
Abstract
The concept that life emerged where alkaline hydrogen-bearing submarine hot springs exhaled into the most ancient acidulous ocean was used as a working hypothesis to investigate the nature of precipitate membranes. Alkaline solutions at 25–70°C and pH between 8 and 12, bearing HS−±silicate, were injected slowly into visi-jars containing ferrous chloride to partially simulate the early ocean on this or any other wet and icy, geologically active rocky world. Dependent on pH and sulfide content, fine tubular chimneys and geodal bubbles were generated with semipermeable walls 4–100 μm thick that comprised radial platelets of nanometric mackinawite [FeS]±ferrous hydroxide [∼Fe(OH)2], accompanied by silica and, at the higher temperature, greigite [Fe3S4]. Within the chimney walls, these platelets define a myriad of micropores. The interior walls of the chimneys host iron sulfide framboids, while, in cases where the alkaline solution has a pH>11 or relatively low sulfide content, their exteriors exhibit radial flanges with a spacing of ∼4 μm that comprise microdendrites of ferrous hydroxide. We speculate that this pattern results from outward and inward radial flow through the chimney walls. The outer Fe(OH)2 flanges perhaps precipitate where the highly alkaline flow meets the ambient ferrous iron-bearing fluid, while the intervening troughs signal where the acidulous iron-bearing solutions could gain access to the sulfidic and alkaline interior of the chimneys, thereby leading to the precipitation of the framboids. Addition of soluble pentameric peptides enhances membrane durability and accentuates the crenulations on the chimney exteriors. These dynamic patterns may have implications for acid-base catalysis and the natural proton motive force acting through the matrix of the porous inorganic membrane. Thus, within such membranes, steep redox and pH gradients would bear across the nanometric platelets and separate the two counter-flowing solutions, a condition that may have led to the onset of an autotrophic metabolism through the reduction of carbon dioxide. Key Words: Dendrites—Framboids—Greigite—Hydrothermal chimneys—Mackinawite—Origin of life. Astrobiology 11, 933–950.
1. Introduction
B
Presuming such a scenario, flow reactor experiments were undertaken that involved the interaction (serpentinization) of mafic rock in the presence of iron monosulfide at 100 bar pressure and temperatures up to 120°C. These produced sulfidic and siliceous alkaline effluent (Mielke et al., 2010). As expected, both chimneys and geodes were generated on passing these solutions into a simulacrum of the ferrous iron-rich Hadean Ocean (Kump and Seyfried, 2005; Russell and Hall, 2006). Comparable membranes or walls to such compartments hold a potential of more than half a volt for several hours (Filtness et al., 2003; Russell and Hall, 2006). Such exothermic serpentinization also generates the fuels and some of the materials required for the onset of metabolism: H2, CH4, NH3, HS−, Mo, and W (Russell et al., 1994, 2010; Kelley et al., 2001, 2005; Proskurowski et al., 2006, 2008); and would deliver them in alkaline solutions to compartments at comparable temperatures to those exhaling at Lost City in the North Atlantic (average 70°C, Proskurowski et al., 2006) and Eyjafjordur in northern Iceland (also inferred to average 70°C, Marteinsson et al., 2001). Because the compartment walls would restrict diffusion of these hydrothermal fluids, they might be expected to focus reactions with the invasive carbonic ocean at, or within, their boundaries. Indeed, these boundaries may act as “low-entropy” traps whereby compatible products would be forced to interact through their proximity (Nitschke and Russell, 2010; Russell et al., 2010; Simoncini et al., 2010). Within limits, the smaller the compartment the more effective the trap. Crucially, without compartments there could be no focusing of the electrochemical gradients required to drive a metabolic pathway over the initial, thermodynamically unfavorable steps of carbon dioxide reduction (Maden, 2000). Nor could organic products be retained and concentrated for further interactions (Russell et al., 1988, 1994; Schoonen et al., 1999; Nitschke and Russell, 2009; Berg et al., 2010; Lane et al., 2010; Mast and Braun, 2010). Thus, the compartments are the devices that solve the thermodynamic problem met at the onset of metabolism (cf. Schrödinger, 1944; Sievers et al., 2004; Russell et al., 2010).
Such semipermeable and semiconducting compartment walls that comprise ferrous hydroxide, silica gel, and a complex of iron, nickel, and cobalt sulfides with adsorbed molybdenum or tungsten, or both, and act as a multifunctional catalyst would, along with their interiors, be the most beneficial sites for emergent biochemistry. The molybdenum or tungsten atoms act as two electron transfer agents; while one electron transfers to a high potential electron acceptor such as ferric iron or nitrate, the other now has enough energy to reduce carbon dioxide in an otherwise highly endergonic reaction (Russell and Hall, 1997, Maden, 2000; Ducluzeau et al., 2009; Nitschke and Russell, 2009). Although silica is not generally dissolved in alkaline spring waters exhaling from serpentinizing ultramafic terrains, it does occur to the extent of around 2 millimoles in such springs exhaling from basaltic crust and has been found at such a level in our own experiments with basalt (Kelley et al., 2001; Marteinsson et al., 2001; Mielke et al., 2010). Moreover, silica can be extremely enriched in alkaline springs that transect previously carbonated environments (Shibuya et al., 2010). Ephemeral carbonate (primarily as siderite, FeCO3) is also a possible initial precipitate, though it would soon be redissolved or transformed to a hydroxide around springs issuing from the seafloor into the deep (perhaps around 8 km) and mildly acidic Hadean ocean (Macleod et al., 1994; Bounama et al., 2001; Elkins-Tanton, 2008). Brucite, another hydroxide [Mg(OH)2], is also a likely precipitate (Kelley et al., 2001). We assume, in the absence of firm evidence, that the salinity of the early ocean was much like today's, because while the absence of evaporite basins in the Hadean might suggest higher concentrations, there was probably twice as much water on the planet at that time (Bounama et al., 2001; Elkins-Tanton, 2008; Tarduno et al., 2010). And given that the salinity of modern ocean water is little affected during hydrothermal circulation that feeds off-ridge springs (Kelley et al., 2001), we also assume an ocean water–like salinity of similar springs in the Hadean and thus no sodium cation gradient. The alternative view, that the first ocean was magnesium sulfate–dominated rather than sodium chloride–dominated, was not acted on in these experiments (Izawa et al., 2010).
