
Abstract
Because of their ubiquity and resistance to spacecraft decontamination, bacterial spores are considered likely potential forward contaminants on robotic missions to Mars. Thus, it is important to understand their global responses to long-term exposure to space or martian environments. As part of the PROTECT experiment, spores of B. subtilis 168 were exposed to real space conditions and to simulated martian conditions for 559 days in low-Earth orbit mounted on the EXPOSE-E exposure platform outside the European Columbus module on the International Space Station. Upon return, spores were germinated, total RNA extracted, fluorescently labeled, and used to probe a custom Bacillus subtilis microarray to identify genes preferentially activated or repressed relative to ground control spores. Increased transcript levels were detected for a number of stress-related regulons responding to DNA damage (SOS response, SPβ prophage induction), protein damage (CtsR/Clp system), oxidative stress (PerR regulon), and cell envelope stress (SigV regulon). Spores exposed to space demonstrated a much broader and more severe stress response than spores exposed to simulated martian conditions. The results are discussed in the context of planetary protection for a hypothetical journey of potential forward contaminant spores from Earth to Mars and their subsequent residence on Mars. Key Words: Bacillus—Mars—Planetary protection—Spaceflight—Spores. Astrobiology 12, 469–486.
1. Introduction
The well-characterized spore-forming bacterium Bacillus subtilis has long been used as a model organism for astrobiological studies (reviewed in Nicholson et al., 2000, 2005, 2009; Horneck et al., 2002, 2010) and as a representative spacecraft contaminant in planetary protection studies (Nicholson et al., 2000, 2009; Fajardo-Cavazos et al., 2007). Most historical studies in which spores have been exposed to the space environment have concentrated mainly on measuring spore survival and, to a lesser extent, mutation frequency of selected markers (Horneck et al., 1984, 1994). In the PROTECT experiments presented in this volume, we attempted for the first time to elucidate some of the molecular responses of spores exposed to the conditions of outer space and simulated martian surface conditions. In addition to evaluating the spectrum of mutations induced in the rpoB gene (Moeller et al., 2012), we attempted to capture a broad range of cellular damages caused to spores by space or simulated Mars exposure, using the following rationale. It is known that spores are dormant and metabolically inactive and thus cannot repair cellular damage suffered during dormancy (Nicholson et al., 2000, 2005). Exposure to space or simulated martian conditions would therefore be expected to cause cumulative damage to cellular components. We reasoned that, upon germination, spores would detect such damages and activate the transcription of specific genes responsible for repair of each type of damage. The identification of such repair genes would thus indicate the particular type of damage inflicted on the spore by space or Mars exposure, or both. This data can enhance our understanding of how terrestrial microorganisms cope with transfer through space to an environment such as Mars and possibly become viable forward contaminants. In this communication, we present the results of these experiments.
2. Materials and Methods
2.1. Bacillus subtilis sporulation and spore purification
The laboratory strain of B. subtilis, strain 168 (trpC2) (Zeigler et al., 2008), was from the corresponding author's strain collection and was used throughout these studies. Spores were obtained by cultivation at 37°C for 5–7 days on solid agar plates of Schaeffer sporulation medium (Schaeffer et al., 1965) and were purified and stored in distilled water as described previously (Nicholson and Setlow, 1990). Spore preparations were free (>99%) of growing cells, germinated spores, and cell debris as determined by phase-contrast microscopy.
2.2. Spore sample preparation
Spacecraft-qualified, chemfilm-treated aluminum 6061 coupons (13 mm in diameter×1 mm thick; Titusville Tool & Engineering, Titusville, FL, USA) were dry-heat sterilized (132°C, 16 h) prior to use. Aliquots (20 μL) of a suspension of B. subtilis 168 spores containing ∼1×108 colony-forming units were deposited aseptically onto the surface of each coupon. Spores were allowed to settle, and coupons were air-dried for 1 day under ambient laboratory conditions (temperature 20°C±2°C and relative humidity 33±5%), which resulted in layers ∼5–10 spores thick.
2.3. Spaceflight experiment
Spore samples on aluminum coupons were stacked in triplicate inside the compartments of two trays of the EXPOSE-E facility. EXPOSE-E was installed on the outside of the International Space Station (ISS), and the samples were exposed to outer space and simulated martian conditions for 559 days before being retrieved and analyzed in the laboratory. Spores in the two trays were exposed to the following parameters: (i) space vacuum (∼10−4 Pa), temperature (−20°C to +59°C), and galactic cosmic radiation (155 mGy); (ii) simulated martian atmospheric composition (1.6% argon, 0.15% oxygen, 2.7% nitrogen in CO2), martian surface pressure (103 Pa), temperature (−20°C to +59°C), and galactic cosmic radiation (134–142 mGy). Further details on the radiation dosimetry, applied UV fluences, doses of ionizing radiation, pressure values, temperature fluctuations, gas compositions, and so on are given by Horneck et al. (2012) and Rabbow et al. (2012). Only spores shielded from solar radiation (i.e., the middle and bottom coupons in triplicate stacks) were analyzed in this study. Laboratory ground controls consisted of an identical set of spore-loaded coupons stored at Earth-ambient conditions shielded from light for the duration of the experiment. Exposed spores and ground control samples were processed simultaneously.
2.4. Spore recovery from aluminum coupons
Spores were recovered from the aluminum coupons by stripping with polyvinyl alcohol as described previously (Lindberg and Horneck, 1991; Moeller et al., 2012). Spores were washed by repeated centrifugation in sterile distilled water and stored in water at 4°C. For enumeration, spores were diluted serially 10-fold in phosphate-buffered saline (PBS) (Nicholson and Setlow, 1990). Total spores recovered were enumerated by direct counts in a Petroff-Hauser cell counting chamber (Gerhardt et al., 1981), and viable spores were enumerated by spreading of appropriate dilutions on Luria-Bertani (LB) agar plates (Miller, 1972) and counting colonies after overnight incubation at 37°C.
