
Abstract
Cryptoendolithic microbial communities and epilithic lichens have been considered as appropriate candidates for the scenario of lithopanspermia, which proposes a natural interplanetary exchange of organisms by means of rocks that have been impact ejected from their planet of origin. So far, the hardiness of these terrestrial organisms in the severe and hostile conditions of space has not been tested over extended periods of time. A first long-term (1.5 years) exposure experiment in space was performed with a variety of rock-colonizing eukaryotic organisms at the International Space Station on board the European EXPOSE-E facility. Organisms were selected that are especially adapted to cope with the environmental extremes of their natural habitats. It was found that some—but not all—of those most robust microbial communities from extremely hostile regions on Earth are also partially resistant to the even more hostile environment of outer space, including high vacuum, temperature fluctuation, the full spectrum of extraterrestrial solar electromagnetic radiation, and cosmic ionizing radiation. Although the reported experimental period of 1.5 years in space is not comparable with the time spans of thousands or millions of years believed to be required for lithopanspermia, our data provide first evidence of the differential hardiness of cryptoendolithic communities in space. Key Words: Astrobiology—Lithopanspermia—Radiation resistance—Survival—Vacuum. Astrobiology 12, 508–516.
1. Introduction
Shock recovery experiments performed to test phase-I of lithopanspermia showed that spores of Bacillus subtilis and the lichen Xanthoria elegans could survive pressures up to 40 GPa, which are comparable to those experienced by the martian meteorites (Stöffler et al., 2007; Horneck et al., 2008). Space technology provided the opportunity to study a variety of biological specimens after exposure to space. Among the systems tested, bacterial spores (B. subtilis) and the lichens Rhizocarpon geographicum and X. elegans stood out due to their high resistance to the hostile space environment (Sancho et al., 2007; de la Torre et al., 2010; Horneck et al., 2010). However, eukaryotes have never been studied after long-term exposures to space conditions.
To investigate the fate of lithic organisms and communities during long-term travel in space, we used ESA's EXPOSE-E facility (Rabbow et al., 2009, 2012). This facility was attached to the balcony of the Columbus module of the International Space Station (ISS) (Fig. 1A). EXPOSE-E was designed to expose a variety of biological systems to selected parameters of space over time spans of one year and more. All biological test systems of the LIFE (

Experiment hardware and biological samples of the LIFE experiment. (
2. Material and Methods
2.1. Experiment hardware and biological samples of the LIFE experiment on board the International Space Station
The EXPOSE-E facility is part of the European Technology Exposure Facility (EuTEF) (Fig. 1A), which was designed for testing different materials under selected parameters of space. On 7 February 2008, EuTEF with EXPOSE-E accommodating the biological samples of the LIFE experiment (Fig. 1B) was launched on board Space Shuttle STS-122 for the ISS. On 15 February 2008, EXPOSE-E was mounted onto the outside balcony of the Columbus module by extravehicular activity. EXPOSE-E was decommissioned on 1 September 2009, retrieved by extravehicular activity on 2 September 2009, and returned to Earth on 12 September 2009 with STS-128. During the 1.5 years mission, the samples were exposed to space vacuum (10−7 to 10−4 Pa) (Horneck et al., 2010), galactic cosmic radiation (≤190 mGy) (Berger et al., 2012), and the full spectrum of solar extraterrestrial electromagnetic radiation (λ>110 nm) with fluences of 9.19×105 J m−2 (below a 0.1% transmission neutral density filter) and 6.34×108 J m−2 (100% transmission insolated samples). All fluences were calculated for the biologically active UV range of 200 nm<λ>400 nm, depending on the orientation of the ISS to the Sun. Temperature varied between −21.5°C and +59.6°C (Rabbow et al., 2012).
2.2. Test systems of the LIFE experiment
The lichen Xanthoria elegans was collected from alpine habitats between 2000 and 3000 m altitudes at Zermatt, Switzerland (45°59′N, 7°48′E) (de Vera et al., 2003, 2008); Rhizocarpon geographicum was collected from the Plataforma de Gredos, Spain (40°17′N, 5°14′W) at an altitude of 2020 m (de la Torre et al., 2010); sandstone fragments colonized by a stratified cryptoendolithic microbial community were collected by L. Zucconi at Battleship Promontory (76°54′37.6″S, 160°55′27.5″E), Southern Victoria Land, Antarctica, in January 2004; and the microcolonial black yeast-like fungi Cryomyces antarcticus CCFEE 515 and Cryomyces minteri CCFEE 5187, both dwelling cryptoendolithically, were isolated from sandstone collected in McMurdo Antarctic Dry Valleys (Selbmann et al., 2005). The survival of those cryptoendolithic organisms is of special interest in terms of lithopanspermia because rocks may supply an additional external protection to face the impact-driven ejection into space (Horneck et al., 2008; Meyer et al., 2011) and transfer from one planet to another.
