
Abstract
Candida albicans is an opportunistic fungal pathogen responsible for a variety of cutaneous and systemic human infections. Virulence of C. albicans increases upon exposure to some environmental stresses; therefore, we explored phenotypic responses of C. albicans following exposure to the environmental stress of low-shear modeled microgravity. Upon long-term (12-day) exposure to low-shear modeled microgravity, C. albicans transitioned from yeast to filamentous forms at a higher rate than observed under control conditions. Consistently, genes associated with cellular morphology were differentially expressed in a time-dependent manner. Biofilm communities, credited with enhanced resistance to environmental stress, formed in the modeled microgravity bioreactor and had a more complex structure than those formed in control conditions. In addition, cells exposed to low-shear modeled microgravity displayed phenotypic switching, observed as a near complete transition from smooth to “hyper” irregular wrinkle colony morphology. Consistent with the presence of biofilm communities and increased rates of phenotypic switching, cells exposed to modeled microgravity were significantly more resistant to the antifungal agent Amphotericin B. Together, these data indicate that C. albicans adapts to the environmental stress of low-shear modeled microgravity by demonstrating virulence-associated phenotypes. Key Words: C. albicans—LSMMG—Amphotericin B—Irregular wrinkle colony morphology—HARV bioreactor. Astrobiology 11, 825–836.
1. Introduction
The most prominent bacterial pathogen examined in low-shear microgravity (ground-based and in spaceflight) is the invasive enteric pathogen Salmonella enterica, serovar Typhimurium (S. Typhimurium), although Escherichia coli, Streptomyces levoris, Bacillus subtilis, Methylbacterium organophilium, Methylosinus methanica, Clostridium butyricum, and Pseudomonas aeruginosa have also been shown to undergo alterations consistent with increased pathogenicity (Leys et al., 2004; Guadarrama et al., 2005; Wilson et al., 2007; Crabbé et al., 2008, 2010; Vukanti et al., 2008; Wilson et al., 2008). Several studies support the notion that E. coli responds to the effects of microgravity with a generalized stress response and increased biofilm formation (Lynch et al., 2004, 2006; Vukanti et al., 2008). Biofilms, defined as structurally heterogeneous and genetically diverse cellular communities encased in a polymeric matrix, are potent causative agents of disease, in part due to increased antimicrobial resistance. It is also worth noting that not all prokaryotic pathogens analyzed, such as Yersinia pestis, respond to LSMMG with increased virulence (Lawal et al., 2010). The notion of increased virulence of pathogenic microbes is alarming from the perspective of astronaut health, considering studies showing that microgravity exposure adversely affects the human immune system (Kaur et al., 2005; Sonnenfeld, 2005; Aponte et al., 2006).
Eukaryotic microbes also respond to the microgravity environment, and early studies in both spaceflight and LSMMG demonstrated that Saccharomyces cerevisiae experienced cell clumping and transition from a bipolar to a random budding phenotype (Walther et al., 1996; Purevdorj-Gage et al., 2006; Van Mulders et al., 2011). Several genes associated with cell separation and bipolar budding were differentially expressed following culture in LSMMG, consistent with the observed morphological changes (Sheehan et al., 2007). Recent studies have demonstrated that C. albicans also undergoes morphological transformation upon exposure to LSMMG and spaceflight, including an abnormal budding pattern and an increase in filamentation (Altenburg et al., 2008; unpublished data). Hyphal formation, a form of filamentation, is associated with pathogenicity in C. albicans, an endogenous opportunistic human pathogen responsible for a wide array of diseases, such as oral thrush, vaginitis, biofilm-associated infections, and disseminated candidiasis (Lo et al., 1997; Gullo, 2009; Lopez-Martinez, 2010). In an immunocompromised host, the normally commensal fungus can spread rapidly and cause serious infections. Mortality from systemic infections involving the blood stream and internal organs is quite high (Pfaller and Diekema, 2007). This eukaryotic pathogen also maintains populational heterogeneity, which enables quick responses, via stimulation of signaling programs, gene activation, and overcoming silencing mechanisms, to protect against environmental stress (Rustchenko-Bulgac et al., 1990; Soll, 2002; Ramage et al., 2009). Cellular responses to environmental adaptations often present as increased yeast-to-hyphal transition (filamentation) and high-frequency phenotypic switching (we focus on switching as it relates to colony morphology herein), which are correlated with drug resistance, decreased antigenicity, and reduced sensitivity to host scavenger cells and oxidants.
In the present study, we tested the hypothesis that the environmental stress of LSMMG induces phenotypic adaptations in C. albicans. Specifically, we evaluated cellular morphology, biofilm formation, phenotypic switching, and antifungal resistance following long-term culture of C. albicans in LSMMG.
2. Materials and Methods
2.1. Strains and growth conditions
The C. albicans clinical isolate SC5314 was used throughout these studies. Cells were maintained as colonies on YPD agar (1% yeast extract, 2% peptone, 2% glucose, 1% agar; Becton Dickinson, NJ, USA) and were re-plated every 4–6 weeks. Overnight cultures were initiated from a single colony into YPD broth and grown in shaker conditions at 30°C for 16–18 h. YPD medium containing penicillin (100 units/mL) and streptomycin (100 μg/mL) (HyClone, UT, USA) was used for these studies to prevent bacterial contamination throughout the duration of long-term (12-day) HARV experiments. Preliminary testing determined that incorporation of these antibacterial agents into the growth medium did not affect parameters analyzed in these studies.
2.2. Propagation and analysis of cultures
To initiate LSMMG experiments, 2 mL of an overnight culture (OD600>2) was added to 60 mL YPD medium and transferred to a sterile bioreactor (HARV), which resulted in an initial cell density of 2.5×106 cells/mL. Each experiment consisted of 4 HARV bioreactors, duplicates in both the experimental and control orientations. Cells were grown in HARVs with rotation in either a parallel (continuous sedimentation, LSMMG) or perpendicular (control) orientation to the gravitational vector at 30°C in an upright 14 ft3 (0.4 m3) incubator. Every 24 h, 1 mL of each stationary culture was removed from the bioreactor and inoculated into 60 mL fresh medium (plus penicillin and streptomycin). The bioreactors were then fully drained and reinoculated with the freshly inoculated culture medium for propagation with continued rotation in the appropriate orientation. At the indicated time points and during the regular medium change, cells were analyzed by light microscopy for filamentation and plated to evaluate colony morphology; three 2 mL aliquots of the planktonic culture were harvested from each HARV, pelleted by brief centrifugation, and the cell pellets were stored at −80°C for RNA isolation and further microscopic analyses. Antifungal drug resistance testing was performed at 5, 8, and 12 days. For biofilm analysis, cells were cultured in HARVs for 12 days, and bioreactor membranes were isolated and processed for imaging.
