
Abstract
The discovery of polygonal terrain on Mars underlain by ice heightens interest in the possibility that this water-bearing habitat may be, or may have been, a suitable habitat for extant life. The possibility is supported by the recurring detection of terrestrial microorganisms in subsurface ice environments, such as ice wedges found beneath tundra polygon features. A characterization of the microbial community of ice wedges from the high Arctic was performed to determine whether this ice environment can sustain actively respiring microorganisms and to assess the ecology of this extreme niche. We found that ice wedge samples contained a relatively abundant number of culturable cells compared to other ice habitats (∼105 CFU·mL−1). Respiration assays in which radio-labeled acetate and in situ measurement of CO2 flux were used suggested low levels of microbial activity, though more sensitive techniques are required to confirm these findings. Based on 16S rRNA gene pyrosequencing, bacterial and archaeal ice wedge communities appeared to reflect surrounding soil communities. Two Pseudomonas sp. were the most abundant taxa in the ice wedge bacterial library (∼50%), while taxa related to ammonia-oxidizing Thaumarchaeota occupied 90% of the archaeal library. The tolerance of a variety of isolates to salinity and temperature revealed characteristics of a psychrotolerant, halotolerant community. Our findings support the hypothesis that ice wedges are capable of sustaining a diverse, plausibly active microbial community. As such, ice wedges, compared to other forms of less habitable ground ice, could serve as a reservoir for life on permanently cold, water-scarce, ice-rich extraterrestrial bodies and are therefore of interest to astrobiologists and ecologists alike. Key Words: Ice wedge—Cryo-environments—Astrobiology—Microbial respiration—Biodiversity. Astrobiology 12, 347–360.
1. Introduction
Polygon wedges occur beneath the troughs of tundra polygonal features visible at the soil surface (see Fig. 1) and cause dome-like protrusions in the polygon centers (Jorgenson et al., 2006). The V-shaped, subsurface wedges form over thousands of years of repeated influx of meltwater or debris, or both, into millimeter-sized cracks created by the thermal contraction of the surrounding soil. Depending on moisture levels and soil composition, wedges are composed of a variety of infill materials, with some wedges consisting predominantly of water (ice wedges), small particulate matter such as sand (sand wedges), or a mixture of both, such as ice-cemented soil (sublimation wedges) (Levy et al., 2009). In wet environments, like our study site, ice wedges form at the permafrost table (soil frozen for greater than two consecutive years) and are typically between 2 and 4 m across at the top, forming over thousands of years. An ice wedge can be overlain by 5–15 cm of soil, referred to as the “transient layer,” which typically undergoes annual thaw (active layer) but can remain frozen depending on inter-annual temperature variation (Shur et al., 2005).

Comparison of the morphological features of Arctic (
Polygonal terrain is common on the martian surface and is broadly distributed at latitudes >30°. According to a recent survey based on satellite photography, the majority of martian polygon morphotypes resemble either sand or sublimation wedges, such as those found in the McMurdo Dry Valleys, which are consistent with the arid conditions on Mars (Levy et al., 2009). These observations are supported by the Phoenix lander's discovery of ice-cemented rock at the polygon site instead of larger ice deposits (Smith et al., 2009). However, satellite photographs also display a number of stark, high-rimmed polygons that are characteristic of terrestrial ice wedge polygons (Levy et al., 2009). This discovery, together with growing evidence for a more dynamic hydrology on Mars (i.e., liquid brine on the martian surface), improves the probability of finding ice-rich subsurface wedges and the ongoing hydrological processes involved in their formation (McEwen et al., 2011). Martian ice wedges are high-priority targets in the search for life beyond Earth, and to expand our understanding of the habitability of such ice environments, we elected to study analog ice wedge environments from the Canadian high Arctic.
The microbial communities of a variety of terrestrial cryo-environments have been studied, such as those found in glacier ice (Skidmore et al., 2000; Miteva et al., 2004), subglacial environments (Yde et al., 2010), cryoconite holes (Säwström et al., 2002), Arctic permafrost soil (Steven et al., 2009; Wilhelm et al., 2011), Antarctic Dry Valley soils and endoliths (Pointing et al., 2009), cryopegs (Gilichinsky, 2002), ice shelves (Bottos et al., 2008), high Arctic saline perennial spring environments (Perreault et al., 2007; Niederberger et al., 2009, 2010), and various other ice environments (Lacelle et al., 2011). These studies reveal complex cold-tolerant microbial communities, some of which exhibit active metabolism at in situ temperatures in ice and brine (−5°C) (Niederberger et al., 2010; Bakermans and Skidmore, 2011) and permafrost (−15°C) (Steven et al., 2008). Ground ice, such as ice wedges, is one of the most inhospitable of cryo-environments due to a lack of sunlight, scarcity of liquid water, scarcity of energy sources, and subzero temperatures, and may in fact be a poor microbial habitat (Steven et al., 2008). However, ice vapor or ice brine may provide the necessary solvent for life in habitats beyond Earth, with ice commonly found in environments within the Solar System (Prockter, 2005): on planetary bodies such as Europa, Enceladus, and Mars (Rathbun et al., 1998; Fairen et al., 2010), and on satellites such as Tempel 1 (Sunshine et al., 2006).
On Earth, microbial life has been isolated from ice environments, such as glacier ice, ground ice, and snow environments, many of which were thousands of years old (Table 1). Ice contains a network of liquid water veins (brine veins) that can transport soluble and insoluble particles and support life within the ice (Mader et al., 2006). Isotopic analyses of gas trapped in ancient ice wedges have demonstrated that active microbial respiration occurs within the ice structure (Lacelle et al., 2011). Two preliminary studies have examined the culturable microbial community and the total diversity of ice wedges: one study involved a 25,000-year-old Alaskan ice wedge (Katayama et al., 2007) that contained a relatively abundant population (105 to 106 CFU·g−1 ice), and another involved a 5,000- to 6,000-year-old Yukon ice wedge (Lacelle et al., 2011) that yielded a relatively diminutive population (∼103 CFU·mL−1 melted ice). In comparison, a variety of other ground ice types have not yielded any culturable cells (Table 1). Yet despite a relative abundance of culturable cells, the Alaskan ice wedge bacterial 16S rRNA clone library had low taxonomic diversity, which consisted almost entirely of Gammaproteobacteria (93%) and is, to date, the only assessment of ice wedge bacterial diversity. Two studies of similar environments, a high Arctic massive ground ice (Steven et al., 2008) and a Yukon pingo (Lacelle et al., 2011), discovered a markedly different diversity of ice-inhabiting microorganisms, with taxa belonging predominantly to Firmicutes (∼60%). There appears to be distinct variability in the microbiota of ice-rich environments, which suggests that conditions within ice can significantly differ.
Viable cell counts are measured in CFU·mL−1 or CFU·gdw−1 (colony-forming units per gram dry weight), while total cell counts are in cells·mL−1 or cells·gdw−1. Act, Actinobacteria; Fir, Firmicutes; Prot, Proteobacteria; D.T., Deinococcus-Thermus.
Mineralization of [1−14C] acetate was observed.
Obtained isolates by alternate culturing methods.