2.3. Viability assays of the LIFE test systems
The photosynthetic activity of the lichens X. elegans and R. geographicum was measured after reactivating the samples in a climatic chamber under the following controlled conditions: constant temperature of 10°C, 12 h light and 12 h dark cycles for 96 h (X. elegans) or 72 h (R. geographicum). Irradiation with photosynthetically active light was performed by using a mercury lamp with a 100 μmol m−2 s−1 photosynthetic photon flux density. For rehydration, samples were sprayed twice a day with deionized water. After reactivation, the activity of the photosystem II (PSII) of the photobiont was measured with a Mini-PAM fluorometer (Heinz Walz GmbH), as described previously (Sancho et al., 2007; de Vera et al., 2010). Lichens were rewetted immediately before each measurement. The optimum quantum yield of chlorophyll a was determined by fluorescence measurements after 20 min of dark adaptation, according to the equation
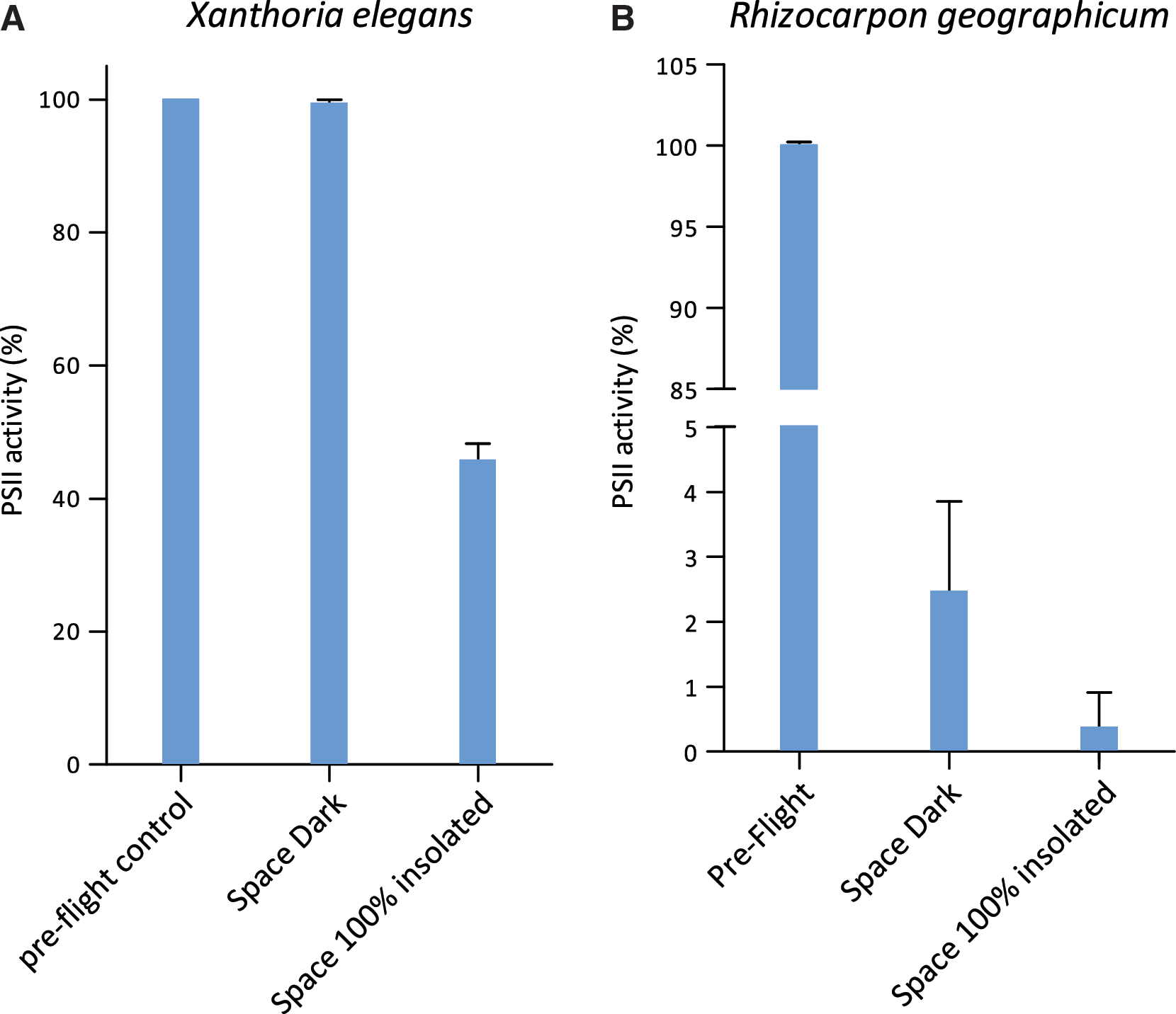
with Fv=variable fluorescence yield, Fm=maximal fluorescence yield, and Fo=minimal fluorescence yield. This optimum quantum yield of PSII was taken as an indication of the PSII activity of the photobiont of the lichen system after the exposure to space parameters. The percentage (n=2 for space 100% insolated and for space 0.1% insolated, and n=4 space dark samples) of PSII activity was determined from the ratio of the Fv/Fm of the same flight sample before and after flight. Control data are given for the same sample measured before spaceflight (preflight).