2.5. Spore germination and total RNA isolation
To synchronize germination, spores in water (∼1×109) were heat activated at 70°C for 30 min and then harvested by centrifugation. To initiate germination, spores were resuspended in 100 mL of 2× LB medium containing the germinant L-alanine (10 mM final concentration) in a 250 mL Klett flask and agitated vigorously in a rotary water bath shaker at 37°C. Germination was monitored by loss of optical density as measured in a Klett-Summerson photometer fitted with the No. 66 (660 nm; red) filter. After 60 min of germination, spores were harvested by centrifugation, and the pellets stored frozen at −20°C until further analysis. Total RNA was isolated from germinated spores by using the Ribo-Pure Bacteria Kit followed by treatment with RNase-free DNase, using the protocols supplied by the manufacturer (Ambion-Life Technologies, Grand Island, NY, USA).
2.6. Microarray analyses
Total RNA samples (20 μg) were sent to the University of Florida Interdisciplinary Center for Biotechnology Research (UF-ICBR). Samples were converted into Cy3- or Cy5-labeled cDNAs, and identical quantities of cDNAs were used to probe 15,000-feature custom microarrays (Agilent) prepared from the B. subtilis strain 168 genomic sequence (Kunst et al., 1997). Background subtraction, quantile, and Loess normalization were performed by the UF-ICBR. The microarray contained 15,209 data points equivalent to 4,103 genes with an average number of 3.7 measurements per gene; the oligos were designed such that each open reading frame (ORF) was sampled at multiple locations (n=3 or 4). Log-transformed data sets for each ORF from space- or simulated Mars-exposed spores were compared to the same ORF from the ground control spores by using one-way analysis of variance (ANOVA). Data showing ≥3-fold differences with P values of ≤0.05 were considered statistically significant.
2.7. Microarray data accession number
Complete microarray data have been deposited in Gene Expression Omnibus under accession number GSE37124 (
3. Results
3.1. Spore survival and germination
From the total viable versus direct spore counts, it was determined that 6.1% and 97.6% of B. subtilis 168 spores survived 559 days in outer space or under simulated martian conditions, respectively, if shielded from solar UV radiation. Note that in our samples, space-exposed spores exhibited an apparently lower survival value (6.1%) than in the samples reported by Horneck et al. (2012) (∼55%). This discrepancy is likely due to the different techniques used to assess spore viability in our two experiments (see Horneck et al., 2012 in this issue for comparison). However, these observations are in good general agreement with the results of previous flight and ground-based laboratory experiments (reviewed in Horneck et al., 2010). Making the reasonable assumption that spore inactivation is a first-order process (Nicholson, 2003), these survival rates can be converted into decimal reduction (D) values of ∼458 days (1.25 years) in space and ∼53,000 days (∼145 years) on the martian surface.
Spores were germinated in 2× LB liquid medium containing the potent germinant L-alanine (Fig. 1). It was noted that spores stored at ambient Earth conditions germinated normally, losing ∼50% of their initial optical density within the first 30 min, whereas spores exposed to space or simulated martian conditions aboard PROTECT demonstrated only ∼30% loss in optical density over the same time period (Fig. 1).
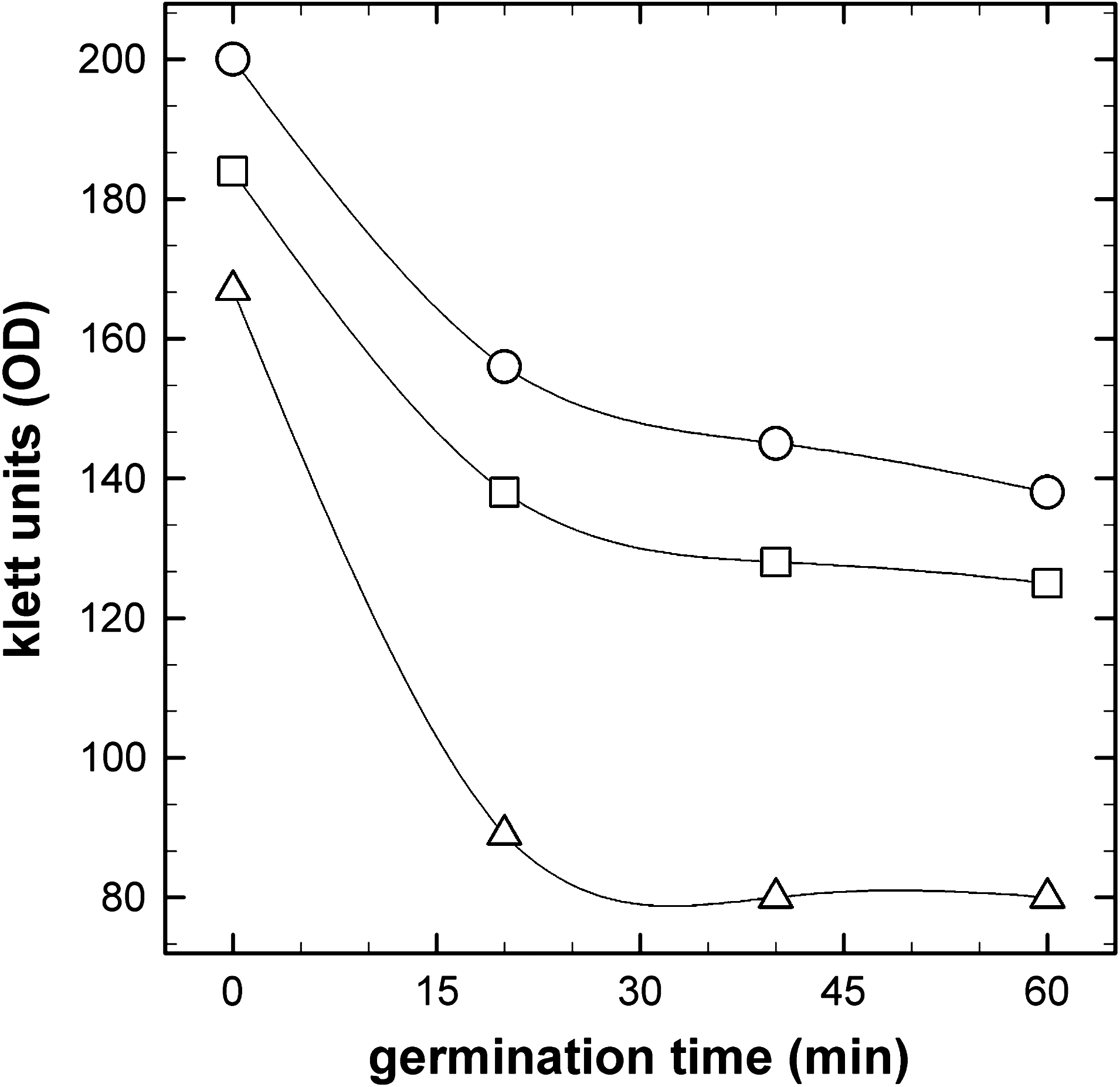
Germination of B. subtilis spores exposed to space (open squares), simulated martian conditions (open circles), or Earth conditions (open triangles) for 559 days. OD, optical density. See text for details.
