. So, the question is how could sufficiently complex polymers have been synthesized to get life started. Well, under certain conditions, synthesis of ester bonds is spontaneous. All you need to do is make conditions dry enough so that water itself becomes a leaving group. It seemed to me that the conditions I was seeing in volcanic pools largely fulfilled the “five Cs and an E” defined above. All around the edges of the pools there were cycles of wetting and drying that would concentrate any potential reactants in the solution. Energy would be added during drying, producing a chemical potential that could drive ester bond formation by condensation. All I had to do was to add lipid vesicles, the compartments, along with monomers, then see whether polymers could be produced.
But how could adding compartments promote polymerization reactions? Ever since we first trapped solutes in lipid vesicles with drying and wetting cycles, I have had the notion that this is very different from simply drying down in the absence of lipid. Drying a solution turns typical solutes into a solid film; but, if a lipid is present, the solute becomes concentrated and organized within the two-dimensional plane of the lipid layers. Not only that, but lipids are liquid crystals, which means that the solute molecules are not immobilized in a solid but instead can diffuse around within the two-dimensional planes of the lipid matrix. This led to a new idea, which is central to the hypothesis we are currently testing: What if the lipid structures impose order on the monomers in such a way that specific polymerization is promoted during condensation reactions? If so, such conditions would be expected to lead to the synthesis of long polymers. Furthermore, the polymers would be back in solution as soon as water is added to the system, and they would be encapsulated in lipid vesicles. Figure 4 shows how we imagine mononucleotides would look when they are organized and concentrated between lipid bilayers in a multilamellar lipid matrix. It is easy to see how order can be imposed on the nucleotides and why the ordering makes it much more likely that they will be able to form phosphodiester bonds that link them into nucleic acid molecules. When the nucleotides are in solution, they are in random motion, and each is surrounded by thousands of water molecules. But in the lipid matrix the water is gone, and the molecules are confined within a two-dimensional plane.

Adenosine monophosphate (AMP) is one of the nucleotide monomers of nucleic acid. (A molecular model of AMP is shown in top and side views in the inset.) When dried in the presence of lipid vesicles, the vesicles fuse to form multilamellar sheets that capture and organize the AMP molecules between alternating layers. Color images available online at
We have promising preliminary results to describe. First, I want to say something about the way a research laboratory works in a university setting. Lab groups are very international, and we work as a team composed of graduate students, post-doctoral associates, technicians, and what is called a principal investigator, or PI. Doctor Sudha Rajamani (from India) did much of the hands-on work, Dr. Felix Olasagasti (from Spain) did some of the preliminary studies, Seico Benner (from Japan) and Amy Coombs (USA) carried out several of the technical assays, and Dr. Alexander Vlassov (from Russia) did some of the analytical gels that finally convinced us there was a real phenomenon. My job, of course, was to come up with the original idea (which I first began to work on 25 years ago) and to write the proposal to get the grant that supported this team for 2 years.
The basic experimental approach is fairly straightforward but had to be repeated quite a few times under different conditions to be convincing. We made up dispersions of lipid vesicles in water (actually a dilute acid at pH 3 to simulate the acidity of volcanic hot springs). Mononucleotides, the monomers of RNA, were added so that there was about one nucleotide per lipid in the solution. The mixture was warmed to 90°C for 2 hours while it was dried with a gentle stream of carbon dioxide to simulate the prebiotic atmosphere. A small amount of water was then added, the mixture was stirred for a few seconds to disperse the lipid vesicles, and the cycle was repeated up to 7 times. The idea was to simulate the conditions of a volcanic hydrothermal area on early Earth, similar to the sites in Kamchatka described earlier, in which a continuous drying and wetting process occurs at the edges of the pools. The water would be fairly hot (80–90°C) and weakly acidic.
Well, what happened? We had a wonderful surprise. When we tested the solution for the presence of polymers, we found that RNA-like molecules had been synthesized, ranging from 20 to 100 nucleotides in length (Rajamani et al., 2008). The yields were very low by the standards of organic syntheses: less than 0.1% of the nucleotides had been linked up into longer polymers, representing a few micrograms of product from the milligram quantities of nucleotides present in the mixture. However, I see no particular reason to think that there were high-yield polymerization reactions occurring in the prebiotic environment. The reactants were complex mixtures of hundreds of different organic compounds, so the reactions leading to early biopolymers almost certainly produced very low yields.
