
Abstract
Tardigrades are tiny (less than 1 mm in length) invertebrate animals that have the potential to survive travel to other planets because of their tolerance to extreme environmental conditions by means of a dry ametabolic state called anhydrobiosis. While the tolerance of adult tardigrades to extreme environments has been reported, there are few reports on the tolerance of their eggs. We examined the ability of hydrated and anhydrobiotic eggs of the tardigrade Ramazzottius varieornatus to hatch after exposure to ionizing irradiation (helium ions), extremely low and high temperatures, and high vacuum. We previously reported that there was a similar pattern of tolerance against ionizing radiation between hydrated and anhydrobiotic adults. In contrast, anhydrobiotic eggs (50% lethal dose; 1690 Gy) were substantially more radioresistant than hydrated ones (50% lethal dose; 509 Gy). Anhydrobiotic eggs also have a broader temperature resistance compared with hydrated ones. Over 70% of the anhydrobiotic eggs treated at either −196°C or +50°C hatched successfully, but all the hydrated eggs failed to hatch. After exposure to high-vacuum conditions (5.3×10−4 Pa to 6.2×10−5 Pa), the hatchability of the anhydrobiotic eggs was comparable to that of untreated control eggs. Key Words: Tardigrades
1. Introduction
Ultimately, if we are to assess the ability of tardigrades to survive transfer among planets or to thrive in extreme environments, they must be able to reproduce. Thus, the present study focuses on the hatchability of tardigrade eggs after exposure to environmental extremes. In contrast to considerable studies of the tolerance in adult tardigrades, the survival of anhydrobiotic tardigrade eggs after exposure to the vacuum of space, space UV radiation, and cosmic radiation has only been investigated in two studies. Jönsson et al. (2008) reported that desiccated adults of two tardigrade species, Richtersius coronifer and Milnesium tardigradum, survived exposure to the vacuum of space, and a small portion of the tested individuals survived the combined exposure to the vacuum of space and UV radiation for 10 days in low Earth orbit. In addition, they demonstrated that desiccated eggs exposed to the vacuum of space alone hatched at the same level as the controls (Jönsson et al., 2008), which suggests that tardigrade eggs in anhydrobiosis tolerate space environmental stresses. In another flight experiment, Persson et al. (2011) demonstrated that eggs of M. tardigradum tolerated low levels of cosmic radiation (less than 4 Gy) during a 10-day flight in low Earth orbit. Also, eggs of Ramazzottius oberhauseri in anhydrobiotic state survived for a longer period of time, with a maximum of 9 years, than did adult Ramazzottius oberhauseri and other tardigrade species (Guidetti and Jönsson, 2002). Here, we set out to estimate the reproductive potential of tardigrade eggs in possible extraterrestrial environments by measuring their hatchability after exposure to ionizing radiation, temperature, and vacuum.
2. Materials and Methods
2.1. Animal culture and egg collection
The tardigrade species Ramazzottius varieornatus, which was originally collected from moss samples in Hokkaido, Japan (43°03′48″N, 141°22′25″E), was used in this study. The life cycle of R. varieornatus was revealed in our previous study (Horikawa et al., 2008). Juvenile R. varieornatus become mature adults subsequent to the molting processes (ecdysis). On average, the life span of R. varieornatus is 35 days, the number of eggs produced by an animal is 7.85, and the hatching time of eggs is 5.7 days when the animals are cultured at 25°C (Horikawa et al., 2008). In the present study, tardigrades were cultured at 22–25°C on 1.5% w/w agar plates with distilled water and the green alga Chlorella vulgaris (Chlorella Industry Co., Ltd., Tokyo, Japan) as food. Culture dishes were checked at 1–2 day intervals, and newly laid eggs were transferred onto fresh agar plates containing distilled water but not algae. Dark-colored eggs that failed to develop were excluded, and normally developing eggs (2–3 days post-deposition) were used for experiments.
2.2. Preparation of anhydrobiotic egg samples for exposure experiments
Here, we employed several desiccation methods that are slightly different among types of exposure experiments as described below. This was due to a limited availability of incubators with identical temperature, a difference in sample stage in each apparatus among those exposure experiments, and a requirement for sample transportation in the vacuum exposure experiments.