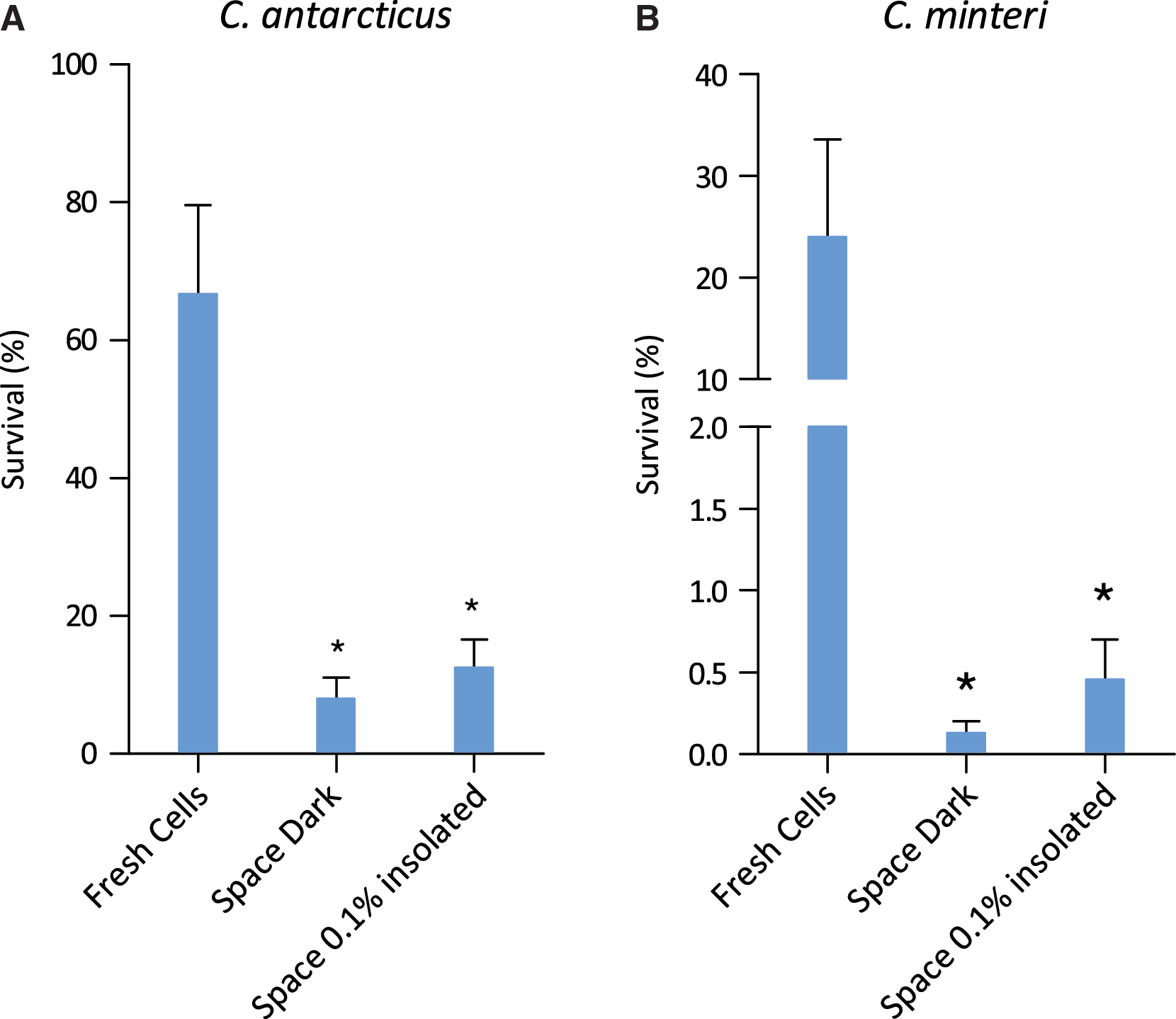
The survival of C. antarcticus and C. minteri was determined from their colony-forming ability as percentages of colony-forming units (CFU). Growth tests were performed by suspending fungal cells from rehydrated colonies in a 0.9% NaCl solution, inoculating them on malt agar Petri dishes (five replicates), and incubating them at 15°C for 30 days. Control data were obtained from fresh colonies. Statistical analyses were performed by one-way analysis of variance (ANOVA) and pairwise multiple comparison procedure (Tukey test), which was carried out by using the statistical software SigmaStat 2.0 (Jandel, USA). The means (n=5)±standard deviation (s.d.) are plotted. *P=0.001; power of performed test with α=0.050: 1.000.
