
Abstract
The Life Marker Chip (LMC) instrument is part of the proposed payload on the ESA ExoMars rover that is scheduled for launch in 2018. The LMC will use antibody-based assays to detect molecular signatures of life in samples obtained from the shallow subsurface of Mars. For the LMC antibodies, the ability to resist inactivation due to space particle radiation (both in transit and on the surface of Mars) will therefore be a prerequisite. The proton and neutron components of the mission radiation environment are those that are expected to have the dominant effect on the operation of the LMC. Modeling of the radiation environment for a mission to Mars led to the calculation of nominal mission fluences for proton and neutron radiation. Various combinations and multiples of these values were used to demonstrate the effects of radiation on antibody activity, primarily at the radiation levels envisaged for the ExoMars mission as well as at much higher levels. Five antibodies were freeze-dried in a variety of protective molecular matrices and were exposed to various radiation conditions generated at a cyclotron facility. After exposure, the antibodies' ability to bind to their respective antigens was assessed and found to be unaffected by ExoMars mission level radiation doses. These experiments indicated that the expected radiation environment of a Mars mission does not pose a significant risk to antibodies packaged in the form anticipated for the LMC instrument. Key Words: Life-detection instruments—Planetary habitability and biosignatures—Radiation—Mars—Life in extreme environments. Astrobiology 12, 718–729.
1. Introduction
For use in a planetary exploration context, there is concern that antibodies and other immunoassay reagents will not be sufficiently robust to survive the various environments and stresses encountered without extensive protection. Limited experiments have been performed to demonstrate that some conditions encountered in space missions do not pose a challenge; one of the first examples is work by Maule et al. (2003) with experiments on antibody performance in conditions of altered gravity. Among the various factors that could have a detrimental effect on immunoassay performance, the radiation environment is generally considered to be one of the most threatening aspects of a space mission, and any biological materials must be tested for their ability to survive exposure to various types of radiation in an interplanetary mission context. Irradiation of affinity receptors with particle radiation has not been extensively studied, with the exception of Le Postollec et al. (2009) with irradiation of polyclonal IgG with neutron radiation, Baqué et al. (2011) with proton irradiation of nucleic acid aptamers, and more recently de Diego-Castilla et al. (2011) with the study of the effects of gamma radiation and temperature shifts on antibody performance. Both the microgravity study (Maule et al., 2003) and the irradiation experiments demonstrated that affinity receptors (antibodies and aptamers) were able to resist inactivation under these types of stress, at the levels and doses studied in each case. Despite the positive results from these experiments, there is a need for more studies that involve exposure of affinity receptors to space radiation with the intent to gain a better understanding of the effects it has on their survivability and overall performance.
The driving force behind the radiation studies described herein is the ongoing development of the Life Marker Chip (LMC) instrument, an antibody-based instrument that is part of the Pasteur instrument payload of ESA's ExoMars rover mission (Vago et al., 2006). The rover mission's primary goals are to search for evidence of past or present life on Mars and to cache samples for Earth return at a later date; the rover is currently scheduled to launch in 2018.
The LMC is frequently described in the general media as a “pregnancy test for Mars,” given that both the LMC and commercial off-the-shelf pregnancy tests are based on lateral flow immunodiagnostic technology (Wilson and Howell, 2002). Thus, the LMC uses (i) multiple dried-down fluorescently labeled antibodies that (ii) dissolve into an aqueous-based liquid extract of a martian regolith or crushed rock sample containing extracted organic molecules and (iii) initiates a multiplexed inhibition immunoassay before (iv) being flowed over a microarray composed of immobilized copies of the various multiplexed immunoassay targets. For a given antibody, (v) if the corresponding target molecule is present in the liquid extract, the antibody binding site is occupied and thus inhibited from binding to the corresponding microarray spot, which (vi) subsequently does not fluoresce. Conversely, if the relevant target molecule is not present in the liquid extract, the antibody binding site is left vacant and therefore able to bind to the corresponding microarray spot that fluoresces. The immunoassay target molecules chosen as LMC targets are those representative of ancient, preserved biomarkers of extinct life, biomarkers of extant life, and those that represent meteoritic input and other abiotic organic molecules as well as organic molecular markers of rover- and instrument-borne Earth contamination (Parnell et al., 2007).
One of the main concerns for the reagents at the core of the LMC is the exposure to space radiation, and two classes of molecular materials are of interest within the context of their stability to the cumulative radiation environment of a Mars mission. One class, fluorescent dyes, has been studied elsewhere (Thompson et al., 2006). The second class, antibodies, is the focus of both the ground-based radiation simulation described herein and an experiment flown on board Biopan-6, the experimental platform for the exposure of biological material to conditions, including radiation, in low-Earth orbit (Derveni et al., unpublished data).
The key radiation elements to be considered for the LMC as part of the ExoMars mission are the radiation en route to Mars (cruise phase) and the radiation environment on the planet's surface (surface phase). Other environments, such as those in low-Earth orbit, and transition through Earth's radiation belt are insignificant due to the short duration of exposure for the proposed mission trajectory. The radiation environment to which all mission instruments are exposed while en route to Mars is mainly due to galactic cosmic rays (GCR) and solar particle events (SPE). GCR exposure is expected to be approximately constant, while radiation associated to SPE will be more sporadic (Hands and Rodgers, 2006—see Supplementary Material available online at
In terms of the radiation profile on the surface of Mars [modeled by Dartnell et al. (2007)], a number of different types of radiation are present that can have an effect on surface exploration missions like ExoMars. The component of the surface radiation environment that is most relevant to simulate for the current study is the neutron environment. The surface neutrons are produced via interactions between cosmic rays and the upper layer of regolith, scattering secondary neutrons back toward space. SPE protons play a much less significant role on the surface than they do during the cruise phase due to significant attenuation from the thin martian atmosphere (Tripathi and Nealy, 2008). Other types of radiation (UV, mineral radiation, GCR) are also present; some of them are heavily attenuated by the thin martian atmosphere and others by the rover's outer shielding, which makes them secondary to the effect of neutron radiation.