Anhydrobiotic eggs for scanning electron microscopy (SEM) were prepared by placing them on double-sided plastic tape and desiccated at 25°C under 85% relative humidity (RH) for 1 day, followed by 0% RH for more than 2 days. RH was controlled by glycerol (Invitrogen, Carlsbad, CA, USA) in water according to Johnson (1940).
For the irradiation experiment, 15–20 hydrated eggs in 100 μL of distilled water were placed on filter paper (1.33 g, diameter 35 mm) in a plastic Petri dish (diameter 35 mm) and desiccated at 25°C under 0% RH for 2–7 days.
For the temperature experiment, 15–30 hydrated eggs in 100 μL of distilled water were placed on filter paper (0.55 g, 2×2 cm) in a plastic Petri dish (diameter 35 mm) and desiccated at 25°C under 33.8% RH for 1 day and then desiccated at 25°C under 0% RH for 4–6 days in a desiccator.
For the vacuum tolerance experiments, 25–100 hydrated eggs in 100 μL of distilled water were placed on filter paper (0.55 g, 2×2 cm) in a plastic Petri dish (diameter 35 mm) and desiccated at 22°C under 33.8% RH for 5 days and then kept at 22°C under 0% RH for 9 days.
It should be noted that, irrespective of desiccation methods, the difference in the hatchability of control anhydrobiotic eggs was statistically insignificant (Tukey's multiple comparison test, p>0.05: 88.8±5.8% for SEM, 93.9±0.9% for irradiation experiments, 89.7±8.4% for temperature experiments, and 87.6±7.6% for vacuum experiments; values are the means of three independent experiments).
2.3. Scanning electron microscopy
We examined the morphology of hydrated or anhydrobiotic eggs of R. varieornatus with SEM. Hydrated eggs in 500 μL distilled water in a 1.5 mL microtube were frozen at −80°C in a freezer and then dried in a vacuum system (FDU-810, EYELA, Tokyo, Japan). Anhydrobiotic eggs were prepared as described above. Both hydrated and anhydrobiotic egg samples were coated with gold or platinum palladium and observed with the VE-7800 scanning electron microscope system (Keyence, Osaka, Japan).
2.4. Irradiation
Hydrated eggs were placed on filter paper with 200 μL of distilled water in a plastic Petri dish (diameter 35 mm). The Petri dishes were covered with a 7 μm thick polyimide Kapton film (Dupont-Toray, Tokyo, Japan) to prevent air-drying, and the film was sealed with parafilm (Pechiney Plastic Packaging, Inc., Chicago, IL, USA) on the side of the dishes. Anhydrobiotic eggs were placed in a similar way except without water. Both hydrated and anhydrobiotic egg samples were irradiated with 250–2000 Gy of 4He (50 MeV, 16.3 keV/μm) delivered from the azimuthally varying field cyclotron installed at the Takasaki Ion Accelerators for Advanced Radiation Application (TIARA) facility, Japan Atomic Energy Agency, as described previously (Hamada et al., 2006; Horikawa et al., 2006). The absorbed dose was calculated as follows: Dose (Gy)=1.6×10−9×LET (keV/μm)×fluence (particle/cm2). Within 1 h after irradiation, 2 mL of distilled water was added to the dishes, and then the eggs were incubated at 22°C.
In this experiment, three replicates were considered. To determine the 50% lethal dose (LD50), the linear regression analysis was performed between the dose and the hatchability. Spearman's correlation coefficient by rank test was used to test correlations between the dose and the hatchability. Hatchability was compared between anhydrobiotic eggs and hydrated ones by using a chi-square test with a Yates' correction.
2.5. Temperature experiments
In eighteen 1.5 mL microtubes, hydrated eggs were suspended in 1 mL of liquid paraffin (Wako, Tokyo, Japan). In a second set of 18 microtubes, anhydrobiotic eggs were also suspended in 1 mL of liquid paraffin. Three microtubes from each of the two sets were then exposed to a temperature of −196°C, −80°C, +22°C, +50°C, +80°C, and +100°C, all for a total of 1 h. For exposure to −196°C, microtubes containing the egg samples were instantaneously frozen in liquid nitrogen. For the samples to be exposed to −80°C, they were placed in a freezer. Cooling rates were −140±6.1°C/min in liquid nitrogen and −7.7±1.0°C/min in the freezer, as determined by recording temperature inside a microtube with a thermocouple element connected to a logger inserted in liquid paraffin. In the case of +50°C, +80°C, and +100°C, the microtubes were heated in a dry heat block. Samples kept at +22°C (room temperature) were considered as controls.