It is notable that simple organic reactions of the kind that may have led to the emergence of metabolism are favored by hot alkaline conditions. For example, Huber and Wächtershäuser (1997) demonstrated the catalytic potential of entities such as NiS and NiSO4 in the production of activated acetate from methyl sulfide and carbon monoxide. The same authors showed that Fe(OH)2 is equally active as FeS in catalyzing the amination of other carboxylic acids to amino acids (Huber and Wächtershäuser, 2003). Yamanaka et al. (1988) dimerized glycines with trimetaphosphate in Mg2+-bearing alkaline solution at 38°C, and 4- to 6-mer peptides have been generated at the same temperatures but at the lower pH that might have occurred at the exterior of ancient hydrothermal mounds (Rabinowitz et al., 1969; Yamanaka et al., 1988; Yamagata and Inomata, 1997; Gao et al., 2008). In life itself, iron±nickel sulfide clusters, along with molybdenum or tungsten, play a critical role as active centers of proteins in many biological processes (Beinert et al., 1997; Adams, 1998; Hoke et al., 2004; Volbeda and Fontecilla-Camps, 2006; Rothery et al., 2008; McGlynn et al., 2009). For example, ferredoxin derivatives and iron-(nickel)-sulfur cluster proteins can act as hydrogenases; and electron donors, acceptors, and transfer agents in all domains of life are found even in the deepest evolutionary roots to both the bacteria and the archaea (Eck and Dayhoff, 1966; Hall et al., 1971; Baymann et al., 2003; Martin and Russell, 2007; Say and Fuchs, 2010). Even in the absence of organic ligands, metal sulfide clusters may have catalyzed the synthesis of simple organic molecules through reactions between the hydrogen, ammonia, and bisulfide produced from serpentinization reactions and the carbon dioxide and phosphate introduced from the Hadean ocean (Walker, 1985; Yamagata et al., 1991; Hagan et al., 2007; Smirnov et al., 2008)—reactions possibly energized by the ambient proton motive force and redox energies that acted across the inorganic membranes (Russell et al., 1994, 2010). In particular, the ambient proton motive force might have driven pyrophosphate synthesis which, in turn, drove the polymerization of amino acids within the membrane, thus improving its durability and functionality (Russell et al., 1994; Milner-White and Russell, 2010). These and other organic anions so generated would tend to have been trapped within the compartments and further concentrated through thermal diffusion, driven by the ambient temperature gradient that also acted across the margins of the hydrothermal mound (Baaske et al., 2007; Budin et al., 2009; Mast and Braun, 2010). Such interactions between a diverse set of reactants may have led to the onset of metabolism. While the metal sulfides had the makings of proto-metalloenzymes, the silica could have trapped and even helped in the synthesis of the sugars (Lambert et al., 2004, 2010). Eventually, RNA-based coenzymes might have replicated and then been polymerized through a natural convective polymerase reaction and concentrated through thermal diffusion (Baaske et al., 2007; Yarus, 2010). These RNA polymers may then have infected contiguous compartments and encoded the proteins that ushered in Darwinian evolution proper (Koonin and Martin, 2005). Such an evolutionary continuum, from the mineral to the living world, was first advocated in broad terms by Leduc (1911).
It is the aim of the present study to understand the effects of varying four important chemical conditions for iron sulfide or hydroxide precipitation in such a compartmentalized hydrothermal mound: (i) addition of silica in rising concentrations, (ii) pH levels, (iii) temperature, and (iv) the addition of pentameric peptides. One variable impossible to assess was the effect of pressure. However, given that no volatiles were involved in our experiments and only hydrogen and minor amounts of methane are evolved at Lost City and Eyjafjordur, this is unlikely to compromise our conclusions. Moreover, the effect of pressure on, for example, peptide stability, increases rather than decreases at higher pressure (Shock, 1992), which again is a trend that would be expected only to improve the results of our simulation.
2. Methods
In the laboratory simulation of alkaline seepages, aqueous alkaline solutions between 25°C and 70°C (70°C is the average temperature of submarine alkaline springs recorded so far) containing sulfide±silicate were injected into iron (II) solutions by way of motorized syringe pipettes or a peristaltic pump, at rates between 0.08 mL/h and 0.2 mL/h for the syringe (Fig. 1) and at 7.2 mL/h for the peristaltic pump. Chemicals, including FeCl2·4H2O and Na2S·9H2O, were purchased from VWR and used without further purification. Oxygen-free water (milli grade; 18.2 MΩ) was used throughout these experiments. Four main experimental approaches were taken, as follows: (i) sodium orthosilicate concentrations in syringe solutions were varied to observe the precipitated membrane mineralogical and structural changes; (ii) the pH within the injected solution was varied, and the resulting structures again were observed and analyzed; (iii) change of temperature; (iv) the effect on membrane structure was investigated through the addition of different simple and soluble pentameric peptides that comprise residues likely to have been generated early in the development of metabolism (Eck and Dayhoff, 1966; Hennet et al., 1992; Brooks et al., 2002; Davis, 2002). The growth, structure, and elemental composition of the resulting iron-sulfide precipitate membranes were analyzed by environmental scanning electron microscopy (ESEM), energy dispersive X-ray spectroscopy (EDX), X-ray powder diffraction, and Raman spectroscopy.

The experimental apparatus used for the production of metal-bearing chimneys. The crimp-top Wheaton bottle (125 mL) was cut open to more easily harvest the chimneys from the “Ocean” receptacle. Parafilm was tightly wrapped around the receptacle's open top, and a gaseous nitrogen purge line was used to create an oxygen-depleted headspace. All solutions were purged with gaseous nitrogen before use in experiments. A 10 mL syringe connected with Teflon tubing to a 1/16-inch (1.59 mm outer diameter) stainless steel tube was used to inject the “Hydrothermal” solution into the “Ocean” receptacle through the Teflon septa. Specially designed devices were used to minimize structural damage to the chimneys during harvesting and transport to the particular analytical instruments.
All receptacle and injection solutions were prepared simultaneously under the same environmental conditions at standard temperature and pressure (STP). Receptacle solutions were prepared inside glass Wheaton vials that had their base removed with a wet tile saw, were stoppered and inverted, and after the newly created opening was covered with parafilm, solutions were continuously sparged with gaseous nitrogen (N2) to create anaerobic solutions. Sparging was begun at least 30 min before any reactants were added to millipore (18.2 MΩ) water to displace dissolved oxygen. After water sparging, sodium chloride was added to each solution to achieve a 600 millimolar concentration similar to modern ocean water (see Introduction).
All pH values were determined with a Thermo Orion 550A pH meter. Injection solutions were pumped upward through an acid-cleaned (1% HCl; 3×H2O rinses) 1/16-inch stainless steel capillary tube into the receptacle solutions (Mielke et al., 2010, figure 8) with a peristaltic pump at a rate of 7.2 mL/h. For slower injection rates of 0.08 to ∼0.2 mL/h, a Razel Model A-99 syringe pump was used with a plastic 10 mL syringe and 16G1½ needle (Becton-Dickinson Medical). Experiments were duplicated or triplicated for each set of conditions. Similar runs were undertaken in a positive pressure nitrogen-filled glove box to check on any differences in product (data not shown). During the injection process, nitrogen gas was continually introduced into both solutions to ensure the exclusion of oxygen from the experimental environment and maintain positive pressure.
(i) Varying silicate concentrations. Primary reactants were added simultaneously to each solution to achieve a 10 millimolar iron (II) chloride receptacle solution (∼ pH 5.0) and a 10 millimolar sodium sulfide injection solution. The injection solutions were then slowly titrated with 4% hydrochloric acid to obtain a pH of 11 for all the silicate experiments. Once these constant reactants were completely dissolved, varying concentrations of sodium orthosilicate (Na4SiO4), ranging from 0.01 to 416 millimolar, were added to only the injection solution. This alkaline injection solution was allowed to flow into the receptacle solution for approximately 20–24 h at ∼0.2 mL/h.