Detection of cellular respiration is an important line of evidence for whether cryo-environments, ice wedges in particular, can support active life or whether they merely preserve it. Microbial respiration in ice wedges has already been inferred based on isotopic ratios of Ar-O2-CO2 (Lacelle et al., 2011), though these methods did not determine the timeframe of the microbial activity, that is, whether respiration occurred during the initial stages of ice crystallization or it occurred periodically or steadily over time. The mineralization of radioisotope-labeled substrates at in situ temperatures has been used to detect microbial respiration with high sensitivity in a number of cryo-environment samples, including ground ice (Steven et al., 2007, 2008), and were used in the present study to detect activity in ice wedge samples.
From an astrobiological perspective, whether ice wedges support active microbial communities will be important in identifying possible refugia beyond our hospitable planet. Based on two ice wedge cores obtained from high Arctic polygon terrain, our objective was to determine the microbial composition along a depth gradient, using culture-dependent and culture-independent approaches, and determine whether ice wedge microbial communities are metabolically active in situ. The conditions within terrestrial ice wedges, while severe, are inviting to life when compared to the physical conditions expected in martian wedge environments, where microorganisms would have to manage water scarcity, drastic daily temperature fluctuation, low pressure, and high levels of radiation (Gómez et al., 2010). Therefore, the characterization of in situ respiration and the microbial community of terrestrial ice wedge habitats represents a best-case scenario in the search for life on Mars and will broadly improve our understanding of the ecology of ice-rich habitats.
2. Methods
2.1. Description of site and sampling methods
Sampling was conducted in July 2009 on polygonal terrain located near the McGill Arctic Research Station on Axel Heiberg Island, Nunavut, Canada. The mean annual air temperature for the region, recorded at the closest weather station in Eureka, NU, was −17.6°C in 2009. Ice wedge samples were taken in a polygon field characterized by acidic soil (pH 4.5–5) adjacent to an acidic lake (pH 3.7), with pH driven by iron-sulfur chemistry (Buttle and Fraser, 1992). Two troughs running alongside a soil polygon, a feature created by the pressure exerted during ice wedge formation, were chosen for coring (AH1: 79°24.963′N, 90°45.759′W; AH2: 79°24.965′N, 90°45.802′W). Both troughs were cracked down the center, which indicates that the ice wedges beneath are still actively forming. The active layer was 45 cm deep at the time of sampling, while the transient layer was 10 cm thick overlying the ice wedge. Cores were taken by using a combination of a Hilti hammer drill fitted with a 3-inch corer and a Pioneer portable hydraulic drill (Multi-Power Products Ltd., Kelowna, BC, Canada). Each corer was sterilized with ethanol prior to extraction of each sample. The AH1 borehole reached a depth of 345 cm, whereas the AH2 hole reached 155 cm deep. Cores were transported in sterile Whirl-Pak bags (Nasco Laboratory Sampling Products, Modesto, CA, USA) and kept frozen at −20°C until processing.
The interior of each core was subsampled to ensure the material used in our analyses was not subject to contamination. This was performed according to the process outlined by Juck et al. (2005) with minor changes. Fluorescent microspheres were not used, but rather the exterior of each core was painted with a liquid culture (∼108 cells·mL−1 in TSB) of a kanamycin-resistant, green fluorescent protein-labeled (gfp) strain of Pseudomonas, strain Cam1-gfp2. To verify that there had been no contamination, a portion of the subsample was melted and plated on LB agar +50 μg/mL kanamycin to verify that no kanamycin-resistant colonies grew. Additionally, following each DNA extraction, a polymerase chain reaction (PCR) with primers specific to the gfp gene (gfpF, gfpR) was performed to verify the absence of gfp gene artifacts in the sample; PCR conditions and primers were as previously described. The percentage of solid material in the ice wedge was calculated based on mass, where 15 g of ice from AH1 at a depth of 245–260 cm was dried at 100°C for approximately 24–32 h and weighed. Conductivity, salinity, total dissolved solids, and oxidation/reduction potential were also measured from this sample with the YSI 556 Multi Probe System (YSI Incorporated, Yellow Springs, OH, USA).
2.2. Community DNA extraction and 16S rRNA gene 454 pyrosequencing analysis
An ice wedge sample (AH1 2.2.5) from the deepest portion of recovered ice (325 cm) was chosen for comprehensive community analysis by 16S rRNA gene pyrosequencing. To concentrate biomass prior to DNA extraction, 15 mL of melted ice was passed through a Sterivex GP 0.22 μm filter (Millipore, Billerica, MA, USA). The filter was then sectioned into 10, and DNA was extracted by using the UltraClean Soil DNA Isolation Kit's (Mo Bio, Carlsbad, CA, USA) alternative protocol for maximum yields. The 10 individual extractions were pooled onto a single spin column prior to the final elution step to maximize DNA yields. DNA samples were sent to Research and Testing Laboratories, LLC (Lubbock, TX, USA) for tag-encoded FLX-titanium amplicon pyrosequencing (Roche 454). 16S rRNA gene taxonomic profiles for Bacteria and Archaea were created with primers “Assay b.2: 28F-519R,” for Bacteria (28F: 5′-GAG TTT GAT YMT GGC TC & 519R: 5′-GWA TTA CCG CGG CKG CTG), and “Assay a.1: 340F-806R,” for Archaea (5′-CCC TAC GGG GCG CAG CAG & 5′-GGA CTA CCA GGG TAT CTA AT). The Ribosomal Database Project (RDP) pyrosequencing pipeline was used to pre-process 16S rRNA sequences (ensuring only high-quality sequences were used in analysis) and to provide the diversity estimates and classification of all taxa (at 97% sequence similarity) (Cole et al., 2009).
2.3. Total cell and culturable heterotrophic cell counts
Total cell counts were made on the transient layer (55–56 cm) and shallow ice wedge samples (125–135 cm) from both cores with a 4,6-diamidino-2-phenylindole (DAPI) cell stain and fluorescence microscopy. DAPI was added to 0.5 mL replicate melted ice sample to a final concentration of 100 ng/mL. The stained samples were then filtered onto white polycarbonate filters, washed with 2 mL of ddH2O to remove background stain, and visualized (λ=461 nm) with a Nikon Y-FL epifluorescence microscope at 1000×magnification. Cell counts were averaged over 10 randomly chosen fields of view.
Culturable heterotrophic cell counts were performed on 14 sections of the AH1 and AH2 ice cores by using a variety of media and at two incubation temperatures, 25°C and 5°C. Samples were serially diluted in 0.1% sodium pyrophosphate and plated in triplicate onto R2A (7.2±0.2), R2A supplemented with 5% w/v NaCl, R2A (pH 5.5±0.1), and TSA (7.2±0.2). Incubation times varied and were left until no new growth was observed, corresponding to 14–21 days for incubations at 25°C and 40–60 days for incubations at 5°C. Samples were plated on LB agar with 50 μg/mL kanamycin to test for contamination from the Pseudomonas Cam1-gfp2 tracer strain; any sample with a positive contamination was reprocessed. A one-way ANOVA was performed comparing colony-forming units (CFU) per core (AH1, AH2) and temperature (5°C and 25°C) at each depth with ezANOVA (
2.4. Selection, identification, and characterization of bacterial isolates
Isolates were characterized from ice wedge core AH1 from all media types and incubation conditions based on colony morphology, color, growth media, incubation temperature, and time of appearance. Each colony was streaked for isolation three times to ensure purity. The classification of bacterial isolates was made by PCR amplifying and sequencing 16S rRNA genes according to Steven et al. (2008) with the following primers: 27F (5′- AGA GTT TGA TCC TGG CTC AG-3′) and 758R (5′- CTA CCA GGG TAT CTA ATC C-3′). PCR product was sequenced by Sequencage Laval (University of Laval, Quebec City, QC, Canada) with an ABI Prism 3130XL genetic analyzer. Isolates were tested for growth on R2A at various temperatures (−5°C, 5°C, 25°C, 37°C) and salt amendments (1%, 2%, 3%, 4%, 5% w/v NaCl).