2.3. RNA isolation and quantitative reverse transcription–polymerase chain reaction (qRT-PCR)
For gene expression analyses, frozen cell pellets were thawed on ice and total RNA extraction was performed according to the RNeasy miniprotocol (Qiagen, CA, USA), following the mechanical disruption protocol supplied by the manufacturer. Homogenization of the samples was completed with a Mini-Beadbeater-8 (Biospec Products, OK, USA) with 0.5 mm glass beads. RNA integrity and concentration were evaluated at 260/280 nm with a Nanodrop (Thermo Scientific, MA, USA).
Quantitative reverse transcription–polymerase chain reaction was completed with the SuperScript III Platinum SYBR Green One-Step qRT-PCR kit (Invitrogen, CA, USA). Twenty-five microliter reactions contained 0.2 mM of each dNTP, 0.5 μM sense and antisense primer, 0.25 μL RT-Taq mixture, and 0.4 μg RNA. Primers were designed with Invitrogen Oligo Perfect Designer and obtained from Invitrogen (see Table 1 for primer sequence). Amplicons ranged in size from 75 to 200 nucleotides. Gene products were amplified with a Rotor-Gene 3000 (Corbett Research, CA, USA) by incubation at 50°C for 10 min (cDNA synthesis); 55°C for 3 min; 95°C for 5 min; and 40 cycles of 95°C for 15 s, 60°C for 30 s, 72°C for 30 s. The melt curve analysis was carried out immediately after amplification in 1°C increments from 72°C to 95°C, holding for 5 s at each step. Data were normalized to PDA1, and fold changes were calculated for LSMMG relative to control conditions by using the
2.4. Biofilm microscopy
Confocal scanning laser microscopy was performed on biofilm communities associated with the siliconized rubber membranes of the HARV following 12 days of continuous culture in the control or LSMMG orientation. The membranes were isolated from each HARV, rinsed with water, and stained with 25 μM FUN-1 [2-chloro-4-(2,3-dihydro-3-methyl-(benzo-1,3-thiazole-2-yl)-methylidene)-1 phenylquinolinium iodide] (Molecular Probes–Invitrogen, CA, USA; see dye description in the Antimicrobial resistance section) for 30 min. Biofilm communities were imaged with a Leica TCS SP2 AOBS laser scanning confocal microscope (Leica Microsystems, Inc., IL, USA) with a 63×, 0.9NA water immersion objective lens. Excitation was performed at 488 and 561 nm, and emission was collected at 506–533 nm for green and 611–727 nm for red. Image analysis was performed with Imaris software (Bitplane).
2.5. Colony morphology
To evaluate the effect of LSMMG on colony morphology, cells were removed from treatment conditions (control or LSMMG) daily, diluted (approximately 1:106), spread on YPD plates, and allowed to grow for 3 days at 30°C. Samples were prepared in biological and technical duplicate, and colony morphology was quantified by visual observation. Individual colonies were imaged with a stereoscope (Nikon SMZ1500) at 10× magnification, and images were obtained with an Infinity 2 camera (Lumenera Co., Ontario, Canada).
To observe the cellular morphology characteristics of each colony type, colonies generated from cells cultured for 12 days in control or LSMMG conditions were mechanically disrupted, and cells were fixed in 4% paraformaldehyde. Cells were imaged with a Photometrics CoolSnap digital camera with light microscopy (Nikon Eclipse E800) at 600× magnification.
2.6. Antimicrobial resistance analysis by flow cytometry
The fluorescent probe FUN-1 was used to quantify cell viability secondary to damage incurred following Amphotericin B (AmB) exposure. In metabolically active cells, FUN-1 is converted from a diffuse, yellow-green dye to compact cylindrical intravacuolar structures (CIVS) characterized by fluorescence in the red wavelength. A previously described antifungal susceptibility testing method was used with slight modifications (Wenisch et al., 1997). Briefly, serial 2-fold dilutions of AmB (Acros, NJ, USA) were prepared in a 96-well plate with drug concentrations ranging from 4 to 0.03125 mg/L in standard YPD medium. C. albicans cell suspensions were created by diluting control- or LSMMG-exposed cells to 107/mL in YPD medium. Diluted cell suspensions (100 μL) were added to 100 μL of each drug dilution in biological and technical duplicate for a total of four replicates per treatment condition. Positive (no AmB added) and negative (50% household bleach) viability controls were included in each analysis. Following a 30 min incubation period (at 30°C), FUN-1 was added to each well to a final concentration of 5 μM. Cells were incubated for 30 min at 30°C in the dark. Each reaction mixture was diluted in 1 mL ice-cold phosphate-buffered saline and analyzed immediately by flow cytometry.
Samples were analyzed with a FACScalibur (Becton Dickinson, NJ, USA) flow cytometer; Cell Quest software was used for data acquisition and analysis. The instrument settings were as follows: forward scatter, 1.50 linear gain; side scatter, 385 V; fluorescence (FL-2), 550 V log; and threshold value, 52. Scatter parameters were established based on a population of cells not exposed to AmB or FUN-1, and FL-2 parameters were determined based on cells stained with FUN-1 but not exposed to drug. For each sample, 10,000 events (cells) were analyzed for forward scatter, side scatter, and fluorescence (FL-2) at a rate of 500–1000 cells/s.
To evaluate the viability of cells exposed to various concentrations of AmB, a viability threshold was established with the positive control FL-2 histogram. The threshold was determined such that 99±1% of untreated cells had fluorescence values above the determined limit. This threshold was then applied to all histograms to determine the percentage of viable and dead cells. Impairment of cell metabolism as a result of AmB exposure is evident by decreased FUN-1 staining, visualized as a leftward shift on the x axis of the FL-2 histogram. The percentage of viable cells following AmB exposure was compared between control- and LSMMG-exposed cells to quantify fold change in AmB resistance. Data representative of two separate HARV experiments are shown.