(ii) Varying injection solution pH. Primary reactants were added simultaneously to each solution to achieve a 10 millimolar iron (II) chloride receptacle solution (∼ pH 5.0) and a 10 millimolar sodium sulfide injection solution. The injection solutions were then slowly titrated with 4% hydrochloric acid to reach a range of pHs from 7.98 to 11.7. For all values of alkaline pH tested, the sodium orthosilicate concentration was kept consistent at 0.1 millimolar.
(iii) Heated precipitation experiments. To better mimic the hydrothermal environment as described in the Introduction, receptacle solutions were heated to hydrothermal temperatures (70°C) before the injection solution was allowed to flow. An acidic salt solution (600 millimolar NaCl, 10 millimolar FeCl2 at pH∼5) mimicking the Hadean ocean was prepared in a 125 mL round-bottom flask with attached condenser and purged with N2. As described in previous experiments, an inverted glass bottle was used to prepare an oxygen-sparged alkaline hydrothermal solution containing 600 millimolar NaCl, 10 millimolar Na2S, and 2 millimolar Na4SiO4. The acidic solution was heated to 70°C by a resistor-controlled electric heating jacket. Once the solution reached 70°C, the pH of the solution was recorded. The alkaline hydrothermal solution was slowly injected at two different rates, ∼0.2 mL/h over a 20–24 h period (Mielke et al., 2010, figure 8) and ∼0.08 mL/h over a 68–72 h period.
Upon the formation of an inorganic tube or bubble (up to 20 h, Fig. 2a, 2b), injection was halted, and the precipitate sample was extracted into a glass vial via either a 1000 μL pipette whose disposable tip had been trimmed to make a large opening or a Gilson 5000 μL pipette. This was accomplished by placing the pipette tip around the chimney and simultaneously knocking the structure free while drawing the precipitate upward into the pipette, along with the surrounding solution. The resulting mixture was moved to an Eppendorf tube and allowed to settle. The majority of the solution phase was removed with a pipette, and 2.0 mL of oxygen-free water was added to rinse the precipitate. This rinsing process was performed four additional times to remove aqueous ions and minimize excess precipitation of reagent salts. The chimney was applied directly from this solution to an ESEM stub, or alternatively the vial was then sealed and purged with nitrogen gas for 10 min or until the sample was dry. Samples were then either immediately subjected to ESEM or Raman analysis or stored on the bench top for not more than 1 h under a nitrogen purge.

(
For ESEM analysis, samples were placed on an aluminum tab within 15 min and then inserted into the ESEM (FEI XL30 FEG) under a water vapor pressure of 3.8 torr, an accelerating voltage of 20 kV, and a working distance of 10.2 mm. The gaseous secondary electron detector was used at a chamber pressure of 3.8 torr to allow imaging without subsequent coating steps. The spot analysis feature was used for an energy dispersive X-ray spectrometer (EDAX, Inc.) to analyze X-ray signals with the Genesis program. Additional samples were washed, dehydrated, and embedded in Eponate resin before ultrathin sections of 60 nm were made and imaged with a scanning transmission electron microscope. An accelerating voltage of 30 kV and a working distance of 6.8 mm was used on the ESEM as described above (Werlin et al., 2011).
For X-ray powder diffraction analysis, the glass vials containing samples were immediately sealed after the three rinses and purged with nitrogen gas for 10 min or until the sample was dry. Samples were then placed on a sample cell between two layers of micro-thin Kapton film and placed under a Siemen's D500 diffractometer, equipped with a Cu anode tube. The energy beam was a Co-Kα Fe-filtered radiation. X-ray diffraction analysis was by a Peltier cooled Kevex Si(Li) (lithium-drifted silicon) X-ray detector. Mineral identification was made by reference to the 2009 ICDD database for organic and inorganic phases in conjunction with the JADE PLUS Version 9.1.1 XRD analysis program to identify the diffraction responses in the spectra.
Raman spectroscopy was performed on samples precipitated at both STP and 70°C. After injection, precipitated chimney structures were cut off from the steel capillary from which they grew with a steel scupula. Still intact, the chimney and remaining ocean solution were poured out into a Petri dish and rinsed six times with N2-purged Millipore (18.2Ω) water to remove excess salt. After extraction and rinsing, the precipitates were placed on a 3×1 glass slide and partially crushed with a spatula to expose the interior of the chimney. The slide was placed in a nitrogen-purged cryochamber at 10°C to limit oxidation of the sample. Raman spectra were obtained on a LabRam HR (Horiba Jobin Yvon) with use of a 532 nm laser and 600 g/mm grating with a resolution of ∼2 cm−1. The cryochamber was placed on a 3-axis motorized stage and the sample illuminated through a sapphire window. The instrument was calibrated by using the 520 cm−1 silicon line and zero order.
(iv) Addition of pentameric peptides. Primary reactants were added simultaneously to each solution to achieve a 10 millimolar iron (II) chloride receptacle solution (∼ pH 5.0) and a 5 millimolar sodium sulfide injection solution. The solutions involved in the peptide experiments were not manually buffered, and their initial pHs were measured as 4.1 and 11.7, respectively. Control experiments were performed without addition of peptide. For this series of experiments, a 5 millimolar sulfide solution was utilized in conjunction with a 10 millimolar ferrous iron solution, while concentrations of 5-mer peptides were added into the sulfide solution to obtain 100 micromolar concentration. Peptides were obtained from ANASPEC with no further purification. Peptides selected for investigation were AEGEA, EEGEE, and GGGGG on the basis of their simplicity, plausibility, and solubility (Hennet et al., 1992). All solutions were at a 600 millimolar sodium chloride concentration.
3. Results and Interpretation
3.1. Basic iron sulfide chimney structure
Analysis by ESEM and light microscopy revealed membrane bilayers with an overall thickness of ∼20 μm (though ranging between ∼4 and ∼100 μm). The inside is composed of globular clusters of higher sulfur content (always considered relative to iron) and the outside composed of more rigid shards of lower sulfur content (Figs. 2 and 3). The outermost crystalline layer's iron:sulfur ratios are relatively high (∼2.0 to 8.0), and the globular aggregations are relatively low (∼0.75–2.0). These atomic ratios lead us to assume that the outermost crystalline layers are primarily composed of ferrous hydroxide [Fe(OH)2] with an admixture of nanocrystalline ferrous sulfide [FeS], that is, mackinawite, whereas the globular clusters that comprise, or are attached to, the inner walls consist of mackinawite with greigite. These clusters, ∼1–10 μm in diameter, resemble framboids similar in appearance to those found in the interiors of fossil hydrothermal chimneys (Fig. 3c, 3d) (Larter et al., 1981; Boyce et al., 1983).

ESEM micrographs (
In experiments where no silica was added, ESEM revealed regular and symmetrical crenulations parallel to growth around and within the chimney walls (Fig. 4). At first, the crenulations reminded us of “capping and escape” effects comparable to chemical garden growth, whereby the top of the chimney is initially capped by a precipitate membrane that, in time, bursts or is at least sloughed off to the sides, leaving the outer edges as an additional rib to the growing chimney (cf. the growth rings of Stone and Goldstein, 2004). However, the crenulations' regularity, intricate structure that comprises well-ordered microdendrites (Fig. 4b), and sinuous appearance, along with the fact that periodicities were discerned at ∼4 μm (Fig. 11), all suggest a certain self-ordering—a physicochemical wave pertaining perhaps to the dissipation of the steep pH gradient (cf. Tóth et al., 2007, figure 2C). Indeed, the orientation of these dendrites normal to banding suggests a response to inward and outward diffusive flow through the chimney walls (cf. Jeong et al., 2001) (Fig. 2d). The crenulations were produced both at the slower and faster pump rates.