2.5. Detection of microbial activity at cold temperatures
The microbial activity of ice wedge, active layer, and transient layer communities was compared by using radio-respiration assays at low temperatures (−10°C, −5°C, and 5°C) as described for high Arctic permafrost samples by Steven et al. (2007, 2008). Due to a limited supply of ice sample, only one radio-labeled substrate was used, [1−14C]acetate, which has been used successfully in radio-respiration assays of accretion ice over Lake Vostok in Antarctica, permafrost bacteria, and glacial microorganisms. Four samples were analyzed: active layer from above AH1 (depth of 30 cm); transient layer from above AH1 (55–56 cm); and two ice wedge samples—shallow (AH2: 60–75 cm) and deep (AH1: 245–260 cm). These were prepared in triplicate with sterile controls for each temperature. A total of 5 mL of sample was added to each microcosm along with radio-labeled acetate, which yielded an average of 57,285 disintegrations per minute per microcosm. Non-radio-labeled acetate was included to a final concentration of 15 mM to stimulate growth. Incubations were sampled periodically over a period of 164 days. The percentage of radio-labeled acetate mineralized to 14CO2 was compared between sample and controls by using an unpaired t test.
2.6. In situ measurements of CO2 flux from surface of field sites
Net CO2 flux was measured from the soil surface at the field site with a LiCor LI 8100 Field Survey Unit (LI-COR Biosciences, Lincoln, NE, USA), according to manufacturer's instructions. Flux measurements recorded with the LiCor system accounted for variation in surface soil temperature (∼5 cm deep) and atmospheric CO2 concentration at the time of sampling. Measurements were performed during 2 days in the summer (mid July, 2009) and late winter (late April, 2010). Readings of net CO2 flux (3×3 min replicates) were measured at three different sites on the surface of polygon troughs, where ice wedge samples were extracted, and on the surface of polygon centers, underlain by permafrost. Mean CO2 flux values were compared per sampling year/season by using a student t test for the polygon center versus trough surface soil.
2.7. Nucleotide sequence accession numbers
The accession numbers for the nucleotide sequences deposited in the GenBank database are as follows: bacterial isolates (JN221486–JN221547), bacterial 16S rRNA gene library (JN217243–JN219938), and archaeal 16S rRNA gene library (JN219939–JN221485).
3. Results
3.1. Ice wedge characteristics and DNA extraction
The age of the ice wedges is estimated to be at least 4000 years before present, based on the Holocene deposits in which they have formed (Aitken and Bell, 1998) and ice wedge dating at regionally proximal sites (Fortier and Allard, 2004). Vertical cracking patterns within the ice core structures were apparent and are consistent with the process of ice wedge formation (Fig. 1). Other ground ice structures observed included either horizontal layering with no striations or horizontal layering with random striations (resulting from the crystallization of snow, i.e., firnification). The in situ temperature of the ice wedge was −16°C during the summer sampling period. Measurements of pH for all ice wedge and transient layer sections ranged from 5.2 to 5.8. The salinity, conductivity, oxidation/reduction potential, and total dissolved solids of the ice wedge sample were as follows: 0.20 ppt, 373 μS·cm−1, +186.0 mV, and 0.274 g/L. On average, the ice wedge contained extremely low percentages of solid mass (∼0.2% w/v±0.07). The PCR screen to detect the intentional contaminant strain, Pseudomonas Cam1-gfp2, was negative during all molecular analyses, which indicates sample purity.
3.2. Abundance of total and culturable cells
Total cell counts ranged across one order of magnitude, from 8.4×107 to 9.7×108 cells·mL−1, while culturable cell counts were highly variable, ranging from 70 CFU·mL−1 to 1.1×106 CFU·mL−1 (Fig. 2). The greatest abundance of culturable aerobic heterotrophs were cultivated at 5°C from the AH1 transient layer soil. Culturable counts at 5°C were significantly higher (p=<0.1) than those at 25°C for nearly all profile depths, which indicates that the heterotrophic ice wedge community consists predominantly of cold-adapted bacteria. Culturable cell counts along the core profile revealed an inverse relationship between the number of culturable heterotrophs and the sampling depth. Culturable versus microscope-based “total” cell counts differed, at most by a factor of 104, which indicates the presence of a largely uncultured population. No clear delineation was found between the abundance of culturable and total cells between the transient layer soil and ice wedges, since both enumerations were greater in the ice wedge for core AH2, while the opposite was true of AH1. The type of solid growth media used, that is, R2A, R2A +5% NaCl, R2A +pH 5.5, and TSA, influenced the recovery of culturable cells. In all cases, the greatest growth was observed on R2A plates, and these cell counts form the basis for Fig. 2.