The microtubes were then removed from the devices and left in ambient atmosphere for up to 2 h to set back to room temperature. After being washed one time in distilled water, the eggs were transferred to Petri dishes (diameter 35 mm) and then rehydrated with 2 mL of distilled water. Three independent experiments were carried out for each temperature condition. A chi-square test with a Yates' correction was used to compare the survival between anhydrobiotic and hydrated groups.
2.6. Vacuum experiments
Anhydrobiotic samples were placed in a vacuum chamber where the atmosphere was decompressed from atmospheric pressure with a turbo-molecular pump (PT-500, Mitsubishi Heavy Industries, Ltd., Tokyo, Japan) connected to the chamber. An ion gauge (GI-TL3, ULVAC, Inc., Kanagawa, Japan) inserted in the chamber indicated that pressure in the vacuum chamber was 5.3×10−4 Pa and 6.2×10−5 Pa at 1 h and 7 days, respectively, after starting decompression. Control eggs were kept at room temperature under atmospheric pressure at 0% RH for 7 days. After returning to atmospheric pressure, egg samples were kept with silica gel in a tightly enclosed plastic bag. The samples were rehydrated with 2 mL of distilled water 7 days after shipment from H. Hashimoto's laboratory (Tsukuba University, Japan) to L.J. Rothschild's laboratory (NASA Ames Research Center, USA).
Three independent replicate experiments were performed. The survival of eggs exposed to vacuum and those that comprised the untreated control were compared by a chi-square test with a Yates' correction.
2.7. Egg hatching
After rehydration, egg samples were transferred with a glass pipette onto agar plate dishes (diameter 35 mm) containing a small amount of distilled water that covered the agar surface with a thin film. For the irradiation experiment, eggs were observed under a stereomicroscope with transmitted illumination (SZ60, Olympus, Tokyo, Japan, or STEMI 2000-C, Zeiss, Jena, Germany) every day. For the temperature and vacuum experiments, they were viewed under the stereomicroscope at 1–5 day intervals. Since the maximum development time of R. varieornatus eggs that has been reported is 9 days after deposition (Horikawa et al., 2008), the egg hatching was observed up to 10 days after rehydration. Hatched juveniles were removed for the convenience of counting newborns each day.