The objective, therefore, of this study has been to demonstrate that suitably packaged antibodies can survive exposure to the expected radiation environment of a Mars mission. This will be achieved by taking antibodies representative of those likely to be in the final version of the LMC and packaging them in a format similar to that intended for the final LMC flight instrument before exposing them to various levels and combinations of proton and neutron radiation (including levels representative of those expected for the ExoMars mission) produced by a cyclotron. Post-exposure, the effect of the radiation on the antibodies' binding ability will be assessed.
2. Materials and Methodology
2.1. Estimation of radiation exposure during a Mars mission
As described previously, the dominant radiation threat to the LMC molecular reagents during a mission to Mars will be high-energy protons during the mission cruise phase and neutron radiation during surface operations. By using standard modeling tools (SPENVIS and CREME96)1, the proton fluence for a 2-year cruise phase was calculated with a 95% upper confidence limit at 2×1010 protons/cm2. Given that the preceding value represents a spectrum of energies that is unrealistic to simulate at a cyclotron source, normalization to a mono-energetic value was required and calculated to be equivalent in terms of total ionizing dose to a 50 MeV mono-energetic fluence2 of 4×1010 protons/cm2 (hereafter “mission dose”) (Hands and Rodgers, 2006—see Supplementary Material). This figure was doubled to provide a suitable minimum “overtest” fluence of 8×1010 protons/cm2, which corresponds to a total ionizing dose of approximately 14 Krad (140 Gy) (Hands, 2008b—see Supplementary Material).
Modeling of the radiation environment at the surface of Mars was performed with the use of a Monte Carlo simulation as part of ESA's MarsREM project (Gonçalves et al., 2009). For a 6-month-long operational period on the martian surface, the neutron dose was calculated, after normalization, at ∼20 millisieverts (mSv). At the test facility used, this value was equivalent to a fluence of 1.8×107 neutrons/cm2 at an energy of 47 MeV, rounded to 2×107 neutrons/cm2 (hereafter “mission dose”) and doubled to give an experimental minimum “overtest” fluence of 4×107 neutrons/cm2 (Hands, 2008a—see Supplementary Material).
2.2. Choice of radiation exposure levels
The simulated Mars mission radiation studies took place in two stages, Phase I (February 2008) and Phase II (September 2008). The reasons for having two rounds of irradiations included the staged availability of antibodies against biomarkers of interest (availability of sample antibodies) and the development of additional lyoprotection and radioprotection methods for the antibodies.
In Phase I, the irradiations focused on investigation of the effects of combined proton and neutron irradiation, both in doses that simulated the “mission dose” for ExoMars and exceeded the mission dose 20-fold, as an overtest by which to test the antibodies' survival limits. Some samples also received the “mission dose” of protons or neutrons in isolation to clarify whether either type of radiation would dominate the damage caused to the samples.
Phase II was designed to provide further information about irradiation damage to a number of antibodies that were not available during Phase I and confirm the findings of the previous round for all antibodies used in Phase I. A further test during Phase II was the irradiation of antibody samples at 250 times the mission dose for protons, an extreme dose to help identify the upper threshold of exposure for antibody inactivation. Neutron irradiation at 250 times the ExoMars dose was not performed at this stage due to limited beam time and, as the effect of neutron radiation had been deemed negligible, after analysis of the results from Phase I.
2.3. Selection of antibodies for study
Antibody selection was based on two main criteria; they had to be representative of the broad range of types of antibodies that could be included in the final format of the LMC, and they had to be part of existing working assays within the lead authors' group. Availability was another factor that affected the choice of antibodies for each round of experiments. Table 1 provides a list of the antibodies used in the present study against a broad range of antigens—whole cell, protein, and small molecule—as well as IgG antibody types/formats—polyclonal, monoclonal, and recombinant fragments.
The anti-GroEL antibody was purchased from Sigma-Aldrich (G6532), and the single-chain antibody fragment against atrazine was provided by ImmunoSolv Ltd. The mouse monoclonal antibody to peptidoglycan from Staphylococcus aureus was purchased from Abcam (ab20002), as were the rabbit polyclonal antibody against phytanic acid (ab51309) and the rabbit-raised polyclonal anti-FITC antibody (ab19491-100).