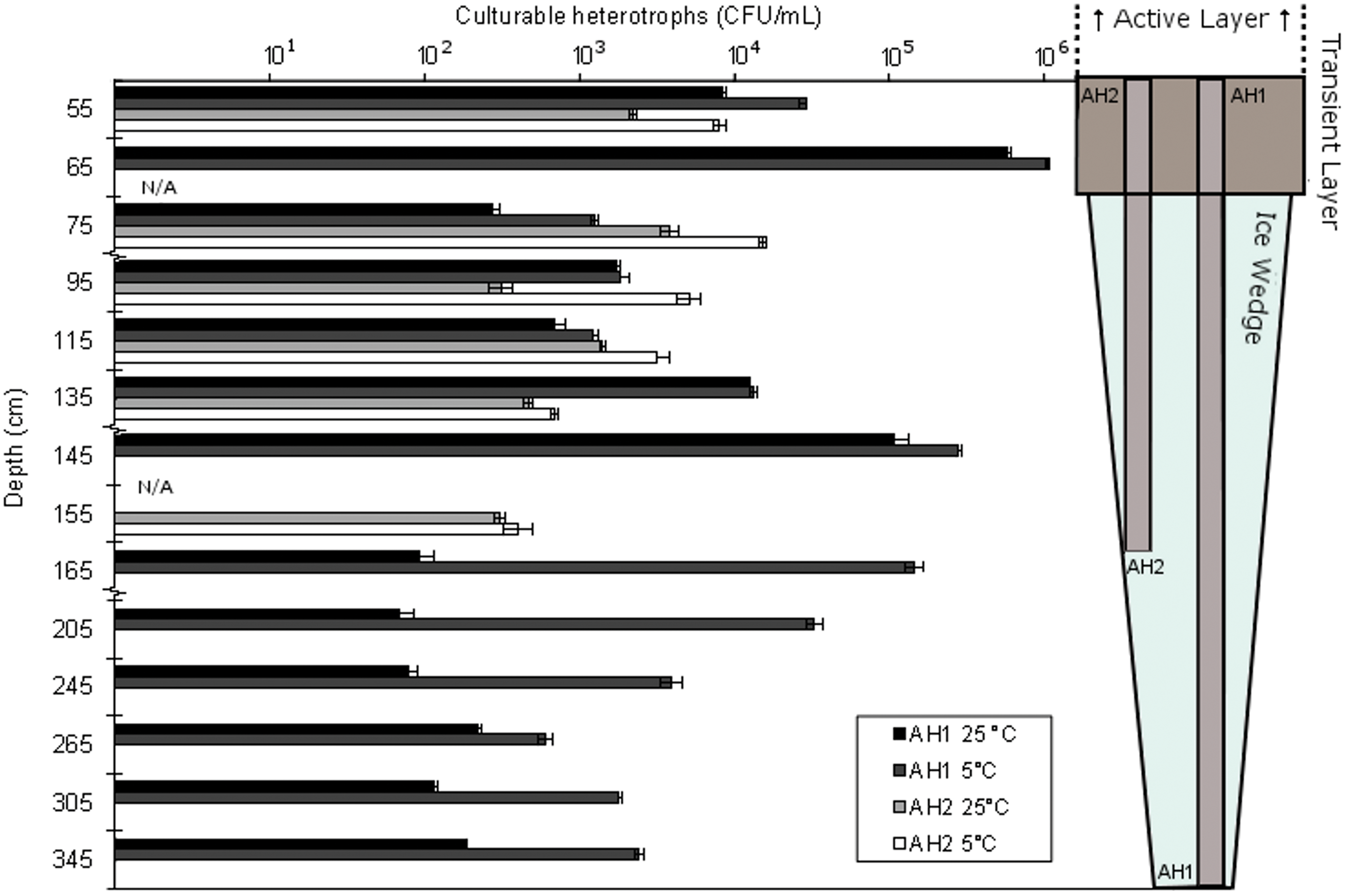
Heterotrophic plate counts on R2A growth medium at 5°C and 25°C along the depth profile of the two ice wedge cores (AH1 and AH2). Error bars represent standard error across triplicates. Note that the y axis is not to scale. Color images available online at
3.3. Isolate identification and characterization
A total of 25 transient layer and 37 ice wedge isolates were selected from AH1 growth plates for 16S rRNA gene sequencing and characterization. Arthrobacter sp., Paenibacillus sp., Shigella sp., and Curtobacterium sp. were the most abundant (each constituted ∼10% of total isolates) and most cosmopolitan species isolated and occurred in both transient layer and ice wedge samples (Table 2). A number of isolates were also detected in the 16S rRNA gene bacterial library (in bold type in Table 2), including one Pseudomonas sp. (P. migulae) that constituted ∼50% of all bacterial sequences. All isolates were capable of growth at 25°C, and nearly all isolates could grow at 5°C. Three isolates were capable of growth at 37°C, two of which (Pseudomonas sp. and Arthrobacter sp.) exhibited the broadest temperature tolerance and grew at 5°C as well. Isolates incubated at −5°C did not grow after 2 month's incubation. The majority of isolates (65%) demonstrated mild halotolerance (3–5% NaCl), while 33% were capable of growth at 5% NaCl.
Classifications are based on the Ribosomal Database Project and top cultured BlastN hits, which corresponded for the majority of isolates. Isolates with greater than 95% sequence similarity were aggregated and, as a result, growth temperatures and salt tolerance values represent the broadest range observed for each aggregated genotype. Isolates in bold share 97% similarity to sequences found in the 16S rRNA gene library.
Classification derived solely from top cultured BlastN hit.
N/A, not available.
3.4. 16S rRNA gene community analysis
Archaeal and bacterial 16S rRNA gene pyrosequencing was performed on core subsection 2.2.5, the deepest ice wedge sample, to provide a description of the abundance and community structure therein. The bacterial community library was composed of 2690 sequences and exhibited high diversity based on the Chao (872) and Shannon indices (4.10). The bacterial library provided reasonable coverage of the diversity of organisms present in the ice wedge sample. A total of 591 operational taxonomic units (OTUs) were present at 97% sequence similarity with a Good's percent coverage suggesting an estimated sampling of 75.2% of the community (Good, 1953). The community was dominated by Gammaproteobacteria, classified as Pseudomonas, and comprised ∼50% of the total of classifiable Bacteria. The remainder of the taxa were distributed across 10 other phyla, the most abundant of which were Actinobacteria (15.2%), Acidobacteria (8.3%), and Alphaproteobacteria (7.9%) (Fig. 3). The majority of 16S rRNA sequences classified as Pseudomonas belong to a single OTU, the closest relative of which is Pseudomonas migulae (98% similarity), a pseudomonad isolated from a low-nutrient mineral spring in France (Verhille et al., 1999). This type strain is capable of denitrification, possesses a flagella, and grows at 4°C. In analyzing all OTUs occupying greater than 0.5% of the total library, the following environments were represented: Antarctic and Arctic soils, aquatic environments (lakes, springs, wastewater), permafrost, alpine soils, agricultural soil, and human skin. A substantial proportion of the bacterial 16S rRNA gene library (12.4%) consisted of unclassifiable sequences.
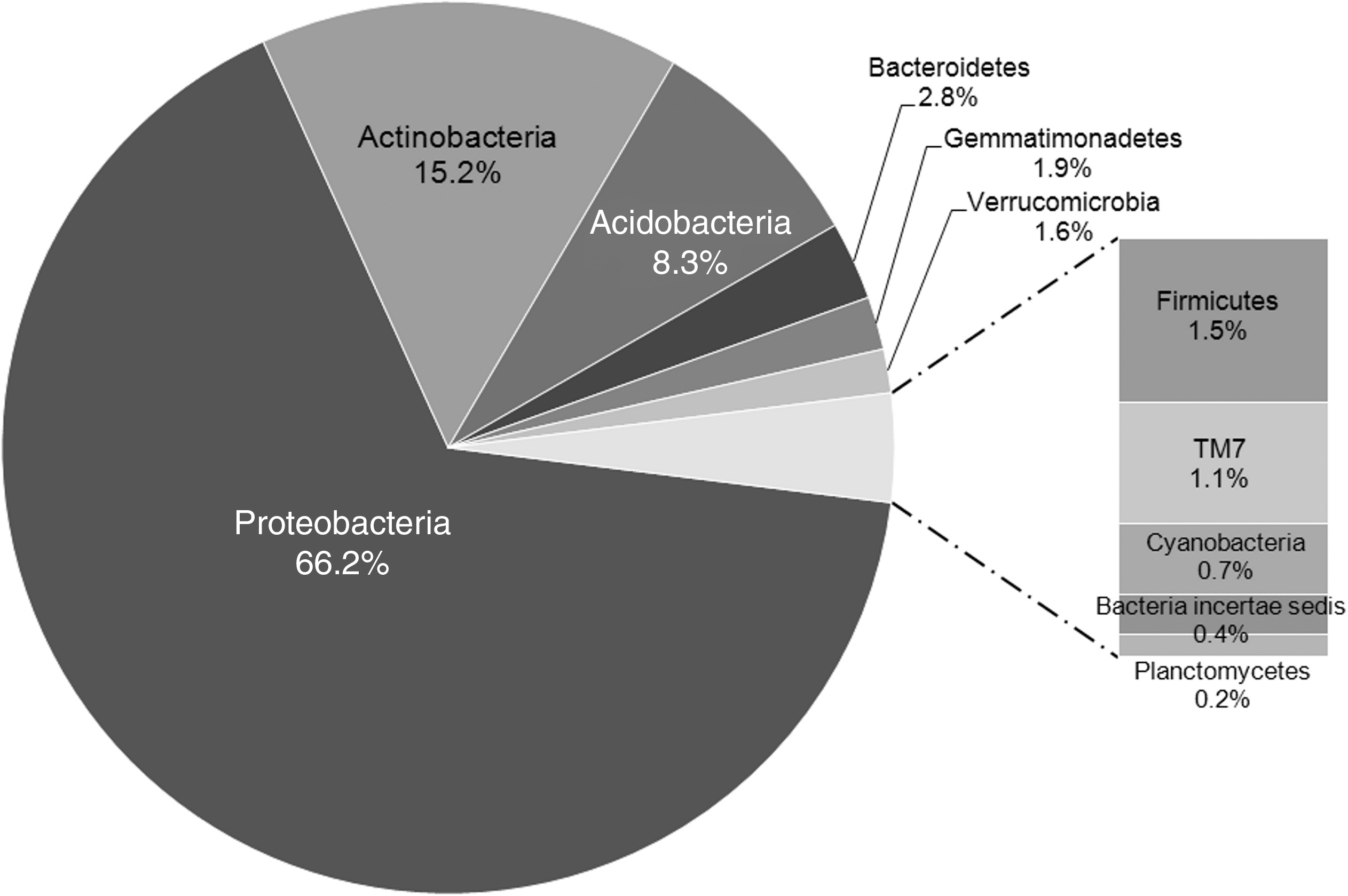
Distribution of bacterial taxa, at a depth of 325 cm in the ice wedge, displaying the predominance of Proteobacteria and range of bacterial diversity (n=2396). The phylum Proteobacteria was comprised of two major classes: Gammaproteobacteria (83%) and Alphaproteobacteria (12%). Unclassifiable 16S rRNA gene sequences were not included in the analysis (12% of all sequences).