2.7. Statistical analysis
For all data sets, means±standard deviations are shown. For each experiment, the number of replicates performed is indicated in the appropriate figure legends. Determination of the difference between mean values for each data set was assessed by Student t test. Significance testing compared LSMMG to control values and was established at p values <0.05, with more specific p values provided in the appropriate figure legends.
3. Results
3.1. Expression of biofilm- and hyphal-related genes following LSMMG
We have previously shown that exposing C. albicans to LSMMG for 5 days induces a filamentous morphology (Altenburg et al., 2008). Indeed, a temporal analysis of cell morphology demonstrated a time-dependent increase in filamentation that was initially observable by 4–5 days of LSMMG (Fig. 1A, dotted line). LSMMG-induced filamentation continued to increase throughout the 12-day time course. The visually observed increase in filamentation was confirmed by expression analyses of form-specific genes, namely, the yeast-specific gene Yeast Wall Protein 1 (YWP1, gray bars) and the hyphal-specific gene Hyphal Wall Protein 1 (HWP1, white bars; Fig. 1b) (Sundstrom, 1999; Granger et al., 2005). YWP1 expression is maximally repressed within 5 days and remains at that level throughout the 12-day LSMMG exposure. HWP1, on the other hand, is increased by over 2-fold within 8 days and continues to increase to almost 4-fold by the 12 day-time point, mirroring the cellular filamentous morphology trend.
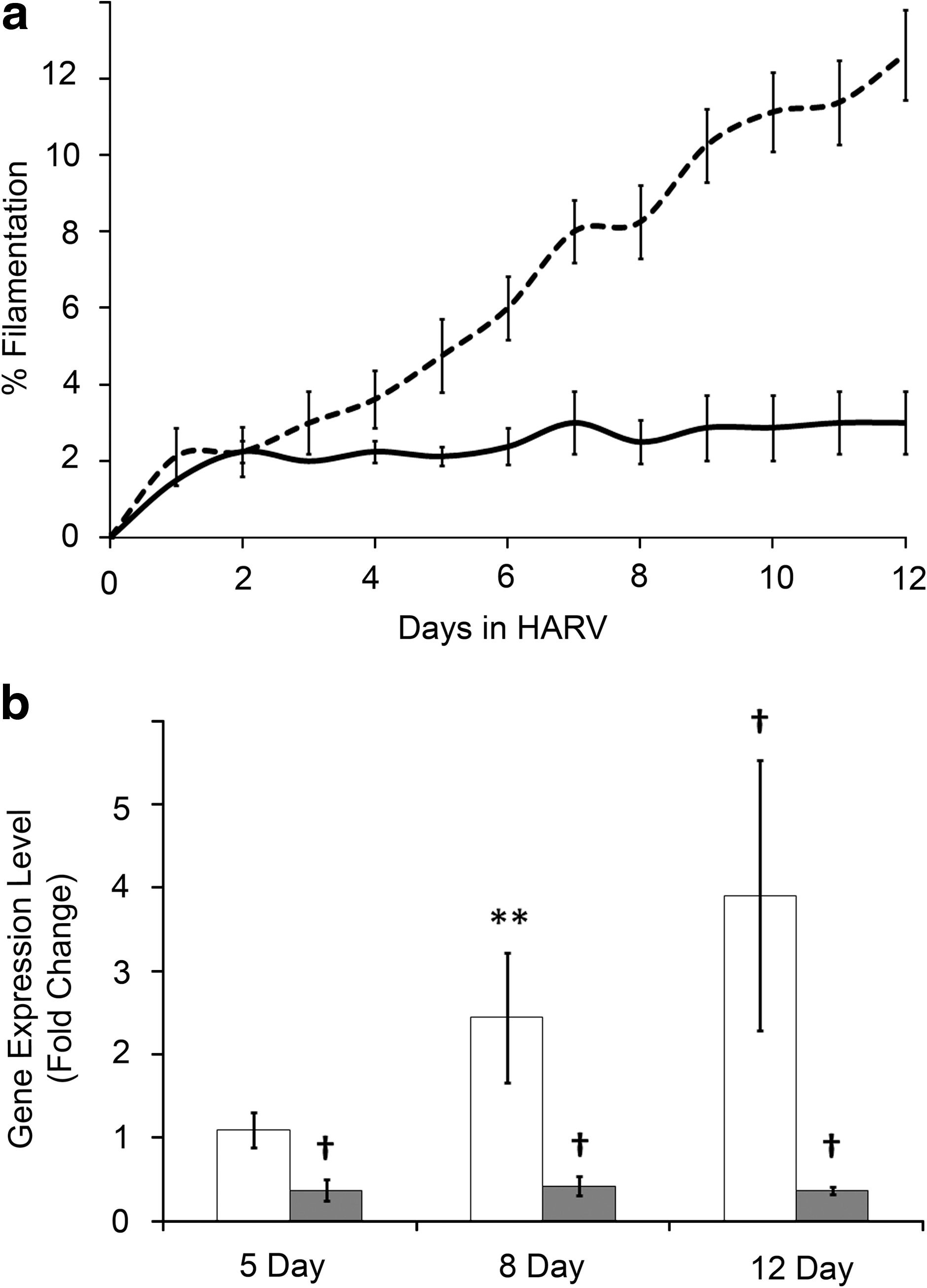
Temporal analysis of filamentation and associated gene expression. (
To determine whether other genes associated with filamentation or biofilm formation were differentially expressed, selected genes from the molecular pathway that regulates HWP1 expression were interrogated in cells exposed to control or LSMMG conditions for 12 days (Fig. 2). Only genes in this pathway known to be transcriptionally regulated (indicated in white) were analyzed (Fig. 2a). Similar to HWP1, expression of both ALS1 and ALS3 was increased upon exposure of C. albicans to LSMMG (Fig. 2b). Furthermore, the expression of BCR1, a transcription factor that contributes to the positive regulation of the adhesin gene family and is associated with biofilm formation was also increased following growth in LSMMG. Surprisingly, the positive regulator of BCR1 gene expression, TEC1, was minimally affected by LSMMG exposure.

