
Abstract
In the Universe, oxygen is the third most widespread element, while on Earth it is the most abundant one. Moreover, oxygen is a major constituent of all biopolymers fundamental to living organisms. Besides O2, reactive oxygen species (ROS), among them hydrogen peroxide (H2O2), are also important reactants in the present aerobic metabolism. According to a widely accepted hypothesis, aerobic metabolism and many other reactions/pathways involving O2 appeared after the evolution of oxygenic photosynthesis. In this study, the hypothesis was formulated that the Last Universal Common Ancestor (LUCA) was at least able to tolerate O2 and detoxify ROS in a primordial environment. A comparative analysis was carried out of a number of the O2-and H2O2-involving metabolic reactions that occur in strict anaerobes, facultative anaerobes, and aerobes. The results indicate that the most likely LUCA possessed O2-and H2O2-involving pathways, mainly reactions to remove ROS, and had, at least in part, the components of aerobic respiration. Based on this, the presence of a low, but significant, quantity of H2O2 and O2 should be taken into account in theoretical models of the early Archean atmosphere and oceans and the evolution of life. It is suggested that the early metabolism involving O2/H2O2 was a key adaptation of LUCA to already existing weakly oxic zones in Earth's primordial environment. Key Words: Hydrogen peroxide—Oxygen—Origin of life—Photosynthesis—Superoxide dismutase—Superoxide reductase. Astrobiology 12, 775–784.
1. Introduction
Nevertheless, there are some geochemical and geophysical data, theoretical model considerations, molecular phylogenetic and biochemical comparative analyses that suggest a higher oxygenation of both the young Earth's surface and atmosphere than is commonly accepted (McKay and Hartman, 1991; Castresana et al., 1994; Towe, 1996; Draganić, 2005; Hoashi et al., 2009; Kornaś et al., 2010; Haqq-Misra et al., 2011). The modeling of the photochemistry of some realistic atmospheres (including present and past Earth and Mars) has revealed O2-rich atmospheres (up to 5%) as a result of abiotic H2O and CO2 photolysis (Selsis et al., 2002). It has also been proposed that abiotically produced reactive oxygen species (ROS), such as H2O2, could be the primordial source of molecular oxygen (O2). This assumption has been supported by experiments that have shown that pyrite/aqueous suspensions generate H2O2 in the absence of O2 (Borda et al., 2001). Additional arguments that suggest a more oxidized state of the young Earth may be adduced from astrophysical studies concerning the surface of Mars. The Viking mission in the 1970s showed that the surface of Mars is highly oxidized (Klein, 1978; Hartman and McKay, 1995), in contrast to its atmosphere, which contains only traces of molecular oxygen (ca. 0.13%) and a vast amount of CO2 (ca. 95%) (Moroz, 1998; Selsis et al., 2002). Recently, a geochemical model for the pyrite-dependent aqueous formation of ROS, such as OH· and H2O2 in the subsurface of Mars, has been presented (Davila et al., 2008). Houtkooper and Schulze-Makuch (2007) suggested that hypothetical martian organisms might use a H2O2-H2O mixture as an intracellular fluid to be optimally adapted to the highly oxidized martian environment. Moreover, it has been reported that the surface of Europa, the icy satellite of Jupiter, contains H2O2, and both radiolysis and photolysis of its icy surface leads to the formation of oxygen and ROS (Carlson et al., 1999; Johnson et al., 2003). Recently, the presence of abiotically formed O2 has also been found in the atmosphere of Saturn's icy moon Rhea (Teolis et al., 2010). These data are important in the context of hypothetical life-forms present on Europa (Chyba and Phillips, 2001), which could be equipped with some components of enzymatic antioxidant systems. All these very speculative models and some experimental data may suggest that the early stages of biological evolution on Earth occurred in a more oxic environment.
A view commonly presented in handbooks is that primordial organisms were strict anaerobes (Halliwell and Gutteridge, 1999; Alberts et al., 2002; De Las Rivas et al., 2004; Shaw, 2008). At present, there is a growing number of fully sequenced genomes of organisms that belong to three domains of life (Archaea, Bacteria, and Eukarya). This has allowed comparative analysis of the biochemical processes of the hypothetical Last Universal Common Ancestor (LUCA) and the metabolic background of presently living organisms (Peregrin-Alvarez et al., 2003; Ouzounis et al., 2006). Currently, we have access to primary databases of the small-molecule metabolism (e.g., BioCyc, KEGG), containing many experimentally verified metabolic pathways, along with enzyme information derived from the scientific literature (Caspi et al., 2008; Kanehisa et al., 2008).
Anaerobes, mainly prokaryotic organisms, still exist today; but as a rule, their growth is inhibited, or they are killed by exposure to the present O2 atmospheric level (Halliwell, 2006). Nevertheless, the available literature and data stored in public domains indicate that components of antioxidant response systems are present in strict and in facultative anaerobes (Brioukhanov and Netrusov, 2004; Imlay, 2008). With this in mind, we have formulated the following hypotheses: (1) LUCA was not an obligate anaerobe, but it had genes that encoded enzymes that utilized ROS, for example, H2O2, and the earliest protocells were able to tolerate O2; (2) on the ancient surface of the young Earth, ca. 3.5 Ga, microenvironments could have existed where minor, but sufficient, amounts of ROS and O2 were generated in the generally anoxic environment of early Earth. In silico analysis of selected O2/H2O2-dependent pathways deposited in the primary BioCyc database (Caspi et al., 2008) was used to test whether the above hypothesis could be a promising pointer for further studies.