3. Results
3.1. Morphology of anhydrobiotic eggs
The typical shapes of hydrated and anhydrobiotic eggs desiccated under laboratory conditions are shown in Fig. 1. The desiccated eggs showed an erythrocytic form with the projections bent onto the egg surface (Fig. 1A), while hydrated eggs had a spherical shape with numerous projections (Fig. 1B).
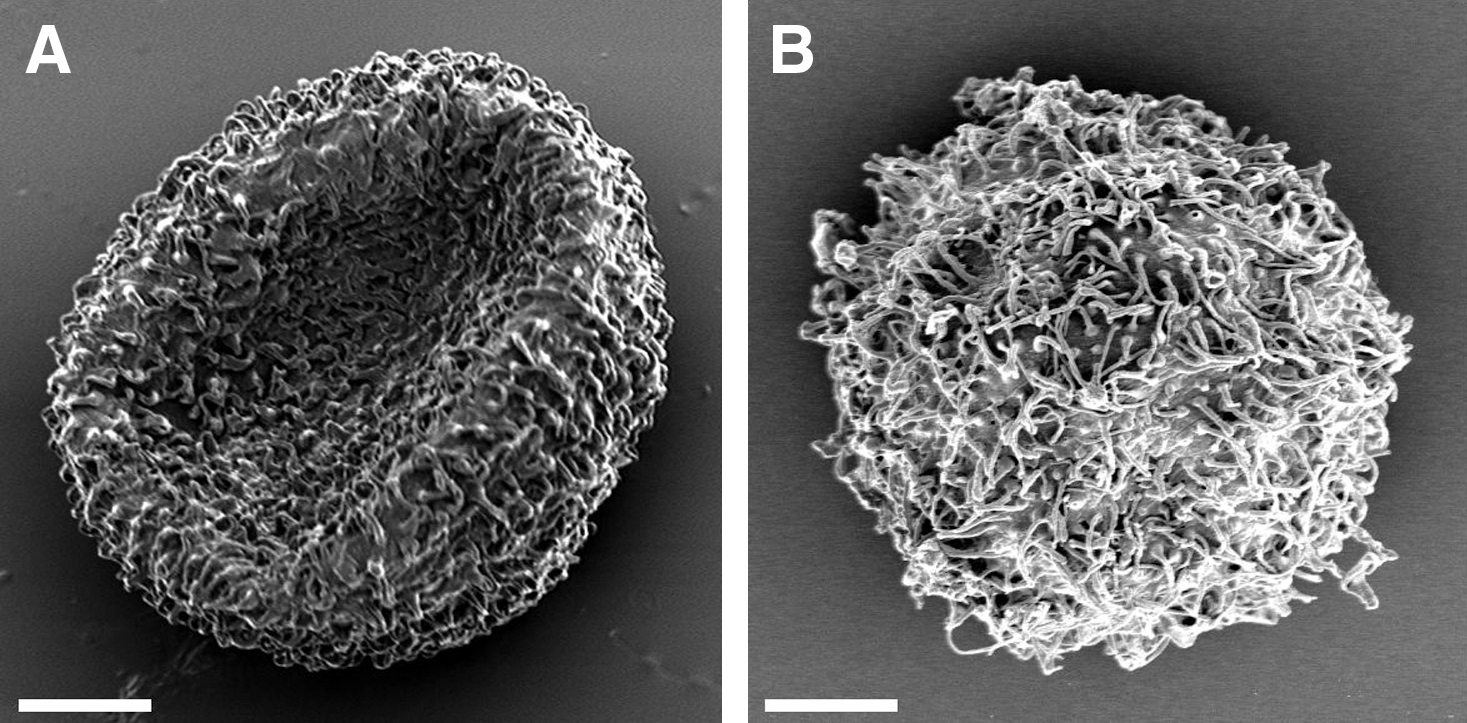
The representative scanning electron micrograph of anhydrobiotic eggs (
3.2. Radiation tolerance of tardigrade eggs
The hatchability of anhydrobiotic eggs was over 80% at doses up to 1000 Gy, whereas that of hydrated eggs was decreased from 100% at 0 Gy to approximately 5% at 1000 Gy in a dose-dependent manner (Fig. 2). Irradiation with over 1000 Gy decreased the hatchability of anhydrobiotic eggs to approximately 30% at 2000 Gy. The hatchability of anhydrobiotic eggs was significantly higher than that of hydrated ones at all radiation doses tested (P<0.001), and the LD50 values were 509 Gy in hydrated eggs and 1690 Gy in anhydrobiotic ones.

Changes in hatchability following 4He irradiation (250–2000 Gy) of anhydrobiotic and hydrated eggs of the tardigrade R. varieornatus. Error bars indicate plus and minus one standard deviation (N=3–4). Asterisks indicate significant differences in hatchability between anhydrobiotic and hydrated eggs (*P<0.001).
3.3. Temperature tolerance
The hydrated eggs hatched successfully at +22°C, while only a very small proportion of eggs hatched after the +50°C treatment (Fig. 3). In contrast, in anhydrobiotic eggs, the hatchability was relatively high (>79.0%) over a broad temperature range, from −196°C to +50°C, although it markedly decreased at +80°C (Fig. 3). None of 100°C-treated eggs hatched. Anhydrobiotic eggs hatched at a significantly higher rate than hydrated ones at low (−196°C and −80°C) and high (+50°C and +80°C) temperatures.