2.4. Protective matrices
To increase the resistance of the antibodies to inactivation prior, during, or after exposure to radiation, three protective matrices (solute mixes) were used in the freeze-drying process, in addition to samples that were freeze-dried in the buffer solutions in which they were provided by their manufacturers. These matrices were mixtures of sugars, proteins, and free radical scavengers, and are well established for use in the pharmaceutical/diagnostics industry, which allowed us to assess the effectiveness of these additives on the stability and post-rehydration activity of the antibody samples. The decision to include sugar/protein mixtures as protective agents within an instrument employed to detect, among others, whole molecules and/or oligomers of sugars and proteins may appear counter intuitive. Initial testing has shown, however, that correct integration and thorough cross-reactivity testing can ensure the minimization of any contamination issues to below the detection limits of the assays. Two matrices were used in Phase I of the present work; one of the mixtures included the dye Blue Dextran (“Blue Dextran–based matrix”), and the second mixture included sucrose (“Sucrose-based matrix”). A third matrix was added during Phase II, a mixture that included the dye Allura Red (“Allura Red–based matrix”). The Allura Red matrix was included due to codevelopment of the LMC instrument, where Blue Dextran was found to interfere optically with the fluorescence-based immunoassay format used and the high-molecular-weight nature of Blue Dextran affected the viscosity of the resultant assay solution. Hence, Allura Red was used as a low-molecular-weight visual tracer dye for microfluidic development reasons and did not interfere with the fluorescence-based immunoassay format used.
The disaccharide (sucrose) and polymer (Blue Dextran) used in these mixtures were added as lyoprotectants, and bovine serum albumin (BSA) served as an anti-aggregation agent. Tween 80 is considered protective during freezing and thawing and has been suggested to prevent protein adsorption to ice/water interfaces during freezing and assist protein refolding during thawing. Also, ascorbate is believed to be an effective radical scavenger (Niki, 1991). A list of the reagents used in each matrix can be found in Table 2.
The list of components for the lyophilization matrices included BSA, which was used in all three lyophilization matrices and was purchased from Sigma-Aldrich (A4503). The HEPES buffer was also from Sigma-Aldrich (H7523), as was the sucrose (S7903), the Blue Dextran (D5751), Allura Red (458848), the ascorbate (Fluka 95210), and Tween 80 (W29, 170-6-K).
2.5. Sample integration
Independently of the current work, it has been decided to integrate antibodies into the LMC instrument as freeze-dried preparations (to increase reagent stability and radioresistance), that is, they were freeze-dried directly into laser-cut glass fiber pads (to ease antibody integration during LMC assembly). This format was maintained during the radiation studies as it allowed radiation exposure in a flight-model representative format.
In terms of the preparation of the pads, glass-fiber rectangles (2×4×0.6 mm3) were laser-cut with a Fenix CO2 Laser Marker (SYNRAD) from glass microfiber filters (Whatman GF/D, 47 mm ø). A volume of 6 μL of each antibody in buffer or lyophilization matrix was dispensed onto a pad and allowed to wick into it. The volume fully saturated the pad. Antibodies were used either directly as the fluid sample supplied or diluted 1:1 with lyophilization matrix (when used). After sample addition, pads were immediately frozen by immersion into liquid nitrogen. Due to the pads' small size and little thermal inertia, it was observed that removing them from liquid nitrogen prior to the freeze-drying step of the process resulted in rapid thawing. The problem was overcome by keeping the frozen pads in liquid nitrogen, inside 20 mL plastic beakers, which were then placed in the vacuum chamber of a bench-top freeze dryer (Coolsafe 55-4 bench-top freeze dryer from ScanVac). Upon establishment of the vacuum inside the chamber, the liquid nitrogen quickly boiled away. The samples were dried overnight (approx. 12 h). The lyophilized samples were then loaded into 384-well microtiter plates from Nunc (cat. No 242765) within a central square area of 32×32 mm that was compatible with the approximately 8–10 cm diameter of the particle beam (Fig. 1).

View of a 384-well microtiter plate with antibody sample pads loaded in the central square area of 32×32 mm (plate area is 12.5×8 cm).
Each antibody sample was freeze-dried into a separate glass fiber pad, and for each set of experimental conditions three replicates of each antibody pad were used to confirm the reproducibility of the results.
In addition to the microtiter plates with samples for exposure to radiation, a number of control antibody pads were prepared and packed identically as were the samples for exposure. One set of controls was kept at Cranfield University, in ambient temperature and atmosphere, for the duration of the study prior to final analysis (“storage” controls). A second set of control pads was shipped alongside the samples for exposure to the radiation facility, but they were not irradiated (“shipping” controls). Both sets of control pads were analyzed in parallel with the radiation-exposed samples to determine differences in antibody functionality caused solely by exposure to radiation, after subtraction of the effects of the storage and shipping process on the samples.
2.6. Exposure facilities
The two radiation test campaigns took place at the Theodor Svedberg Laboratory (TSL) in Uppsala, Sweden. The accelerator of the laboratory is the Gustaf Werner cyclotron (Prokofiev et al., 2007).
The proton beam is collimated, and the diameter is stable at 8 cm, with a top-hat radial distribution. The neutron beam, which originates from a point source (a lithium target), is best described as cone-shaped, with a diameter of approximately 10 cm at the samples' distance from the source. The samples' exposure to both radiation beams was considered uniform (in the case of the neutron beam, exposure is not uniform in theory, but given the samples' dimensions and distance from the source, this was considered a reasonable approximation). Table 3 shows a detailed description of the samples used in each phase and each level/combination of radiation exposure.
2.7. Antibody recovery and ELISA for assessment of antibody binding activity
After sample return from TSL, the antibodies in the pads were recovered and used in enzyme-linked immunosorbent assays (ELISA). To recover antibodies, each pad was placed inside the narrow tapered end of a shortened 10 μL plastic pipette tip and placed inside a 1.5 mL Eppendorf tube. Six microliters of phosphate-buffered saline solution was added to one end of the pad and wicked into the pad. This was followed by centrifugation, which separated the pad from the resulting solution and allowed the efficient collection of the solution with any rehydrated and washed-out antibody. Each pad was subjected to 10 washes, which yielded a ∼60 μL solution with an expected antibody dilution factor of 10, assuming 100% washout efficiency.
