
Abstract
As part of the PROTECT experiment of the EXPOSE-E mission on board the International Space Station (ISS), the mutagenic efficiency of space was studied in spores of Bacillus subtilis 168. After 1.5 years' exposure to selected parameters of outer space or simulated martian conditions, the rates of induced mutations to rifampicin resistance (RifR) and sporulation deficiency (Spo−) were quantified. In all flight samples, both mutations, RifR and Spo−, were induced and their rates increased by several orders of magnitude. Extraterrestrial solar UV radiation (>110 nm) as well as simulated martian UV radiation (>200 nm) led to the most pronounced increase (up to nearly 4 orders of magnitude); however, mutations were also induced in flight samples shielded from insolation, which were exposed to the same conditions except solar irradiation. Nucleotide sequencing located the RifR mutations in the rpoB gene encoding the β-subunit of RNA polymerase. Mutations isolated from flight and parallel mission ground reference (MGR) samples were exclusively localized to Cluster I. The 21 RifR mutations isolated from the flight experiment showed all a C to T transition and were all localized to one hotspot: H482Y. In mutants isolated from the MGR, the spectrum was wider with predicted amino acid changes at residues Q469K/L/R, H482D/P/R/Y, and S487L. The data show the unique mutagenic power of space and martian surface conditions as a consequence of DNA injuries induced by solar UV radiation and space vacuum or the low pressure of Mars. Key Words: Bacillus—Spores—Planetary protection—Space—Mars—Mutagenicity. Astrobiology 12, 457–468.
1. Introduction
In the framework of the EXPOSE-E project “Resistance of spacecraft isolates to outer space for planetary protection purposes (PROTECT),” spores of B. subtilis 168 were exposed in low-Earth orbit (LEO) to the conditions of outer space and simulated martian surface conditions. In previous spaceflight and ground-based space-simulation experiments, spores of this laboratory strain of B. subtilis were used as a model organism for astrobiological research to determine spore survival, quantify induced DNA damage, and probe the mutagenic effects of various space parameters (Horneck et al., 1984, 1994, 2001, 2002, 2008, 2010; Nicholson et al., 2000, 2002, 2005; Setlow, 2006). It has been shown that exposure of B. subtilis spores to simulated and real space conditions, such as ultrahigh shock pressure, space vacuum, extraterrestrial solar UV, and components of galactic cosmic radiation, leads to increased mutation frequencies in selected genetic markers, for example, amino acid auxotrophy (histidine revertants), sporulation ability (e.g., sporulation deficiency), and resistance to antibiotics (e.g., nalidixic acid or rifampicin or azide; Horneck and Buecker, 1983; Horneck et al., 1984, 1994, 2010; Munakata et al., 1997, 2002, 2004; Fajardo-Cavazos et al., 2005). These experiments have indicated that the mutagenic effect depends on (i) the physical parameter applied (such as the type of radiation, pressure, atmospheric composition, temperature); (ii) the biological variables (bacterial species, strain, genotype, etc.); and (iii) the endpoint of the investigation (such as the type of mutation, induction of enzymes related to mutagenesis; as reviewed in Horneck et al., 2010; Nicholson et al., 2000).
To gather further knowledge on the mutagenicity of outer space and simulated martian conditions, we applied two independent approaches. In the first approach, we specifically looked for mutations in one B. subtilis essential component of the major RNA polymerase, the rpoB gene, encoding the RNA polymerase β-subunit. Certain mutations in rpoB result in increased resistance to the antibiotic rifampicin (Rif) (Wehrli et al., 1968; Campbell et al., 2001). Rifampicin-resistant (RifR) mutations have been isolated from numerous bacterial species and mapped to four small areas within the rpoB gene, referred to as Clusters I, II, III, and N (reviewed in Campbell et al., 2001; Severinov et al., 1993). To date, the vast majority of the RifR mutations isolated in B. subtilis have been found to be single nucleotide substitutions resulting in specific amino acid changes within Cluster I of the rpoB gene (Nicholson and Maughan, 2002; Maughan et al., 2004; Perkins and Nicholson, 2008; Perkins et al., 2008; Moeller et al., 2010). Several lines of evidence indicate profound fundamental connections between the induced Rif resistance and effects on the general microbial physiology, for example, growth, competence, sporulation, and metabolism (Maughan et al., 2004; Perkins and Nicholson, 2008; Perkins et al., 2008). The rpoB gene has shown alterations in its gene sequence after exposure of spores to accelerated heavy ions (Moeller et al., 2010), vacuum (Munakata et al., 1997, 2002), simulated space conditions (Munakata et al., 2004, 2010), and simulated martian climate conditions (Perkins et al., 2008). To obtain more detailed insights into the mutagenicity of the various extraterrestrial conditions in this study, we looked for the mutagenic specificity in the nucleotide composition in the rpoB gene of B. subtilis resulting in amino acid changes in RpoB, one of the major enzymes that regulates the overall synthesis of RNA under the direction of DNA.
In the second approach, we analyzed exposed B. subtilis spores for changes in general physiological behavior, that is, the ability of spore formation (sporulation). The sporulation genes comprise approximately 12% of the whole genome of B. subtilis (Kunst et al., 1997; Maughan et al., 2007). Approximately 520 genes are (directly or indirectly) involved in spore formation, which indicates that deficiency in sporulation (i.e., a Spo− phenotype) can be due to several mutations (Kunst et al., 1997). The loss of sporulation is a dramatic change in the life cycle of B. subtilis cells that leads to cell death when nutrient starvation occurs and spore formation fails. It can be envisioned that in natural environments sporulation-deficient (Spo−) mutants would die and disappear from an environmental niche; thus under some conditions, sporulation deficiency can be considered a potentially lethal mutagenic event.
From the survival and mutagenicity data obtained, we assessed the likelihood of survival and genetic modifications of bacterial spores during an assumed robotic landing mission to Mars, which could have an impact on future planetary protection considerations.
2. Material and Methods
2.1. Bacillus subtilis sporulation and spore purification
Bacillus subtilis strain 168 (DSM 402) was obtained from the German Collection of Microorganism and Cell Cultures (DSMZ) GmbH, Braunschweig, Germany, and was used throughout. Spores were obtained by cultivation under vigorous aeration in double-strength liquid Schaeffer sporulation medium (SSM) (Schaeffer et al., 1965), and the spores were purified and stored as described (Nicholson and Setlow, 1990; Moeller et al., 2006). Spore preparations were free (>99%) of growing cells, germinated spores, and cell debris, as determined by phase-contrast microscopy.
2.2. Spore sample preparation
In the PROTECT experiment, aluminum coupons were chosen as carriers for the spore samples to simulate surface materials of a spore-contaminated spacecraft (Tauscher et al., 2006). The aluminum coupons (13 mm in diameter) were fashioned from spacecraft-qualified, chemfilm-treated aluminum 6061 (Titusville Tool & Engineering, Titusville, FL) and dry-heat sterilized prior to usage. Aliquots (20 μL) of a suspension of B. subtilis 168 spores containing (3.6±0.5)×108 colony-forming units (CFU) were deposited aseptically onto the surface of each coupon. Spores were allowed to settle, and coupons were air-dried for 1 day under ambient laboratory conditions (temperature 20±2°C and relative humidity 33±5%). The resulting spore layer was about 5–10 spores thick on average.