The archaeal pyrosequencing data contained only 21 archaeal sequences and consequently exhibited very low diversity, according to Chao (18) and Shannon indices (2.14). Out of the total 1547 “archaeal” sequences, 1526 sequences were classified as bacterial. According to the experiences of the pyrosequencing facility, poor archaeal primer specificity results when the abundance of archaeal sequences is minor relative to bacterial sequences. The 21 archaeal sequences grouped into 11 OTUs at 97% similarity, the majority of which (9) were related to Crenarchaeota of the class Thermoprotei (clustering to marine Group 1.3b). The closest cultured match (95%) was with a member of Thaumarchaeota, Nitrososphaera viennensis, an ammonia-oxidizing soil archaeon (Tourna et al., 2011). The remaining two OTUs were unclassifiable beyond the level of Euryarchaeota, with top cultured relatives from the genera Methanococcus and Methanosarcina.
Similarities in community structure were apparent following direct comparison of our data to 16S rRNA gene libraries from another ice wedge (Katayama et al., 2007) and from the active layer and permafrost at the same Axel Heiberg field site (Wilhelm et al., 2011). The major Pseudomonas sp. phylotype (∼50% of sequences, n=2690) in the Axel Heiberg ice wedge was also present in the Alaskan ice wedge library (97% similarity), though at a relatively lower abundance (6%, n=79); this was the only overlap between the two ice wedge libraries. A low degree of overlap was also seen with the libraries obtained from the same field site, where 18 bacterial (3% overlap) and 3 archaeal (14% overlap) OTUs were common to both. Members of the bacterial families Bradyrhizobiaceae, Chitinophagaceae, Acidobacteriaceae, Gemmatimonadaceae, and Acidothermaceae were shared; three OTUs of unclassified Bacteria were also shared. All three archaeal OTUs were classifiable to class Thermoprotei and constituted the major archaeal taxa found in both libraries.
3.5. Microbial activity of ice wedge samples by radio-labeled acetate mineralization
The relative mineralization rates of trough and ice wedge samples are summarized in Fig. 4. Both ice wedge samples, shallow and deep, did not show any significant respiration at any temperature. At 5°C, the transient layer demonstrated the greatest total mineralization (17.3%) compared to the active layer (10.6%). The active layer (p=0.01) and transient layer (p=0.01) were significantly different than controls, while both ice wedge samples were not (p>0.05; p>0.05). At subzero temperatures, −5°C and −10°C, no sample differed significantly from its control after 164 days of incubation.
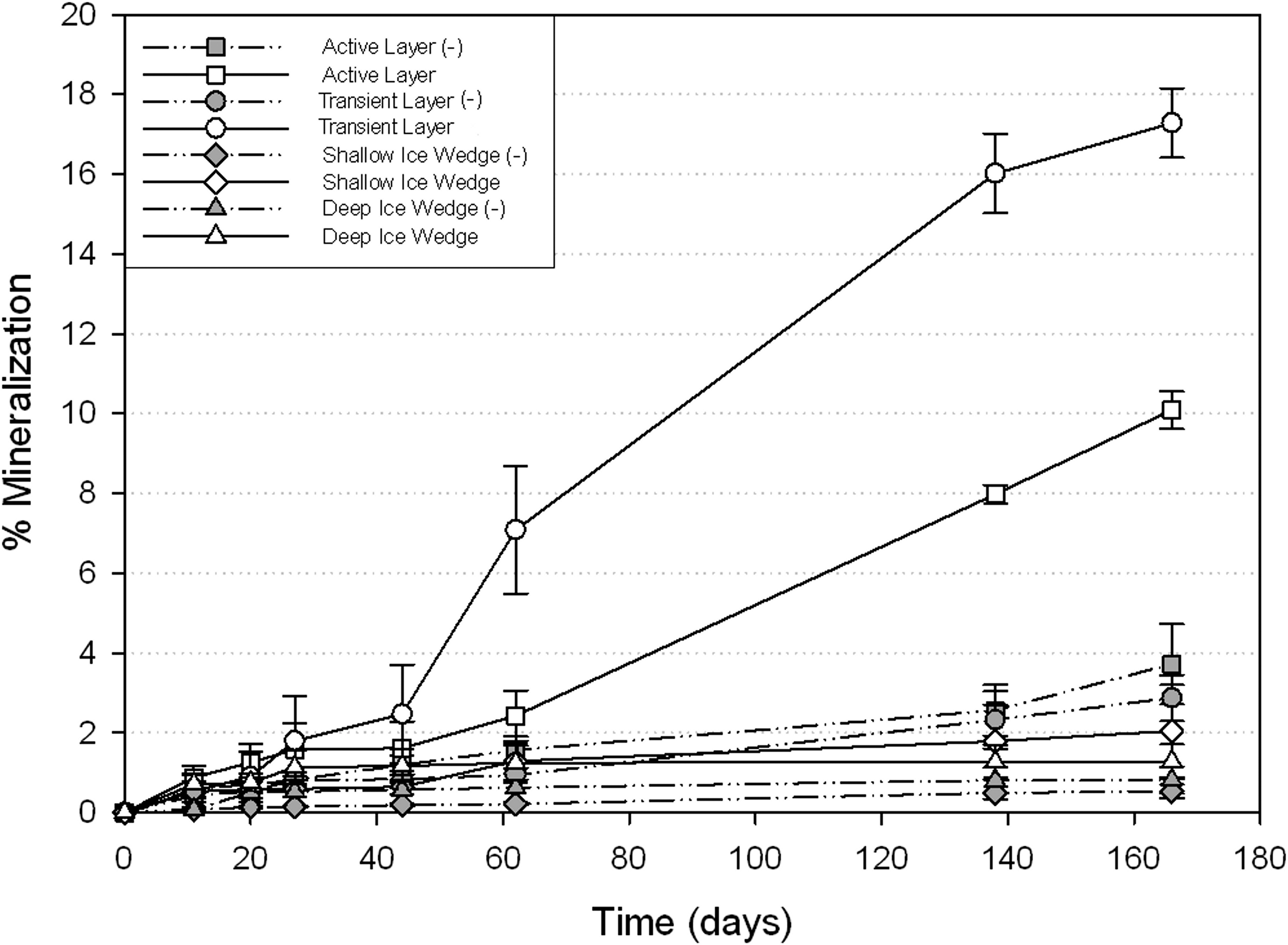
Percentage of radiolabeled acetate mineralized to CO2 at 5°C for active layer soil (depth of 30 cm); transient layer soil (55–65 cm); and two ice wedges—shallow (AH2: 60–75 cm) and deep (AH1: 245–260 cm). Transient layer and active layer samples were significantly different from their controls. The notation (−) denotes sterile negative controls, and error bars represent standard error across triplicates.