The subsequent standard ELISA applied in the analysis of samples followed the same protocol in all cases (see Supplementary Material), the only variable being the antibodies and antigens used in each assay. The ELISA were all binding assays, aiming to test the antibodies' ability to recognize and bind to their respective targets as represented by appropriate protein conjugates or native antigens—no competition or inhibition assays with free antigens were attempted.
3. Results
3.1. Radiation dose measurement
To represent the radiation dose that corresponds to each sample-loaded pad (2×4 mm2), “sample fluence” (which is the particle fluence at the mounted sample's distance from the beam exit point) was used. Table 4 shows the total dose per pad for the proton and neutron irradiations in Phase I and Phase II.
N/A=not available.
3.2. ELISA results
The ELISA results from both Phase I and Phase II were collated and plotted against “standard” curves that were obtained from ELISA with the use of fresh stock solutions of all antibodies. Due to availability limitations, only the anti-GroEL antibody was used in radiation plates 2 to 4. Figure 2 summarizes the results of the Phase I irradiations.
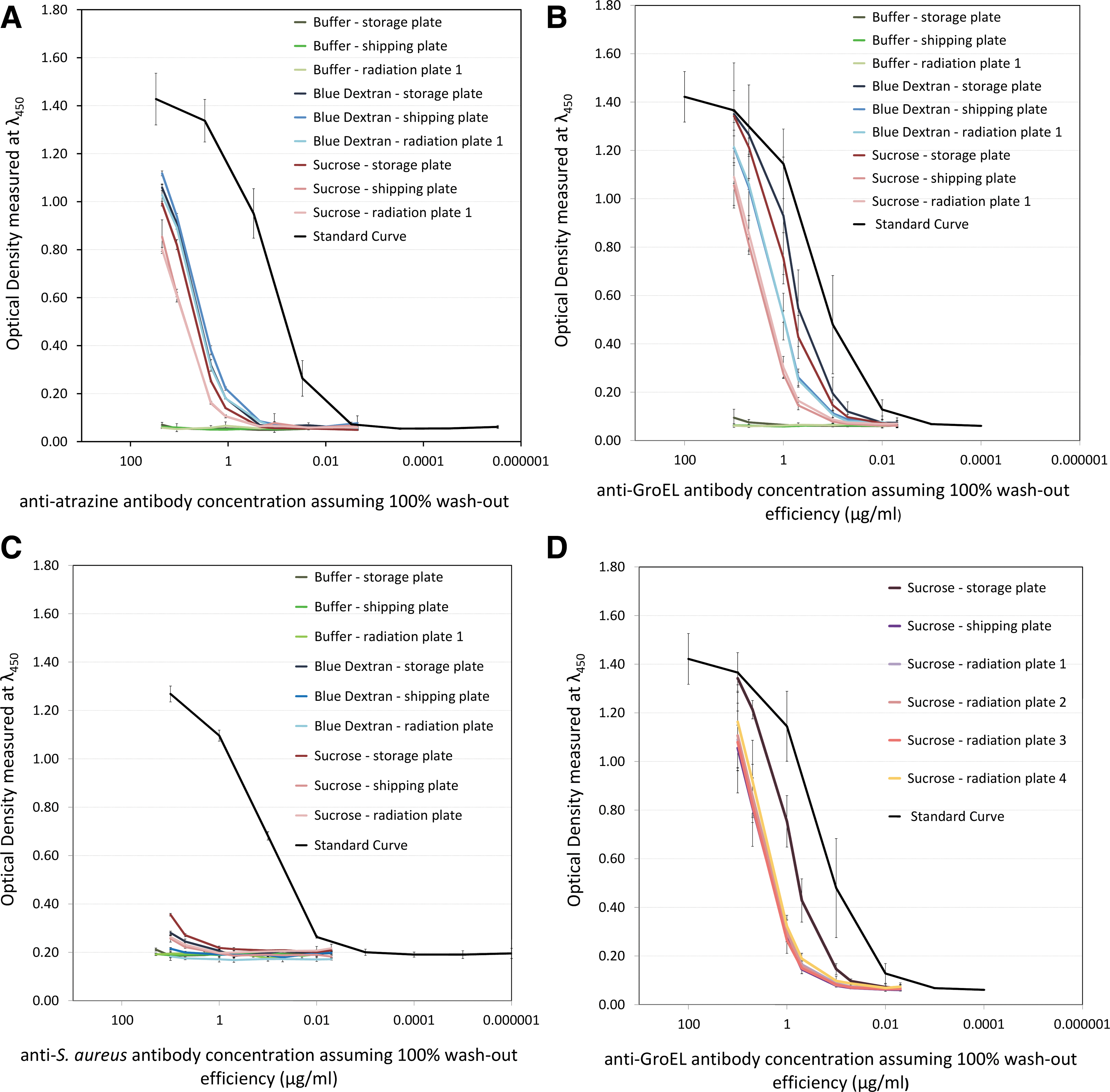
Binding performance of the anti-atrazine (
From Fig. 2, the performance of all the antibodies that have been freeze-dried appears significantly lower to that of nonlyophilized stocks (standard curves). In terms of specific performance, the monoclonal antibody to S. aureus (Fig. 2C) presented the poorest activity, regardless of the lyophilization matrix used. In the case of the anti-atrazine antibody, there appears to be no significant differences between the controls and radiation-exposed samples, while the anti-GroEL antibody presents a clearer distinction of binding activity between controls and irradiated samples.