An important outcome of the experiment was that when the last cycle of hydration was completed by adding water, the lipid captured the RNA in vesicles. I consider such vesicles to be the first step toward cellular life, that is, microscopic membrane-bounded compartments containing complex mixtures of polymers that have the potential to be both catalysts (ribozymes) and carriers of information. But we can't get too excited yet. It's good to find a plausible way to make polymers resembling RNA, but they are not like biological RNA because the monomers are linked by 3′−5′ phosphodiester bonds (the bond of biological RNA) but also unnatural 2′-5′ bonds. This mix of bonds means that the molecules will not readily fold into structures that can have catalytic activity like a ribozyme. Furthermore, there is no obvious process by which the molecules can reproduce themselves. What we need to do now is to isolate and amplify those few strands that have catalytic activity.
Maybe Even Replication?
We have just completed a second series of experiments testing an obvious possibility that arises from the results described above. If polymerization of nucleotide monomers into polymers like RNA can be promoted by lipid matrices, the next step is to see whether the same conditions could drive template-directed synthesis. We are now including synthetic nucleic acid templates in the mixture of mononucleotides and lipid vesicles. During drying, it is conceivable that the monomers will line up on the template to form phosphodiester bonds. Felix Olasagasti carried out the experiment in which we included a synthetic nucleic acid template along with four 5′-deoxynucleotides (AMP, TMP, GMP, and CMP) and did the usual wet-dry cycles up to 5 times at 85°C, with phosphatidic acid as the membrane-forming phospholipid. We then had the products sequenced in the new DNA-sequencing facility established by Nader Pourmand at University of California, Santa Cruz. The amazing result is that short DNA strands of variable lengths were produced, but they were not random polymers. Instead, it was very clear that they contained base sequences that would be expected if complementary sequences had been synthesized on the template (Olasagasti et al., 2011).
If this works, it would represent a plausible way for replication to begin on prebiotic Earth, driven non-enzymatically by anhydrous cyclic condensation reactions and directed by the sequence of bases in the template strand. Significantly, such a process would be equivalent to a prebiotic polymerase chain reaction, because the elevated temperature when water is added back to the dry matrix would cause the double-stranded nucleic acid to melt back into two separate chains, just as it does in the PCR devices we use every day in the laboratory. The result would be growth and replication of the original polymers, together with amplification of the sequences in the nucleic acids. This, of course, is a very exciting result, but as always in science, there is more to do. The next and most important step is to find conditions that permit the cycle of life to begin, in which a genetic molecule directs the synthesis of a catalytic molecule, and the catalyst in turn speeds up the synthesis of the genetic molecule. This is where our current knowledge ends and speculation begins. But speculation can turn into a useful hypothesis, and the simulation I described earlier can provide the test.
So, that's what it's like to spend 35 years trying to figure out how life began. I will finish up by saying that as the years accumulate in a scientific lifetime, something interesting happens, which is that seemingly diverse bits of information accumulate in one's brain and patterns begin to emerge. All too often, the patterns are just ideas that don't match reality when tested by experiment. But rarely, perhaps once in a lifetime, a pattern suggests a novel way to understand life's processes that withstands critical scrutiny. In this brief essay, I have tried to show how such a pattern can emerge. I don't know yet whether the basic idea is right or wrong, but as long as I wake up in the morning thinking about another way to test it, I can be happy.
Footnotes
Acknowledgments
I want to acknowledge the graduate students and postdoctoral associates who were brave enough to spend a few years in my lab. From A to Z, my special thanks go to Charles Apel, Gail Barchfeld, Ajoy Chakrabarti, Will Hargreaves, Dmitri Kirpotin, Sarah Maurer, Pierre-Alain Monnard, Trishool Namani, Felix Olasagasti, Sudha Rajamani, Sara Singaram, Roscoe Stribling, Sasha Volkov, and Helmut Zepik. The research described here was supported by the NASA Exobiology and Astrobiology programs.