3.2. Microarray data: overall trends
To visualize overall differences in the germination transcriptome of B. subtilis spores exposed to space or simulated martian conditions, microarray data were subjected to global comparisons and displayed as scatter plots (Fig. 2). As a measure of reproducibility, the complete sets of germination microarray data were compared from the two Earth control samples used in the experiment (Fig. 2A), and the two data sets demonstrated a high correlation coefficient (r=0.9781), which indicates that the methods used were inherently reproducible. When the complete sets of microarray data from space-exposed spores (Fig. 2B) or simulated Mars-exposed spores (Fig. 2C) were compared to their respective Earth controls, considerable divergence was observed (correlation coefficients of r=0.8967 and r=0.8445, respectively), which indicated that exposure to space or simulated martian conditions had resulted in alterations in the global B. subtilis germination transcriptome.
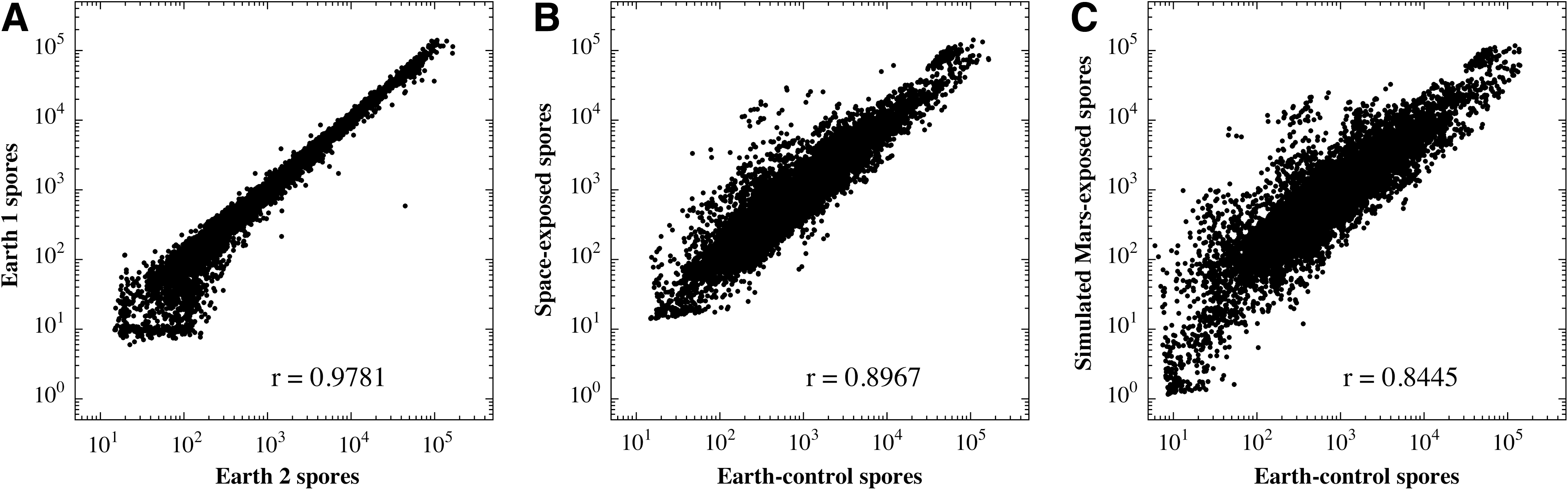
Scatter plot summaries of microarray data. Germination microarray comparisons of B. subtilis 168 spores exposed to the following conditions. (
Analysis of microarray data from space- and simulated Mars-exposed spores identified genes which were significantly up- or down-regulated relative to the ground control spores. Visual inspection of the summarized data (Fig. 3) yielded the following observations. Spores exposed to full space conditions exhibited more than twice as many genes with altered expression levels (236) than did spores exposed to simulated martian conditions (112) (Fig. 3). It is particularly notable that simulated Mars exposure led to significant down-regulation of only 20 genes (Fig. 3). Furthermore, it was observed that in all spores exposed to either space or simulated martian conditions, greater than 50% of genes with altered expression belonged to Category 5, genes of unknown function (Fig. 3). The subsets of B. subtilis genes found to be significantly up-regulated or down-regulated during germination of space-exposed and simulated Mars-exposed spores are listed and compared in Tables 1 and 3, respectively. According to our hypothesis, searching through these genes should offer clues to the type(s) of cellular damage suffered by spores exposed to space or simulated martian conditions.