Concerning the stand-alone effect of radiation, Fig. 2D shows the performance of the anti-GroEL antibody in the sucrose-based matrix, after exposure to various doses and combinations of proton and neutron radiation (for details, see Table 3), and shows that all radiation-exposed samples and the shipping control samples had similar levels of retained antibody activity, though the storage control had retained significantly more activity than the other samples. To quantify the differences in each antibody's binding activity due to the use of specific protective matrices and/or irradiation, the data is summarized in Table 5.
n/a=not available.
For Phase II, two antibodies, in addition to those used in Phase I, were added to the sample set: a polyclonal antibody against phytanic acid and a polyclonal antibody against fluorescein isothiocyanate (FITC). Both antibodies were available in limited amounts, and priority was given to samples lyophilized in the sucrose-based matrix, which was considered the most efficient at the time of studies. An oversight in the handling of the storage control samples rendered them unsuitable for further consideration3; for this reason they are not included in the results of Phase II. Given the difficulty in repeating the experiments to replace the storage controls, and judging by the results in Phase I where the storage and shipping controls gave similar results, the shipping controls were considered adequate as a control sample set. Figure 3 shows the ELISA results for the anti-GroEL (Fig. 3A) and anti-phytanic acid (Fig. 3B) antibodies.
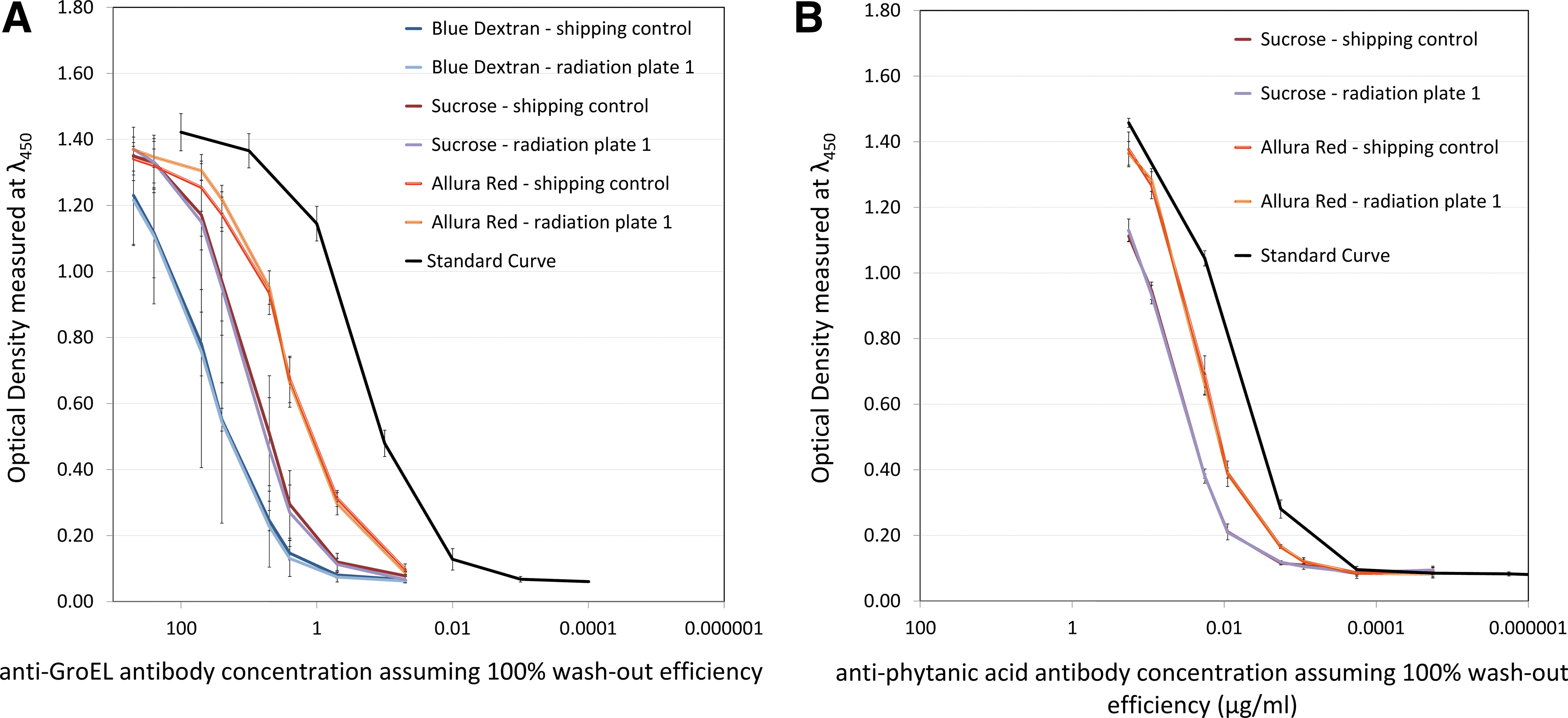
Binding performance of the anti-GroEL antibody (
Figure 3 shows how the use of the Allura Red–based matrix resulted in higher signals than those obtained from the use of the sucrose-based matrix, while the Blue Dextran–based matrix appeared to result in the lowest signals from the three matrices.