2.3. Spaceflight experiment
The PROTECT spore samples on aluminum coupons were stacked in triplicate inside the compartments of two trays of the EXPOSE-E facility on board the International Space Station (ISS), as described in Horneck et al. (2012 in this issue). In brief, tray 1 allowed access to space vacuum (10−7 to 10−4 Pa), cosmic radiation (up to 168 mGy depending on mass shielding), and—for the top layer of the sample stack—the full spectrum of solar extraterrestrial radiation at wavelengths >110 nm reaching after 1.360–1.860 h of insolation a total fluence of (5.5±0.6)×105 kJ/m2 for 100% transmission and 798±84 kJ/m2 for 0.1% transmission. Tray 2 provided simulated martian surface conditions, including atmospheric composition (1.6% argon, 0.15% oxygen, 2.7% nitrogen in carbon dioxide with a total pressure of 1000 Pa), cosmic radiation (up to 160±20 mGy), and—for the top layer of the sample stack—a simulated UV radiation climate of Mars at wavelengths >200 nm reaching a total fluence of (3.9±0.4)×105 kJ/m2 for 100% transmission and 525±44 kJ/m2 for 0.1% transmission. EXPOSE-E was installed on the outside of the ISS, and the samples were exposed to outer space and simulated martian conditions for 559 days (Rabbow et al., 2012 in this issue) before retrieval and analysis in the laboratory. Further details on the radiation dosimetry, applied UV fluences, doses of ionizing radiation, pressure values, temperature fluctuations, gas compositions, and so on are given in Tables 1 and 2 of Horneck et al. (2012), in Berger et al. (2012), and in Rabbow et al. (2012).
F, forward primer; R, reverse primer. Numbers denote the nucleotide coordinates at the end of the final PCR product. For example, the PCR product resulting from primer pair A would span from −24 to +737 of the rpoB coding sequence (the first nucleotide of the translation initiation codon is defined as +1).
Codon positions (CP) using B. subtilis 168 coordinates.
aa change=amino acid change.
One- and three-letter code for the amino acids: R arg (arginine), D asp (aspartic acid), Q gln (glutamine), H his (histidine), L leu (leucine), K lys (lysine), P pro (proline), S ser (serine), Y tyr (tyrosine).
Forty-seven RifR mutations were obtained and sequenced from B. subtilis 168 spores exposed to space and martian conditions in the frame of PROTECT (21 from the flight and 26 from the ground reference experiment) after cultivation (germination and outgrowth) of their vegetative cells.
Percentages of the induced RifR mutations were individually calculated for the flight (F) or the MGR experiment of PROTECT, respectively.
Tested space and martian conditions: space 100% UV=below 100% optical transmission (T) filter (>110 nm); space 0.1% UV=below 0.1% optical transmission (>110 nm); space vacuum=UV shielded; Mars 100% UV=100% T (>200 nm); Mars 0.1% UV=0.1% T (>200 nm); and Mars atmosphere=shielded from UV.
An identical set of samples was used as mission ground reference (MGR) for the EXPOSE-E flight experiment. In the MGR, the spore sample coupons were placed in two EXPOSE-E trays in an identical arrangement as in the flight experiment. For exposure to simulated space and martian surface conditions, the Planetary and Space Simulation facilities at the German Aerospace Center (DLR, Cologne, Germany) were used, as described by Rabbow et al. (2009, 2012). The test conditions mimicked the flight protocol (temperature profile, UV irradiation) to the best extent possible. In addition, a laboratory control was prepared in a similar manner as the flight samples and stored under laboratory conditions (20±2°C, 33±5% relative humidity) during the whole EXPOSE-E mission. All samples—flight samples, MGR, and laboratory controls—were analyzed simultaneously.
2.4. Survival assay
Spores were recovered from the aluminum coupons by the PVA recovery method (Horneck et al., 2001; Moeller et al., 2007). In brief, the spore layer was covered by a 10% aqueous polyvinyl alcohol solution (PVA), and after drying for 1 day under ambient laboratory conditions (20±2°C, 33±5% relative humidity), the spore-PVA layer was stripped off the aluminum coupons and resuspended in 1 mL distilled sterile water, which resulted in >95% recovery of the spores. To ensure optimal spore recovery, this process was repeated three times. Spores in 10-fold serial dilutions (in distilled sterile H2O) were spread on nutrient broth (NB) agar plates (Difco, Detroit, USA) and incubated overnight at 37°C; survival was determined from the resulting CFU. To control contamination, serial dilutions were plated on Spizizen minimal medium agar plates with 0.5% (w/v) glucose as the carbon source and supplemented with, and without, L-tryptophan (50 μg/mL). Note that strain B. subtilis 168 is a tryptophan auxotroph and requires 50 μg tryptophan per milliliter to grow on selective minimal medium.
2.5. Assay for mutations to rifampicin resistance
Spores in appropriate aqueous dilutions were plated on NB agar plates containing 50 μg rifampicin/mL (Merck KGaA, Darmstadt, Germany) as described previously (Nicholson and Maughan, 2002). The colonies that arose after overnight incubation at 37°C were counted as RifR mutants.
2.6. Sequencing and characterization of rpoB mutations
As mentioned previously, RifR mutations have been mapped to four small areas within the rpoB gene called Clusters I, II, III, and N (reviewed in Campbell et al., 2001; Severinov et al., 1993). Cluster N is located near the beginning of rpoB, and mutations consist of changes that alter the single amino acid (aa) V135 (Note: B. subtilis coordinates are used throughout). In the middle of the rpoB coding sequence are three closely linked regions: Cluster I spanning amino acids 463-GSSQLSQFMDQTNPLAELTHKRRLSALG-490; Cluster II spanning amino acids 519-TPEGPNIGLIN-529; and Cluster III consisting of the single amino acid R645. To amplify these regions of interest, we designed two pairs of primers (Table 1). Primer pair A amplified a 713–base pair (bp) region spanning the N cluster, and primer pair B amplified a 681 bp region spanning Clusters I, II, and III.
For nucleotide sequencing, RifR mutants were cultivated overnight in liquid Luria-Bertani medium (LB) containing 50 μg/mL rifampicin (LB+Rif), and a 1 mL sample was harvested by centrifugation in a microfuge. Chromosomal DNA suitable for PCR was isolated from RifR mutants by a rapid heating protocol as described previously (Nicholson and Maughan, 2002). Each PCR reaction was performed on four μL template DNA (0.15–0.2 μg DNA/μL), with each 1 μL (20 pmol) of the primer pairs, 5 μL H2O, and 10 μL of a 2×PCR Master Mix (Promega Corporation, Madison, WI). The regions of rpoB spanning all four RifR clusters were amplified with the primer pairs A or B (Table 1) in separate reactions. PCR reactions were performed in a PTC-200 thermal cycler (MJ Research, Waltham, MA) or in a peqStar thermocycler (peqlab, Biotechnologie GmbH, Erlangen, Germany), with a 2 min preincubation at 95°C, followed by 35 cycles of denaturation for 1 min at 95°C, annealing for 2 min at 55°C, and elongation for 3 min at 72°C. After a final incubation for 10 min at 72°C, the PCR products were purified with a Qiaquick PCR purification kit (QIAGEN), and the purified products were sequenced at the University of Florida Interdisciplinary Center for Biotechnology Research (ICBR, Gainesville, FL) or LGC Genomics GmbH (Berlin, Germany).