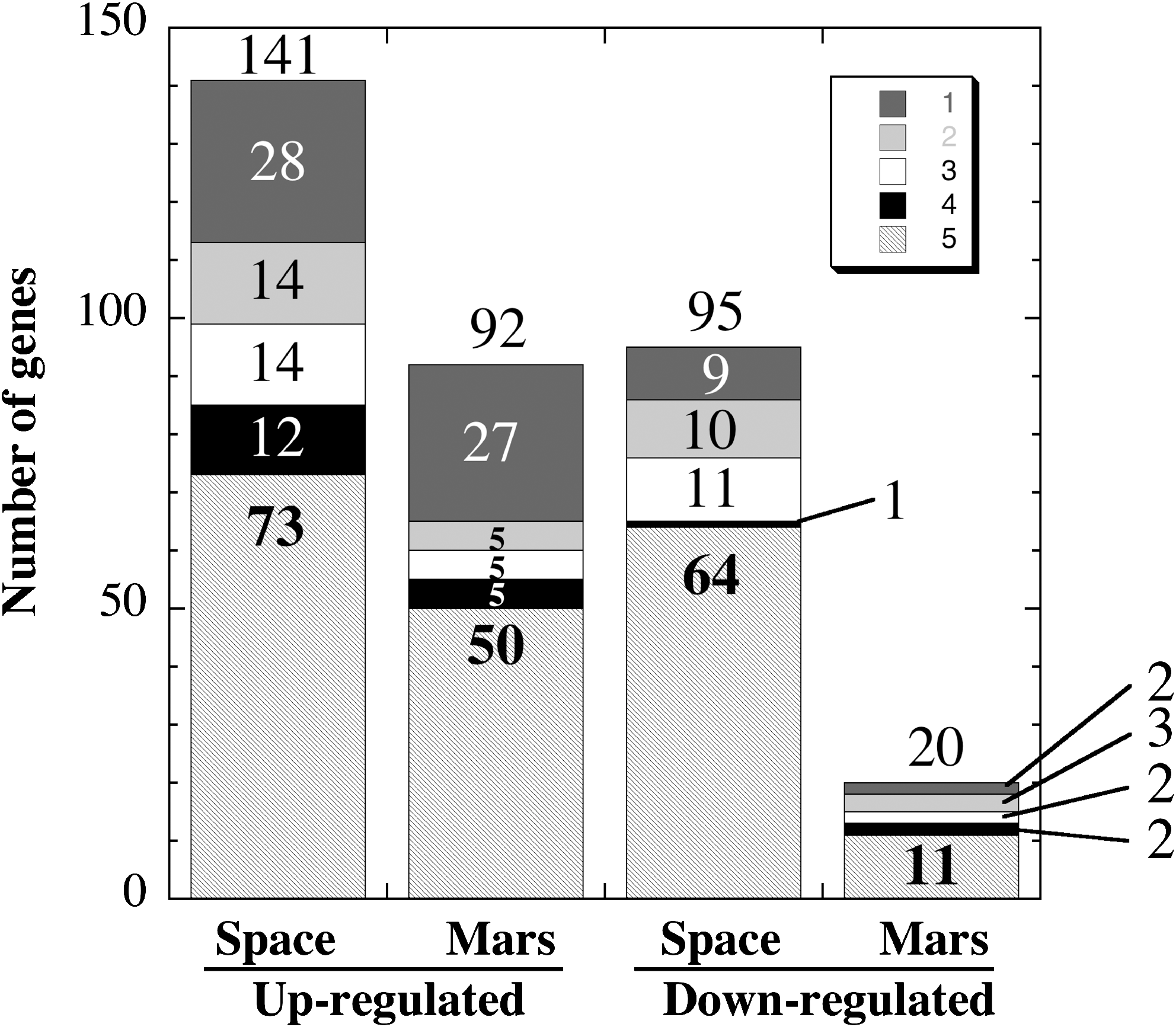
Summary of genes identified by microarray experiments as significantly up- or down-regulated from space- or simulated Mars-exposed spores. Categories are defined as described in Appendix 2 of Sonenshein et al. (2002). Categories are Cell envelope and cellular processes (1, dark gray bars); Intermediary metabolism (2, light gray bars); Information pathways (3, white bars); Other functions (4, black bars); and Unknown (5, hatched bars). See text for details.
Microarrays were probed with Cy3- or Cy5-labeled cDNA isolated from 60 min germinated space-exposed or simulated Mars-exposed spores and compared to 60 min germinated ground control spores.
Annotated functions assigned using the Bacillus subtilis Genome Database BSORF (
Genes showing an average≥3-fold up-regulation (n=4) with a P≤0.05 were considered significant.
nsd, not significantly different.
3.3. Up-regulated genes in space- and/or simulated Mars exposed-spores versus ground control spores
All bacteria, including B. subtilis, encode a large number of cross-regulated and interlocking regulons devoted to coping with environmental stresses (reviewed in Storz and Hengge, 2011). In B. subtilis, these stress regulons are controlled by a number of “master regulator” proteins such as RecA/LexA, sigma-B, CtsR, Fur, Spx, PerR, HrcA, ECF sigma factors, and so on (Tam et al., 2006; Elsholz et al., 2010). We found elevated transcript levels of several of these master regulators, including recA, sigV, hrcA, and ctsR; in most (but not all) cases, induction of these master stress-responsive genes was higher in space-exposed spores than in simulated Mars-exposed spores (Table 1).
3.3.1. RecA and the SOS regulon
Visual inspection of the data immediately revealed a striking up-regulation of genes involved in DNA repair and the SOS response in space-exposed spores but not in simulated Mars-exposed spores (Table 1). This result confirmed several earlier observations that exposure of spores to space conditions damages DNA (see Moeller et al., 2012 for details). The SOS regulon has been well characterized at the molecular level in vegetative cells of B. subtilis, including its response to DNA-damaging agents such as UV radiation, mitomycin-C, and mild ionizing radiation (Au et al., 2005; Goranov et al., 2006). These previous studies enabled us to compare transcriptional activation of SOS genes in space-exposed or simulated Mars-exposed spores to known genes involved in the SOS response.
The classic SOS regulon has been defined by genes whose expression was induced significantly by exposure of vegetatively growing B. subtilis cells to the DNA-damaging agents mitomycin-C (which causes mainly interstrand crosslinks) and 254 nm UV (which mainly causes pyrimidine dimers) (Au et al., 2005; Friedberg et al., 2006). We therefore compared the DNA repair and SOS regulon genes identified from germinated space- or simulated Mars-exposed spores (Table 1) to the set of genes previously identified by microarray experiments as belonging to the classic B. subtilis SOS regulon (Au et al., 2005) (Fig. 4). We observed that out of a total of 38 SOS-regulon genes defined by Au et al. (2005), 23 SOS genes were not significantly up-regulated during germination of either space- or simulated Mars-exposed spores (Fig. 4, white arrows). Fifteen SOS regulon genes were identified as induced significantly during germination of spores exposed to space but not in spores exposed to simulated martian conditions (Fig. 4, black arrows). A single SOS regulon gene (dnaE) was significantly up-regulated during germination of both space- and simulated Mars-exposed spores (Fig. 4, gray arrow with black outline). The yqjWXYZ operon was identified as belonging to the SOS regulon (Au et al., 2005) and was significantly up-regulated in space-exposed, but not simulated Mars-exposed, spores (Fig. 4). The first gene of the operon, yqjW, has been identified as encoding the homologue of the Y-family translesion bypass DNA polymerase UmuC (Sung et al., 2003; Rivas-Castillo et al., 2010), but the functions of the downstream genes yqjX, yqjY, or yqjZ are currently unknown (Table 1). Upstream from the yqjWXYZ operon is located the divergently transcribed yqzH gene, and downstream from yqjWXYZ are located the yqkABC operon and the yqkED divergon (Fig. 4). The functions of these flanking genes are currently not known, nor were they previously identified as SOS regulon genes. It is interesting to note, however, that expression of all genes in this region was specifically up-regulated during germination of space-exposed, but not simulated Mars-exposed, spores (Table 1; Fig. 4, gray arrows).