Figure 4 shows the ELISA results for the same two polyclonal antibodies, anti-GroEL (Fig. 4A) and anti-phytanic acid (Fig. 4B), but this time the varying factor is the dose and combination of radiation to which each sample was exposed.
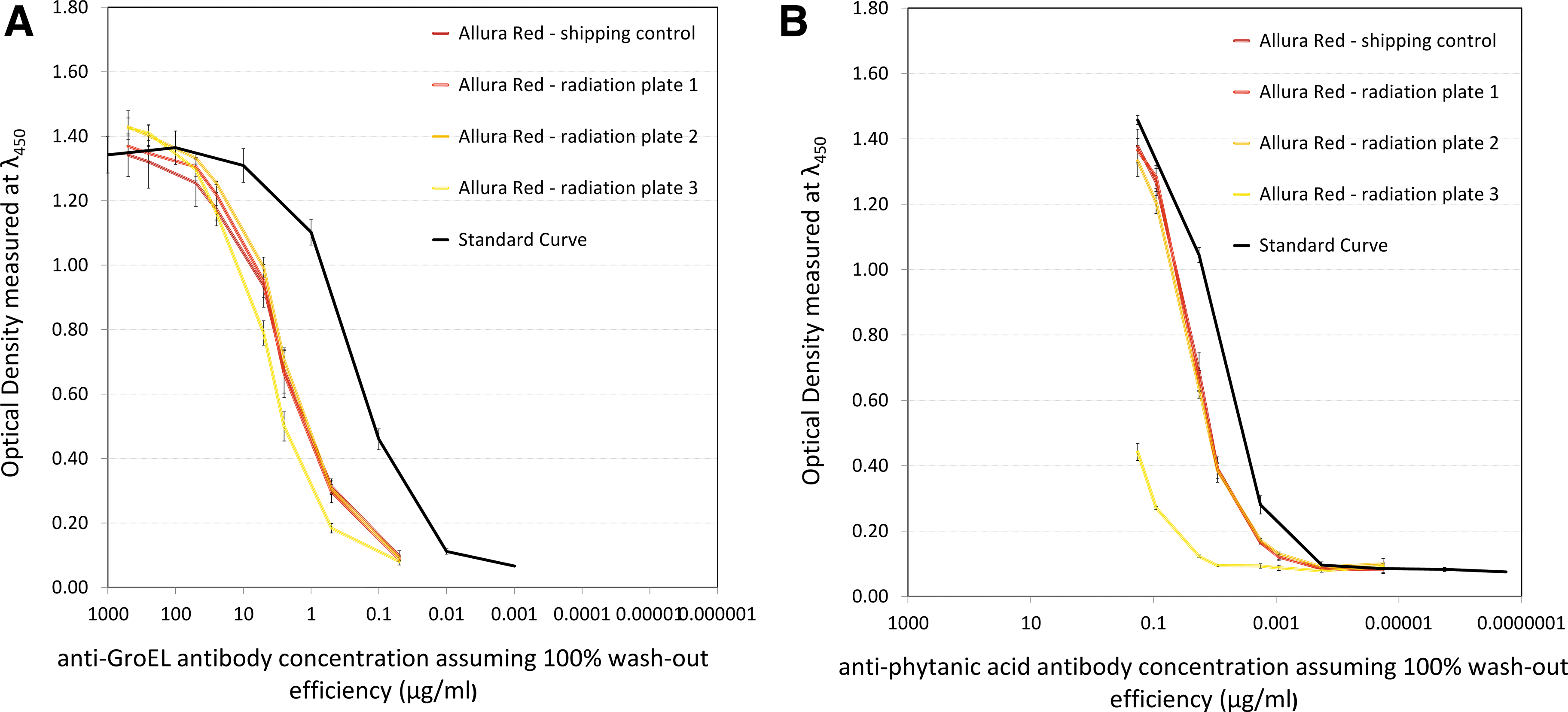
Binding performance of the anti-GroEL antibody (
The different reaction of antibodies to radiation exposure can be seen in Fig. 4, where the anti-GroEL antibody seems to maintain its binding activity even after exposure to 250 times the ExoMars envisaged proton radiation dose, while the anti-phytanic acid antibody shows visibly reduced activity after being exposed to the same dose of radiation.
Figure 5 shows the ELISA results obtained for the two remaining antibodies, the anti-S. aureus monoclonal (Fig. 5A) and the polyclonal anti-FITC (Fig. 5B). Both of them were only freeze-dried in the sucrose-based matrix, so the varying factor in Fig. 5 is the level of radiation to which each sample was exposed. From Fig. 5, it is clear that these two antibodies showed different levels of activity retention (compared with the standard curves which serve as an indication of the “maximum” antibody activity), with the anti-S. aureus antibody being heavily affected, similarly to Phase I. To quantify the loss of activity for all antibodies used in Phase II, the data is summarized in Table 6.