Mutations in rpoB were identified from multiple sequence alignments of the PCR products performed with use of the program ClustalW available online through the European Bioinformatics Institute (
2.7. Detection of sporulation deficiency
To determine mutations to sporulation deficiency (Spo−), spores in appropriate dilutions were plated onto SSM agar plates and incubated at 37°C for 7 days (Fajardo-Cavazos et al., 2005). This method leads to opaque, brownish-pigmented colonies of uniform morphology (Fajardo-Cavazos et al., 2005; Moeller et al., 2008), which is an indicator of spore formation (Hullo et al., 2001). A decrease in colony pigmentation and appearance of translucence are characteristic of Spo− mutants (Piggot and Coote, 1976; Fajardo-Cavazos et al., 2005). To verify the Spo− mutation, after growth for 3–4 days on NB agar plates, 10 Spo− colonies of each sample set (i.e., experimental condition) were individually transferred into 5 mL of fresh SSM medium and incubated for 24 h at 37°C. To determine the number of spore-forming units, each culture was plated before and after heat shocking (80°C for 10 min) as described in detail by Maughan et al. (2007).
2.8. Numerical and statistical analysis
The surviving fraction of B. subtilis spores was determined from the quotient N/N
0, with N=the number of CFU of the exposed sample and N
0 that of the nontreated (laboratory) controls. The frequency of induced RifR mutations was measured and calculated according to Baltschukat and Horneck (1991) with the following equation:
with CFU - RifR=colony formers grown on NB agar plates plus 50 μg rifampicin/mL - carrying induced RifR mutations (after exposure) or (control: spontaneous RifR mutations of laboratory control sample); total CFU: total colony formers (after exposure) grown on (NB) agar plates.
The frequency of induced Spo− mutations was determined by
with M=the total number of Spo− mutants of the exposed sample, N=the surviving spores of the exposed sample, and m S=the rate of spontaneous Spo− mutations. The residual sporulation efficiency of the induced asporogenous mutants was determined by dividing the titer after heat shock (spores) by the titer before heat shock (viable cells and spores).
For each experimental condition, that is, space 100% transmission (T) UV, space 0.1% T UV, space dark (i.e., UV shielded), Mars 100% T UV, Mars 0.1% T UV, and Mars dark, three stacks (with three sample layers in each stack) were analyzed. Data were obtained and calculated from the top, middle, and bottom layer of the following samples: space 100% T UV (sample ID: 83, 84, 88), space 0.1% T UV (sample ID: 91, 92, 96), space dark (sample ID: 67, 72, 75), Mars 100% T UV (sample ID: 211, 212, 216), Mars 0.1% UV (sample ID: 219, 220, 224), and Mars dark (sample ID: 195, 200, 203). Data shown are expressed as averages±standard deviations. The results were compared statistically via the Student t test and analyses of variance (ANOVA). Values were analyzed in multigroup pairwise combinations, and differences with P values ≤0.05 were considered statistically significant (Moeller et al., 2007, 2008, 2010).
3. Results
To study the mutagenicity of outer space and simulated martian conditions, air-dried multilayers of spores of B. subtilis 168 on board the EXPOSE-E platform were exposed for 1.5 years to extraterrestrial environmental parameters. In addition, an MGR experiment was performed at the Planetary and Space Simulation facilities at the DLR (Cologne, Germany). After retrieval, survival and mutations to RifR and Spo− were analyzed.
3.1. Spore survival