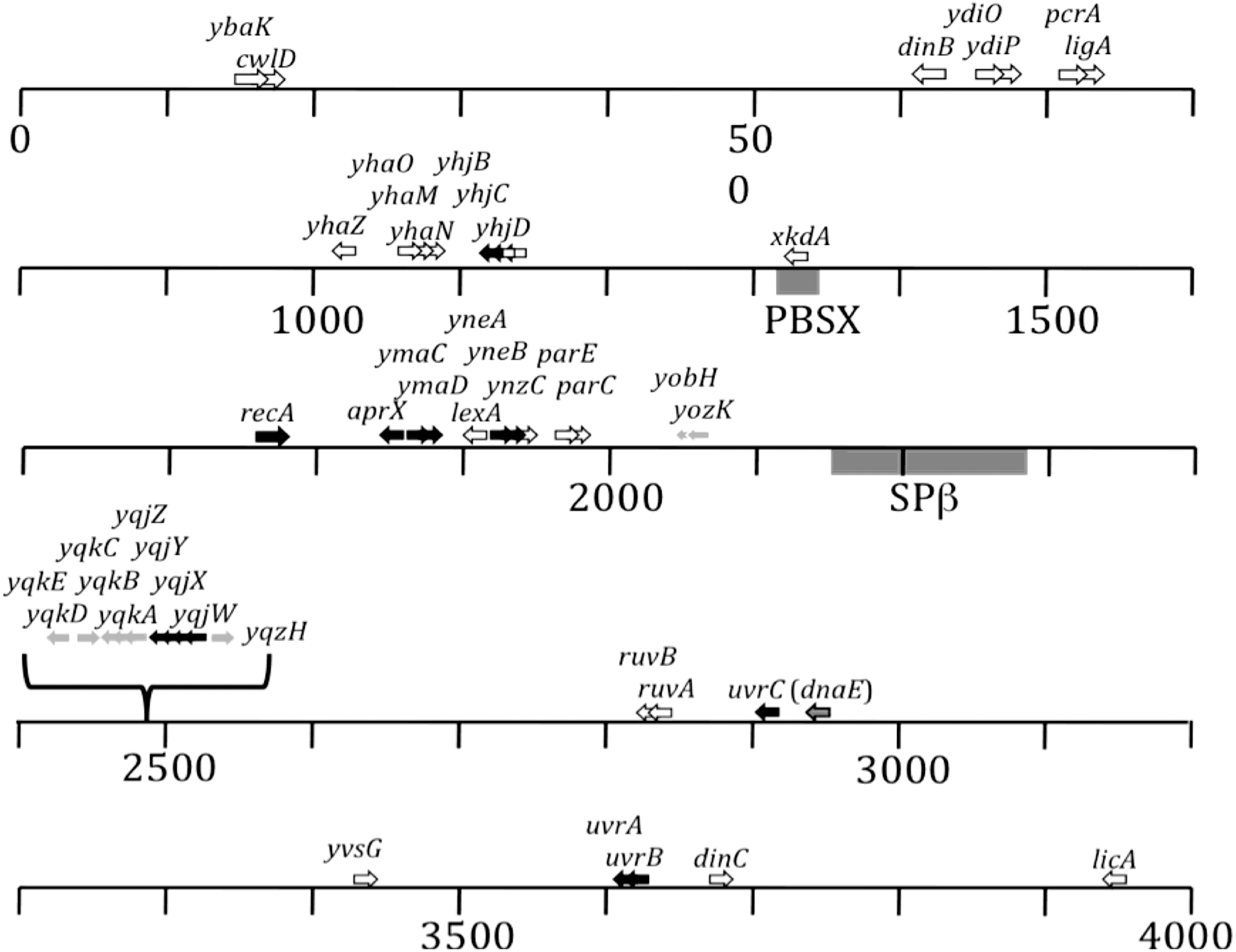
Map of the B. subtilis genome summarizing the locations of SOS regulon genes (modified from Au et al., 2005). Scale is in kilobase pairs. Shown are SOS regulon genes whose expression was not induced by exposure of spores to space or simulated martian conditions (white arrows); SOS regulon genes that were up-regulated in spores exposed to space, but not simulated martian, conditions (black arrows); genes whose expression was up-regulated in space-exposed spores, but not in Mars-exposed spores or defined in the SOS regulon (gray arrows); and the dnaE gene, which was identified as an SOS regulon gene and whose expression was up-regulated in both space- and simulated Mars-exposed spores (gray arrow with black outline). Overlapping arrows denote genes that belong to putative operons. The gray bars denote the locations of prophages PBSX and SPβ.
A novel set of SOS genes induced by treatments that introduce double-strand (ds) breaks in DNA was recently uncovered, including yneAB, msm, dinB, aprX, ymaC, yhaZ, lexA, and recA (Simmons et al., 2009). It is of interest to note that we observed significant up-regulation of a subset of these genes (yneAB, aprX, ymaCD, and recA) during germination of space-exposed, but not simulated Mars-exposed, spores (Table 1), which supports earlier findings that space exposure causes ds DNA breaks in spores (reviewed in Nicholson et al., 2000).
A dramatic consequence of the classic SOS response in B. subtilis is prophage induction, which results from activated RecA cleavage of prophage repressors (Yasbin et al., 1993). Bacillus subtilis strain 168 carries two resident prophages, SPβ and the defective prophage PBSX (Zahler, 1993). No up-regulated PBSX genes were identified in germinating space-exposed or simulated Mars-exposed spores; however, a large number of SPβ genes were up-regulated (Table 2). Consistent with an SOS-like response, out of a total of 186 SPβ genes (Sonenshein et al., 2002), 68 were up-regulated in space-exposed spores, and a subset of 38 were also up-regulated in simulated Mars-exposed spores (Table 2).