3.6. Microbial activity by in situ net CO2 flux
Carbon dioxide flux was measured to estimate in situ microbial activity (respiration) at different locations along polygon and trough surfaces in summer (2009) and late winter (2010) (Fig. 5). Active flux of CO2 was observed during both sampling periods at significantly higher levels above atmospheric values, successfully identifying the product of microbial respiration in situ. Variability in CO2 flux was greater in summer and showed significantly higher flux between trough versus polygon active layer surfaces, averaging 2118±805 mg/m2/day and 853±548 mg/m2/day, respectively (p=0.01). In late winter, flux values for the polygon (1637±579 mg/m2/day) were similar (p>0.05) to the trough (1682±446 mg/m2/day). Throughout the measurement period, the maximal single flux value observed was 4600 mg/m2/day at the trough surface. The greatest average flux value was measured at the surface of an ice wedge positioned directly over a borehole with a depth of 155 cm (2604±456 mg/m2/day) (Fig. 5) (p<0.01). During summer flux analyses, the soil temperature profile ranged from 1°C at a depth of 45 cm to 14°C in the upper 5 cm of the soil. During late-winter flux analyses, the ground was completely frozen, and soil temperatures were measured at −16°C at 40 cm deep and −9°C in the upper 5 cm of soil.
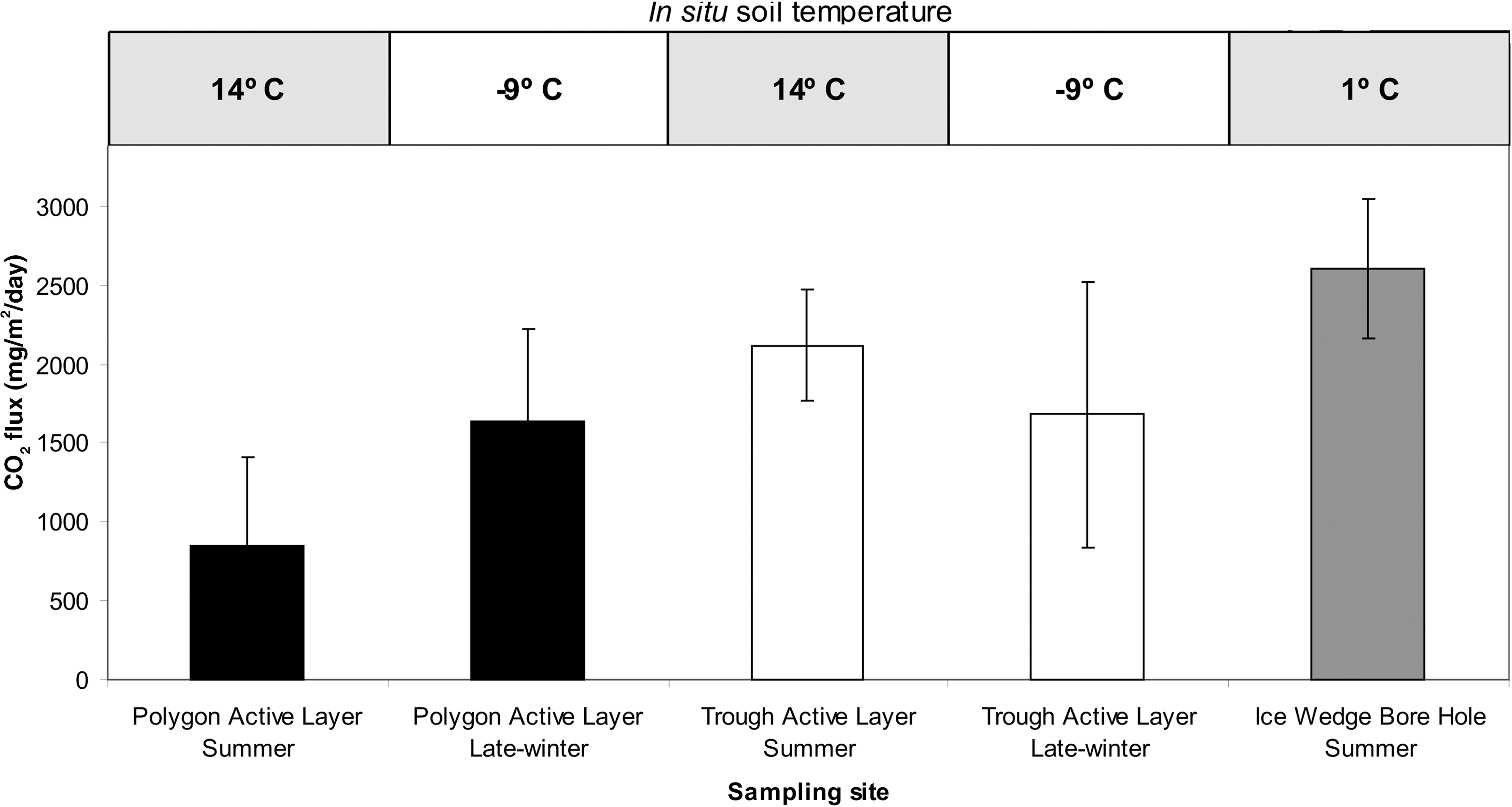
Mean net CO2 flux from three site types during summer (2009) and late winter (2010): polygon center, polygon trough (ice wedge), and the ice wedge surface above a borehole. Error bars represent the standard deviation of the mean from the total data collected per site type.
4. Discussion
Pursuing evidence of extraterrestrial life in ice environments on Mars, or elsewhere, is possible through characterizations of analogous environments on Earth (Fairen et al., 2010). Terrestrial analogue environments provide improved logistical foresights, reveal priority targets like hotbeds of biological activity, and are useful in broader criteria development for assessing habitable zones on planetary bodies. By studying the microbial community capable of surviving the conditions in ice wedge environments, where liquid water is at its limit of stability, we gain insight into (i) the physiological limits of cold-tolerant life, (ii) characterizing the visual features of extremophilic microorganisms, and (iii) studying the biosignatures left by certain types of microbial activity. Community differences may also be used to infer geochemical and geological differences in ice environments, and provide insight into more habitable versus less habitable ice conditions.