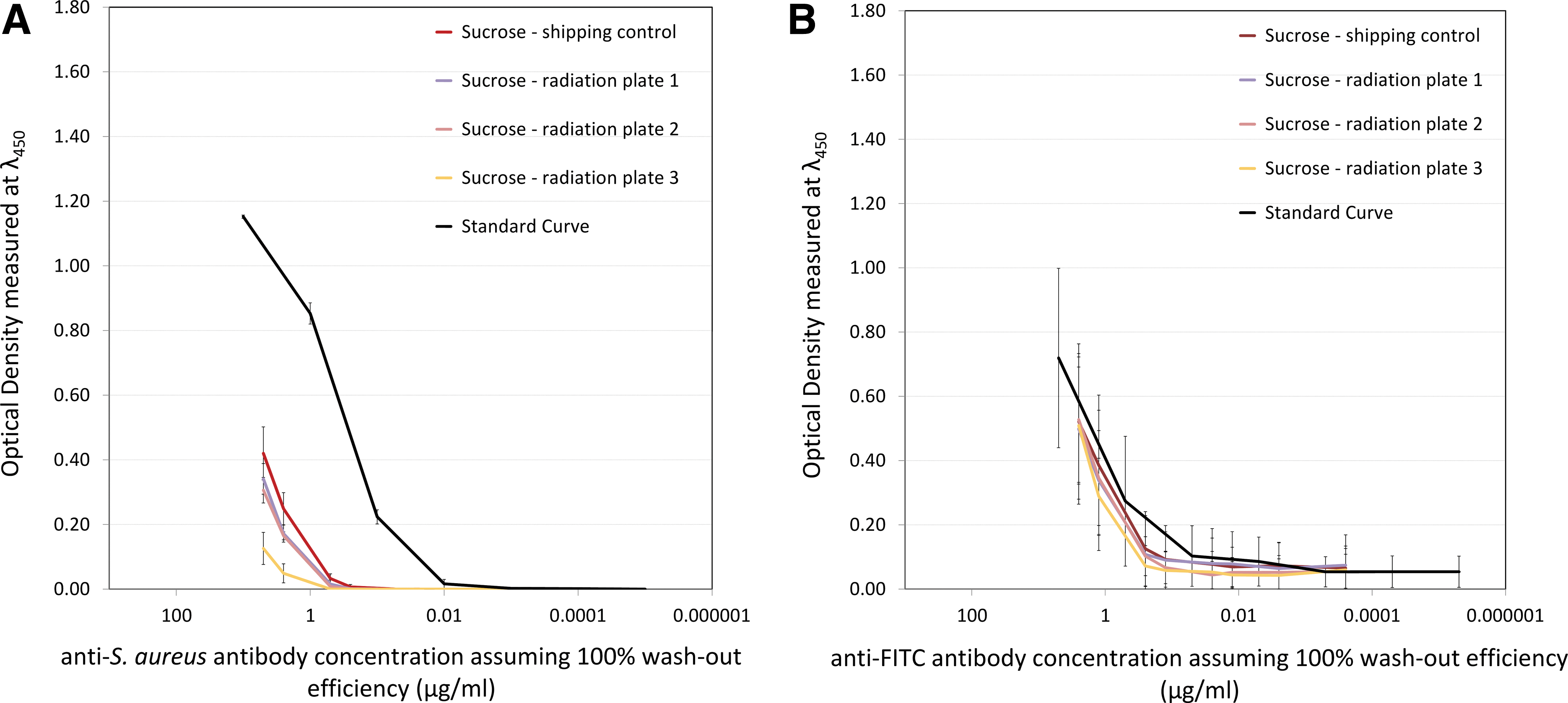
Binding performance of the anti-S. aureus (
n/a=not available.
4. Discussion
Ground-based simulations are subject to a number of limitations and approximations and therefore never fully represent all the events during a real mission. However, if appropriately designed, such experiments are very useful and can be the only way to study the effect of factors (in this case, radiation) that could have a harmful effect on the performance of an instrument. In the modeling and implementation of the radiation environment, we assumed that SPE-related proton radiation is the dominant threat en route to Mars, ignoring the radiation hazard by GCR ions and other forms of radiation (e.g., gamma). Similarly, the composition of the radiation environment on the surface of Mars was simplified to include only the secondary neutrons. Given the expectancy that these types of radiation will have, the dominant effects on the operation of the LMC, and the unrealistic nature of trying to simulate the heterogeneity of the real radiation environment, these simplifications were considered acceptable and a satisfactory representation of the envisaged mission conditions.
Mission duration is another factor to consider; at the time of modeling it was assumed that the Earth-Mars cruise phase would take place over a 2-year period, with an additional 6-month period of surface operation. This estimation was based on the early ExoMars mission design and is a significant overtest of the current (circa 2010) ExoMars mission design, which has a 9-month transit period.
For the dose estimation, the present experiments were designed with estimated radiation doses during a mission to Mars that were the result of modeling of the radiation environment based on “worst-case scenarios” for every type of radiation that forms part of this environment; for example, assume solar maximum and worst-week scenario for a solar proton event. Consequently, they do not necessarily offer a representation of the actual levels of radiation that will be encountered by the ExoMars mission. The advantage of this simulated environment is that it results in a conservative estimate of the dose (i.e., higher radiation dose than expected) that could be incident on the mission instruments; it is therefore another overtest of the expected conditions.
Concerning experimental dose deposition rates, for a real mission the total dose of radiation would be accumulated over the mission's duration period, while in the present experiments the samples were exposed to the same dose of radiation in a period ranging over some minutes. This experiment design assumed that radiation damage to the antibodies is dose rate–independent. This is highly likely but difficult to prove in a realistic manner given the limited access to cyclotron beam time. There is a possibility that a longer exposure time (for the same dose) could have a reduced effect on antibody performance due to annealing of radiation damage over time, so the simulation was viewed as an overtest situation.