Footnotes are as described in Table 1.
3.3.2. Non-SOS regulon DNA repair genes
Further inspection of the data in Table 1 revealed a number of genes involved in DNA repair and recombination, not previously identified as part of the SOS regulon, that were significantly up-regulated in space-exposed but not simulated Mars-exposed spores (Table 1). The addABsbcDCyisB operon encodes the AddAB helicase/nuclease, the homologue of the E. coli RecBCD recombination repair protein (Kooistra et al., 1993; Yeeles et al., 2011), and sbcDC encode the SbcCD complex important in repair of strand breaks and crosslinks in B. subtilis (Sharples and Lloyd, 1993; Mascarenhas et al., 2006). The recUponA operon encodes RecU, a Holliday junction resolvase important for both double-strand break repair (Sanchez et al., 2005) and chromosome segregation (Pedersen and Setlow, 2000; Carrasco et al., 2004), and PonA, a penicillin binding protein (PBP-1) important in forming the cell division septum (Pedersen et al., 1999). The addABsbcDCyisB operon was only significantly up-regulated during germination of space-exposed spores, while the recU gene (but not the ponA gene) was up-regulated in both space- and simulated Mars-exposed spores (Table 1). In addition, the radA and disA DNA repair and integrity-scanning genes embedded in the ctsR operon (see Section 3.3.3 below) were induced only in space-exposed spores. Germination induction of these genes further supports the notion that DNA is a major target of damage by space exposure and, to a much lesser extent, by simulated Mars exposure.
3.3.3. Other stress response regulators
Various environmental stresses provoke protein damage and misfolding, and B. subtilis possesses a system for monitoring protein quality within the cell. A number of ATPases (ClpC, ClpE, ClpX, ClpY) bind to misfolded proteins and either act as molecular chaperones or direct the misfolded protein to the associated proteases ClpP or ClpQ for degradation (Gerth et al., 2004). The Clp protease system is regulated by the CtsR repressor encoded in the ctsRmcsABclpCradAdisA operon, which was found to be significantly up-regulated in space-exposed, but not simulated Mars-exposed, spores (Table 1). However, the Clp ATPase-encoding clpC and clpE genes, and the Clp protease-encoding clpP gene, were all significantly up-regulated in both space- and simulated Mars-exposed spores (Table 1). Particularly notable was the dramatic (>50-fold) up-regulation of clpE, perhaps due to the dual role of ClpE in protein quality monitoring and proteolytic regulation of CtsR activity itself (Miethke et al., 2006). The data suggest that protein damage/denaturation is also a likely consequence of exposure to space and, to a lesser extent, simulated martian conditions.
The HrcA repressor controls expression of the class I heat shock regulon, which consists of only two operons coding mostly for protein chaperones for refolding denatured proteins, hemNhrcAgrpEdnaK and groESgroEL (Schumann, 2003). Interestingly, only the hrcA transcript itself, not its target operons, was significantly up-regulated in both space- and simulated Mars-exposed spores (Table 1).
The PerR regulon consists of a group of genes induced in response to oxidative stress, including mrgA, katA, ahpCF, a heme biosynthetic operon (hemAXCDBL), a zinc uptake system (zosA), fur, and perR itself (Herbig and Helmann, 2002). Although perR, ahpCF, and fur transcripts were not significantly elevated, we did observe significant up-regulation of mrgA and katA during germination of both space- and simulated Mars-exposed spores. Also, significant up-regulation of hemAXCDBL was observed in space-exposed, but not simulated Mars-exposed, spores (Table 1). Interestingly, we also noted significant up-regulation of the hemEHY operon during germination of space-exposed, but not simulated Mars-exposed, spores (Table 1). The prior observation that heme biosynthesis requires all nine products from both hemAXCDBL and hemEHY operons (Hansson and Hederstedt, 1992) and that both hem operons were induced during germination of space-exposed spores (Table 1) suggests that the hemEHY operon may also be part of the PerR regulon.
The sigV gene encodes SigV, one of seven so-called extracytoplasmic function (ECF) sigma factors involved in response to antibiotics and other cell envelope-stressing stimuli (Guariglia-Oropeza and Helmann, 2011). A major operon induced by SigV is the autoregulated sigVrsiVoatAyrhK operon encoding SigV itself, which we found up-regulated by 4- to 6-fold during germination of both space- and simulated Mars-exposed spores (Table 1). However, another major SigV-dependent operon, dltABCDE (Guariglia-Oropeza and Helmann, 2011), was not significantly up-regulated in either set of spores.
3.3.4. Sporulation genes
It has previously been reported that a number of transcripts encoded by sporulation genes were found in the dormant spore, including the unknown genes ykzE, ymfJ, yhcV, yqfX, ythC, ythD, and coxA; genes encoding the minor small, acid-soluble spore proteins (SASPs) (sspN, tlp, and sspO); and the major SASP-γ gene sspE (Keijser et al., 2007). It was postulated that these represented abundant late sporulation transcripts that persisted into the dormant spore, consistent with the observation that these transcripts were rapidly degraded and disappeared within the first 30 min of spore germination (Keijser et al., 2007). We also found a number of sporulation genes apparently preferentially up-regulated during germination of both space- or simulated Mars-exposed spores. As reported by Keijser et al. (2007), we also noted increased transcript levels of the unknown genes yhcV, ykzE, ymfJ, yqfX, and ythC (but not ythD); genes encoding minor SASPs of the α/β-type (sspF, sspJ, sspM, sspNtlp, and sspOP) (Setlow, 2007); and the major SASP-γ gene (sspE) (Vyas et al., 2011) (Table 1). In addition, however, we noted significant up-regulation of the cgeAB operon, products of which are involved in maturation of the outermost spore layer (Roels and Losick, 1995); cotG, cotT, and cotVWXY, products of which are components of the spore coat (Driks, 1999; Kim et al., 2006); the safAcoxA operon, products of which are involved in spore morphogenesis and spore cortex formation (Ozin et al., 2000); and rapAphrA, products of which are involved in initiation of sporulation (Pottathil and Lazazzera, 2003) (Table 1).