ESEM micrographs of the flanged walls to iron sulfide/hydroxide chimneys produced when 5 millimolar alkaline sulfide solution is injected into a slightly acidic 10 millimolar iron (II) chloride solution. Experiments were conducted at a rate of 7.2 mL/h with a peristaltic pump. Micrographs (
Under injection at 0.2 mL/h, chimneys were observed to completely cap off with a substantial covering of iron sulfide precipitate, which halted elongation (Fig. 5). This process is thought to start off in the same way as the aforementioned capping and escape cycle, with the exception that the fluid pressure is dissipated through the greater area of iron sulfide membrane in longer chimneys. This pressure dispersal obviates bursting, though it causes the cover to thicken and strengthen from additional layers of precipitates. This type of permanent capping has been followed by spongiform overgrowth in some cases (Fig. 2A). Experiments where chimneys appeared capped usually also spawned secondary chimneys branching from their sides or base (Fig. 5). This suggests that, on fouling, the pressure once again exceeds chimney strength and causes a portion of the membrane to burst, which thereby allows further exhalation and chimney growth. We could view this phenomenon as revealing how reproduction was initiated in the mound and how organic molecules generated in one compartment could be transported to the offspring.

Series of frames (top) captured from video of separate experiments with varying injection solution pH (i.e., sodium sulfide solution). Notice the increasing bulbous appearance and eventual deterioration of the FeS precipitate as pH is lowered and the lack of chimney formation that occurs at pH 8.03. Above pH 11 the exterior membrane surfaces comprise green rust (iron oxyhydroxides), and chimneys are characteristically taller and often branched. The graph (bottom) shows the ESEM–energy dispersive X-ray spectroscopy (EDX) analytical results for iron:sulfur atomic ratios of multiple analyses per chimney cross section and multiple chimneys at the various pHs. Note that this experiment was run under ambient laboratory conditions with reactant concentrations specified in the Methods section. For a scale of reference, the stainless steel tubing possesses an outer diameter of 1/16 inch (1.59 mm) (3 mm scale bar). Color images available online at
3.2. Varying injection solution pH
According to the ESEM-EDX data, chimneys formed from injected solution with a pH that exceeded 11 are composed of ferrous oxyhydroxide. Below pH 11, the membranous precipitates mainly comprise stoichiometric FeS (Fig. 5, graph), a result consistent with expected FeS/Fe(OH)2 equilibria. However, the lower the pH, the weaker and more bulbous is the structure (Fig. 5). These bulbous structures also possess more fibrous clusters of precipitate on the outside than the standard chimneys. Based on observations of broken samples and ESEM images of bulb walls, these fibers are also hollow. Because of their smaller size relative to the chimneys, it appears that the membranes of the bulbs are more permeable than the chimney membranes, or at least more fluid and less brittle, as they do not rupture from injection pressure. Under ESEM, they are visibly more permeable and spongy. Below a pH of 8.03, no precipitate formed, neither was sulfide found present in the solution mixture, having volatilized as hydrogen sulfide gas.
3.3. Varying sodium orthosilicate concentrations and temperature
Increasing the concentration of sodium orthosilicate (Na4SiO4) generated more bulbous or geodic structures (Fig. 6). It appears that a concentration of 6 millimolar Na4SiO4 generates Fe:S ratios that approach 1:1, which effectively facilitates iron monosulfide precipitation, whereas higher concentrations appear to facilitate ferrous hydroxide precipitation.

Series of frames (top) captured from video of separate experiments involving various concentrations of sodium orthosilicate added to the sodium sulfide solution at an unadjusted pH of ∼11.7 and injected into a solution of iron (II) chloride. The left-hand data point here is equivalent to right-hand data point in Fig. 5. The precipitate structures become more bulbous with increasing sodium orthosilicate concentrations. The graph (bottom) of ESEM-EDX analyses of iron:sulfur atomic ratio indicates that the highest sulfur content relative to iron is found in these structures when the concentration of sodium orthosilicate in the sodium sulfide injection solution is ∼7.15 millimoles/liter. For a scale of reference, the stainless steel tubing possesses an outer diameter of 1/16 inch (1.59 mm) (3 mm scale bar). Color images available online at
Under ESEM, the membranes of the silicate-induced bulbous structures are quite similar to those membranes created by lowering the injection solution pH. The mineralogy of the iron sulfide nanocrystalline precipitates was investigated with X-ray powder diffraction spectrometry. Mackinawite was strongly indicated with some suggestion of greigite (Fig. 7). Raman spectroscopy revealed three peaks in the spectra that correspond to crystalline mackinawite at STP, and the higher-temperature (70°C) chimneys were found to be, as expected, partially composed of greigite (Figs. 8 and 9) (Krupp, 1994; Lennie et al., 1997; Rickard et al., 2001; Bourdoiseau et al., 2008, 2010; Rémazeilles et al., 2010).

X-ray diffraction spectra of a precipitate formed via the injection of 2.0 millimolar sodium orthosilicate and 10.0 millimolar of sodium sulfide solution into a receptacle solution of 10 millimolar iron (II) chloride at STP. The main mineral phase was found to be mackinawite. The two strong peaks at ∼32° and ∼46° are associated with unknown minerals that may be forms of ferrous silicate (cf. Bourdoiseau et al., 2008; Rémazeilles et al., 2010). All solutions contain 600 millimoles/liter sodium chloride. Color images available online at

Raman spectrographic analysis shows evidence for partially oxidized mackinawite (iron (III)-containing mackinawite) in iron sulfide chimney grown at 70°C. The small shoulder peak at ∼309 cm−1 and the broad hump centered around ∼354 cm−1 are similar to the spectral peaks identified by Bourdoiseau et al. (2008) as representing partial oxidation of iron (II) in mackinawite to iron (III). The partial oxidation of iron (II) is demonstrated to lead to the formation of greigite (Rickard et al., 2001, and see Fig. 9). Color images available online at

Raman spectrographic analysis shows evidence for greigite and native sulfur in iron sulfide chimneys grown at 70°C. Greigite is expected at higher temperatures through the loss of iron to solution, and the native sulfur is possibly produced by the same process, though we cannot rule out the production of sulfur by oxidation occurring during transfer of the sample to the Raman system. The large greigite peak around 350 cm−1 is separated into two bands occurring at ∼351 and ∼352 cm−1, closely resembling the greigite Raman spectrum of Rémazeilles et al. (2010, and see Wei and Osseo-Asare, 1995; Benning et al., 2000; Rickard et al., 2001.) Color images available online at
3.4. Effects of pentameric peptides on structure
Experiments in which peptides were added to the injected alkaline solutions were conducted to investigate their effects as metal chelating agents on growth, structure, and morphology at STP. Three contrasting pentameric peptides (AEGEA, EEGEE, or GGGGG) were added at 100 micromolar concentration to 5 millimolar sulfide-bearing alkaline solution at STP. This composite solution was then injected into a 10 millimolar ferrous iron solution at a flow rate of 7.2 mL/h. Tubular structures comprising multiple distinct iron sulfide mineralogical phases were produced as judged by ESEM contrast as well as EDX. Sulfur-rich iron sulfide spheres, again with the appearance of framboids and Fe:S ratios of ∼1.5 to 3, and darker, nonspherical material of lower Fe:S ratios of around 6, were the major constituents observed. However, in examples where 5-mer glycine was added, particular locations had Fe:S ratios that approached 18 (Fig. 10C). The other pentameric peptides induced Fe:S ratios that reached as high as ∼8 and ∼12 (Fig. 10A and 0B, respectively). Addition of these peptides also enhanced the development of the crenulations on the outer walls of the chimneys (compare Fig. 4 with Fig. 10). Thus, results of these experiments indicate that iron:sulfide ratios of the mixed mineral precipitate increase with the addition of the three peptides, though the effect of the 5-mer glycine was most marked (Fig. 10C).