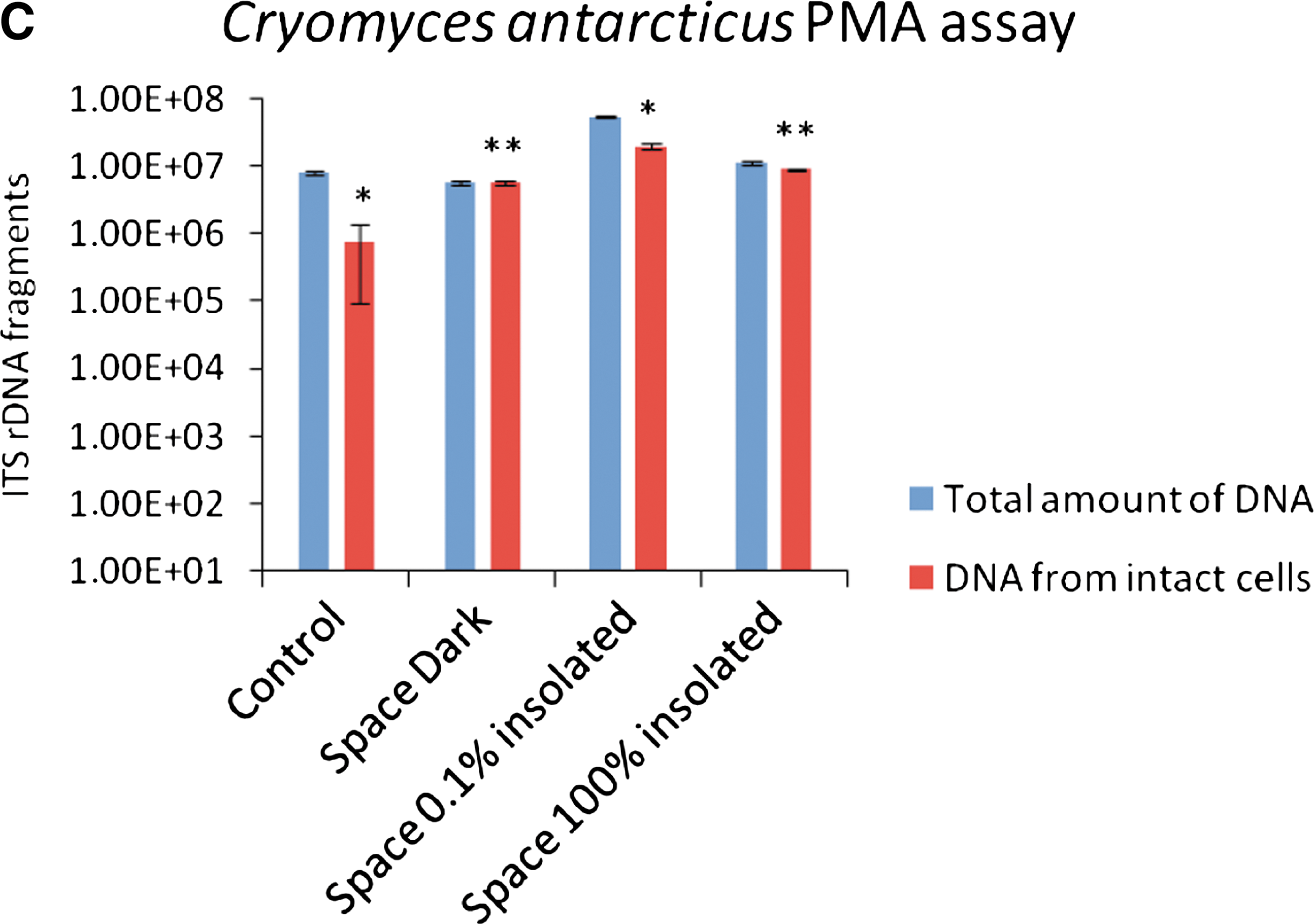
The propidium monoazide (PMA) assay was used to check the integrity of the cell membranes after spaceflight. Fractions of DNA extracted from intact cells of cryptoendolithic fungi C. antarcticus and C. minteri colonies were compared with the fraction of DNA extracted from cells isolated from the space-exposed colonized sandstone fragments. It was performed by adding PMA (Biotium, Hayward, CA) at a final concentration of 200 μM to the rehydrated fungal colonies or to powdered rock suspensions in phosphate-buffered saline solution. PMA penetrates only damaged cell membranes, cross-links then to DNA after light exposure, and thereby prevents polymerase chain reaction (PCR). Following DNA extraction and purification (Maxwell 16 automated DNA extraction instrument, Promega, Madison, WI), quantitative PCR (Bio-Rad CFX96 real time PCR detection system) was used to quantify the number of fungal internal transcribed spacer (ITS) ribosomal DNA fragments present in both PMA treated and non-treated samples. For all reactions, 1 μL of purified genomic DNA was added to 23 μL of PCR cocktail containing 1X Power Sybr-Green PCR Master Mix (Applied Bios, Foster City, CA), as well as NS91 forward (5′-gtc cct gcc ctt tgt aca cac-3′) and ITS51 reverse (5′-acc ttg tta cga ctt tta ctt cct c-3′) primers, each at 0.02 M final concentration. These primers amplify a 203 bp product spanning the 18S/ITS1 region of rRNA-encoding genes.