3.4. Down-regulated genes in space- and/or simulated Mars-exposed spores versus ground control spores
In addition to genes up-regulated during germination, a smaller subset of significantly down-regulated genes was found (Table 3). Only 22 and 9 genes of known function were significantly down-regulated during germination of space- and simulated Mars-exposed spores, respectively (Fig. 2). Many of the down-regulated genes encoded products involved in the following: the cell envelope, such as transport proteins, lipid biosynthetic enzymes, and membrane-associated nuclease; the translational apparatus, such as a few ribosomal proteins and tRNA synthetases; and cold-shock proteins. Large-scale expression patterns of known regulons could not be readily discerned from the data without heavy speculation; thus the data are presented but not discussed in full detail.
Footnotes as in Table 1.
4. Discussion
4.1. Microarray analysis of PROTECT spore samples
Dormant B. subtilis spores were exposed to the space environment in low-Earth orbit and to a simulation of martian surface conditions for 559 days aboard the PROTECT experiment on the ISS. These environmental parameters were designed to simulate a hypothetical journey to, and residence on, Mars of spacecraft-contaminant spores (see Horneck et al., 2012). It has been known from previous experiments spanning the past 40 years that spore exposure to the space environment causes both lethal and mutagenic DNA damage to spores (reviewed in Nicholson et al., 2000; Horneck et al., 2012; Moeller et al., 2012). More recent research has also shown significant lethal and mutagenic DNA damage to spores exposed to simulated martian surface conditions (Schuerger et al., 2003, 2006; Perkins et al., 2008; Fajardo-Cavazos et al., 2010; Moeller et al., 2011). However, very little attention has been devoted to studying damage to other cellular constituents by space or Mars exposure. We knew that dormant spores lack metabolism and thus cannot repair cellular damage until metabolism is re-activated during germination (Nicholson et al., 2000). We reasoned that germinating spores would induce the expression of specific cellular repair pathways in response to the specific type of cellular component damaged; we could thus infer the type(s) of damage spores suffered by identifying which genes were specifically induced. To accomplish this goal, we performed transcription microarray analyses on RNA isolated from 60 min germinated space- or simulated Mars-exposed spores, versus RNA isolated from 60 min germinated spores from unexposed ground control samples. We found that germinating spores exhibited a strong stress response by inducing the expression of several known stress-related regulators including recA, sigV, hrcA, and ctsR. In general, two interesting observations were made. (i) Induction of these master stress-responsive genes was generally stronger and more extensive in space-exposed spores than in simulated Mars-exposed spores, which indicates that spore exposure to the space environment caused a wider spectrum and more severe cellular damages than spore exposure to simulated martian conditions. (ii) Induced expression of a particular master regulator usually caused induction of only a subset of that regulator's putative target genes, which implies that space or Mars exposure exerted a unique set of stresses upon dormant spores. It can be imagined that some of these stresses (e.g., ultrahigh vacuum or low-pressure CO2 atmosphere) are outside the range of conditions normally encountered by spores on Earth and thus provoke novel combinations of both cellular damages and stress responses. Specific responses will be discussed below.
4.2. SOS response
By far the major stress regulon induced in space-exposed (but not simulated Mars-exposed) spores was the SOS regulon. The recA and lexA gene products serve as the master regulators of the classic SOS regulon in B. subtilis (Au et al., 2005). The LexA protein is a transcriptional repressor of SOS-inducible genes. DNA damage provokes production of single-stranded DNA, which binds to RecA and converts it into a protease specific for cleavage of repressors of the LexA family. Cleavage of LexA leads to derepression of ∼50 LexA-repressed SOS target genes (including recA and lexA themselves).
We observed that expression of the recA gene was induced 7.5-fold in space-exposed spores but was not significantly induced in simulated Mars-exposed spores. Surprisingly, however, the microarray data failed to detect a statistically significant elevation in lexA expression in either space- or simulated Mars-exposed spores. This observation was puzzling because in previous spore germination microarray experiments we observed significant induction of both recA (5-fold) and lexA (3-fold) transcription. Bacillus subtilis 168 spores germinated for 60 min after irradiation with 254 nm UV (Moeller et al., unpublished), as would be expected for a classic SOS response. The data suggest that space exposure of spores may have induced a partial SOS response that was apparently independent of LexA cleavage. It is instructive to recall that the PROTECT spores analyzed in these experiments were shielded from solar UV (an inducer of the classic SOS response) but were subjected to space vacuum, extreme desiccation, and ionizing radiation, each of which are known to cause ds DNA breaks (reviewed in Nicholson et al., 2000). We know that the SOS response in B. subtilis is considerably more complex than its counterpart in E. coli (Yasbin et al., 1993) and that the two systems have been demonstrated to respond differently to ds DNA breaks induced in vegetative B. subtilis cells either artificially or by mild doses of ionizing radiation (Simmons et al., 2009). Furthermore, it has been shown that the DNA repair responses to ds DNA breaks in B. subtilis are substantially different in vegetative cells versus sporulating cells and spores (Moeller et al., 2007; Simmons et al., 2009). Therefore, it is not unreasonable to suppose that the circuitry controlling the SOS response during germination of spores exposed to space conditions could exhibit substantial differences from the classic SOS response. This might also explain the induction of non-SOS DNA repair genes such as addABsbcDC or recU. Regardless of the molecular details, the results strongly indicate that DNA is a major target of damage in spores exposed to the space environment and furthermore suggest that ds DNA breaks are involved.
4.3. Interlocking stress responses
From the PROTECT microarray experiments it became apparent that spores exposed to the space or simulated martian environment accumulated damage to cellular components in addition to DNA. Certainly spore proteins also sustained damage, as evidenced by partial induction of the CtsR/Clp chaperone/protease systems in both space- and simulated Mars-exposed spores. What environmental parameter(s) may have caused protein damage? Proteins can be denatured by heat or by extreme desiccation resulting from ultrahigh vacuum (Jaenicke, 1981), or they can become inactivated by ionizing radiation and reactive oxygen species (Berlett and Stadtman, 1997). In exposure to space on PROTECT, spores encountered cycles of heating and cooling, ultrahigh vacuum, and ionizing radiation, any of which could damage or denature proteins. In addition, contact with water and oxygen during subsequent germination of space-exposed spores would be expected to generate reactive oxygen species in levels higher than those experienced during the normal oxidative burst in germinating spores (Ibarra et al., 2008). Many individual stress regulons are controlled by partially overlapping control circuitries that respond to diverse environmental stimuli (reviewed in Herbig and Helmann, 2002; for recent examples, see Chaturongakul et al., 2011; Faulkner and Helmann, 2011; Hesketh et al., 2011). Therefore, it is perhaps not surprising to observe partial induction of overlapping stress regulons such as PerR, HrcA, CtsR, and SigV during germination of space- or simulated Mars-exposed spores.