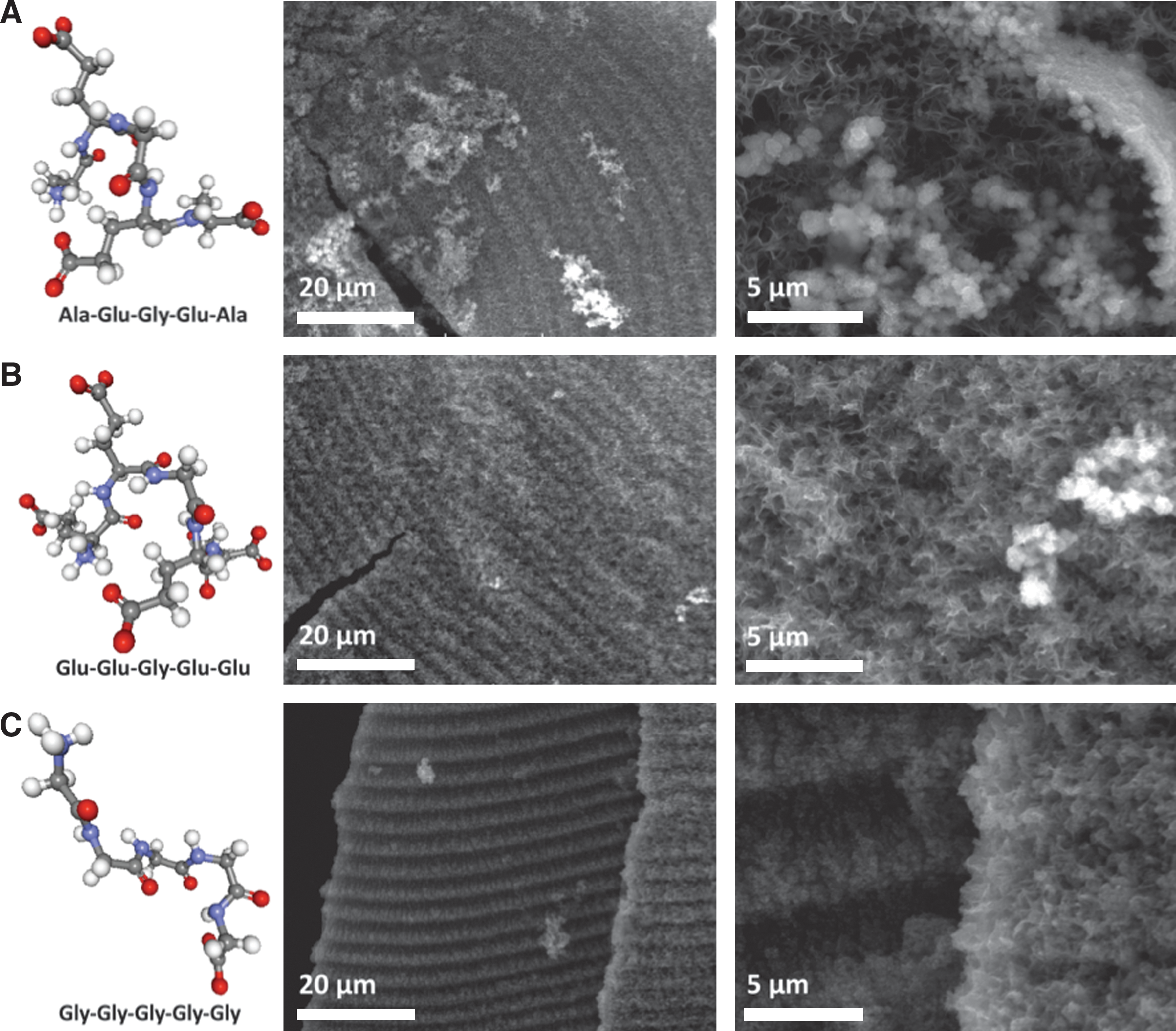
ESEM images of flanged walls on ferrous sulfide/hydroxide chimneys grown by the injection of a 5 millimolar alkaline sodium sulfide (pH∼11.7) solution bearing 100 micromolar pentameric peptides into a 10 millimolar slightly acidic iron (II) chloride solution. The chimneys were produced at an injection rate of 7.2 mL/h, and the different pentameric peptides are identified below the ball-and-stick illustrations. All three peptide experiments formed flanged walls that had a similar periodicity (wavelength) of ∼4 μm. The three peptides gave structures topographically unique from one another as judged by ESEM. All solutions contained 600 millimolar sodium chloride and no sodium orthosilicate. Ball-and-stick images produced with BALLView 1.4.
The small peptides utilized in these studies comprise the simplest and earliest amino acids (Brooks et al., 2002; Davis, 2002). Apart from pentaglycine, these peptides contain a number of functional groups capable—as multi-dentate carboxyl, amine, and carbonyl ligands—of coordinating metal ions. Thus, each peptide could potentially ligate to several metals or other inorganic entities with multiple bonds (Harford and Sarkar, 1999; Watson and Milner-White, 2002; Cox et al., 2005; Milner-White and Russell, 2008; Torrance et al., 2009).
The propensity of peptides to chelate metal ions in solution and at solution/mineral phase boundaries, together with the observed change in morphological and elemental composition of iron sulfide/hydroxide chimneys, suggests the possibility that these polymers direct epitaxial growth of particular iron-rich minerals by specific metal ion chelation. As an initial iron sulfide zone is formed, polymers present in the spring solution may be exposed at the liquid-membrane interface. In addition to this, the high porosity of these structures allows the possibility that, as the solution continues to flow, organic polymers are transported into the membrane. These processes together would make for an iron sulfide wall that bears an organic functionality capable of metal ion chelation. The presence of these metal cation-binding sites in the form of an organic matrix increases local concentrations of iron near the outer wall, which creates a unique chemical environment that contrasts with environments where organic functionalities are absent (cf. Sikes et al., 1991; Silverman et al., 1995).