2. Materials and Methods
The comparative analysis was performed according to BioCyc (
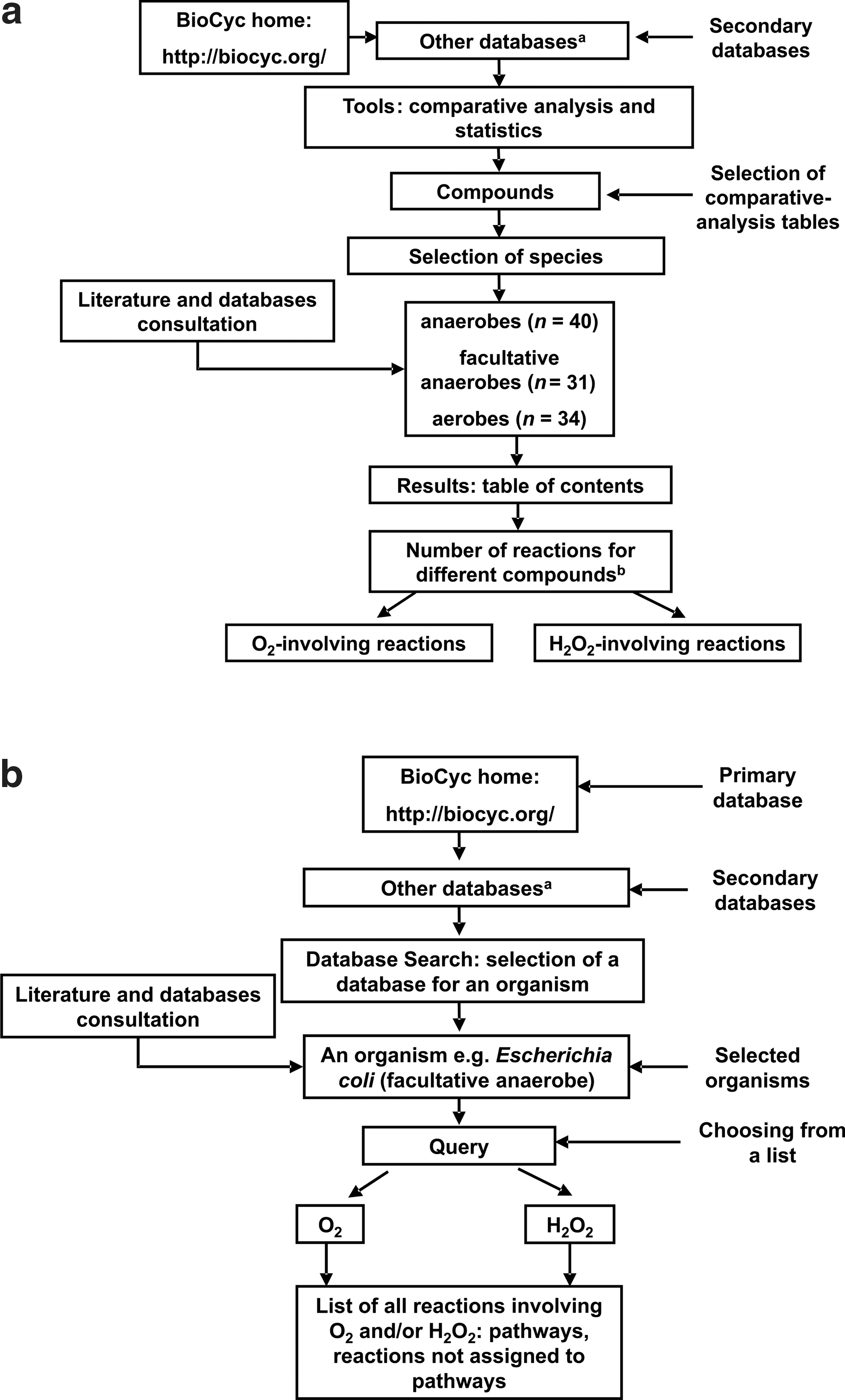
Two schemes (
The classification of organisms by oxygen tolerance was derived from database NCBI Microbial Genome Project—http://www.ncbi.nlm.nih.gov/genomes/lproks.cgi—and from the literature (Vieira-Silva and Rocha, 2008; Mentel and Martin, 2008).
Organisms performing anoxygenic photosynthesis.
Organisms performing oxygenic photosynthesis.
where xi
is the number of O2/H2O2-involving reactions for the i
th organism derived from databases (Table 1), xt(i)
is the total number of all enzyme-catalyzed reactions that utilize unique small molecules for the i
th organism, and n is the entire number of analyzed organisms of each group: anaerobes, facultative anaerobes, and aerobes (Table 1). Moreover, two crucial biochemical pathways where O2/H2O2 is involved were selected: (1) aerobic respiration, (2) removal of ROS. Simplified criteria for linking the above pathways to the organisms listed in Table 1 were used. In this study, the aerobic metabolism as a process of electron transport to molecular oxygen as a final acceptor was defined. Finally, all the anaerobes and facultative anaerobes that have the terminal electron acceptors known as dioxygen reductases were classified as organisms that show aerobic respiration. The reduction of O2 to water is catalyzed by dioxygen reductases belonging to two superfamilies, cyt. bd oxidases and heme-copper oxidases, for example, cytochrome c oxidases (Pereira et al., 2001; Brochier-Armanet et al., 2009), according to the reaction:
Among analyzed obligate and facultative anaerobes (Table 1), the percentage of organisms possessing at least one type of dioxygen reductase was derived not only from the BioCyc database but also from molecular phylogenies obtained previously by Blank (2009) and Brochier-Armanet et al. (2009). A ROS removal system was defined by the occurrence of at least one of the antioxidant enzymes, superoxide dismutase (SOD), superoxide reductase (SOR), or catalase (CAT). These enzymes are responsible for the detoxification of ROS, such as superoxide anion radical and H2O2, in the following reactions:
In light of this classification, “complete” aerobes are organisms having both aerobic respiration and ROS removal pathways.