Approximately 60% of the B. subtilis 168 spores survived 559 days in outer space if totally shielded from solar electromagnetic radiation (Fig. 1B) This survival rate was decreased by up to 3 orders of magnitude if the spores were simultaneously exposed to the full spectrum of solar radiation. Attenuation of the solar flux by a factor of 1000 resulted in an increase in the surviving fraction by approximately a factor of 10 over those exposed to the full radiation spectrum. Likewise, under simulated martian conditions, about 70% of the spores survived 559 days exposure if kept in the dark with zero UV flux (Fig. 1C). Simultaneous exposure to simulated martian UV radiation climate (100% or 0.1% transmission) reduced the survival rate by a factor of about 10 or 3, respectively. The survival rates of irradiated spores of the MGR were for the most part higher than those obtained for the corresponding irradiated in-flight samples. However, it should be noted that under MGR conditions the polychromatic spectrum was cut off at 200 nm and did not extend to λ=110 nm as in the flight experiment. The laboratory control showed 100% survival, which indicates the long shelf life of B. subtilis 168 spores (Fig. 1A). The data clearly show the lethal effects of extraterrestrial solar UV radiation, whereas other environments, for example, space vacuum and galactic cosmic radiation, exerted only a minor lethal effect on the spores. To speculate on the long-term viability of terrestrial spores in the different space environments, the reasonable assumption could be made that spore inactivation is a first-order process (Nicholson, 2003); the survival rates (Fig. 1) can be converted into decimal reduction (D) values of ∼0.5 days (λ>110 nm, with 100% transmission),∼6 days (λ>110 nm, with 0.1% transmission),∼309 days (UV-shielded exposure to space vacuum, temperature shifts, and galactic cosmic radiation) in space, and ∼27 days (λ>200 nm, with 100% transmission),∼112 days (λ>200 nm, with 0.1% transmission), ∼413 days (with zero UV flux) on the martian surface. These observations are in good agreement with the results of previous flight and ground-based laboratory experiments (reviewed in Horneck et al., 2010). A genetic marker test proved that all colonies arose from the initial B. subtilis 168 strain and not from any contaminants.
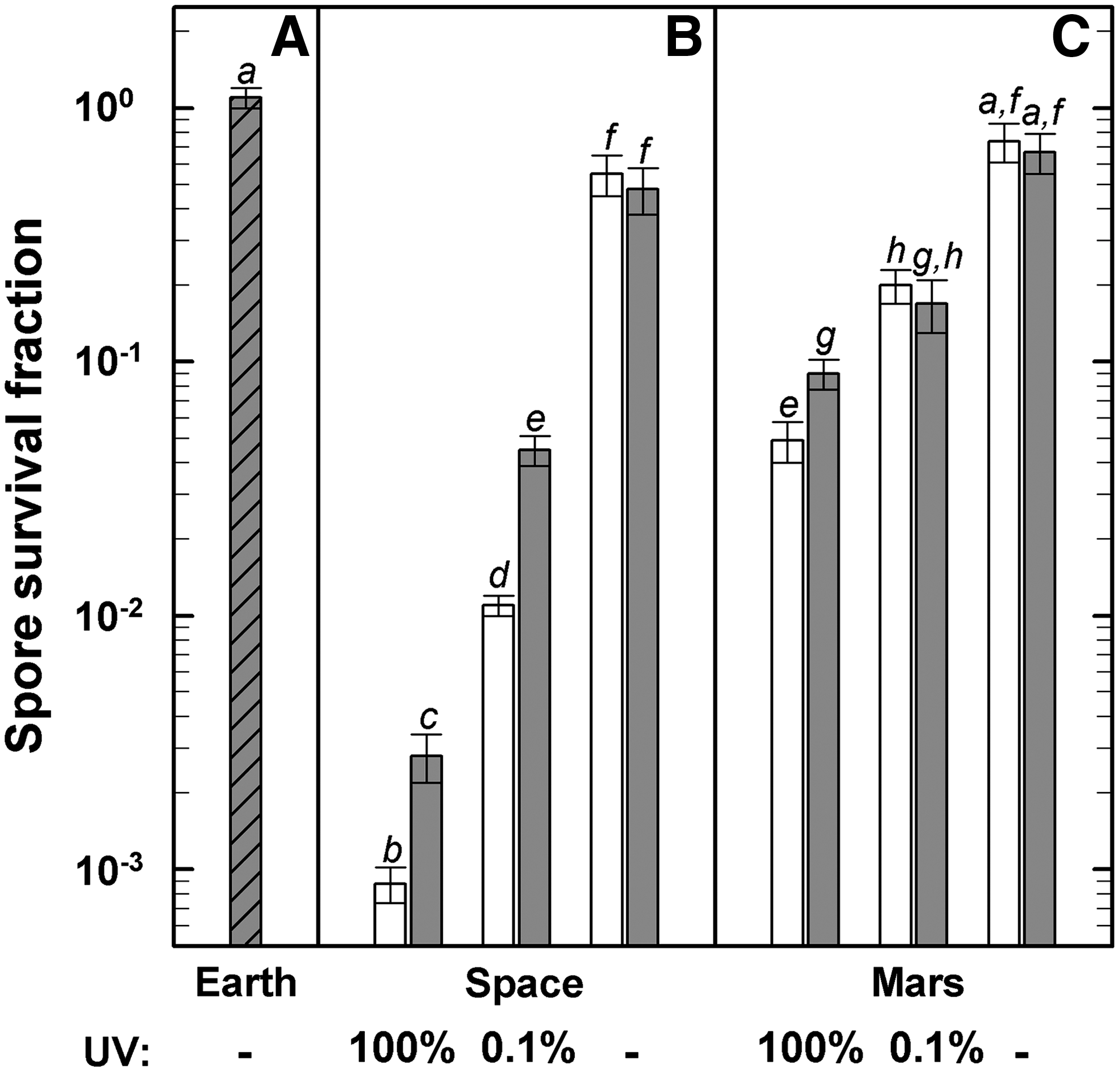
Survival rates of B. subtilis 168 spores in response to the exposure to laboratory control conditions (“Earth,”
3.2. Mutations to rifampicin resistance
The spontaneous mutation rate of RifR mutations, as determined from the laboratory control, was in the range of (1.2±0.3)×10−9 (Fig. 2A), which is in good agreement with data from Nicholson and Maughan (2002) for wild-type B. subtilis spores. This mutation rate was substantially increased in all space-exposed samples (Fig. 2B), that is, by more than a factor of 100 in the dark (UV-shielded) samples, and up to nearly 4 orders of magnitude in samples exposed to the full spectrum of solar extraterrestrial UV radiation. The corresponding “Mars samples” showed a similar increase in RifR mutations (Fig. 2C). These findings confirm previous studies on mutation induction in B. subtilis spores by simulated and real space parameters (reviewed by Horneck et al., 2010). Interestingly, the RifR mutation rates of all corresponding MGR samples were significantly lower than those of the flight samples (Fig. 2B and 2C).
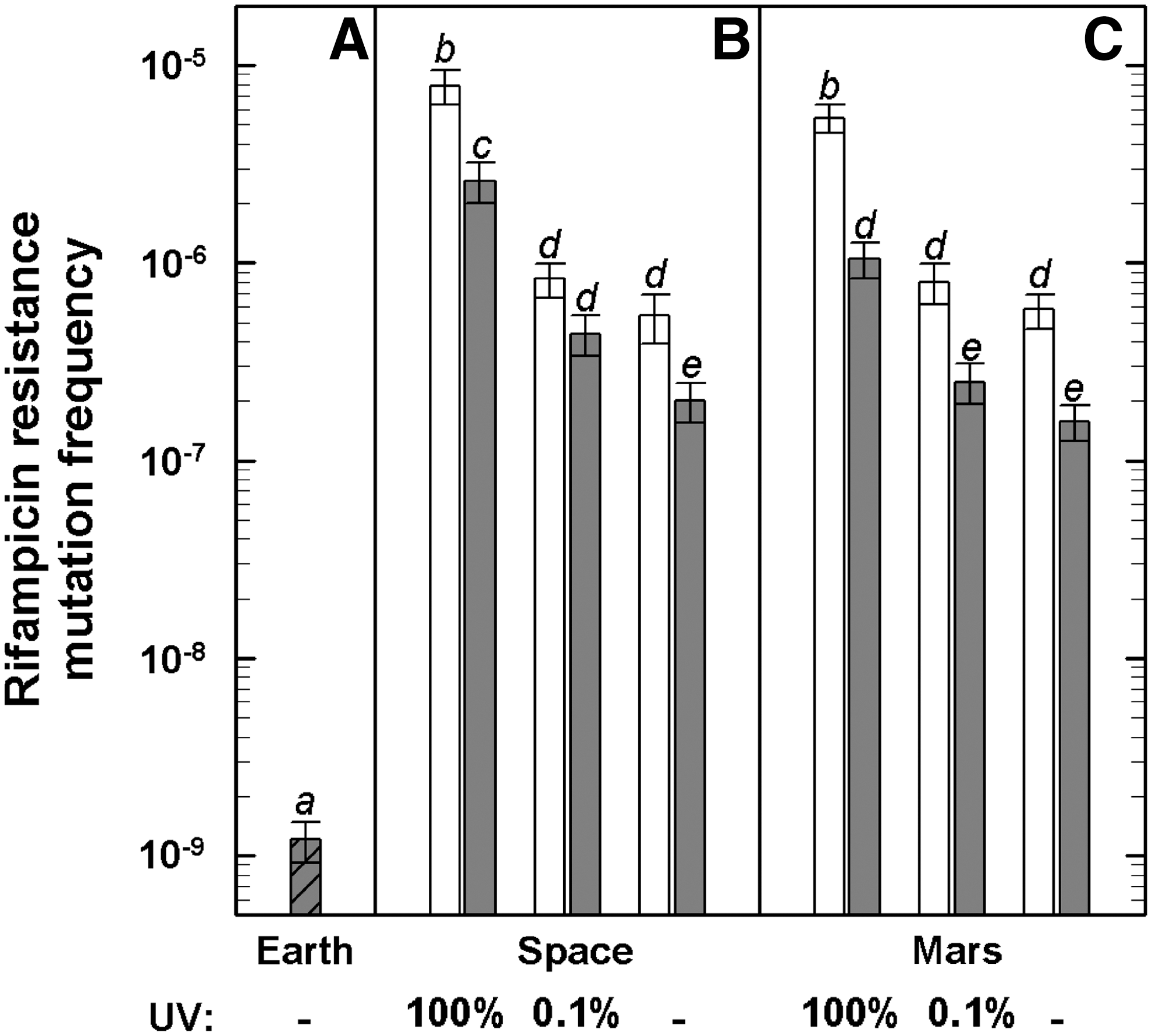
Mutation induction to rifampicin resistance in B. subtilis 168 spores exposed to laboratory control conditions (“Earth,”
3.3. Spectrum of RifR mutations in the rpoB gene of B. subtilis
DNA sequencing was performed on a total of 47 isolates, which consisted of 21 and 26 RifR mutants isolated from the flight and the MGR experiment, respectively. All mutations (47/47) were localized to Cluster I, in codons Q469 (10/47), H482 (35/47), and S487 (2/47) (Fig. 3). No mutations were found in the N cluster, Clusters II or III (data not shown). The codon H482 (CAC) was the definite hotspot for RifR mutations in the 21 isolated samples obtained from the flight experiment and accounted for 100% of the total, whereas the codons H482 and Q469 (CAA) were definite hotspots for RifR mutations in isolated samples of the MGR experiment; and each accounted for ∼54% and ∼38% of the total, respectively. The codon S487 (TCA) accounted for ∼8% of the total from MGR isolates (Fig. 3).