4.4. Sporulation genes
As noted above (Section 3.3.3), a number of genes usually associated with sporulation were found to be up-regulated during germination of spores exposed to space or simulated martian conditions. Transcripts for some of these genes (ykzE, ymfJ, yhcV, yqfX, ythC, ythD, coxA, sspN tlp, sspO, and sspE) were previously found in dormant spores and disappeared rapidly within 30 min of initiation of germination (Keijser et al., 2007). In the PROTECT samples, were these sporulation transcripts simply left over from the previous round of sporulation, or were they synthesized de novo during germination? It is difficult to envision the former possibility for the following reasons. (i) Our RNA samples were harvested at 60 min of germination, by which time these same transcripts had disappeared in the germination experiments of Keijser et al. (2007). (ii) The induction levels we report here were calculated relative to RNA from non-treated ground control spores also harvested at 60 min of germination, thus controlling for any elevated transcript levels in non-treated spores. (iii) We found up-regulation of transcripts not found in the dormant spore, such as: sporulation genes expressed early in sporulation (rapAphrA); the additional SASP genes sspF, sspJ and sspM; and genes normally expressed from sigma-K promoters only in the mother cell (cgeAB, cotG, cotT, cotVWXY). Why space- or simulated Mars-exposed spores would up-regulate transcription of some sporulation genes is a mystery at present and can be resolved only by closer analysis of the expression of individual genes during germination with an independent method such as lacZ gene fusions.
4.5. Down-regulated genes
Although several apparently down-regulated genes were identified during germination of space- or simulated Mars-exposed spores, closer examination failed to find unifying regulons to which these genes belonged. There are likely a number of reasons for this. The majority of down-regulated genes belonged to the Unknown function category. Few genes of known function were found to be significantly down-regulated in space-exposed (22 genes) or simulated Mars-exposed (9 genes) spores, and most of these known genes were only modestly (3- to 5-fold) down-regulated. Previous investigations in the literature included measurement of the mRNA levels expressed by the same gene with two complementary techniques, microarray and quantitative reverse transcriptase-polymerase chain reaction, and concluded that the correlation between the two techniques was much greater for up-regulated genes (0.700; Spearman's Rho, P<0.0001, n=169) than for down-regulated genes (0.356; Spearman's Rho, P=0.0002, n=108); furthermore, the differences between the two groups were highly statistically significant (ANOVA, P=0.0042, n=10) (Morey et al., 2006). In other words, the authors concluded that microarray analysis is much better at identifying up-regulated than down-regulated genes (Miron et al., 2006; Morey et al., 2006).
4.6. Conclusions
Regarding planetary protection, the collective spore survival data (Horneck et al., 2012; Moeller et al., 2012; and Section 3.1 in this communication) indicate that spores shielded from solar UV radiation would likely suffer ∼50–90% reduction in viability during the cruise phase to Mars but could potentially survive for decades once deposited on the martian surface. The data presented in this communication demonstrate that microarray profiling can yield a substantial amount of insight into the types of cellular damages suffered by spores during long-term exposure to space or to a simulated martian environment. The results (i) confirm earlier investigations that indicated that spore DNA is a major molecular target for damage, (ii) demonstrate that the UV-shielded martian surface environment is much less harsh than is the UV-shielded space environment, and (iii) indicate that germinating spores are faced with repair of oxidative damage likely resulting from long-term exposure to ionizing radiation, vacuum, extreme desiccation, or a combination of these parameters. From a planetary protection perspective, the results indicate that a fraction of forward-contaminant spores on robotic spacecraft could survive an Earth-to-Mars transfer but would suffer substantial cellular damage during the journey.
(Other) Members of the PROTECT Team and Their Affiliations
Jean Cadet: Laboratoire “Lésions des Acides Nucléiques,” SCIB-UMR-E n°3 (CEA/UJF) Institut Nanosciences et Cryogénie CEA/Grenoble, Grenoble, France.
Thierry Douki: Laboratoire “Lésions des Acides Nucléiques,” SCIB-UMR-E n°3 (CEA/UJF) Institut Nanosciences et Cryogénie CEA/Grenoble, Grenoble, France.
Rocco L. Mancinelli: Bay Area Environmental Research Institute, NASA Ames Research Center, Moffett Field, California, USA.
Corinna Panitz: Institut für Flugmedizin, Technical University RWTH, Aachen, Germany.
Elke Rabbow: German Aerospace Center (DLR), Institute of Aerospace Medicine, Radiation Biology Department, Cologne, Germany.
Petra Rettberg: German Aerospace Center (DLR), Institute of Aerospace Medicine, Radiation Biology Department, Cologne, Germany.
Andrew Spry: Jet Propulsion Laboratory, Pasadena, California, USA.
Erko Stackebrandt: German Collection of Microorganisms and Cell Cultures GmbH (DSMZ), Braunschweig, Germany.
Parag Vaishampayan: Jet Propulsion Laboratory, Pasadena, California, USA.
Kasthuri J. Venkateswaran: Jet Propulsion Laboratory, Pasadena, California, USA.
Footnotes
Acknowledgments
We thank Patricia Fajardo-Cavazos for insightful discussions, Samantha Waters for careful examination of the microarray analyses, and the UF-ICBR for performing the microarray experiments described in this work. This study was supported in part by grants from the NASA Planetary Protection Office (NNA06CB58G and NNX10AV22G) to W.L.N. and by the DLR grant DLR-FuE Projekt ISS-Nutzung in der Biodiagnostik, Programm RF-FuW, Teilprogramm 475 to R.M.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
ANOVA, analysis of variance; COSPAR, the Committee on Space Research; ds, double strand; ECF, extracytoplasmic function; ISS, International Space Station; LB, Luria-Bertani; ORF, open reading frame; PBS, phosphate-buffered saline; SASP, small, acid-soluble spore protein; UF-ICBR, University of Florida Interdisciplinary Center for Biotechnology Research.