4. Discussion
The inorganic compartments produced in these experiments appear to satisfy one of the expectations of the alkaline hydrothermal theory for the emergence of life in that they could act as chemiosmotic and semiconducting containers for organic reactions and their reactive products (Russell et al., 1994; Lane et al., 2010). Moreover, we know from previous studies that the compartments can hold a potential of more than half a volt for several hours, in theory quite enough to contribute to a putative autotrophic metabolism (Filtness et al., 2003; Russell and Hall, 2006). However, their thicknesses, from 4 to 100 μm, are generally orders of magnitude greater than the 5–10 nm typical of the prokaryotic membrane. Thus, at first sight the pH and redox gradients would be too low to drive some of the otherwise endergonic reactions that frustrate the earliest steps in CO2 reduction (Maden, 2000; Michalkova et al., 2011). Regarding pH, it might be expected that protons find their way through the membrane to the alkaline interiors, perhaps via rotational/translational diffusion of water/hydronium molecules that comprise the Langmuir layer on sulfide, hydroxide, or silica surfaces oriented normal to the membrane (Fig. 2) (da Silva and Williams, 1991, p 103). Of course, the pH gradient is logarithmic, and thus a fairly steep gradient will obtain at some locale within the membrane. Judging from comparable studies by Bähr et al. (1984), this is likely to be situated toward the outermost margins of the membrane. And like any response to ionic gradients, the protonic flow is likely to be as waves and as such carry packets of energy along the wave fronts (cf. Stone and Goldstein, 2004). As the membranes themselves are spatially heterogeneous at the micron level and perhaps below, the transportation of ionic species would likely be heterogeneous as well, resulting perhaps in the buildup of steep gradients at some locales. At the same time, the two electrons stemming from hydrothermal hydrogen and reducing molybdenum (or tungsten) VI to IV bifurcate and could be conducted along the iron layers (Fe-Fe=2.6 Å) or tetrahedra that comprise the “ab” crystallographic plane of mackinawite (Russell and Hall, 2006; cf. Ferris et al., 1992; Kwon et al., 2011) from the interior, one to an electron acceptor such as ferric iron phases, sulfur, nitrate, or nitric oxide on the exterior, while the other now has the energy to reduce CO2 (Russell and Hall, 1997; Ducluzeau et al., 2009; Nitschke and Russell, 2009). In this way, molybdenum-bearing mackinawite nanocrysts—possibly ligand modified—act as a protohydrogenase (Russell and Hall, 2002; Nitschke and Russell, 2009; and see McGlynn et al., 2009).
Results of these experiments also reveal patterns of precipitation of the kind normally considered to result from conditions far from thermodynamic equilibrium, that is, from supersaturation attendant upon fluid mixing. The first example is that of the framboids found within the chimney interiors, which presumably comprise aggregates of magnetic FeIII-bearing mackinawite and greigite (Sawlowicz, 2000; Mitra-Delmotte and Mitra, 2010; cf. Larter et al., 1981) (Figs. 3, 8, and 9). The second is the periodic equi-spacing of the crenulations that mark the exteriors and interiors of many of the chimney walls when grown from more alkaline solution or on those where peptides were added to the influx (Figs. 4 and 0). Given the fidelity and regularity of the crenulations and their orthogonality to flow, an early thought was that they were Chladni plates (Lévy, 2006, figure 1), that is, artifacts that respond to vibrations transmitted along the capillary tube that feeds the receptacle solution. However, because these structures were generated in experiments involving both peristaltic and syringe pumps, and because we employed different lengths of plastic tubing, all giving the same length between rill and ridge, we considered the crenulations more likely to be an outward manifestation of hidden chemical self-ordering. After all, flanges at much larger scale around hydrothermal chimneys that comprise beehive heat exchangers are clearly generated far from equilibrium (Fouquet et al., 1993, figure 4; Koski et al., 1994; Rickard et al., 1994, figure 1; Ludwig et al., 2006, figure 2b; Rickard and Luther, 2007, figure 39). Those so-called beehives consist of regularly spaced ribs of anhydrite, pyrite, and iron oxides that encircle a black smoker chimney. The conditions are imposed by the temperature, redox, and pH contrasts across the chimney walls. Fluid flow is organized radially both inward and outward. Rickard et al. (1994) suggested that anhydrite, being inversely soluble, is precipitated along with iron oxides as seawater—presumably drawn toward the chimney by the Venturi effect—is heated by the outgoing hydrothermal fluids from which pyrite and chalcopyrite are precipitated. Once the pattern is set, it is reinforced by feedback effects to be expected of far-from-equilibrium systems. We speculate that the regularly spaced flanges around our micro-chimneys are a result of flow and counterflow, guided and reinforced through a comparable positive feedback. In this case, the outflow of hydroxyl and hydrosulfide ions of the alkaline solution perhaps leads to the precipitation of the external ferrous hydroxide ridges that comprise the rows of microdendrites, while the inflow of hydronium may be focused in the rills between. We envisage this two-way ionic and electron transport through the chimney wall as being controlled by spontaneously precipitated central dividers 10–100 nm thick that comprise platelets of FeS or Fe(OH)2, or both, and separate the permeable channels ∼2 μm deep (Fig. 11). If this tentative interpretation were to prove correct, the scene would be set for the reduction of CO2 or hydrothermal formate and, concomitantly, a high potential electron acceptor, within the nanopores of the membrane rather than in the compartment's interior. Regarding CO2 reduction, the simplest pathway for emerging biochemistry is via two tributaries, one the hydrogenation of carbon dioxide to a methyl entity and the other to form a carbonyl group. These components are then assembled as activated acetate at acetyl coenzyme-A synthase, the nickel iron sulfide–bearing enzyme that gives this pathway its name (Fuchs, 1989; Ljungdahl, 1994; Russell and Martin, 2004).