Genes implicated in C. albicans filamentation and biofilm formation. (
Not all genes in the HWP1 regulatory pathway were investigated. Although TUP1 is an important negative regulator of the pathway, its activity is controlled post-transcriptionally through an interaction with DNA-binding proteins; therefore, it was not analyzed in this context. Additionally, CPH1 and EFG1 were not analyzed in these studies, since they are primarily regulated through phosphorylation via the mitogen-activated protein kinase (MAPK) and the protein kinase A (PKA) signaling pathways, respectively. In addition to the regulatory changes analyzed herein, we cannot rule out the possibility that, due to serial passage of C. albicans over 12 days, heritable genetic changes may also occur. Mutation analysis will be addressed in future studies.
3.2. Biofilm formation following LSMMG exposure
Multicellular biofilm communities could be detected on the HARV siliconized rubber membranes after as little as 8 days of propagation in LSMMG and were analyzed following 12 days of continuous culture. A representative confocal image that demonstrates the three-dimensional and complex structure of an intact biofilm from cells exposed to LSMMG conditions is shown in Fig. 3b relative to those formed in control conditions (Fig. 3a). Biofilms formed under control conditions were comprised of cells primarily in the yeast form, with occasional filaments. The biofilm was relatively uniform in thickness and density. Due to the orientation of the control HARV, the formation of this biofilm is likely initiated by cellular sedimentation. Conversely, biofilms formed under LSMMG conditions contain a higher percentage of filamentous cells. Additionally, and consistent with prior findings, LSMMG-induced biofilms, while not necessarily thicker, show a greater range of cellular density and biofilm thickness (Fig. 3) (Woolley et al., 2010). The orientation of the LSMMG HARV (membrane is parallel to the gravitational vector) prevents sedimentation on the membrane; therefore, biofilms formed under LSMMG conditions must depend on a more active community-building process than those formed under control conditions.

Representative confocal images of C. albicans biofilms formed under control (
3.3. Morphologic switching to “hyper” irregular wrinkle colony morphology following LSMMG exposure