This estimation was used to clarify the data presentation and to discuss the obtained results. It was assumed that all the analyzed aerobes take advantage of aerobic respiration and a ROS removal system. In the case of anaerobes and facultative anaerobes, the percentage of all organisms having these pathways was determined by the equation:
where n p is the number of analyzed organisms possessing the components of a pathway, that is, aerobic respiration or/and a ROS removal pathway, and n is the total number of analyzed organisms, anaerobes, facultative anaerobes, and aerobes (listed in Table 1).
Statistical analysis was performed by using a nonparametric Kruskal-Wallis test. Dunn's test was used as a post test.
3. Results
3.1. Mean number of reactions involving O2 and H2O2
An analysis of 105 different organisms was performed, including 40 obligate anaerobes, 31 facultative anaerobes, and 34 aerobes (Table 1).
The global comparison of total amount of O2- and H2O2-involving reactions showed that aerobes perform significantly more reactions where O2 and H2O2 are engaged than obligate anaerobes (Fig. 2). On the other hand, no significant differences were found between facultative anaerobes and aerobes, as far as reactions involving O2/H2O2 are concerned (Fig. 2a, 2c). If the comparative analysis was carried out with use of the percentage of O2-involving processes, a significant increase in such reactions was observed between all the analyzed groups of organisms (Fig. 2b). A significantly higher number of H2O2-involving reactions identified in aerobes in comparison to obligate anaerobes was observed only when the data were expressed as a percentage of all reactions involving H2O2 in each group of analyzed organisms (Fig. 2d).
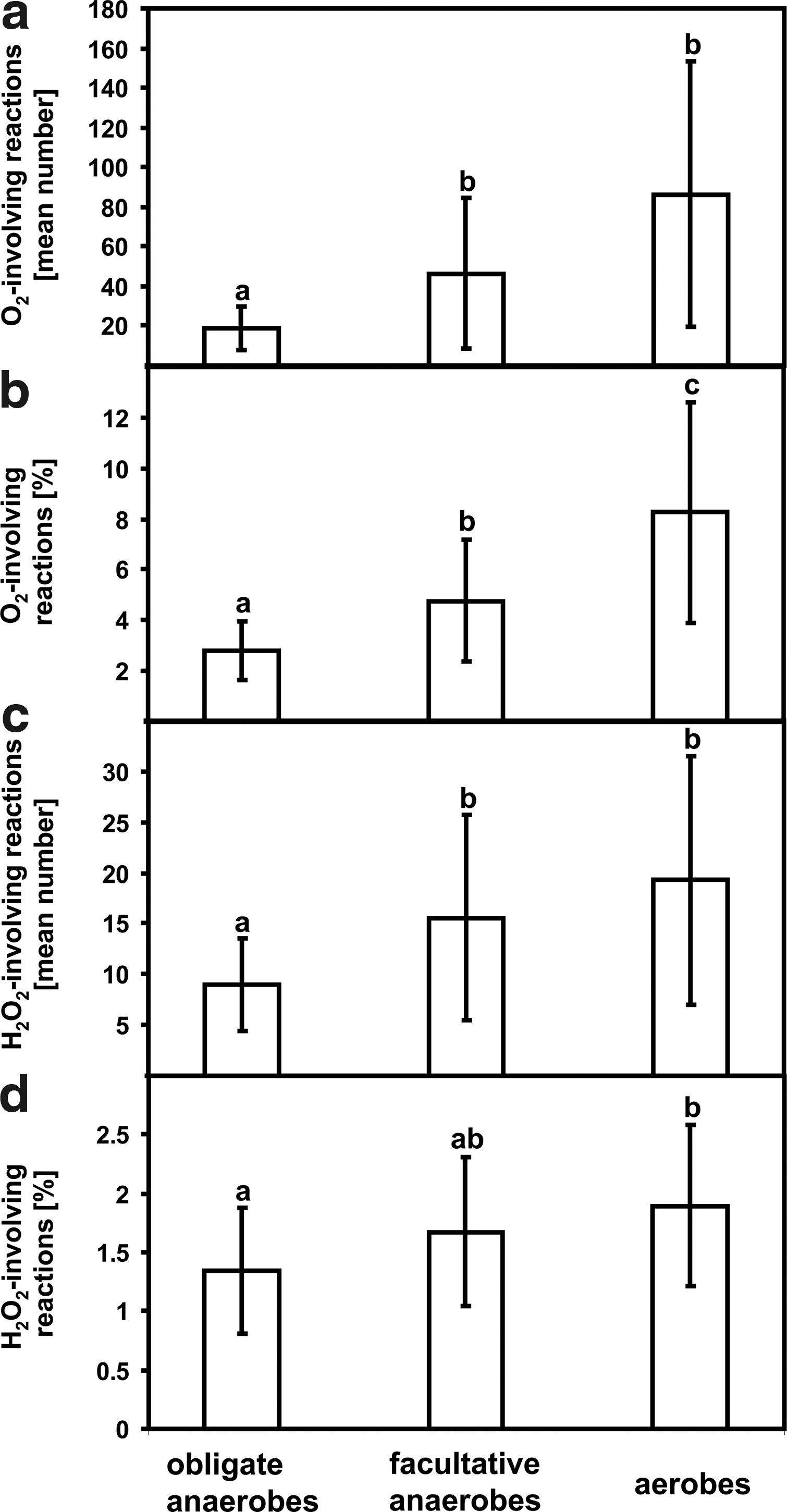
Reactions involving O2/H2O2. (
3.2. Reactions involving O2 and H2O2 in Bacteria, Archaea, and Eukarya