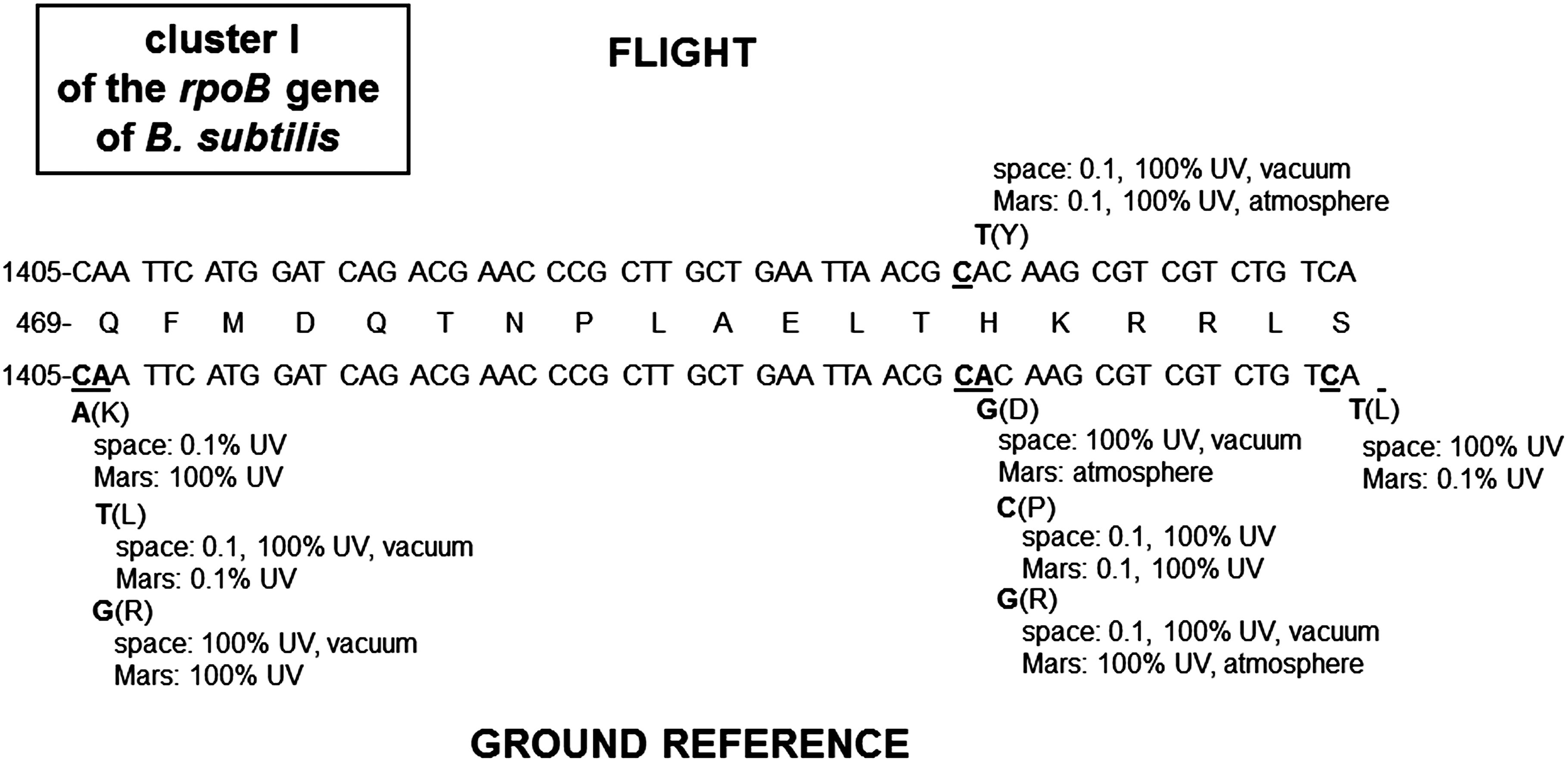
Mutations in Cluster I of B. subtilis 168 rpoB gene from spores exposed to space and simulated martian conditions in the flight (top) and MGR experiment (bottom) of the PROTECT experiment. The central line depicts the wild-type amino acid sequence in Cluster I, with the wild-type nucleotide sequence above and below. Nucleotides suffering changes are denoted by underlined letters. Nucleotide changes are indicated, and the resulting amino acid changes are in parentheses.
All 21 RifR mutations of the flight experiment were C-to-T transitions (Fig. 4A), whereas the majority of the RifR mutations of the MGR experiment were A-to-G transitions (∼35% of the total). The MGR RifR mutations showed further A-to-T transversions (∼19% of the total), C-to-G transversions (∼15% of the total), and A-to-C transversions (∼15% of the total) (Fig. 4B). Less than ∼8% of the total MGR RifR mutations were C-to-T transitions (Table 2).
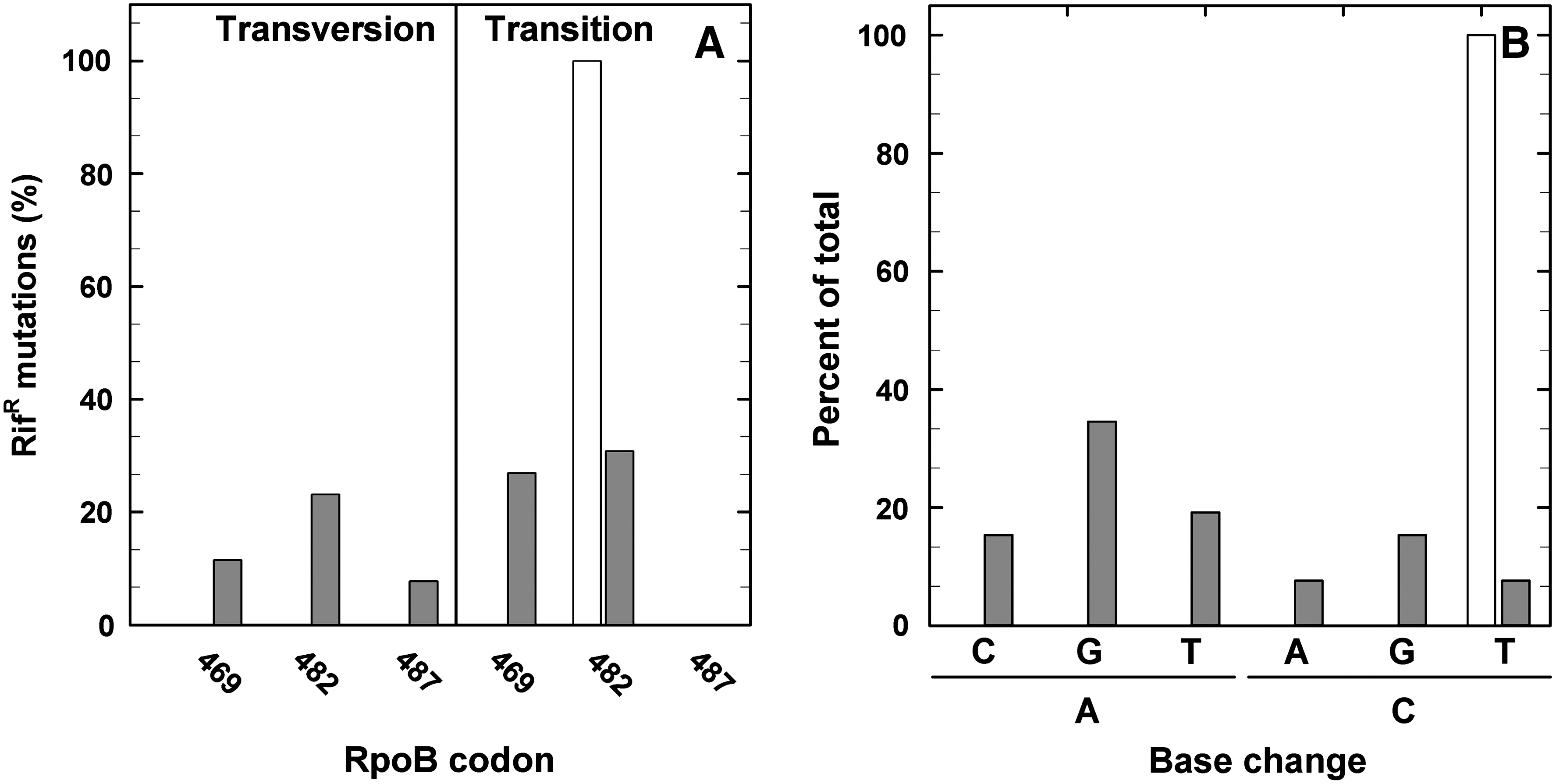
Frequency of real and simulated space- and martian-exposed B. subtilis 168 spores induced RifR mutations (transversion or transition mutation) of the flight (white bar) and MGR experiment (gray bars), determined in the sequenced part of Cluster I of the rpoB gene of B. subtilis 168 (
Interestingly, the C-to-T transitions obtained from the 21 isolated RifR mutations of the flight experiment were localized to only one hotspot within Cluster I. The single C1444T nucleotide substitution found in all 21 RifR mutants from the flight experiment resulted in the amino acid change H482Y, where the weak positively charged amino acid histidine is replaced by the aromatic amino acid tyrosine. In the MGR experiment, the most abundant mutations were A-to-G transitions in two codons: A1406G, resulting in the amino acid change Q469K, and A1445G, resulting in the amino acid change H482R (Fig. 3). The second most abundant mutations were A-to-T transversions in one codon: A1406T, resulting in the amino acid change Q469L (Fig. 4A). The third most abundant categories of mutations were C-to-G and A-to-C transversions (Fig. 4B). All observed eight amino acids suffering changes (Q469, H482, and S487) have been previously described in the B. subtilis rpoB gene (Rothstein et al., 1976; Boor et al., 1995; Ingham and Furneaux, 2000; Nicholson and Maughan, 2002; Perkins et al., 2008; Moeller et al., 2010).
3.4. Mutations to sporulation deficiency