A standard quantitative PCR cycling protocol consisting of a hold at 95°C for 10 min, followed by 40 cycles of denaturing at 95°C for 15 s, annealing at 58°C for 20 s, and elongation at 72°C for 15 s, was performed. Fluorescence measurements were recorded at the end of each annealing step. At the conclusion of the 40th cycle, a melt curve analysis was performed by recording changes in fluorescence as a function of raising the temperature from 60°C to 95°C in 0.5°C per 5 s increments. These protocols were applied to the processing of both fungal colonies and cryptoendolithic sandstone samples.
Control data were obtained for an identical sample stored on ground in the laboratory during the mission (Control). Because cryptoendolithic communities are not uniformly distributed within the rocks, different quantities of total fungal DNA were obtained from different rock samples.
Space data are given for samples shielded from extraterrestrial UV radiation (Space Dark), exposed to λ>110 nm at a fluence of 9.19×105 J m−2 beneath a 0.1% transmission MgF2 filter (Space 0.1% insolated), or at a higher fluence of 6.34×108 J m−2 (Space 100% insolated). Statistical analyses were performed by one-way ANOVA and pairwise multiple comparison procedure (Tukey test), which was carried out by using the statistical software SigmaStat 2.0 (Jandel, USA) *P=0.001; **P>0.05. Power of performed test with α=0.050: 1.000. The means (n=3) and ± s.d. are plotted.