Calculations analogical to those presented in Fig. 2, but independent for Bacteria, Archaea, and Eukarya, were carried out (Fig. 3). The results show that obligate anaerobic bacteria have a significantly lower number and percentage of O2-involving reactions compared to those of aerobic bacteria and eukaryotes (Fig. 3a, 3b). Anaerobic archaea have less O2-dependent reactions than facultative anaerobic bacteria, aerobic bacteria, and aerobic eukaryotes. The analysis showed no significant differences in the mean number and percentage of O2-involving reactions between all groups of Archaea (Fig. 3a, 3b). For H2O2, the common tendency toward an increase of reactions involving H2O2 was similar to that for O2 (Fig. 3c). Moreover, the mean number of H2O2-involving reactions was significantly higher only between obligate anaerobic Bacteria, Archaea, and eukaryotic aerobes (Fig. 3c). If the results are presented as a percentage of reactions involving H2O2, significant differences were found between anaerobic Archaea and both facultatively anaerobic and aerobic Bacteria (Fig. 3d). It should be noted that the lowest mean number of O2/H2O2-involving reactions was very similar for strictly anaerobic, facultatively anaerobic, and aerobic archaea (Fig. 3d).
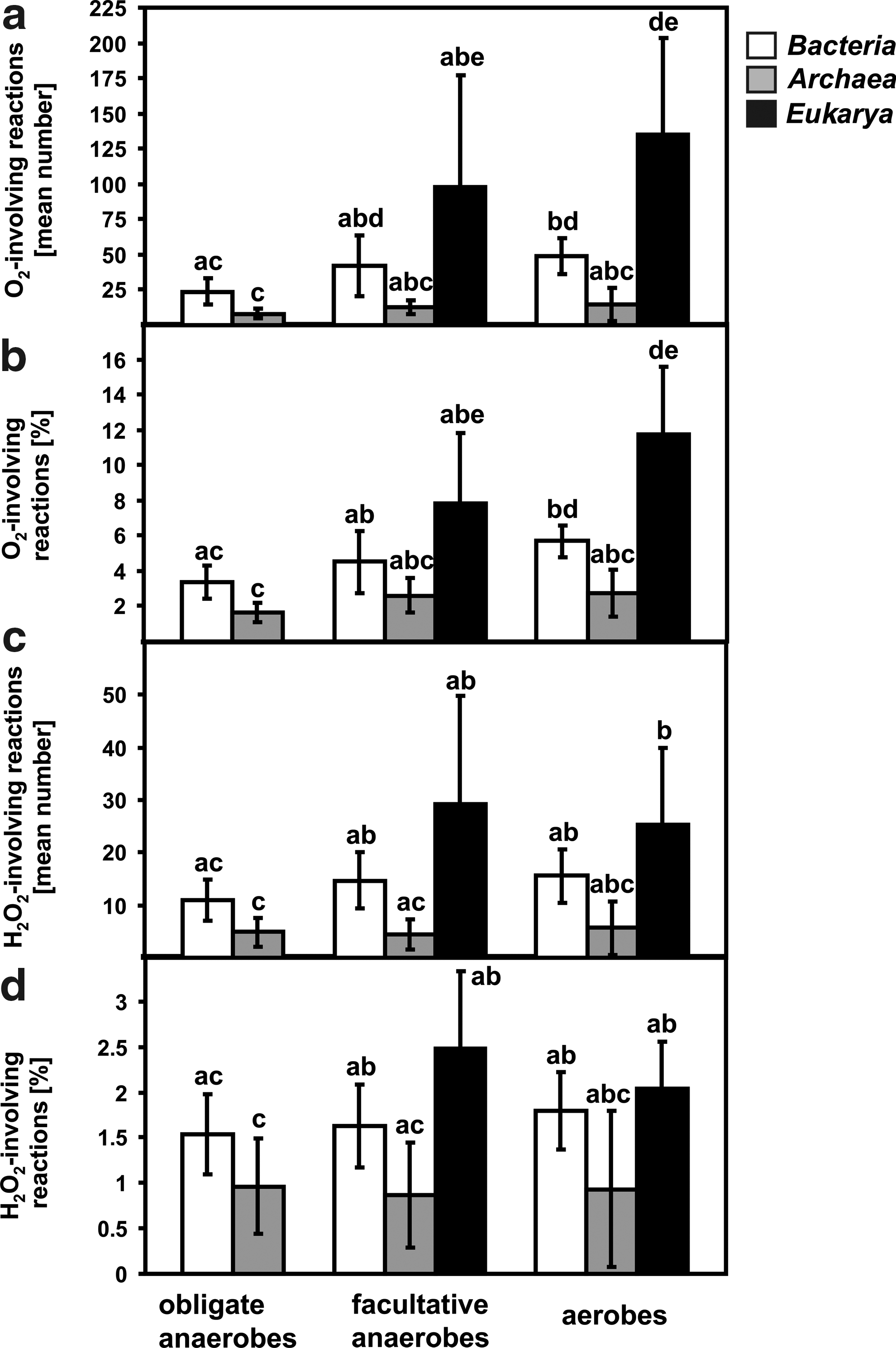
Reactions involving O2/H2O2 in organisms belonging to the three domains of life, Bacteria, Archaea, and Eukarya. (
3.3. The relationship between the mean number of reactions involving O2 and H2O2 and oxygenic photosynthesis
The data show a significant increase in the number of O2-involving reactions only between strict anaerobic bacteria and plants (Fig. 4a, 4b). Unexpectedly, oxygen-producing cyanobacteria showed a significantly lower percentage of reactions involving O2 than plants (Fig. 4b). Moreover, no significant distinctions in the quantity of O2-involving reactions between photosynthesizing obligate anaerobes, facultative anaerobes, and cyanobacteria were noted (Fig. 4a, 4b). For reactions involving H2O2, no significant differences between organisms were observed (Fig. 4c, 4d).