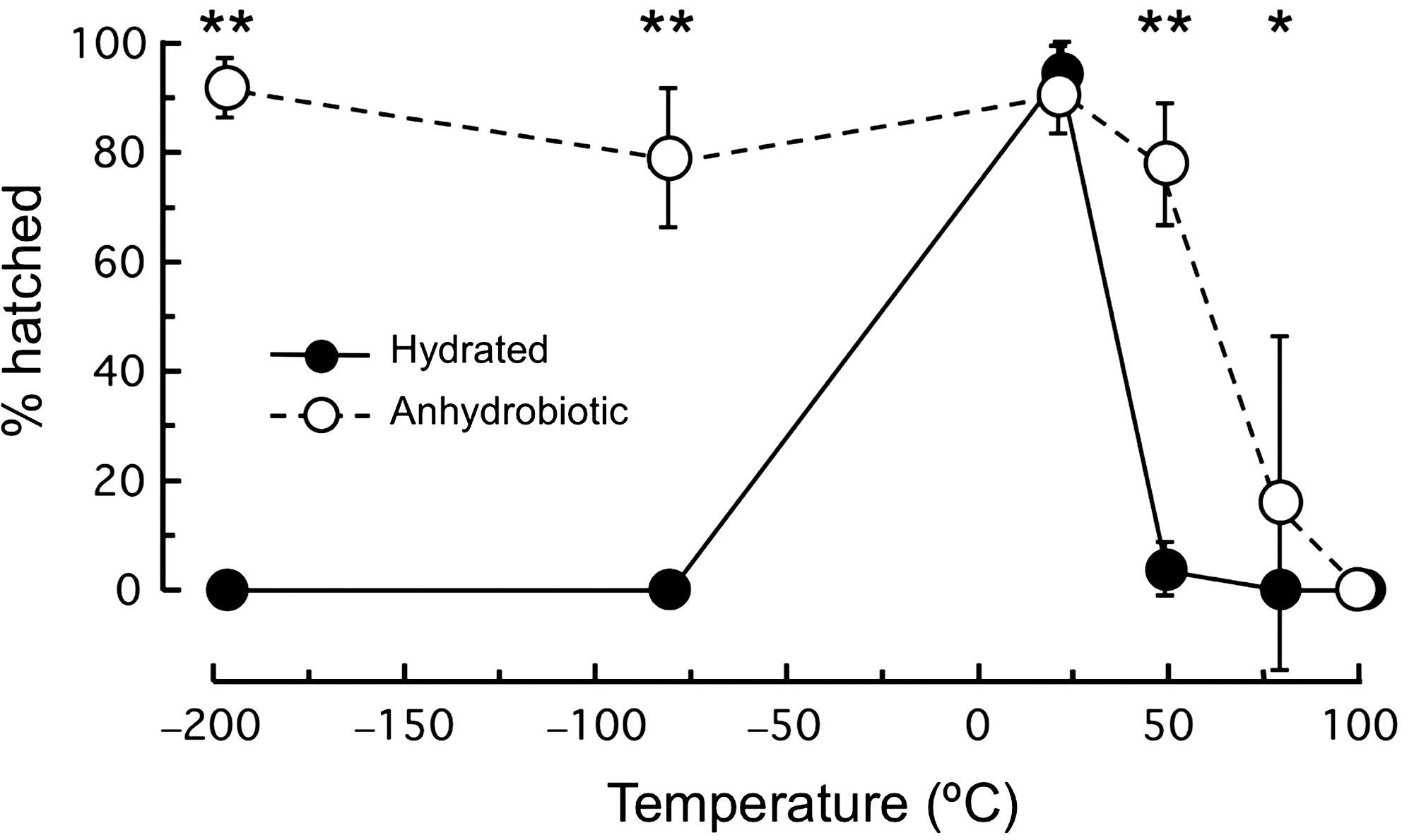
Changes in hatchability of anhydrobiotic and hydrated eggs of R. varieornatus following exposure to a series of temperatures (−196°C to +100°C) for 1 h. Eggs were observed up to 10 days after exposure to temperatures, and mean hatchability was determined for both anhydrobiotic and hydrated groups. Error bars indicate plus and minus one standard deviation (N=3). Asterisks indicate significant differences in hatchability between anhydrobiotic and hydrated eggs (chi-square test with a Yeats' correction; *P<0.05, **P<0.001).
3.4. Vacuum tolerance
There was no significant difference (P=0.71) in the hatching percentage between vacuum treatment (85.9±13.1%) and the control under atmospheric pressure (87.6±7.7%) (Fig. 4).

The ability of anhydrobiotic eggs of R. varieornatus to hatch after exposure to vacuum (5.3×10−4 Pa and 6.2×10−5 Pa) and kept under atmospheric pressure (control) for 7 days. Error bars indicate plus and minus one standard deviation (N=3). There was no significant difference in hatchability between anhydrobiotic and hydrated eggs (chi-square test with a Yeats' correction, P>0.05).
4. Discussion
Anhydrobiotic animals generally show morphological changes when they enter into an anhydrobiotic state. For example, while entering anhydrobiosis, juvenile and adult tardigrades contract their body into a barrel-like shape (tun) (Baumann, 1927; Wright, 1989), and nematodes transition into a coiled shape (Crowe and Madin, 1975). Eggs of R. varieornatus could hatch even after 10-day preservation under extremely dried conditions that caused them to enter into anhydrobiosis (Horikawa et al., 2008).