2.4. Confocal laser scanning microscopy (CLSM) imaging: viability of X. elegans analyzed by LIVE/DEAD staining kit FUN 1
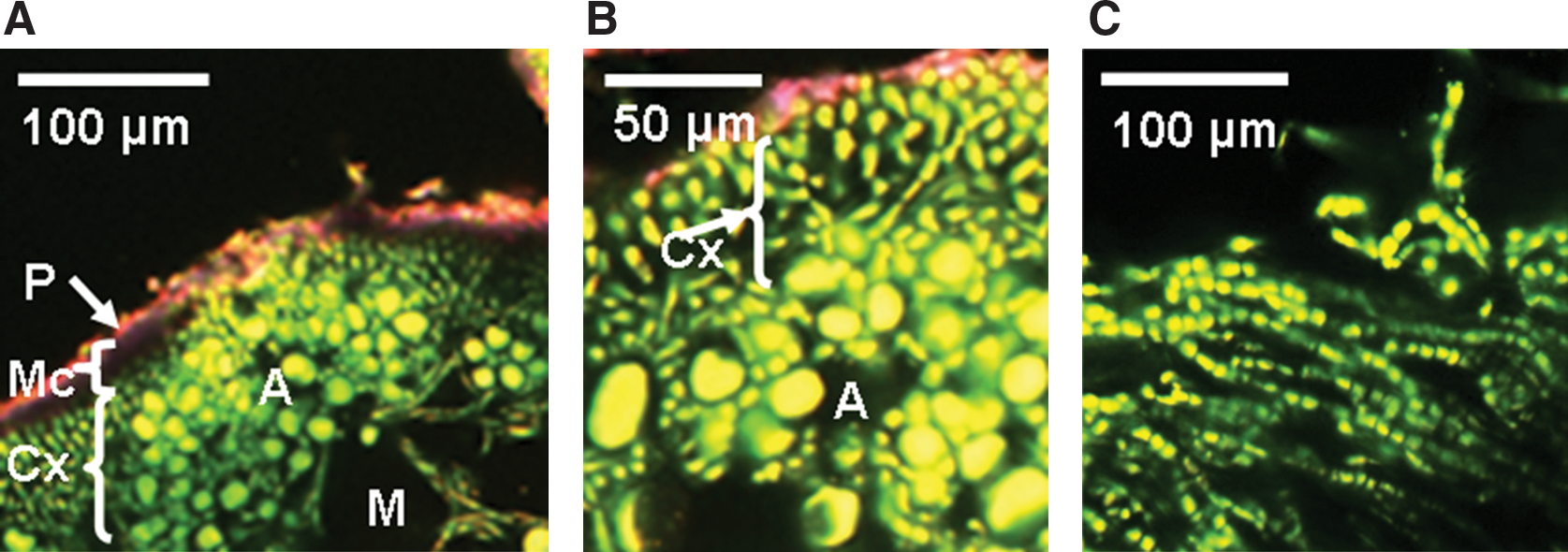
Adult lichen thallus, young thallus, and the isolated mycobiont of X. elegans were stained by FUN 1 to determine their viability. A green and yellow color of the cells indicates that they still maintain vitality. A change from green to yellow in the cytoplasm and from green to red in the vacuoles is an indication of physiological activity expressed by accumulation of the dye in the vacuoles. Dead cells cannot be stained; therefore, red crystals are not formed in the vacuoles.
Instruments used for imaging are as follows: LSM 510 META of Carl Zeiss Mikroskopsysteme Jena GmbH with objective lenses 10× and 40×/oil immersion, scanning resolution 1024×1024 pixels and time 64 μs to 1.76 μs, scan zoom 0.7×−2.8, image bit depth 12 bit, experimental temperature condition 22°C, UV-laser with emission wavelength 488/561/633 nm and VIS-laser HeNe with 633 nm excitation and 5 mW power, filters with ChS1 679–754; Ch2 BP 505–550; Ch3 BP 575–615. For operation of the instrument, the Carl Zeiss Jena GmbH software system at the Institute of Genetics of the Heinrich-Heine-Universität Düsseldorf was used.
3. Results
First visual inspections assured that the “space samples” had not changed in shape and color compared to their preflight appearances. Tailored to each test system, different viability assays were applied: (i) photosynthetic activity of the lichenized alga (photobiont) of X. elegans and R. geographicum (Fig. 2); (ii) colony-forming ability of C. antarcticus and C. minteri (Fig. 3A and 3B); (iii) fraction of DNA amplified from intact cells of C. antarcticus and C. minteri and of cryptoendolithic communities inside sandstone fragments (Fig. 3C, 3D, and 3E); and (iv) viability of X. elegans and the fungus of the lichen (mycobiont, cultured without the algal symbiont and dried) by means of vital staining (Fig. 4).
Viability of the lichens (
Viability of the fungi (

CLSM imaging: viability of X. elegans analyzed by LIVE/DEAD staining kit FUN I. (
Among those space samples that were shielded from extraterrestrial insolation but exposed to space vacuum, cosmic radiation, and temperature fluctuations (Space Dark in Figs. 2 and 3), the lichen X. elegans excelled by a PSII activity of 99.35±0.59% compared to the preflight data of the same samples (Fig. 2A). This high viability was not reached by the other space test systems kept in the dark during the mission: 2.46±1.39% for the PSII activity of the lichen R. geographicum (Fig. 2B), and 8.04±3.05% and 0.13±0.07% surviving cells for C. antarcticus and C. minteri, respectively (Fig. 3A and 3B). PMA assay showed 98±4.67% and 8.14±0.35% of DNA amplified from intact cells from colonies of C. antarcticus and C. minteri, respectively (Fig. 3C and 3D), in comparison to total extracted DNA; the percentage of DNA amplified from intact fungal cells in sandstone fragments was 18±0.18% (Fig. 3E).