Our characterization of Axel Heiberg ice wedges indicated that an extant, diverse, and low-activity microbial community exists within the ice wedge habitat. Ice wedges yield the greatest number of culturable heterotrophic cells compared to other ice environments, including glaciers, pingos, lake ice, ground ice, and snow (Table 1). It is still unclear whether this difference in abundance is due to relatively more hospitable conditions in ice wedges or a reflection of the population sizes in the source of water. It is a reasonable assumption that glacier and snow ice, composed of cells derived from particles carried by wind, may have lower cell abundances compared to ice wedges, which result from water percolating through soil (Xiang et al., 2009). The abundance of culturable heterotrophs in the Axel Heiberg ice wedges is corroborated by findings from an Alaskan ice wedge, which demonstrates that ice wedge populations remain high in the more extreme conditions of the high Arctic. The suitability of the Axel Heiberg ice wedge habitat for microbial life is apparent when contrasted to massive ground ice, which is defined as horizontally extensive subsurface ice greater than 2 m thick (Steven et al., 2008). This proximal high Arctic site yielded zero culturable cells despite significant parallels in culturing technique (Steven et al., 2008). The higher abundance of culturable cells in ice wedges may result from the diffusion of annual influxes of meltwater from the center of the ice wedge through the network of ice veins that carry nutrients and oxygenated water (Mader et al., 2006). The uniformity of the ice wedge culturable cell counts, which vary only 10-fold along the 3 m gradient, may be indicative of the effect this diffusion has on supporting the microbial community throughout the ice wedge. In comparison, permafrost soils show differences of 100- to 1000-fold in abundances across a much shorter distance (∼1 m) (Steven et al., 2006; Gilichinsky et al., 2008). However, it is important to note that the uniformity of ice wedge communities may also reflect the fluctuation or constancy in the deposition rate of annual influxes, an effect that has been correlated to microbial abundances in the formation of glacier ice (Xiang et al., 2009).
The pattern of microbial diversity observed in the Axel Heiberg ice wedge at a depth of 3 m was consistent with other ice environments for both culture-based and 16S rRNA gene-based assays, namely, relatively high bacterial and low archaeal diversity (Steven et al., 2009; Wilhelm et al., 2011). In nearly all studies of the culturable community of ice environments, including the Axel Heiberg ice wedge, Actinobacteria have been the most commonly cultured phylum and Arthrobacter sp. the most common genus. Arthrobacter sp. are known to be hardy, cosmopolitan organisms capable of surviving long periods of high stress such as starvation, exposure to radiation, and temperature shifts, and are well adapted for living in the extreme conditions of ice wedges and other cryo-environments (Nelson and Parkinson, 1978; Mongodin et al., 2006). Arthrobacter psychrolactophilus, a psychrotolerant organism, demonstrated an impressive hardiness when exposed to Mars-like conditions; from a starting population of 109 cells·g−1, a population of 105 cells·g−1 survived 30 days of fluctuating diurnal temperature (−40°C to 24°C), low pressure (13.3 mbar), and high background UV radiation (Johnson et al., 2011). Bacterial diversity estimates based on 16S rRNA gene libraries placed the Axel Heiberg ice wedge community (H’: 4.10) at a similar level of diversity as Axel Heiberg permafrost (H’: 4.00) (Wilhelm et al., 2011) and an Arctic ice shelf microbial mat (H’: 4.41) (Bottos et al., 2008). However, the marked abundance of Pseudomonas sp. in the Axel Heiberg library (50%) indicates an uneven distribution of taxa with a few species that dominate the community. Similar findings were reported in the Alaskan ice wedge (>60% Pseudomonas spp.) (Katayama et al., 2007) and a glacier on Mt. Everest (Zhang et al., 2010). For cryo-environments that preserve extracellular DNA, it is important to gauge diversity estimates carefully, as the species richness can be skewed by singleton sequences that may represent remnants of ancient DNA, sequencing noise (Tedersoo et al., 2010), or perhaps low-abundance organisms of the rare biosphere (Huse et al., 2010).
The abundance of Pseudomonas sp. in all ice wedges studied to date is compelling evidence and affirms the need for examination of their particular role in this niche. The two most predominant Pseudomonas spp. in our 16S rRNA gene library are both most closely related to Pseudomonas miguale (98%). Previous characterizations of P. miguale do not suggest a ready explanation as to why it might be so abundant in ice (Verhille et al., 1999); however, it is a member of the P. fluorescens group (Anzai et al., 2000), known for their capacity to induce ice nucleation (Obata et al., 1987). This phylogenetic evidence implies an adaptive advantage for ice-nucleating bacteria in ice wedge habitats. The benefits conferred by ice nucleation would be two-fold: (i) heat is released during the crystallization of water molecules, (ii) ice nucleation can control the size and growth of ice crystals and thereby protect the cell from physical damage (Zachariassen and Kristiansen, 2000). Ice nucleation has been observed by intact cells at subzero temperatures as low as −4°C (Yankofsky et al., 1981) and is not affected by low pH (≥5.0) (Obata et al., 1987), such as that of the Axel Heiberg ice wedges. The predominance of pseudomonads closely related to ice-nucleating varieties in both Axel Heiberg and Alaskan ice wedges (Katayama et al., 2007) suggests a plausible role for this activity.
The detection of Archaea, which are known for their adaptation to extreme temperature and salinity, was limited by the minimal data recovered during pyrosequencing analyses. The inability to recover archaeal sequences reflects the relatively low abundance of Archaea compared to Bacteria. Based on the sequences recovered, the community was composed almost entirely of Crenarchaeota, which are phylogenetically related to marine Group 1.3b (90%). The same high levels of Crenarchaeota were reported in a study of the neighboring Axel Heiberg wetland soil (Wilhelm et al., 2011) and have been found in a wide variety of cryo-environments, such as Ellesmere Island permafrost (Steven et al., 2008), Norwegian Arctic wetland soil (Hoj et al., 2005), Arctic and Antarctic oceans (Kalanetra et al., 2009), and soils from the Ross Sea region of Antarctica (>99% of clones) (Ayton et al., 2010). The top matching cultured representative to the Crenarchaeota detected in the Axel Heiberg ice wedge is a soil archaeon, Nitrososphaera viennensis (95%), which is capable of autotrophic ammonia-oxidation (Tourna et al., 2011). Ammonia-oxidizing Archaea are thought to contribute significantly to nitrogen cycling on a global scale in both terrestrial and marine environments (Tourna et al., 2011). The recurrence of ammonia-oxidizing Archaea in cryo-environments and in ice suggests their possible ecological importance, though their activity at subzero temperatures is unknown.
The constituency of an ice wedge community is expected to include a significant proportion of immigration from the local environment, as soil particles and meltwater from the surface and active layer percolate through the soil during thermal cracking (Fortier and Allard, 2005). The Axel Heiberg archaeal and bacterial libraries reflect this in the large proportion of taxa that are not present in the Alaskan ice wedge library (Katayama et al., 2007) but are abundant in libraries made from active layer soil at the same study site (Wilhelm et al., 2011). Members of Acidobacteriaceae and Gemmatimonadaceae, for example, were not detected in the ice Alaskan wedge, yet they were prevalent in the Axel Heiberg ice wedge, 18% and 1.9%, respectively, and were dominant community members of soil from the same site (∼50 m of site separation), 30% and 14%, respectively (Wilhelm et al., 2011). A number of sequences recovered from the soil and ice wedge were common to both sites at the species level, which demonstrates that the microbial community, in part, originates from the surrounding environment.