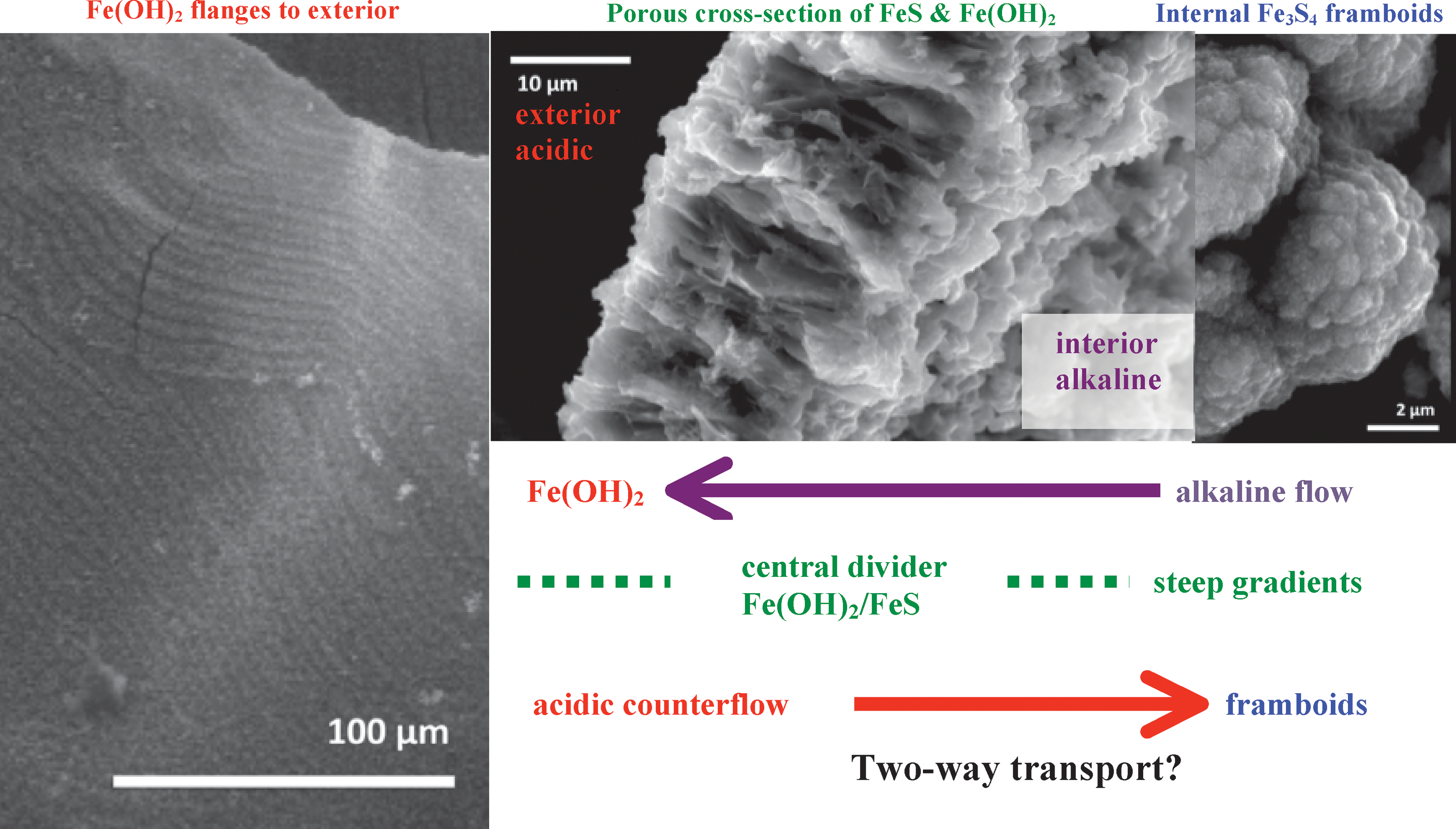
Composite selection of micrographs illustrating the regular flanges that comprise ferrous hydroxide microdendrites (see Fig. 2d for detail) on the exterior of an iron sulfide–bearing membrane (left), a cross section (middle), and framboidal iron sulfides from the interior of a chimney (right). We speculate that flange growth results from the escape of alkaline solution through the membrane along subcompartments oriented normal to growth. Where this solution meets the ferrous iron, ferrous hydroxide is precipitated as ribs, also normal to growth. A counterflow of protons down-gradient to the alkaline interior takes place in the neighboring subcompartments. Platelets of ferrous minerals are spontaneously precipitated at the interface of the two solutions. This is where we imagine the potential exists for carbon dioxide reduction/hydrogenation under the higher pressures obtaining early in the history of wet, rocky worlds—an idea amenable to testing through microfluidics.
While inorganic compartments formed at particular hot springs on early Earth may offer a plausible site for life's emergence (Koonin, 2007), and the hypothesis examined here requires a minimum of contingency, it must be pointed out that sulfide-rich deposition at alkaline springs would have been rare, relying as it would on extensive developments of sulfides in the oceanic crust beneath (Mielke et al., 2010; Russell et al., 2010). More typical would have been precipitates of silica, brucite, and ephemeral carbonates (Shibuya et al., 2010). Ferrous hydroxide and green rust would have been rarer additions to the precipitate mounds, and sulfide deposits rarest of all. Nevertheless, given the thousands of such submarine springs operating at any one time, the hundreds of millions of years between the condensation of the first ocean and the Archean with its signs of life, and the billions of compartments that comprised any one mound, there was an abundance of possibilities for this kind of metabolic process to emerge and evolve into life. The same might be said for Mars and the icy wet moons of Jupiter and Saturn—Europa and Enceladus—prime targets for life-detection missions (Russell and Hall, 1999; Hand et al., 2010; Postberg et al., 2011).
5. Conclusions
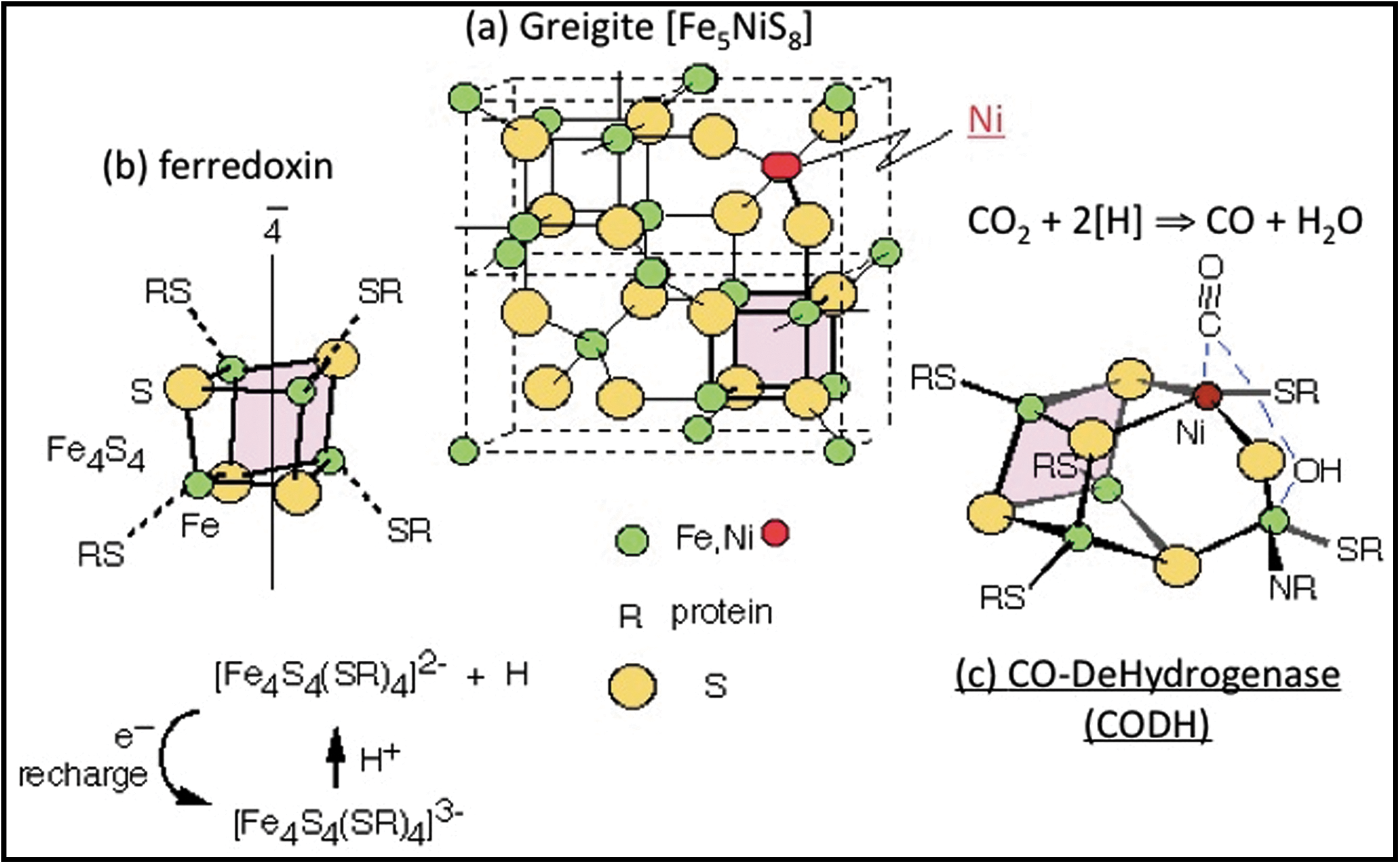
Iron sulfide and hydroxide precipitates formed in experiments that simulated particular, and probably rare, ancient submarine alkaline hydrothermal vents occurred as chimney-like tubes and geodic bubbles. At STP, the exteriors were composed dominantly of crystalline mackinawite (FeS), often with an exterior of dendritic ferrous hydroxide, while framboidal-like iron sulfide spheres tended to coat the interiors. In the higher-temperature chimneys, some of the mackinawite was transformed to greigite (Fe3S4). From a prebiotic perspective, we note that mackinawite has a similar conformation as does the active center of iron-iron hydrogenase, albeit without ligand modification, while the structure of greigite is similar to the active centers of acetyl coenzyme-A synthase and carbon monoxide dehydrogenase as well as to the ferredoxins (Russell and Hall, 1997, 2006; McGlynn et al., 2009) (Fig. 12).