The term “phenotypic switching” encompasses the white-to-opaque switch as well as alterations in colony morphology. In the context of these studies, we did not observe the white-to-opaque switch but did observe a striking change in colony morphology when cells were cultured under LSMMG conditions and plated on YPD agar (Fig. 4). An altered morphology reminiscent of the well-described irregular wrinkle morphology, yet with dramatically more convoluted surface ridges (areas of high cell density and areas devoid of cells), was induced by propagation in LSMMG conditions (Fig. 4c) (Slutsky et al., 1985). The term “hyper” irregular wrinkle (HIW) has been coined to describe this highly convoluted colony morphology, which resembles that previously described as Morphological form A13 (Brown-Thomsen, 1968). Cells exposed to control conditions formed smooth colonies almost exclusively (Fig. 4a), and HIW colonies (or any other altered morphologies) very rarely formed under these conditions, even when cultured for extended periods of time. When analyzed in long-term LSMMG, there was a time-dependent increase in the percentage of cells giving rise to the HIW morphology (Fig. 4e). HIW colonies were detected at minimal levels by 5 days of culture in LSMMG, but by 8 days over 40% of the colonies were of the HIW morphology, and after 12 days over 90% of the colonies were represented by the HIW morphology. The transition from smooth to HIW did not involve intermediate morphologies and was stable (relative to the rate of HIW induction) once cells were removed from the LSMMG environment and exposed to serial passage in liquid or on solid medium (data not shown).
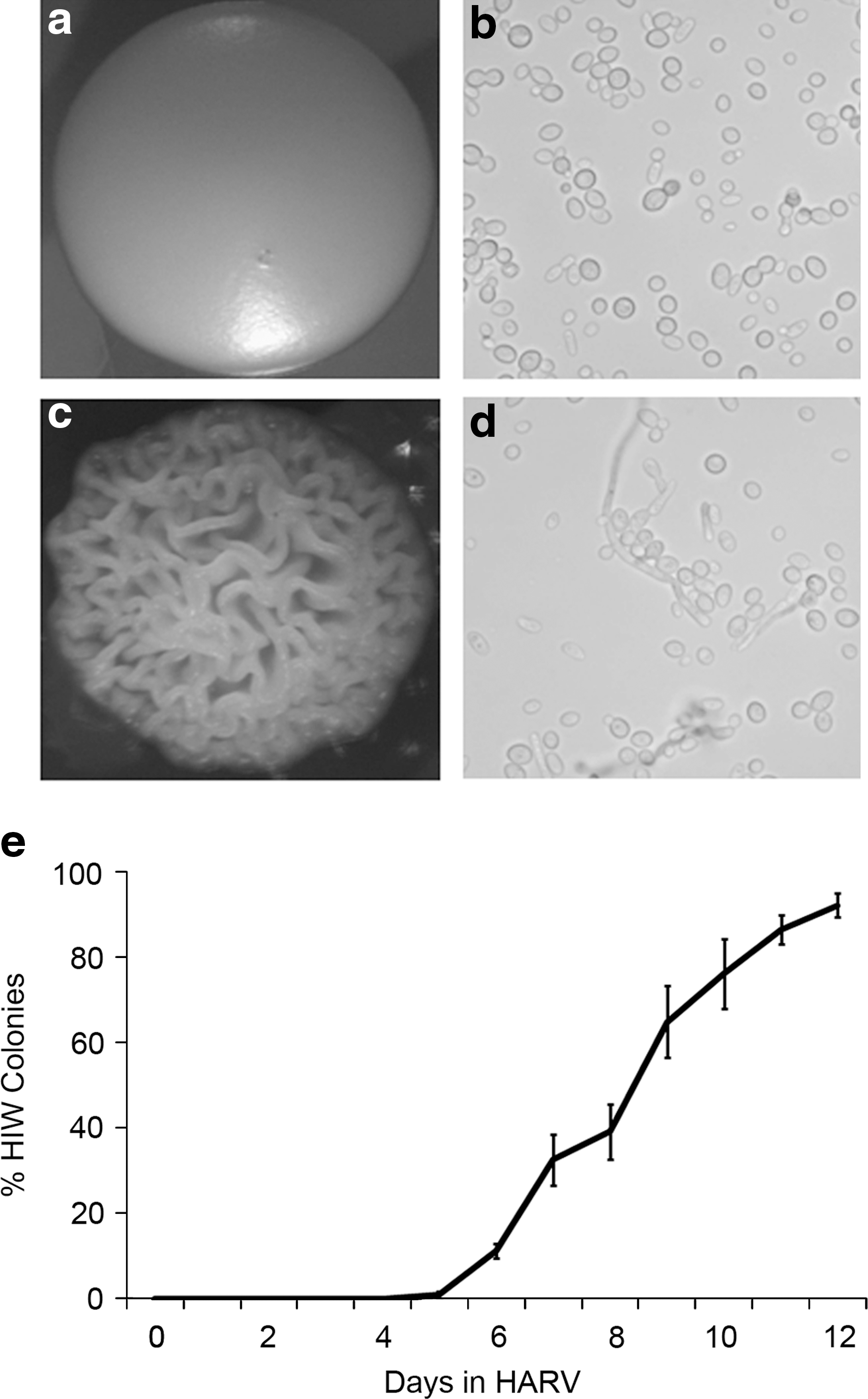
Evaluation of colony morphology arising from C. albicans exposed to LSMMG. (
Predictably, smooth colonies contained exclusively yeast-form cells as illustrated by microscopy (Fig. 4b). Conversely, and consistent with previous descriptions of irregular wrinkle colonies, HIW colonies were comprised of both filamentous and yeast cells (Fig. 4d) (Radford et al., 1994). Gene expression analysis further confirmed the prevalence of filamentous cells in the HIW colonies. Cells isolated from HIW colonies had a 25-fold higher expression of HWP1 and a 1.56-fold lower expression of YWP1 than cells from control smooth colonies. HIW colonies were also more resistant to mechanical disruption than smooth colonies, which suggests a more integrated community structure.
3.4. Resistance of LSMMG-exposed cells to the antifungal agent Amphotericin B
Biofilm communities and colony morphology alterations are consistent with increased antimicrobial resistance; therefore, we explored the effects of LSMMG on antifungal resistance, using flow cytometry (Calderone and Fonzi, 2001; Chandra et al., 2001). For these studies, cell viability following AmB exposure was compared between control and LSMMG conditions to quantify resistance. To ensure that propagation in HARV bioreactors induces similar drug sensitivity as described for C. albicans growth in shaker conditions, cells were grown for 12 days in control-oriented HARV bioreactors and exposed to a serial 2-fold dilution of AmB (Fig. 