The viability of culturable cells was not further decreased in the black fungi (12.5±4.11% for C. antarcticus and 0.46±0.24% for C. minteri) that were insolated with the full extraterrestrial spectrum (λ>110 nm) of the Sun, having received an UV irradiation of 9.19×105 J m−2 (Space 0.1% insolated, Fig. 3A and 3B). The highest fraction of intact fungal cells (35±0.15%) occurred in a sandstone sample that had received the full influx of solar electromagnetic radiation of 6.34×108 J m−2 (Space 100% insolated, Fig. 3E). When comparing all test systems exposed to outer space, including the high influx of full solar extraterrestrial radiation (6.34×108 J m−2), again the lichen X. elegans, with a PSII activity of 45±2.50% (Fig. 2A) and C. antarcticus with 80±0.82% of DNA from intact cells (Fig. 3C), were the most resistant test systems of LIFE. However, black Antarctic cryptoendolithic fungi lost colony-forming ability after exposure to full insolation; no survivors were detected in the space 100% insolated samples. Resistance of X. elegans and its mycobiont after exposure to the combined action of all space parameters tested, including full insolation, was confirmed by vital staining and CLSM. Figure 4 shows the high capacity of the lichenized fungus (part of the symbiotic lichen association) in a young thallus (B), and of the lichen fungus of X. elegans in pure culture (C), to resist space exposure. The fungal cells in the cortex (Cx) of the young thallus were still vital, although a protecting mucilage layer had not been formed, and the parietin layer appeared to be very thin. The same features appeared to (C), the fungal cells of the mycelium.
4. Discussion
The LIFE experiment provided for the first time data on the viability of rock-dwelling organisms and microbial communities after a long-term exposure to space parameters. These conditions cannot easily be simulated in the laboratory, if at all. The test systems collected from hostile conditions, such as Antarctica and high mountain regions, are adapted to cope with high radiation intensities, arid phases, and extreme temperature fluctuations that are in some ways similar to those experienced in space. In C. antarcticus, for example, globular cells are enveloped in a thick melanized cell wall, which protects them from radiation and desiccation. Their meristematic way of producing colonies (i.e., dividing in all directions) further supports their resistance (Onofri et al., 2008, 2009; Selbmann et al., 2005; Sterflinger, 2005). Special protection against environmental extremes is also granted for the photobiont, the green alga Trebouxia sp. of the lichen X. elegans. In this symbiotic organization, the fungus forms a cortex with an upper layer encrusted with parietin and a mucilage layer that envelopes the algal cells in a medulla matrix (Fig. 4A and 4B) (de Vera et al., 2003, 2008).
All organisms selected for the LIFE experiment are poikilohydric, that is, they are able to dehydrate till most biochemical activities stop. In this state, they are highly tolerant to stresses and may resume their metabolism once water becomes available again. Specifically, exposure to vacuum should inhibit any oxidative process related to their metabolism; this particular effect could protect against other damaging effects induced, for example, by solar UV radiation, cosmic ray ions, and temperature extremes.
The LIFE experiment has demonstrated that some, but not all, of those most robust microbial communities from extremely hostile regions on Earth are also partially resistant against the even more hostile environment of outer space. In this experiment, the following species stood out as the most persistent survivors after 1.5 years in outer space: the black fungus C. antarcticus (as determined from PMA assay) and the symbiotic X. elegans (as determined from PSII activity) and its mycobiont (as determined by LIVE/DEAD staining). However, the CFU test did not yield any survivors of C. antarcticus flight samples that were exposed to the un-attenuated solar extraterrestrial spectrum (space 100% insolated) and less than 10% survivors for the space dark samples. This means that even if the cell membrane seemed to be intact, as indicated by the PMA test, the cells had lost their ability to grow and divide.
Earlier studies have shown that the circumpolar and alpine red lichen X. elegans was able to retain its photosynthetic activity almost completely after 14 days in space (Sancho et al., 2007; de la Torre et al., 2010). This observation was confirmed in the present study for a much longer exposure time of 565 days in space, although this high viability was only observed for the lichen that had been shielded from solar electromagnetic irradiation, maintaining 45% PSII activity after 100% insolation exposure. CLSM, in combination with the use of specific fluorescent probes, allowed for the assessment of the physiological state of the cells. Particularly, the mycobiont seemed to play a fundamental role in maintaining the viability of the entire lichen system, because 565 days in space appeared not to have any effect on its physiological activity, even after 100% insolation (Fig. 4C). These observations are in agreement with earlier ones from short-term space exposures (de la Torre et al., 2010); in those studies the spores also maintained a high germination capacity, even after exposure to the full spectrum of solar UV radiation. However, so far, the ability of X. elegans to reproduce after this long-term space exposure remains an open question.