After 7 days of incubation on SSM agar, colonies arising from the laboratory control samples (“Earth”) appeared homogeneous and indistinguishable from those of the wild-type strain (Fig. 5). In contrast, a fraction of the colonies arising from the flight and MGR samples showed distinctly different colony morphologies, including decreased pigmentation and more translucent appearance (Fig. 5); such morphology is characteristic of sporulation-defective (Spo−) B. subtilis mutants (Fajardo-Cavazos et al., 2005). Whereas the spontaneous Spo− mutation rate, as shown by the laboratory control, was 7×10−7 (Fig. 6A), examination of colonies from each flight and MGR condition revealed a high frequency of induced sporulation deficiency; the Spo− mutation rate increased by 1 to 2 orders of magnitude in the UV-shielded space or “Mars” samples (Fig. 6B, 6C) and by nearly 4 orders of magnitude in those space samples exposed to the full spectrum of solar UV radiation. In addition to the visual screening of Spo− mutants by their level of pigmentation, the sporulation frequencies of 10 overnight Spo− colonies from each experimental condition were determined after a heat shock treatment (80°C for 10 min). After transfer in sporulation medium and incubation for 24 h at 37°C, less than 0.01±0.004% of the descendants of Spo− mutants from space and “Mars” samples survived the heat shock treatment, which thereby confirms the stability of the Spo− mutation. Equally treated Spo+ samples of the laboratory control showed sporulation frequencies of 20.3±3.5% after similar overnight incubation.

Colony morphology of B. subtilis 168. Colonies arising on sporulation (SSM) medium: Earth (from laboratory control spores), space-exposed spores (“Space”), and spores exposed to simulated martian conditions (“Mars”). Arrowheads indicate sporulation-deficient (Spo−) mutants (white arrows) and sporulation-capable wild-type (Spo+) colonies (black arrows).
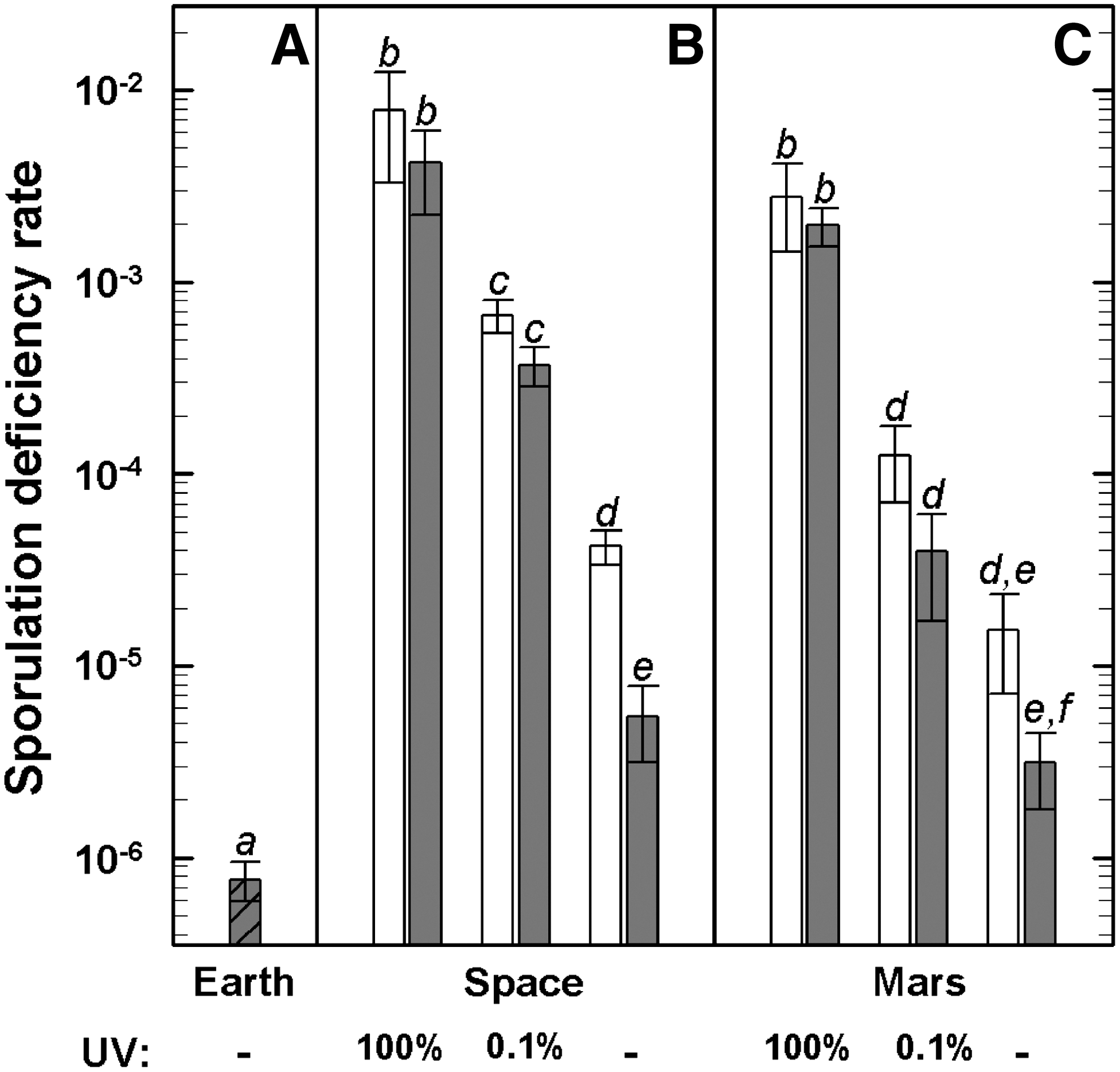
Sporulation deficiency induced in spores: laboratory control conditions (“Earth,”
4. Discussion
It was the aim of the PROTECT experiment of the EXPOSE-E mission to study the fate of bacterial endospores on a hypothetical robotic landing mission to Mars as a proxy of a putative microbial bioburden of the spacecraft. Our part in the PROTECT consortium was to look into the mutagenicity of the environment experienced during this journey.