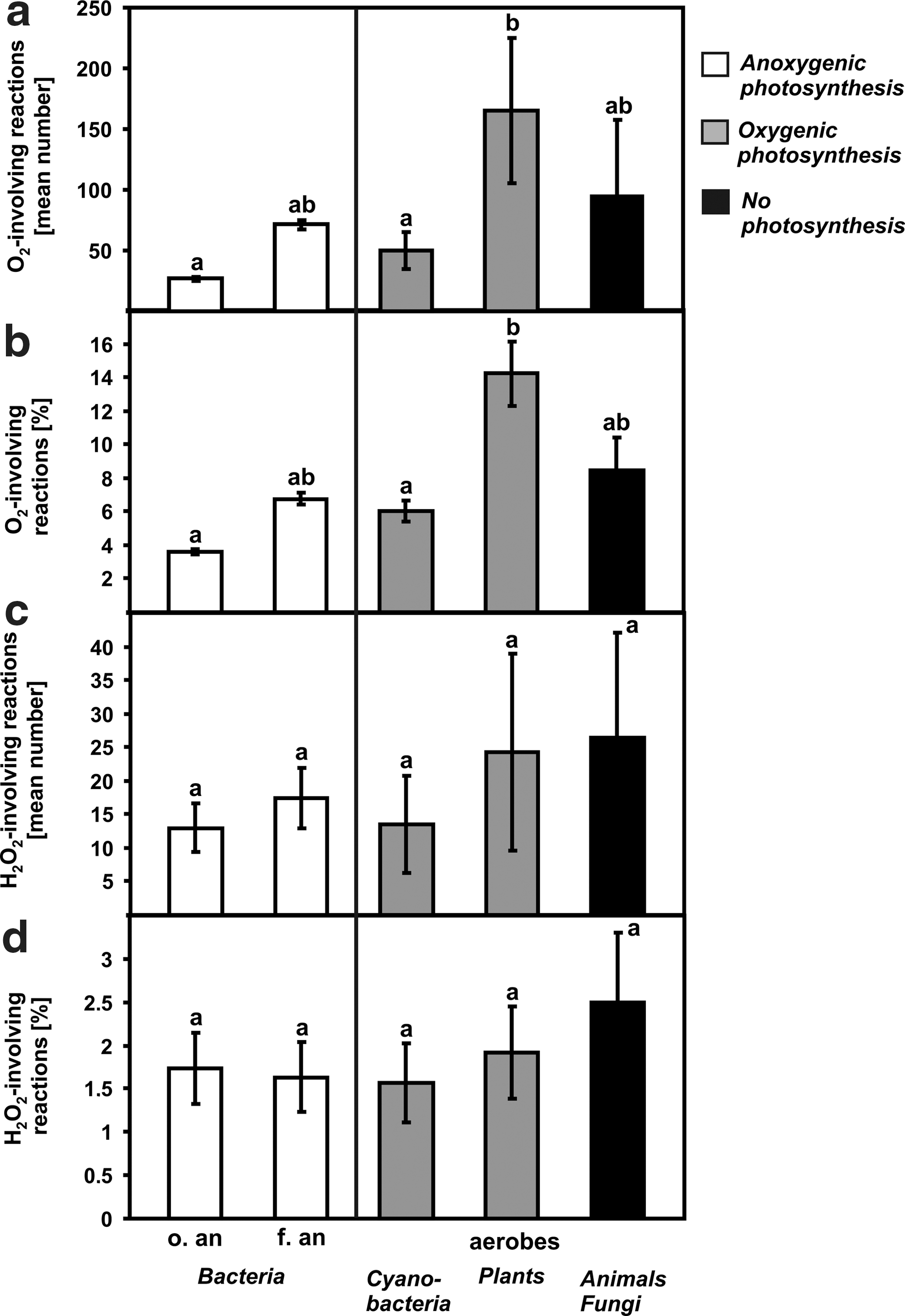
Reactions involving O2/H2O2 in organisms performing anoxygenic photosynthesis (Bacteria), oxygenic photosynthesis (cyanobacteria and plants), and in nonphotosynthesizing organisms (animals and fungi). (
3.4. Aerobic respiration and ROS removal
The percentage of ROS removal systems and aerobic respiration pathways found for aerobes was set to 100% (Fig. 5). Among obligate anaerobes and facultative anaerobes, 50% and 90%, respectively, possess both oxic pathways: aerobic respiration and ROS-removing reactions (Fig. 5c). If the pathways are analyzed separately, aerobic respiration and ROS removal pathways are found in ca. 55%/85% and 90%/93% of obligate anaerobes and facultative anaerobes, respectively (Fig. 5a, 5b).