The use of in situ CO2 flux and lab-based radio-labeled substrate experiments to test for active respiration yielded tentative evidence for current-day microbial respiration in the ice wedge environment. Ice wedge samples showed slightly higher levels of radio-respiration than controls but were not statistically significant. The levels of radio-respiration in our ice wedge samples were extremely low and, taken alone, do not indicate active respiration over the measurement period. However, together with the trends in net CO2 flux observed in situ, which will be discussed in the following paragraph, the low levels of radio-respiration may be demonstrative of activity, albeit inconclusive. The radio-respiration experiment likely requires longer incubation times, and it is also possible that our methods were not adequately adapted for ice, as they were originally designed for sediments and soils (Steven et al., 2007). The amendment of radio-labeled acetate to melted ice prior to refreezing may have resulted in concentrations too dilute for readily detectable rates of respiration, or the acetate may have been frozen in the bulk ice and inaccessible to microorganisms that inhabited microstructures within the ice. An additional confounding factor may have been the loss of microstructures, or brine veins, known to be important for microbial life (Mader et al., 2006) during the brief thaw necessary to incorporate the radio-labeled acetate.
The use of CO2 flux as an index for microbial respiration is a common environmental method (Singh et al., 2011) and has recently been used on Antarctic Dry Valley soil (Ball et al., 2009). Ball et al. (2009) found that the relatively scant populations of plant and soil invertebrates in Antarctic soils simplified the correlation between CO2 flux and microbial activity, a result that is applicable to the high Arctic. Above the Axel Heiberg ice wedges, net production of CO2 above control levels occurred in late winter, which demonstrates that ice wedge communities continue to respire during subzero soil temperatures (between −9°C and −16°C). The detection of significantly higher CO2 flux in polygon troughs, underlain by ice wedges, compared to polygon centers is a strong indication of the effects of ice wedge formation on physical and microbiological parameters. The observed buildup of CO2 in the ice wedge, evident in the high rate of flux from above the ice wedge borehole, affirms the previous finding of biogenic CO2 gas released from ice wedges (Lacelle et al., 2011). The CO2 flux rates obtained from the surface fit within the range of previously reported values for tundra in summer, 960–2520 mg/m2/day (Lee et al., 2010), and are closer to dry, summer tundra soil flux, 958–2660 mg/m2/day, than wet conditions, 3180–3220 mg/m2/day (Jones et al., 1998). Previous studies of in situ CO2 flux from the surface of polygon troughs and centers report a negative flux: −3800 and −4300 mg/m2/day, respectively (Sommerkorn et al., 1999), highlighting the range of possible polygon types, some of which have protruding, drier centers, while others may exhibit a wetter environment produced by thermokarst development (see Fig. 1). Our data suggest that the low levels of respiration activity in ice wedges necessitate more sensitive methods of detection; however, the goal of using laboratory and in situ techniques to validate results has been illustrated here and provides evidence for the capacity of ice wedge habitats for supporting an active microbial community.
For life to adapt and persist in ice, the ice environment would have to be relatively stable over evolutionary timescales. On Earth, the Neoproterozoic Era (“Snowball Earth”) is believed to have played a significant role in the development of cold-adapted life (Vincent et al., 1999; Corsetti et al., 2006) over a period of ∼35 million years (Kirschvink et al., 2000). In the 700–900 million years following the formation of planet Mars, it is believed to have been a cold, wet planet. Remnants of ice-sculpted surface features indicate the past presence of large amounts of surface ice (Fairen et al., 2010). Under current atmospheric conditions on Mars, it is possible that remnants of ground ice could have persisted since the last major ice age 400,000 years ago (Bryson et al., 2008) and provided a plausible refugia for the development of ice-adapted life that may persist today.
Currently, there is no data on the physical conditions beneath martian polygons or the diversity of martian wedge morphotypes, which limits any specific comparison of their habitability to terrestrial wedges. In general, it is certain that any extant life in martian wedges would be subject to considerably harsher conditions than those on Earth, with more arid conditions, drastic daily temperature fluctuation, low pressure, and high levels of radiation (Gómez et al., 2010). Temperatures near the equator (Opportunity landing site) oscillate between −93°C and 17°C during the warmest time of year (Ulrich et al., 2010), and mean surface temperatures decrease significantly from the equator (−43°C) to the poles (−113°C) (Tillman et al., 1994). In comparison, the terrestrial ice wedge environment is less dynamic, with temperature fluctuations that occur on a seasonal basis and in situ temperatures that vary from −15°C to 0°C (Christiansen, 2005). Martian soils are expected to mediate temperature fluctuations with some surface features and regions possessing a high volumetric heat capacity and low albedo (Mellon et al., 2000); however, specific information regarding polygonal features and a temperature-depth profile is unavailable.
The Axel Heiberg ice wedges contained sizeable populations of culturable cells with characteristics of a cold-adapted, halotolerant community. Isolates were not capable of significant growth at subzero temperatures and were not tested at the levels of salinity (∼23% NaCl w/v) necessary for liquid brines to exist at martian temperatures (Ulrich et al., 2010). However, eight isolates did demonstrate halophilic characteristics and required 4% NaCl w/v or higher for growth. Modeling the surface temperatures and the expected salinity of common mineral ions on Mars, Ulrich et al. (2010) reported that the most extremophilic terrestrial microorganisms (metabolically active at −20°C) could be seasonally active in martian climes. While our predominantly psychrotolerant isolates would not be active at such low temperatures, similar psychrotolerant chemoorganoheterotrophs have demonstrated surprising hardiness under simulated martian conditions (Johnson et al., 2011), including a member of genus Arthrobacter that was abundant in our isolates. Furthermore, our selection of isolates does not likely represent the most extremophilic, cold-adapted organisms present in the ice wedge, since significant technical challenges exist for mimicking in vitro ice conditions. Yet we were able to culture one of the most abundant taxa, a Pseudomonas sp., which demonstrated high salt tolerance. Developing more advanced culturing methods for ice-based life should be a priority for astrobiologists, since an account of the true limits to terrestrial life is likely incomplete.
The major questions have yet to be answered regarding the ecological function or limitations of microbial life in ice and ice wedge environments, though this research provides the groundwork for what we might expect. The in situ respiratory activity of heterotrophic ice wedge communities will be very minimal due to both limited access to nutrients and low diffusion of gases through ice. The characterization of terrestrial ice wedges as habitable niches with a plausibly active microbial community has been strengthened by our results. This research also lays the groundwork for future studies at the field site on Axel Heiberg, which is considered a prime analogue for the climate during Mars' second age, when the planet transitioned from a warmer, wetter planet to an increasingly drier and colder one (Pollard et al., 2009; Fairen et al., 2010). Ice wedges are an enticing habitat for the study of terrestrial extremophiles and are likely to contribute insight for developing useful criteria for searching for life in ice-rich environments.
Footnotes
Acknowledgments
Logistical and financial support was provided by the Natural Sciences and Engineering Research Council (NSERC) Discovery Grant, Canada Research Chair Program, Canadian Foundation for Innovation (CFI), the Canadian Space Agency Canadian Analogue Research Network Program (CARN), the Department of Indian and Northern Affairs–Northern Scientific Training Program (NSTP), Polar Continental Shelf Project (PCSP), and McGill University's High Arctic Research Station. We would like to thank Dr. Helen Vrionis for assistance with the radio-labeled mineralization assay.
Abbreviations
CFU, colony-forming units; DAPI, 4,6-diamidino-2-phenylindole; gfp, green fluorescent protein-labeled; OTUs, operational taxonomic units; PCR, polymerase chain reaction.