This study demonstrated that the anhydrobiotic eggs showed cross-tolerance to various extreme environments, as was the case for the adults. It should be noted, however, that a slight difference in the drying conditions from those that were used here could affect the speed of evaporation from the eggs during desiccation. This could result in different degrees of egg tolerant ability in extreme environments among the exposure experiment groups; therefore, careful consideration would be required for direct comparison of the survival ability of the anhydrobiotic eggs among the various exposure experiments. However, the eggs used in this study that underwent dehydration in RH 0% are considered to have entered into an anhydrobiotic state, judging from the desiccating methods used in our previous study (Horikawa et al., 2006). To overcome these issues in future studies, a universal method for desiccating tardigrades should be established.
It is thought that removal of free water from an anhydrobiotic body may fundamentally confer on tardigrades robustness against extreme environmental stressors (Seki and Toyoshima, 1998). Solar and cosmic ionizing radiation is an important potential risk for various organisms in outer space. The irradiation experiments in the present study showed that LD50 values of eggs of R. varieornatus against 4He ions were 1690 Gy in anhydrobiotic eggs and 509 Gy in hydrated ones, which indicates that hydrated eggs are more radiosensitive than anhydrobiotic ones (Fig. 2). This result is consistent with the radiation sensitivity of Artemia salina (brine shrimp) eggs, which, when fully hydrated, are less tolerant to gamma-irradiation than desiccated ones (Engel and Fluke, 1962). On the other hand, hydrated adult tardigrades have been shown to be highly radioresistant, as have anhydrobiotic tardigrades (May et al., 1964; Jönsson et al., 2005; Horikawa et al., 2006, 2008). It is known that cells of tardigrades at the adult stage show less mitotic activity (Bertolani, 1970), and the absence of somatic cell division may lower the lethal effects of radiation (Ducoff, 1972). Tardigrade eggs kept in an anhydrobiotic state for 9 years can successfully hatch following rehydration (Guidetti and Jönsson, 2002), so it is obvious that the cell cycle in these eggs is perturbed during anhydrobiosis. Thus, the higher radiation tolerance anhydrobiotic eggs exhibited over that of hydrated eggs, as shown in this study, might be the result of cell cycle arrest during anhydrobiosis. Jönsson et al. (2008) reported that eggs of R. coronifer and M. tardigradum that were exposed to 4.5 mGy of cosmic radiation combined with the vacuum of space during a 10-day flight in low Earth orbit hatched, as did anhydrobiotic eggs kept at normal pressures. Persson et al. (2011) demonstrated that anhydrobiotic eggs of M. tardigradum that were exposed to 4 Gy of cosmic radiation in low Earth orbit successfully hatched as well. In addition, Rebecchi et al. (2009, 2011) reported that both hydrated and anhydrobiotic adults of tardigrade Macrobiotus richtersi survived irradiation with 1.9 mGy of ionizing radiation inside the spacecraft Foton-M3 during a 12-day flight in low Earth orbit. Results of the present study suggest that anhydrobiotic eggs of tardigrades tolerated much higher doses of cosmic radiation than did the tardigrades that were exposed in low Earth orbit in previous studies.
The present study is the first to evaluate the hatchability of tardigrade eggs following exposure to extremely low and high temperatures. As is the case for anhydrobiotic adults (Becquerel, 1950; Ramløv and Westh, 2001; Horikawa et al., 2008), eggs of R. varieornatus in an anhydrobiotic state tolerate low and high temperatures to a higher degree than do hydrated ones (Fig. 3). Iwasaki (1973) reported that approximately 65% of a sum of dry eggs of A. salina hatched after exposure to 100°C for 1 h, whereas no R. varieornatus eggs hatched with the same exposure (Fig. 3). Although this result may suggest that brine shrimp have a higher tolerance to a temperature of 100°C than do tardigrades, the difference in the way the media in the present study was heated and the way it was heated in the Iwasaki (1973) study might have affected survivability. The liquid paraffin used in the present study has a higher heat transfer efficiency compared to that of air, which was what was used by Iwasaki (1973), who did not utilize a special medium to suspend the heat-treated Artemia eggs. The tardigrade eggs used in this study should, therefore, have received more heat energy, which resulted in lower survivability. It is likely that anhydrobiotic tardigrade eggs are more tolerant to low temperature than they are to high temperature (Fig. 3), as are adult anhydrobiotic tardigrades (Becquerel, 1950; Horikawa et al., 2008). Given this capacity for anhydrobiotic eggs to tolerate a wide range of temperatures, anhydrobiotic eggs could survive temperatures at the martian surface, which range from −123°C to 25°C (Diaz and Schulze-Makuch, 2006). Moreover, the anhydrobiotic tardigrade eggs, juveniles, and adults could survive temperatures below −269°C (Horneck, 1999) in interplanetary space, since it is expected that even such extremely low temperatures would not change the structure of biomolecules in anhydrobiotic organisms.