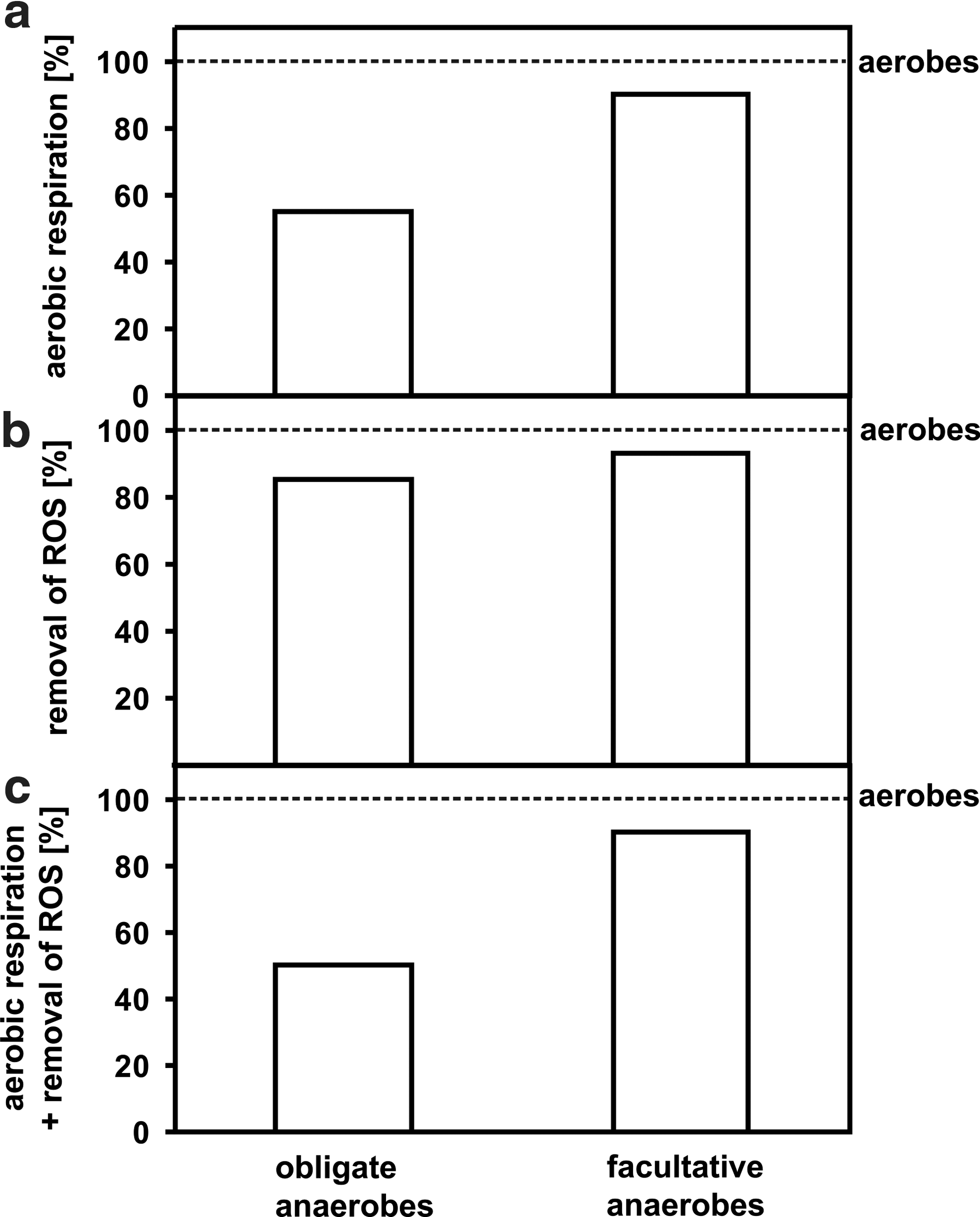
The presence of reactions/pathways representative of aerobes in obligate anaerobes and facultative anaerobes. (
4. Discussion
4.1. Reactions involving O2
In general, as expected, the mean number of O2-involving reactions and the percentage of reactions where O2 is engaged increased from strict anaerobes to obligate aerobes (Fig. 2a, 2b). Moreover, the results concerning O2/H2O2-involving reactions in Bacteria, Archaea, and Eukarya obtained during the analysis might indicate that obligate anaerobes belonging to Archaea are “more obligate” anaerobes than anaerobic bacteria. In this case, the mean number of O2/H2O2-involving reactions of anaerobic archaea is ca. 3- and 2-fold lower for reactions involving O2 and H2O2, respectively, compared to anaerobic bacteria (Fig. 3a, 3c). These results are surprising, because all the analyzed obligate anaerobes actually have metabolic pathways/reactions involving O2. If contemporary anaerobic pathways of obligate anaerobes are represented in their anaerobic ancestors, the occurrence of various O2-involving reactions/pathways in present obligate anaerobes might suggest that O2, as a metabolic reactant, appeared very early in the evolution of life. The results support the previously unorthodox hypothesis that photochemically produced molecular oxygen and aerobic metabolism were already present on early Earth and in primordial organisms, respectively, yet before the evolution of oxygen-producing cyanobacteria (Towe 1978, 1981, 1990; Castresana et al., 1994; Pereira et al., 2001). Recent studies have shown that dioxygen reductases, such as cytochrome oxidases type A (e.g., the contemporary mitochondrial form) preceded other cytochrome oxidases. In this context, the respiration-early hypothesis (Castresana and Saraste, 1995) that early microbiota contained low concentrations of O2 could not be ruled out (Brochier-Armanet et al., 2009). However, many reactions involving O2 may be replaced by analogous O2-independent (anaerobic) reactions, as it has been previously described (Raymond and Blankenship, 2004). Nevertheless, it is most likely that the majority of O2-dependent reactions, including O2 reduction by dioxygen reductases and ROS-detoxifying pathways (see below), cannot be simply substituted by anaerobic equivalents. The analysis of obligate and facultative anaerobes revealed the occurrence of aerobic respiration components in many obligate anaerobes (Fig. 5). It might be suggested that O2 reduction (Eq. 3) is not exclusively limited to strict aerobes only. Moreover, dioxygen may not be the main oxic compound that leads to the evolution of O2-dependent reactions. It cannot be excluded that one class of dioxygen reductases originally reduced other compounds, for example, nitric oxide (for a review see Ducluzeau et al., 2009). Recently, it has been found that facultative anaerobe E. coli is able to perform aerobic respiration at extremely low (≤3 nM) concentration of O2. If the primordial Earth atmosphere contained ca. 10−5 of the present atmospheric level of O2 (see Introduction), such a concentration of oxygen could have supported aerobic respiration, even before photosynthetically induced vast amounts of O2 arose in the atmosphere ca. 2.4 Ga (Stolper et al., 2010). During recently performed metagenomic studies, a new intracellular aerobic pathway in apparently anaerobic denitrifying bacterium Candidatus Methylomirabilis oxyfera was identified (Ettwig et al., 2010). This result reveals that O2 could have been available to the first microorganisms not only from abiotic sources but also from internal biochemical pathways even before oxygenic photosynthesis evolved (Ettwig et al., 2010).