The environment in LEO provided by the ISS is characterized by a high vacuum, an intense radiation environment of cosmic and solar origin, and extreme temperatures (Nicholson et al., 2000, 2005; Reitz, 2008; Horneck et al., 2010). These conditions were used to mimic the environment of the Earth-to-Mars trajectory. To test the mutagenicity of this space environment, we adopted two types of approaches: (i) a specific one targeted toward mutations in a single gene of B. subtilis, namely, the β-subunit of RNA polymerase–encoding gene rpoB, and (ii) a more unspecific approach of determining changes in a variety of genes that lead to loss of spore formation. Spores were either exposed to the full environment of LEO, including solar electromagnetic radiation (λ>110 nm), or shielded from insolation. Both types of mutations, RifR as mutation in the rpoB gene and Spo− indicating loss of sporulation capacity, were substantially induced in all flight samples. Compared to the laboratory control (“Earth” in Figs. 2 and 6), the mutation rates of the dark (shielded) flight spores increased by up to more than 2 orders of magnitude. Space vacuum or cosmic radiation, or both, are suggested to be responsible for this effect. These data confirm earlier mutation studies with spores of B. subtilis exposed to space vacuum (Horneck et al., 1984, 1994) or, in the laboratory, to selected aspects of this extraterrestrial environment, such as accelerated heavy ions as part of galactic cosmic radiation (Baltschukat et al., 1986; Baltschukat and Horneck, 1991; Moeller et al., 2010; Munakata et al., 2010), desiccation (Munakata et al., 1997), or high vacuum (Munakata et al., 2004, 2010). Cosmic particle radiation leads to mutations by damaging the DNA of the spores, mainly through single and double strand breaks and complex (multiple-base substitutions and deletions) damage (Horneck et al., 2006); however, the radiation doses measured for the PROTECT spores (Berger et al., 2012 in this issue) of approximately 160±20 mGy were too low to cause detectable mutations. Hence, desiccation and space vacuum may have been responsible for the increased mutation rates of the flight dark spores. The mechanism of these vacuum-induced mutations is not yet clear and needs further investigation. It may involve substantial conformational changes of the spore DNA, such as its reversible denaturation (Horneck et al., 2010). Additional exposure of the spores to the full spectrum of solar electromagnetic radiation at wavelengths >110 nm resulted in an increase in the mutation rates by almost 4 orders of magnitude (“Space 100%” in Figs. 2 and 6), compared to the laboratory control. This substantial increase in the mutation rates leads to the notion that both the spectrum and fluence of the applied UV radiation are mainly responsible for the highly induced mutagenic consequences.
The second setup of PROTECT dealt with the fate of bacterial endospores as a putative bioburden of a spacecraft landing at the surface of Mars. For this purpose, the spores were exposed for 1.5 years to a simulated martian environment, including atmospheric pressure and composition, cosmic radiation, and solar UV radiation climate (λ>200 nm). It is interesting to note that the extent of induced RifR mutations was comparable to the accordant “space” spores, the dark as well as the insolated ones (Fig. 2). This congruence in the mutation rates of the “space” and “Mars” samples suggests that—besides desiccation—the solar spectral part>200 nm, and not the vacuum UV radiation, is mainly responsible for the UV-induced mutations. This observation is backed up by the action spectrum for mutation induction that peaks around 260 nm, whereas that for inactivation as well as for single strand breaks in DNA shows an additional maximum at 150 nm (Munakata et al., 1991; Horneck and Rabbow, 2007).
We chose to study the mutagenicity of the rpoB gene because it has been shown that mutations in the rpoB gene have dramatic effects on general microbial physiology. It has been well established for the Gram-positive spore-forming bacterium B. subtilis that growth and metabolism, as well as several developmental events (e.g., extracellular enzyme synthesis, motility, chemotaxis, competence, sporulation, spore resistance properties, germination, and outgrowth), are controlled in a large part at the level of transcriptional activity (Maughan et al., 2004). RNA polymerase is a central macromolecular machine that controls the flow of information from genotype to phenotype, and insights into global transcriptional regulation can be gained by studying mutational perturbations in the enzyme (Boor et al., 1995; Campbell et al., 2001; Perkins and Nicholson, 2008). Mutations in the RNA polymerase β-subunit gene rpoB that cause RifR in B. subtilis were previously shown to lead to alterations in the expression of a number of global phenotypes known to be under transcriptional control, such as growth, competence for transformation, sporulation, germination, and nutrient utilization (Maughan et al., 2004; Perkins and Nicholson, 2008).
Exposure of B. subtilis spores to ultrahigh vacuum not only increased mutation frequency in spores but also altered the spectrum of mutations observed (Munakata et al., 1997, 2002, 2004, 2010). Recently, Munakata et al. (2010) reported different types of rpoB mutations arising from ionizing radiation and high-vacuum exposed spores of wild-type and DNA-repair deficient B. subtilis strains. Their spectrum of mutation types involved single-base substitutions, tandem double substitutions, non-tandem double substitutions, multiple-base substitutions, insertions, and deletions (Munakata et al., 2010). Interestingly, in all other publications dealing with mutations in the rpoB gene leading to RifR, including the present work, only single-base substitutions have been reported (Nicholson and Maughan, 2002; Maughan et al., 2004; Perkins et al., 2008; Moeller et al., 2010). It should be noted here that Munakata et al. (1997) isolated their RifR mutations by using a concentration of Rif of only 17 μg/mL, in contrast to the 50 μg Rif/mL used by others (Nicholson and Maughan, 2002; Maughan et al., 2004; Perkins et al., 2008; Moeller et al., 2010). Investigations into RifR mutations in the E. coli rpoB gene have previously demonstrated that selection for resistance to lower concentrations of Rif results in a broader range of rpoB mutations that often affect less-critical amino acids (Severinov et al., 1993).