The vacuum level of open space causes extreme desiccation and is therefore lethal for most organisms. In the present study, the anhydrobiotic eggs of R. varieornatus tolerated high vacuum for 7 days (Fig.4). This is consistent with the results of Jönsson et al. (2008), who reported that anhydrobiotic eggs of other tardigrade species hatched after exposure of 10 days to the vacuum of space. For assessing the effects of high vacuum on anhydrobiotic tardigrades, a longer period of time would be needed to estimate survivability more precisely, since exposure to vacuum for several years is known to reduce survivability, probably due to the accumulation of DNA strand breaks, even in spores of Bacillus subtilis, one of the known most vacuum-resistant bacteria (Horneck, 2003).
Jönsson et al. (2008) demonstrated that adults and eggs of anhydrobiotic tardigrades can tolerate the space environment and that UV radiation is the most severe environmental factor to animals under their flight experiment conditions. In addition, Altiero et al. (2011) showed that exposure to more than 75 kJ/m2 of UV radiation dramatically reduced the survival fraction of adult tardigrades in anhydrobiotic state. In this study, we showed that anhydrobiotic eggs of tardigrades survived extremely low temperatures and space-level vacuum.
Considering the critical effects of UV radiation on tardigrades, these animals may have the capacity to complete interplanetary travel only when they are protected inside a rock that serves as shielding against UV radiation. In fact, when covered with a Mars analog regolith in a Mars-simulated environmental chamber, 70% of adult specimens of R. varieornatus in anhydrobiotic state survived exposure to 3.7×107 J/m2 of UVC radiation (Johnson et al., 2011), a dose that corresponds to more than 1000 times the UV dose that caused the killing of all anhydrobiotic eggs of tardigrades in outer space (Jönsson et al., 2008).
Extremely low temperatures do not seem to reduce survival of anhydrobiotic R. varieornatus, as suggested by this study and our previous study (Horikawa et al., 2008). Anhydrobiotic tardigrades appear to survive longer at low temperature (Sømme and Meier, 1996) than at room temperature (Rebecchi et al., 2006). Tardigrades in the anhydrobiotic state survived up to 9 years under room-temperature conditions (Guidetti and Jönsson, 2002), but it is expected that they can survive much longer at the extremely low temperatures found in outer space. In addition, Gusev et al. (unpublished data) demonstrated that anhydrobiotic larva of the insect Polypedilum vanderplanki showed better survival in the vacuum of space than under atmospheric pressure, which suggests that the vacuum of space is a favorable environmental condition for anhydrobiotic survival. In this regard, evaluation of the tolerance of tardigrades to prolonged exposure to space-level vacuum seems particularly important. This warrants future experiments in which tardigrades would be packed into a rocklike material and exposed to prolonged open space environments. But the survival of tardigrades in space is only of significance to panspermia if they are able to reproduce once they reach a favorable environment. This study provides support for the possibility of successful transfer in the vacuum of space by showing that indeed anhydrobiotic eggs were substantially more resistant to radiation than hydrated eggs.
Footnotes
Acknowledgments
This work was supported by the REIMEI research resources of JAERI (7) and TIARA Cooperative Research Program (51047). This research was also supported by an appointment to the NASA Astrobiology Institute NASA Postdoctoral Program by Oak Ridge Associated Universities. We thank T. Kubo of The University of Tokyo for daily assistance and encouragement and K. Shimada of Hokkaido University for assistance with recording temperatures inside samples in the temperature experiment. We also thank M. Yamashita of JAXA, A. Yamagishi of Tokyo University of Pharmacy and Life Science, and K. Kobayashi of Yokohama National University for arrangement of vacuum exposure experiments.
Abbreviations
LD50, 50% lethal dose; LET, linear energy transfer; RH, relative humidity; SEM, scanning electron microscopy.