The number of reactions involving O2 in organisms that perform anoxygenic photosynthesis (strict and facultative anaerobic bacteria) and oxygenic photosynthesis (cyanobacteria, plants), and in eukaryotic organisms that use O2 for respiration (animals/fungi), could indicate that evolution of cyanobacteria, as compared to strict and facultative anaerobes, was very slow and that the core of O2-involving reactions/pathways remained similar in photosynthesizing anaerobes and facultative anaerobes. Even heterotrophic aerobes, as “consumers” of O2, showed an insignificant quantity of reactions involving O2 in comparison to other photosynthetic organisms (Fig. 4). In contrast, plants that perform the same oxygenic photosynthesis are considerably more “overloaded” by O2-involving reactions/pathways than cyanobacteria (Fig. 4a, 4b). The differences between cyanobacteria and plants, mentioned above, require further study (see discussion below).
4.2. Reactions involving H2O2
Although it is not possible to separate O2-utilizing and ROS-removing reactions completely, in that some of these reactions overlap with each other (e.g., see Eqs. 4 and 6), we observed differences between the amount of O2-and H2O2-involving reactions for various groups of organisms. In general, the data concerning H2O2-utilizing reactions are fairly uniform (Fig. 2c, 2d; Fig. 3c, 3d) when compared to that of O2-utilizing reactions (Fig. 2a, 2b; Fig. 3a, 3b). This phenomenon was most pronounced when anaerobes that perform anoxygenic photosynthesis were compared to oxygen-producing cyanobacteria and plants (Fig. 4c, 4d; see also discussion below). There were no differences in the amount of H2O2-utilizing reactions between cyanobacteria/plants and the other organisms analyzed (Fig. 4c, 4d), which strongly indicates the early evolution of reactions utilizing H2O2, as has been suggested previously (McKay and Hartman, 1991). These results might also suggest that cyanobacteria evolved at the same time and under the same conditions as obligate anaerobes. In light of recent geochemical studies that focused on hematite minerals, cyanobacteria might have even appeared earlier than anaerobes, at least in some isolated areas (Hoashi et al., 2009). Recently, the complete genome of the anaerobic and thermophilic bacterium Candidatus Desulforudis audaxviator was sequenced. Although this organism lives in anoxic and hot conditions (48–62°C) at a depth of ca. 2.8 km below Earth's surface, it shows the presence of a sequence for Mn/FeSOD (Chivian et al., 2008). In line with the presented model, the generation of a superoxide anion radical, a substrate for SOD, might be induced by the radioactive decay of uranium present in the natural environment of this organism (Chivian et al., 2008). The production of H2O2 at the rock-water interface has also been reported, which suggests that ROS must have been generated long before the rise of free O2 in the atmosphere (Balk et al., 2009). The occurrence of ROS removal systems in so many strict anaerobes (Fig. 5) suggests that the production of ROS was unavoidable even under the globally anoxic environment of early Earth.