The use of the Gustav Werner cyclotron at the TSL facility allowed the generation of high-energy particles, but its use was not without some limitations. The entire spectrum of energies is narrowed to a single representative energy value, and the exposure time/runs are based on this value. This was a necessary compromise for both the proton and neutron irradiations and was deemed a satisfactory way to simulate the radiation environment in question.
Excluding activity loss due to sample preparation and handling, the three polyclonal antibodies (anti-GroEL, anti-FITC, and anti-phytanic acid) proved significantly radioresistant, with deactivation appearing only at very high levels of radiation (250 times the nominal mission dose for protons). For the monoclonal antibodies (anti-S. aureus and anti-atrazine scAb), the monoclonal antibody to S. aureus was very sensitive to lyophilization and/or recovery from the glass fiber pads, and this concealed any stand-alone effects of radiation, while in the case of the anti-atrazine recombinant antibody fragment the lyophilization process resulted in over 90% loss of activity, judging from the ELISA results for the storage controls. The 2× combined ExoMars mission dose of radiation (only value tested) did not result in further loss of binding activity, which suggests that this single-chain antibody fragment is able to survive at least mission levels of irradiation.
From the ELISA results, there is no consistent pattern or trend in the way different antibodies react to radiation exposure, even among antibodies of the same type (monoclonal or polyclonal). The conclusion, based on the five antibodies studied, is that while we cannot predict the effect of irradiation on a given arbitrary antibody, in general for any given set of antibodies developed we would expect the majority to easily survive the ExoMars mission dose. This conclusion is based on a small sample size and will require further study. The inability to predict which antibodies will fail under irradiation conditions dictates that all antibodies will need to be validated prior to flight. It is noted that, to allow for a degree of loss of antibody activity, all the antibodies in the final module of the LMC will be loaded into pads at concentrations higher than those required for optimal detection in an inhibition immunoassay, with the acceptance of a necessary level of compromise in the assay detection limits.
In addition to the conclusions on the effect of radiation exposure, a key set of results that emerged from this study concerned the effect of the rehydration of lyophilized antibodies and their washout from the glass fiber. The comparison of antibody binding activity recovered from the storage control samples to the antibody binding activity of the nonlyophilized antibodies (standard curves) allowed the calculation of the amount of loss of binding activity due to the physical effects of sample preparation.
While significant loss occurred, sufficient activity was retained so that the main object of the work could be realized, that is, the retained activity allowed determination of loss due to shipping conditions and radiation exposure. One crucial issue was an inability to differentiate between loss of binding activity due to inefficient physical washout and loss of activity due to antibody denaturation, aggregation, physical adsorption, and so on during lyophilization into the pads.
Of the three protective matrices used in the present study, the Allura Red–based mixture was considered the most successful protectant wherever used, based on the ELISA results obtained. The absence of ascorbate from the mixture did not have a negative effect, which implies that it is not essential for the preservation of antibody functionality under the conditions studied. Following the data analysis from the present work, modified versions of the Allura Red matrix have been integrated in various areas of LMC development (assay optimization, chip-based assays)4.
5. Conclusions
Overall, the ground-based irradiations of immunoassay reagents—specifically antibodies—at simulated radiation levels envisaged for the ExoMars mission, and also at much higher levels, have given valuable information and data input for LMC development. It is apparent that the dominant effect on the loss of activity of five representative antibodies studied was the processing and packaging of the antibodies as freeze-dried preparations in glass fiber pads and their consequent retrieval from the pads for use in ELISA. The subsequent steps of shipping of the samples to an irradiation facility and their irradiation typically contribute far less to the loss of antibody activity. Therefore, it is apparent that further studies of the freeze-drying, packaging, and sample recovery process are required. Initial studies of varying the solutes co-freeze-dried with the antibodies have displayed promise in improving the recovery efficiency. For the radiation effects, the important observation is that, at a representative Mars mission dose, none of the antibodies studied exhibited any evidence of loss of activity due to the radiation. This has been an important step for the LMC instrument development process of demonstrating that the core antibody reagents will survive the radiation environment to be encountered on the ExoMars mission. While this study increases confidence in the use of antibody-based assays in the exploration of the Red Planet, the noticeable loss of antibody activity at higher radiation levels (×250 Mars mission dose) does suggest that further shielding or alterative radiation protection approaches will need to be considered for some other astrobiology targets such as future Europa missions.
Footnotes
Acknowledgments
Funding of the work from the Science and Technology Facilities Council within the UK is gratefully acknowledged. The authors would like to thank Alexander Prokofiev (TSL) and Wojtek Hajdas (PSI).
Author Disclosure Statement
The authors would like to state that no competing financial interests exist.
Abbreviations
BSA, bovine serum albumin; ELISA, enzyme-linked immunosorbent assays; FITC, fluorescein isothiocyanate; GCR, galactic cosmic rays; LMC, Life Marker Chip; SPE, solar particle events; TSL, Theodor Svedberg Laboratory.
1
SPENVIS:
2
The 50 MeV value was chosen taking into consideration the facility's beam capacity.
3
The storage controls were unintentionally transported to Sweden with the shipping control samples.
4
Note that the complex issue of matrix use will be highlighted in a related piece of work by the authors where freeze-drying of antibodies into glass fiber pads for use in a space exposure experiment—flight of the Biopan-6 experiment in low-Earth orbit in 2007—showed survivability when freeze-dried from a buffer-only matrix (unpublished data).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.