5). Cell death was manifested as a leftward-shift in the fluorescence (FL-2) histogram that resulted from decreased conversion of FUN-1 to CIVS and subsequent decreased fluorescence intensity (Fig. 5a). The dose response to AmB of C. albicans cultured in the control HARV bioreactor (Fig. 5b) was similar to that observed from shaker cultures; therefore, the previously described range of AmB concentrations was used for LSMMG studies (Wenisch et al., 1997).
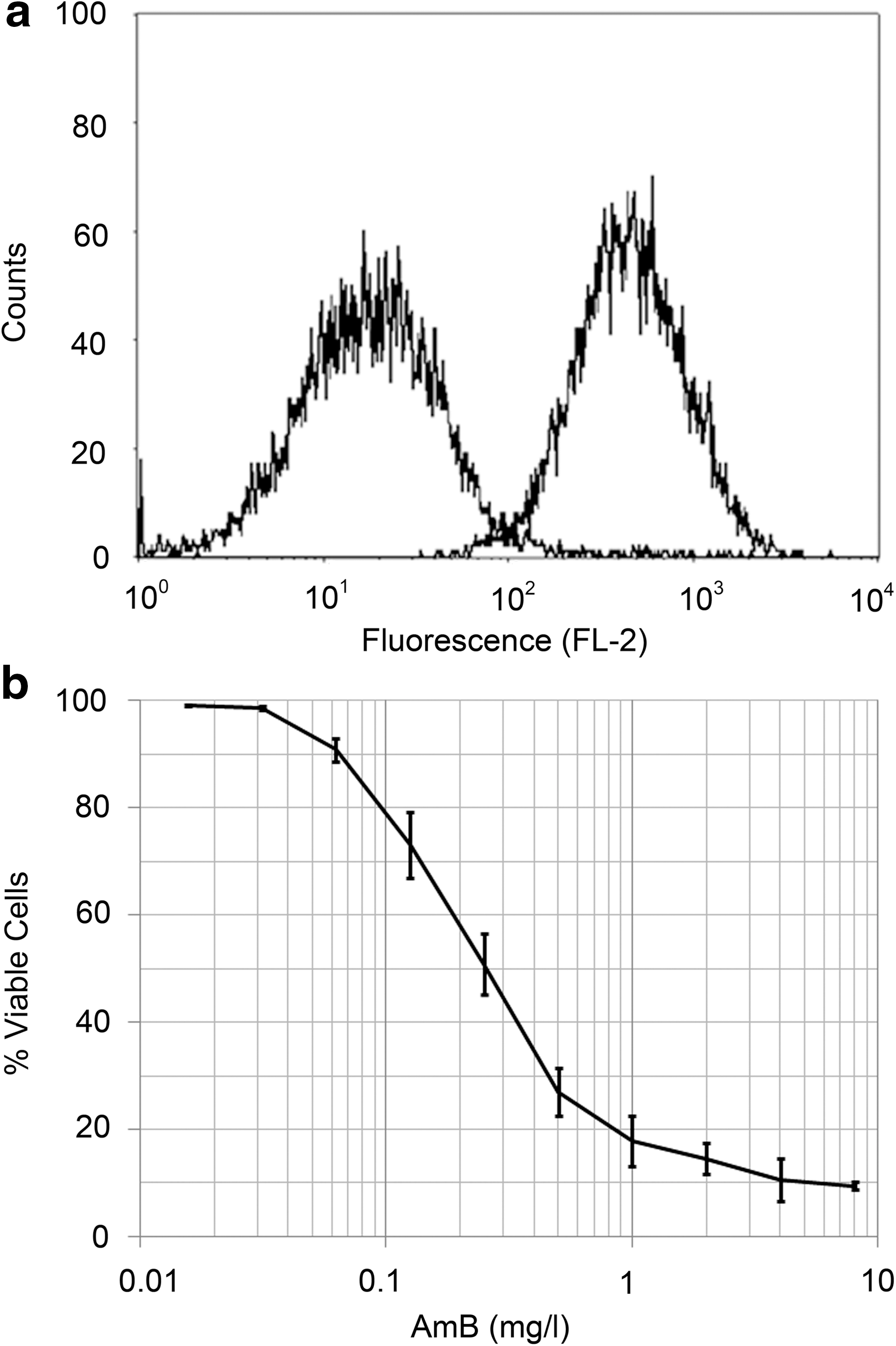
C. albicans viability following AmB exposure. Flow cytometry was used to determine viability of C. albicans upon exposure to AmB following growth in the HARV bioreactor in the control orientation. (
Cells exposed to LSMMG for 12 days were similarly analyzed. Control and LSMMG-exposed cells have similar viability profiles in the absence of antifungal exposure (Fig. 6a). In contrast, cells cultured in LSMMG demonstrate an increased resistance to 2 mg/L AmB compared to control conditions, observable as a separation of the leftward (drug susceptibility) shifting peaks (Fig. 6b). AmB resistance of C. albicans was tested following culture for 5, 8, or 12 days in control or LSMMG conditions (Fig. 6c). Increased time of exposure to LSMMG resulted in enhanced resistance to AmB relative to cells in the comparable control conditions. Maximal resistance (2.1-fold) was observed in cells grown in LSMMG for 12 days and exposed to 2 mg/L AmB (checkered bars). However, even at AmB concentrations as low as 0.25 mg/L (light gray bars), a statistically significant increase in viability can be detected in cells cultured for 12 days in LSMMG compared to control conditions. AmB concentrations lower than 0.25 mg/L elicited no difference in resistance between LSMMG-exposed and control cells (data not shown).
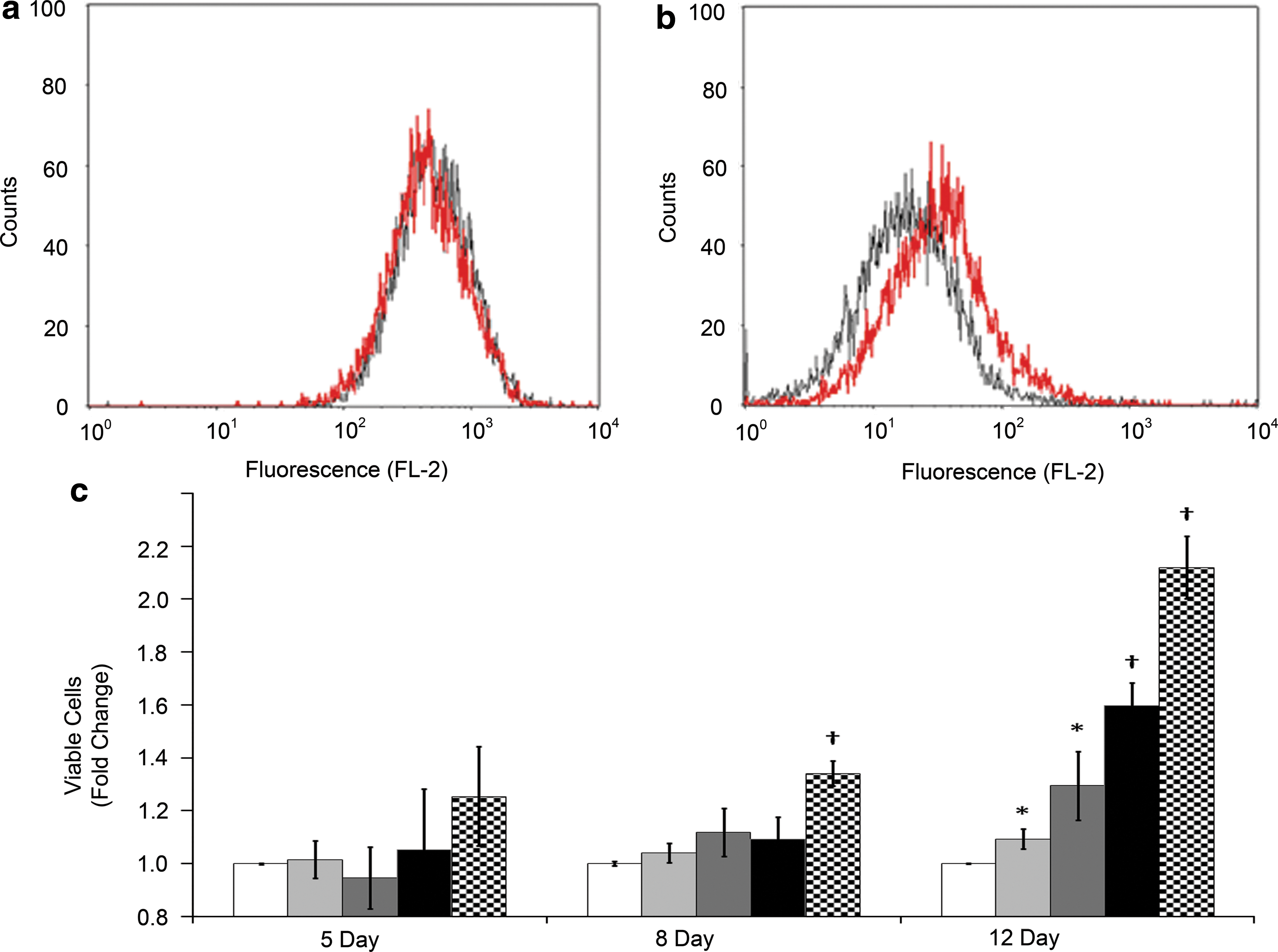
Resistance of LSMMG-propagated C. albicans to AmB. (
4. Discussion
Candida albicans is a commensal organism that often colonizes the human body shortly after birth, and over 25% of the general population are carriers (Hibino et al., 2009). Although studies have failed to isolate hypervirulent strains of C. albicans, the rate of infection is rising due to the growing number of immunocompromised hosts, long-term antibiotic use, and increasing presence of medical implants (Concia et al., 2009; Cataldo and Petrosillo, 2011). Notably, this eukaryotic pathogen is credited with maintaining a populational heterogeneity that allows it to respond quickly to environmental cues (Rustchenko-Bulgac et al., 1990; Soll, 2002). Although no clear evidence has associated specific stress response genes as causative to increased virulence, it has been difficult to separate the two responses and their associated genes (Brown et al., 2007).