It cannot be excluded from these results that ROS, such as H2O2, a hydroxyl radical, and a superoxide anion radical, could also have affected the evolution of oxic pathways. The suggestion of ROS control before the development of oxygenic photosynthesis and before the photosynthetic rise of atmospheric O2 is strictly in line with theoretical data showing that photochemistry could produce ROS, mainly H2O2 on primordial Earth (Towe, 1978; McKay and Hartman, 1991; Liang et al., 2006; Haqq-Misra et al., 2011). Moreover, the analyses performed here strongly suggest that, even if the primordial organism (LUCA) was not able to use O2 as the final electron acceptor (aerobic respiration) or if O2 utilization was limited, it tolerated a local increase in O2 concentration as a result of ROS detoxification, for example, due to CAT activity (Eq. 6).
4.3. The alternative hypothesis and horizontal gene transfer (HGT)
The commonly accepted explanation of processes involving oxygen and ROS removal in strict anaerobes is that they have acquired O2/ROS-utilizing enzymes as a specific adaptation to an environment where oxygen sporadically occurred. It cannot be excluded that genes for O2/ROS metabolism and antioxidant enzymes were acquired by obligate anaerobes from aerobes later during the evolution of life as a kind of preadaptation to possible future transient exposure to oxygen or ROS, or both (Imlay, 2002; Brioukhanov and Netrusov, 2004). In this context, our conclusion concerning the presence of antioxidant enzymes in LUCA might be of less importance if horizontal gene transfer (HGT) had occurred many times during evolution between three domains of life as has been suggested for many other genes (Gribaldo and Brochier, 2009). Interdomain HGT events have been proposed recently for some classes of cytochrome oxidases (Brochier-Armanet et al., 2009). On the other hand, HGT strongly depends on the environmental conditions, such as temperature, pH, salinity, and so on, where organisms live. For example, prokaryotes preferentially acquire genes from other prokaryotes when both share the same aerobic/anaerobic environments (Jain et al., 2003). As evidence for this hypothesis, Passardi et al. (2007) indeed found that HGT for “antioxidant genes” encoding catalase-peroxidases occurred between prokaryotes showing similar O2 tolerance. It should be noted that even if HGT is an important evolutionary phenomenon, it does not affect all genes to the same extent (Gribaldo and Brochier, 2009). In our opinion, such a collective and synchronized transfer of the genes of enzymes of aerobic respiration and antioxidant enzymes (e.g., SOD, CAT) between aerobes and strict anaerobes was considerably less probable than, for example, between aerobes only. Among antioxidant enzymes, superoxide reductase (SOR, Eq. 5) has been identified predominantly in those strict anaerobes and microaerophiles that belong to Bacteria or Archaea (Imlay, 2002; Lucchetti-Miganeh et al., 2011). The very rare occurrence of SORs in aerobic organisms indicates generally that anaerobes could not have acquired these enzymes by recent HGT from aerobes, because they occupied different habitats. Moreover, it suggests that these enzymes could appear long before the rise in O2 in Earth's atmosphere. Based on this, we surmise that the HGT of many “oxic genes” between aerobic and anaerobic Archaea and Bacteria was not essentially responsible for the relatively wide spread of different O2-utilizing reactions and ROS removal pathways in contemporary anaerobes.
One possible explanation of such surprising results is the fact that the analysis did not take into consideration all the strict anaerobes, facultative anaerobes, and aerobes whose genomes have been sequenced and annotated to date. Moreover, not all metabolic pathways are empirically verified, nor are all databases currently updated. Nevertheless, we suggest that an extrapolation made on the basis of ∼100 genomes reveals real differences between these groups of organisms, and it offers a new path for studies concerning the earliest events during the evolution of living beings and for speculation on the attributes of LUCA.
4.4. Conclusions and implications for further research
The presented data suggest that the hypothetical LUCA was “equipped” with reactions involving O2/H2O2. Based on this, the presence of low concentrations of ROS, such as H2O2, and O2 should be taken into account in theoretical models of the early evolution of Archean oceans/atmosphere and life. However, no possible sources of ROS were identified. Nevertheless, the obtained results might indicate that the ability of ROS detoxification did not emerge as a response to an increase in O2 level produced by cyanobacteria but that it was a crucial adaptation to weakly oxic (ROS) primordial microenvironments as a result of abiotic photochemical processes on early Earth. This adaptation could have allowed for the very early appearance of oxygenic photosynthesis. Thus, the hypothesis concerning the occurrence of ROS-utilizing genes/enzymes and at least O2 tolerance by the first living organisms cannot be excluded and requires further study.
Footnotes
Acknowledgments
I.Ś. is grateful for partial support by the Welcome 2008/1 project operated within the Foundation for Polish Science Welcome Program co-financed by the European Regional Development Fund, COST project: 595/N-COST/2009/0, and the Ministry of Science and Higher Education (Poland) grant no. N 303 3569 35. We also thank all reviewers for several suggestions that helped to strengthen this paper. The authors would like to thank Prof. Chris McKay and Prof. Hiroshi Ohmoto for their valuable comments and suggestions to improve the quality of the paper.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
CAT, catalase; HGT, horizontal gene transfer; LUCA, Last Universal Common Ancestor; ROS, reactive oxygen species; SOD, superoxide dismutase (Fe/MnSOD, iron/manganese form of SOD); SOR, superoxide reductase.