Interestingly, it has been earlier shown that X. elegans also resisted shock pressures comparable to those experienced by the martian meteorites during impact ejection (Horneck et al., 2008; Meyer et al., 2011), as required for phase-I of lithopanspermia.
Although we have demonstrated that some rock-dwelling species are capable of partially withstanding the harsh environment of outer space, or certain parameters of it, for at least 1.5 years, the data are insufficient for drawing any consequences for the likelihood of lithopanspermia. The possibility of surviving a much longer journey in space, as would be required for natural travel from Mars to Earth or vice versa, still remains an open question. This especially applies to organisms that dwell at the surface of rocks, like the lichen X. elegans, which would be fully exposed to the lethal spectrum of solar extraterrestrial UV radiation during a hypothetical interplanetary transfer. The only one data point at an exposure time of 1.5 years, resulting in a viability of 45±2.50%, as determined by PSII activity, does not allow any extrapolation over hundreds, thousands, or even millions of years, as would be required for lithopanspermia (Gladman et al., 1996). The situation would be much more favorable for endolithic organisms, because they would be shielded from solar UV radiation by the surrounding rock material. However, the cryptoendolithic organisms tested in this study did not show a high survival rate in space, even if shielded from solar UV radiation. Our experiments also demonstrate that outer space can act as a selection pressure on the composition of microbial communities (Cockell et al., 2011). Further studies are needed with more resistant communities. An example is the vagrant lichen species Aspicilia fruticulosa that, after 10 days in space, showed a complete recovery of space-induced damage after 72 h of reactivation (Raggio et al., 2011).
Natural transfer of microorganisms between planets via lithopanspermia could have occurred over the course of billions of years (Gladman et al., 1996; Nicholson, 2009). In the last 50 years, human activities in space exploration have become another potential source of spreading microorganisms between planets. Nearly 40 robotic missions have been launched with Mars as their destination (Horneck et al., 2007). To prevent the introduction of microbes from Earth to another celestial body or vice versa, a concept of contamination control has been elaborated by the Committee on Space Research (COSPAR) under consideration of specific classes of mission/target combinations, which have been recommended to be followed by each space-faring organization (COSPAR, 2011). Lander missions to Mars require especially strict measures of cleanliness and partial sterility of the spacecraft (COSPAR, 2011). The test systems that have been used in the LIFE experiment, though unlikely to be found on spacecraft, can be considered as models for demonstrating how life, if accidentally transferred from Earth to outer space, may resist and contaminate other celestial bodies and planets (i.e., Mars) and thereby interfere with future life-detection missions.
Footnotes
Acknowledgments
We thank the staff at the European Space Agency for the provision and operations of the EXPOSE-E facility and Thomas Berger for the cosmic ray dosimetry data. We also thank the Italian National Program of Antarctic Research and National Antarctic Museum for funding the collection of Antarctic samples, strains, and sample analyses, as well as the European Coordination Action for Research Activities on life in Extreme Environments (CAREX) for one ToK Grant to G.S. Collection and preparation of samples, and CLSM and photosynthetic activity analyses of X. elegans have been supported partly by the Helmholtz Association through the research alliance “Planetary Evolution and Life” and the BMWi, 50WB0614; the selection and preparation of the Rhizocarpon geographicum samples have been financed by the Space Program of the Spanish Ministry of Science and Education (ESP2005-25292-E) and INTA. We thank Philipp Holzwig, Isabella Halezki, Eva-Maria Posthoff, and the Institute of Genetics at the Heinrich-Heine-Universität Düsseldorf for skillful technical assistance. Special thanks also go to Prof. Giovanni Bignami for his scientific advice.
Author Disclosure Statement
No competing financial interests exist for Silvano Onofri, Rosa de la Torre, Jean-Pierre de Vera, Sieglinde Ott, Laura Zucconi, Laura Selbmann, Giuliano Scalzi, Kasthuri J. Venkateswaran, Elke Rabbow, Francisco Javier Sánchez Iñigo, or Gerda Horneck.
Abbreviations
ANOVA, analysis of variance; CFU, colony-forming units; CLSM, confocal laser scanning microscopy; COSPAR, Committee on Space Research; EuTEF, European Technology Exposure Facility; ISS, International Space Station; ITS, internal transcribed spacer; PCR, polymerase chain reaction; PMA, propidium monoazide; PSII, photosystem II; s.d., standard deviation.