Low-shear fluid dynamics is a key aspect of the modeled microgravity environment and may provide the trigger for an environmental stress response by C. albicans (Gao et al., 1997). We have begun to explore the microenvironmental conditions that contribute to the Candida response to the low-shear environment. Hypoxia-induced genes do not appear to be differentially regulated under LSMMG conditions, nor does a pH response seem to be implicated in the morphological alterations observed (unpublished data). Ultimately, the response to environmental conditions, regardless of the specific trigger, is especially concerning in light of recent data that suggest microgravity increases the pathogenicity of several microbes (Nickerson et al., 2004; Wilson et al., 2007, 2008; Rosenzweig et al., 2010). Others have suggested that the growth environment obtained in the modeled microgravity bioreactors provides low-shear growth cues similar to those encountered in low-shear areas of the body and thus creates a more physiologically relevant environment for studying infection (Guo et al., 2000; Nickerson et al., 2003, 2007). Thus, the responses of C. albicans to low-shear conditions of the modeled microgravity environment may inform a better understanding of yeast virulence in the low-shear environment potentially encountered in vivo.
4.1. LSMMG-induced filamentation
The balance between filamentous and yeast forms of C. albicans is important for virulence due to form-specific contributions to tissue invasion and dissemination, respectively. LSMMG exposure induces a sustained morphological transition from budding cells to pseudohyphal and hyphal forms in C. albicans (Altenburg et al., 2008). The temporal analysis described in Fig. 1 demonstrates that the filamentation response to LSMMG is supported by changes in gene expression, namely, the upregulation of HWP1 and downregulation of YWP1. Preliminary evidence indicates that a filamentation response is also induced when C. albicans is grown in true microgravity experienced during spaceflight (unpublished data). Studies with S. cerevisiae suggest that LSMMG disrupts cell polarity, as evidenced by conversion from a bipolar to random budding pattern, so we continue to explore the role cell polarity plays in the C. albicans filamentation response to LSMMG exposure (Walther et al., 1996; Suzuki et al., 1998; Purevdorj-Gage et al., 2006; Van Mulders et al., 2011).
HWP1 is associated with both filamentation and biofilm formation. Therefore, we investigated whether the expression of other genes associated with both filamentation and biofilm formation, such as members of the agglutinin-like sequence (ALS) gene family (specifically ALS1 and ALS3), were affected by LSMMG. The genes characterized—TEC1, BCR1, HWP1, YWP1, ALS1, and ALS3—are primarily transcriptionally regulated, and most demonstrated differential expression in response to LSMMG. Control of EFG1 involves autoregulation as well as regulation by the PKA signaling pathway (Tebarth et al., 2003; Biswas et al., 2007). Similarly, CPH1 is primarily regulated by phosphorylation via the MAPK pathway (Brown and Gow, 1999). Although many genes associated with the regulatory pathway described in Fig. 2 suggest a responsiveness to LSMMG, the lack of induction of TEC1 (an activator of the pathway) is inconsistent and may indicate that the HWP1 pathway is predominantly upregulated by CPH1 and EFG1. The LSMMG responsiveness of these and other signaling pathways will be the focus of future studies.
4.2. Phenotypic switching
Colony morphology is a sensitive indicator of the dimorphic switch and therefore serves as an independent confirmation of the effects of LSMMG on cellular physiology (Noble et al., 2010). As a reflection of cellular morphology, colony macrostructure is therefore indirectly associated with virulence (Radford et al., 1994). Altered colony morphology is not uncommon in C. albicans, and several different switching systems have been described (Brown-Thomsen, 1968; Slutsky et al., 1985, 1987). Although switching can occur spontaneously at low frequencies (10−4), it occurs at much higher rates in response to environmental signals, and once a switch occurs, the colony morphology is stable and heritable (Soll, 2002). Clinical isolates, especially those isolated from invasive infections, display a higher rate of phenotypic switching (Soll, 1988; Jones et al., 1994; Vargas et al., 2000).
A striking increase in the presence of HIW colonies occurred within 12 days of LSMMG exposure (Fig. 4). The molecular mechanisms responsible for phenotypic switching are not well understood with the exception of the white-opaque switch originally observed in C. albicans strain WO-1 (Slutsky et al., 1987). The most obvious consequence of the white-opaque switch is that it controls the ability to mate, although it also regulates many other cellular functions, including cellular morphology, metabolism, and response to the environment (Slutsky et al., 1987; Lan et al., 2002; Huang et al., 2009). Interestingly, although phenotypic switching as it relates to colony morphology was a common event following exposure to LSMMG, we did not observe an increase in white-opaque switching.
Since the first reports of phenotypic switching, it has been suggested that colony morphology plays a role in virulence, specifically as a way to evade the immune system and alter resistance to antimicrobials (Slutsky et al., 1985). Indeed, mutation analysis has confirmed a relationship between virulence and colony morphology, but the genetic overlap appears to be only partial as some virulence-deficient mutants still undergo phenotypic switching (Noble et al., 2010). Studies have also shown that antimicrobial resistance is specific to each switch phenotype (Kiraz et al., 2000; Vargas et al., 2000, 2004). The molecular mechanisms that regulate phenotypic switching as it relates to colony morphology are considerably less well understood. Notwithstanding, SIR2 (Silent Information Regulator 2) appears to be a key regulator of silencing mechanisms, where diminished expression results in chromatin destabilization and induces dimorphic and phenotypic switching (Perez-Martin et al., 1999). TUP1 has been implicated in phenotypic switching, where it may function downstream of SIR2. Upstream regulators of the pathway may also involve the Ras/c-AMP/Efg1 signaling cascade (Tebarth et al., 2003). Future studies will focus on characterizing the molecular pathways that are responsive to LSMMG and instrumental in the colony morphology response.