In 2008, Perkins et al. (2008) exposed spores to Earth and simulated martian conditions, and as a result of their study they isolated mutations in three rpoB codons (Q469, H482, S487) (Perkins et al., 2008). Munakata et al. (2002, 2004) reported that the predominant rpoB mutation isolated from spores exposed to ultrahigh vacuum was S487F, a tandem CA-to-TT mutation. After exposure to heavy ions and X-rays in a previous study, we found alterations in the rpoB codons L467, Q469, A478, H482, R484, S487, A488, and H507 (Moeller et al., 2010). After exposure to real space and simulated martian conditions on board the ISS and in the Planetary and Space Simulation facilities at the DLR (Cologne, Germany) in the present study, we observed mutations in the following three rpoB codons: Q469, H482, and S487, all of which have been previously isolated after exposure to outer space and martian conditions, indicating that these mutations can be considered preferential hotspot regions in the rpoB gene (Munakata et al., 1997, 2002, 2004; Nicholson and Maughan, 2002; Maughan et al., 2004; Perkins and Nicholson, 2008; Perkins et al., 2008; Moeller et al., 2010).
Mutations in the rpoB gene have been previously reported to lead to significant changes in the global and specific microbial response to changes of the environmental conditions (Maughan et al., 2004; Perkins and Nicholson, 2008). The mutations isolated in this study, in the codons Q469, H482, and S487, have been reported to exhibit dramatic alterations in metabolism patterns in the utilization of various common and novel substrate classes, for example, sugars, carbohydrates, and amino acids (Perkins and Nicholson, 2008). In regard to the differences in the metabolism pattern of specific sugars, for example, D-mannitol, the H482Y mutation, which was the only allele isolated from the PROTECT flight experiment, exhibits dramatic reductions in mannitol utilization to below background level of the wild-type strain (Perkins and Nicholson, 2008). In regard to the other RpoB alleles isolated from the PROTECT MGR experiment, it was intriguing that replacement of S487 by the neutral aa leucine (L), of Q469 by the basic aa lysine (K) versus positively charged aa arginine R or the neutral aa L, or of H482 by the polar aa aspartic acid D versus nonpolar aa proline P or R resulted in such disparate effects (Perkins and Nicholson, 2008).
These, and several other observations, suggest the intriguing possibility that mutations induced in key targets, such as rpoB, during exposure to space radiation might endow the resulting mutants with enhanced fitness in new habitats (e.g., Mars). Such new phenotypes include changes in key microbial events, such as growth, competence for transformation, sporulation, and germination of B. subtilis. For example, mutations in the codons H482 and S487 showed slower growth, reduced (temperature-sensitive) sporulation efficiency (i.e., incubation at 37°C or 48°C in SSM or SSM+Rif), and weaker response to temperature shifts as well as an absence of competence for transformation (Maughan et al., 2004), which implies clearly that mutations in global regulators such as RpoB have selective advantages and/or disadvantages on the microbial propagation when encountering new habitable environments (Rainey, 1999; Cockell et al., 2000; Perfeito et al., 2007; Nicholson, 2009).
Under stressful environmental conditions, such as nutrient starvation, bacterial cells may produce spores that can stay dormant for extended periods (Nicholson et al., 2000). During sporulation, the cell invests a substantial amount of energy and time to form a spore that maximizes its survival potential (Piggot and Coote, 1976). To survive in stressful conditions, spores must function in one of two ways, that is, either (i) prevent cellular damage from occurring during dormancy or (ii) rapidly repair or replace damaged critical components during spore germination before inactivation results in cell death (Nicholson et al., 2000, 2005; Setlow, 2006). It should be noted that several features of spores, such as changed DNA conformation, low cellular water content, or spore-specific DNA repair enzymes, make spores ideal biological survival capsules. One of the most investigated types of mutations in B. subtilis is the loss of the individual sporulation capability, the so-called sporulation-defective (Spo−) mutation as illustrated and described in detail by Fajardo-Cavazos et al. (2005). While the detection of the Spo− phenotype by visual inspection of the colony morphology and pigmentation is quite simple, detailed information on the nature and type of mutations that cause Spo− is less available. The loss of sporulation is a dramatic change in the life cycle of B. subtilis cells; it should be kept in mind that, in B. subtilis, sporulation genes comprise almost 12% of the genome (Kunst et al., 1997; Maughan et al., 2007). Approximately 520 genes are (directly or indirectly) involved in the spore formation, which indicates that deficiency in the sporulation can be due to a variety of mutations (Kunst et al., 1997). Spo− mutants can be considered as a conditionally lethal mutation, as they can lead to cell death when nutrient starvation occurs and spore formation fails. To further verify the Spo− deficiency, we determined the response of Spo− and Spo+ spore survivors to a 80°C wet heat treatment and monitored a significantly lower survivability of the less pigmented Spo− compared to Spo+, indicating that the vast majority of the occurred Spo− mutants were at least blocked at stage IV of sporulation, after which spore heat resistance is developed (Nicholson and Setlow, 1990).
5. Conclusions
To adapt and survive changing environmental conditions, bacteria have global response systems that result in sweeping changes in gene expression and cellular metabolism. The results reported here clearly indicate that mutagenesis in general physiological behavior, that is, spore formation, as well as in specific marker genes like rpoB, shows a differential response to varying conditions in the spaceflight environment; thus these results have profound implications for the interplanetary transport of terrestrial microbes to Mars by impacts or spacecraft (reviewed in Nicholson et al., 2000, 2009). Further investigations on the impact of specific hotspot mutations such as RpoB's H482Y are ongoing with the intent to elucidate detailed insights into microbial physiology and competitive fitness regarding harsh terrestrial and extraterrestrial conditions.
(Other) Members of the PROTECT Team and Their Affiliations
Jean Cadet: Laboratoire “Lésions des Acides Nucléiques,” SCIB-UMR-E n°3 (CEA/UJF) Institut Nanosciences et Cryogénie CEA/Grenoble, Grenoble, France.
Thierry Douki: Laboratoire “Lésions des Acides Nucléiques,” SCIB-UMR-E n°3 (CEA/UJF) Institut Nanosciences et Cryogénie CEA/Grenoble, Grenoble, France.
Rocco L. Mancinelli: Bay Area Environmental Research Institute, NASA Ames Research Center, Moffett Field, California, USA.
Corinna Panitz: Institut für Flugmedizin, Technical University RWTH, Aachen, Germany.
Elke Rabbow: German Aerospace Center (DLR), Institute of Aerospace Medicine, Radiation Biology Department, Cologne, Germany.
Petra Rettberg: German Aerospace Center (DLR), Institute of Aerospace Medicine, Radiation Biology Department, Cologne, Germany.
Andrew Spry: Jet Propulsion Laboratory, Pasadena, California, USA.
Erko Stackebrandt: German Collection of Microorganisms and Cell Cultures GmbH (DSMZ), Braunschweig, Germany.
Parag Vaishampayan: Jet Propulsion Laboratory, Pasadena, California, USA.
Kasthuri J. Venkateswaran: Jet Propulsion Laboratory, Pasadena, California, USA.
Footnotes
Acknowledgments
The authors are very grateful to Andrea Schröder for her excellent technical assistance during the sample analyses. We express thanks to the two anonymous reviewers for insightful comments. R.M. was supported by the DLR grant DLR-FuE-Projekt ISS-Nutzung in der Biodiagnostik, Programm RF-FuW, Teilprogramm 475. W.L.N. was supported by grants from the NASA Planetary Protection Office (NNA06CB58G and NNX10AV22G).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
aa, amino acid; ANOVA, analyses of variance; bp, base pair; CFU, colony-forming units; COSPAR, Committee on Space Research; DLR, German Aerospace Center; ISS, International Space Station; LB, Luria-Bertani medium; LEO, low-Earth orbit; MGR, mission ground reference; NB, nutrient broth; RifR, rifampicin resistance; Spo−, sporulation deficiency; SSM, Schaeffer sporulation medium; T, transmission.