Iron sulfide–bearing precipitates are influenced greatly by the overall chemical environment. Atomic sulfur ratios relative to iron increase with the decrease of injection solution pH (minimum 8.03) and also with the increase of sodium orthosilicate concentrations (maximum 6 millimolar). Moreover, observed differences in cases in which peptides were added to the alkaline solutions reveal a novel functionality for these molecules in that they may have participated as binders and modifiers to chimney structure and reactivity prior to their contemporary roles in biological processes (Milner-White and Russell, 2008; Rabone et al., 2010). The diversity of structural and mineralogical morphologies and elemental stoichiometries of iron and sulfur are consistent with theories in which these elements played a fundamental role in the emergence of chemical complexity through the dissipation of physico- and electrochemical gradients (Russell and Hall, 1997, 2006; Baaske et al., 2007). That is, together they provide for a scenario whereby the close proximity of a number of different mineral types and forms creates a foundry of chemical potential through which the catalysis and gradient-driven chemical condensations required for the interconversion of chemical species could be wrought. In addition, the existence of gel-like to nanocrystalline chimney compartments allows for the sequestering and concentration of reaction products. Put another way, the hydrothermal conduits and their superposed chimneys, geodes, and gels focus and confine the kind of high enthalpy, but low entropy, hydrothermal feed that interacts with the carbonic ocean through the chemiosmotic and semiconducting inorganic membranes that could promote chemical ordering and the emergence of biochemistry through the acetyl coenzyme-A pathway—“order from order” as Schrödinger (1944) would have it (Russell et al., 1994; Nitschke and Russell, 2010; Shock and Canovas, 2010; Stockbridge et al., 2010).
This work suggests that the main site for reduction and polymerization was within the porous and semipermeable membranes themselves rather than in the compartmental voids. However, the compartments may have acted as repositories for some of the products, which in turn may have been entrained and passed on upward by the hydrothermal solution to newly formed compartments, so enriching these in organic anions such as peptides. These peptides may have continued to chelate the ready-made inorganic clusters that acted as ever more effective catalysts or protoenzymes. Finally, these may have coated the inorganic interiors, so bringing about an organic takeover and the formation of budding organic cells (Milner-White and Russell, 2008).
We reiterate that geochemistry could not emerge into biochemistry without the extra energies afforded by the chemiosmotic forces and electron bifurcations that act through the kind of inorganic membranes described here (Russell et al., 1994; Russell and Hall, 1997; Nitschke and Russell, 2009, 2011; Lane et al., 2010; Michalkova et al., 2011). Precipitating at a pH front between the injected alkaline solutions that represent a hydrothermal spring and the ocean-like solvent, there exists the potential across iron sulfide–bearing membranes for relatively steep electrochemical and thermal gradients, for the concentration and storage of organic molecules and an ambient proton motive force, which all autotrophic life appears to exploit (Kell, 1979; Baltscheffsky et al., 1999; Mast and Braun, 2010). These are challenges for experimentation. Further research is also called for to examine whether protons could be put to work condensing monophosphate and acetyl phosphate to pyro- or trimetaphosphate as a natural proton-motive force (Milner-White and Russell, 2010; Barge et al., 2011; cf. de Zwart et al., 2004).
We emphasize that, dependent as it would be upon source rock, temperature, depth, and perhaps proximity to a 400°C hydrothermal spring discharging high concentrations of the d-block elements and a subsidiary quantity of hydrogen sulfide (Douville et al., 2002), the generation of iron sulfide/hydroxide–rich chimneys would have been a rather rare, but inevitable, event at submarine alkaline springs. We say this in the knowledge that there would have been hundreds of submarine alkaline springs operating over the hundreds of millions of years that comprised the Hadean era, each one producing a complex of billions of assorted compartments over their lifetimes of tens of thousands of years (Früh-Green et al., 2003; Ludwig et al., 2006), some of them reproducing and promoting the kind of protometabolic reactions that we consider to have led to the emergence of life.
Footnotes
Acknowledgments
We thank Bill Abbey, Mathieu Choukroun, James Gleixner, and Richard Kidd for help in the laboratory. For discussions we are grateful to Wolfgang Nitschke, Laszlo Roszlo, and Professor Galen Stucky, as well as Laurie Barge, Elbert Branscomb, and Eugenio Simoncini, members of the NAI-sponsored Thermodynamics Disequilibrium and Evolution Focus Group. K.R. thanks the National Aeronautics and Space Administration New York Space Grant Consortium for financial sponsorship. L.W. is supported by the ConvEne IGERT Program (NSF-DGE 0801627). S.E.M. was supported by the Marine Biological Laboratories NASA Planetary Biology Internship Program, an NSF IGERT Fellowship by the MSU Program in Geobiological Systems (DGE 0654336) and acknowledges the NASA Astrobiology Institute–Montana State University Astrobiology Biogeocatalysis Research Center (NNA08CN85A). The research described in this publication was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration, with support by NASA Exobiology and Evolutionary Biology award (NNH06ZDA001N) and supported by the NASA Astrobiology Institute (Icy Worlds).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
EDX, energy dispersive X-ray spectroscopy; ESEM, environmental scanning electron microscope, environmental scanning electron microscopy; STP, standard temperature and pressure.