4.3. Biofilm formation
In addition to phenotypic switching, other multicellular transitions occur in C. albicans, including formation of biofilm communities (Ramage et al., 2009; Uppuluri et al., 2009). These communities are often comprised of both yeast and filamentous forms, the combination of which provides for a more integrated and protective three-dimensional structure, as well as morphologies important for both dissemination and invasion within infected and immunocompromised hosts. Furthermore, studies have shown that biofilms are more resistant to environmental stressors such as pH, temperature, oxidative stress, and antimicrobial drugs (Davis, 2003; Seneviratne et al., 2008; Williams et al., 2009; Vediyappan et al., 2010). Therefore, biofilm formation serves as a potential indicator of increased pathogenicity.
Genes involved in adhesion (ALS1/3, BCR1, SUN41, and others), filamentation (HWP1, EFG1, TEC1, NRG1, and others), antifungal resistance (CDR1, CDR2, MDR1, MKC1, ERG3, ERG11, and others), and intercellular communication (the quorum-sensing molecules farnesol and tyrosol) have been implicated in biofilm development and environmental protection (Sanglard et al., 2003; Green et al., 2004; Ferreira et al., 2005; Nobile et al., 2008; Ramage et al., 2009). The clinical relevance of biofilm communities, combined with the gene expression indicators in Fig. 2, suggest that the C. albicans response to LSMMG may result in increased pathogenicity. Interestingly, high fluid flow has been associated with the development of biofilm structures with increased complexity (Uppuluri et al., 2009). These studies suggest that complex biofilms are also generated in conditions of low shear associated with LSMMG.
Cellular members of biofilm communities tend to have increased resistance to environmental stresses, including antifungal agents. The mechanisms of resistance are not fully understood and appear to be multifactorial. Physical protection from the extracellular matrix, increased cell density, physiological state of the cells, presence of persister cells, and differential gene expression all likely contribute to the increased resistance associated with biofilms. Specifically, genes that encode efflux pump proteins (MDR and CDR) and are involved in ergosterol biosynthesis (ERG) have been implicated in antimicrobial resistance of planktonic as well as biofilm cells (Ramage et al., 2009).
4.4. Antimicrobial resistance
A clinically relevant characteristic of both biofilm communities and altered colony morphologies is an increased resistance to antimicrobial agents (Ramage et al., 2009). Our findings that LSMMG increases biofilm complexity (Fig. 3) and induces morphological switching (Fig. 4) led us to investigate the possibility that LSMMG-conditioned cells are also more resistant to common antifungal agents. To this end, we evaluated resistance of C. albicans to AmB, using the rapid and quantifiable flow cytometry method with FUN-1 viability staining (Pore, 1990; Wenisch et al., 1997; Piña-Vaz et al., 2001). AmB is a commonly used and historically relevant polyene antifungal that associates with ergosterol to form transmembrane channels in the cell membrane. Monovalent ions (Na+, K+, H+, Cl−) leak from these channels and affect cellular homeostasis and, ultimately, viability (James and Rawlings, 1996). AmB has been used clinically for nearly 40 years to treat fungal infections, and emergence of resistant Candida isolates has been reported on rare occasions (Dutcher, 1968). Although it is generally accepted that fungi gain resistance to AmB by alterations in membrane lipid composition, the exact nature of these alterations remains a topic of debate (Athar and Winner, 1971; Hamilton-Miller, 1972; Sanglard et al., 2003; Ferreira et al., 2005). Regardless of the exact molecular mechanism underlying polyene resistance, considering the mechanism of action of AmB, sterol content or synthesis likely plays a role. The data presented in Fig. 6 provide strong evidence that C. albicans exposed to conditions of microgravity experience an increase in resistance to AmB. Future studies will be aimed at understanding the molecular mechanisms that are sensitive to LSMMG and result in this increased antimicrobial resistance.
5. Conclusions and Implications
Candida albicans appears to maintain a semi-stable phenotypic state while retaining the flexibility necessary for rapid adaptation to environmental stresses, including filamentation, biofilm formation, phenotypic switching, and changes in the expression of several hundred genes (Singh et al., 2005; Biswas et al., 2007; Noble et al., 2010). Data presented herein support a time-dependent, LSMMG-induced increase in cellular filamentation, altered colony morphology, biofilm formation and maturation, and antifungal resistance of C. albicans. Individually, and certainly in concert, these phenotypic transitions are indicative of organisms with increased propensity to cause infection and evade therapy. LSMMG appears to increase the potential for pathogenicity of C. albicans and, thus, may also serve as a model system for analyzing the low-shear in vivo environment often associated with C. albicans infections.
The possibility that environmental sensing of the microgravity/low-shear environment triggers a transition to a more pathogenic form of the organism could have clear implications for human health. The host-microbe interactions in spaceflight are important in their own right but can also be extrapolated to similar interactions associated with the common, ground-based population. The immune system becomes generally less resilient upon aging (Murciano et al., 2008). Furthermore, the number of immunocompromised individuals is increasing as a result of the use of chemotherapy and other immunosuppressant drugs (Concia et al., 2009; Cataldo and Petrosillo, 2011). Analysis of the effects of LSMMG on pathogen virulence could provide an informed foundation to identify astronaut risk, as well as provide better understanding of the more generally applicable molecular mechanisms involved in host-pathogen interactions.
Footnotes
Acknowledgments
We thank Betsey Pitts for assistance with microscopy and sample preparation and Larissa Jackiw for assistance with flow cytometry. We thank Scott Deis for his assistance with the colony morphology reversion studies. We appreciate the support of Dr. Bruce Granger for critically reading the manuscript. This work was supported by grant NNH08ZTT003N from the National Aeronautics and Space Administration.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
AmB, Amphotericin B; CIVS, cylindrical intravacuolar structures; HARV, high aspect ratio vessel; HIW, “hyper” irregular wrinkle; LSMMG, low-shear modeled microgravity; MAPK, mitogen-activated protein kinase; PKA, protein kinase A; qRT-PCR, quantitative reverse transcription–polymerase chain reaction.