
Abstract
The survival strategies of one cyanobacteria colony and three terricolous lichen species from the hot subdesert of Tabernas, Spain, were studied along with topographical attributes of the area to investigate whether the protective strategies adopted by these pioneer soil colonizers are related to the environmental stressors under which they survive. A handheld Raman spectrometer was used for biomolecular characterization, while the microclimatic and topographic parameters were estimated with a Geographic Information System (GIS). We found that the survival strategies adopted by those organisms are based on different combinations of protective biomolecules, each with diverse ecophysiological functions, such as UV-radiation screening, free-energy quenching, antioxidants, and the production of different types and amounts of calcium oxalates. Our results show that the cyanobacteria community and each lichen species preferentially colonized a particular microhabitat with specific moisture and incident solar radiation levels and exhibited different adaptive mechanisms. In recent years, a number of studies have provided consistent results that suggest a link between the strategies adopted by those extremophile organisms and the microclimatic environmental parameters. To date, however, far too little attention has been paid to results from Raman analyses on dry specimens. Therefore, the results of the present study, produced with the use of our miniaturized instrument, will be of interest to future studies in astrobiology, especially due to the likely use of Raman spectroscopy at the surface of Mars. Key Words: Hot desert—Raman spectroscopy—Topography—Terricolous lichens—Cyanobacteria—Planetary exploration. Astrobiology 12, 743–753.
1. Introduction
Synthesis of biochemical metabolites, such as oxalates, carotenoids, chlorophyll, parietin, emodin, atranorin, gyrophoric acid, fumarprotocetraric acid, rhizocarpic acid, calycin, usnic acid, and so on, is a common strategy against some extreme conditions. Such biomolecules and pigments play important roles as antidesiccants and water-replacement molecules, UV-radiation screen compounds, antioxidants, DNA repair agents, or energy-quenching molecules (Nealson, 1997; Cockell and Knowland, 1999; Wynn-Williams and Edwards, 2000; Edwards, 2007).
In recent years, there has been an increasing interest in the detection of biochemical metabolites produced by lichens and cyanobacteria colonies in response to external stress as a vital part of understanding life processes under hostile conditions (Jorge-Villar and Edwards, 2010). Nevertheless, despite the critical role of protective pigments in lichen and cyanobacteria survival, little is known about their specific biological role, and in fact, the dynamics of those communities have rarely been studied. Furthermore, few studies have attempted to discern whether environmental factors regulate pigment synthesis in such communities (Garcia-Pichel and Castenholz, 1991; Ehling-Schulz et al., 1997; Sinha et al., 2001; Dillon et al., 2002; Singh et al., 2011).
The study of survival strategies developed by extremophiles would provide significant assistance to the effort of understanding the distribution of life under severe environmental conditions. Knowing which chemicals could serve as signatures of terrestrial life in extreme habitats, such as hot or cold deserts or acidic or alkaline waters, would help in the design of improved instruments for the detection of life on other planetary surfaces.
Raman spectroscopy stands out among analytical techniques as the most suitable for identifying organic and inorganic compounds in these extreme conditions (Edwards et al., 1999, 2003a, 2003b; Wang et al., 2004). It provides molecular information and is able to characterize any mixture of organic and inorganic compounds, including polymorphous and isomorphous differentiation. No chemical or physical sample manipulation is necessary, and micro- as well as macrospecimens can be analyzed. Furthermore, it is a nondestructive technique, so that after analyses the sample can be completely recovered and reused. Development of small commercial handheld Raman spectrometers, with long-lasting batteries and optical fibers, makes it possible to conduct on-site field measurements. These characteristics have made Raman spectroscopy an important technique in astrobiology such that it is currently being considered by NASA and ESA for use in martian surface exploration (Jorge-Villar and Edwards, 2005).
Desert habitats are considered plausible analogues for possible extraterrestrial life. Studying and understanding the adaptive mechanisms of extremophiles in desert environments, and the subsequent recognition of biomarkers, will be of great help in the search for life on extraplanetary surfaces. This is of particular interest for the identification of possible signs of life on Mars, since at this point in its geological evolution there is little or no liquid water on its surface. To date, investigations of biomarkers and survival strategies adopted by extremophiles in which Raman spectroscopy has been used have focused primarily on cold deserts (Wynn-Williams and Edwards, 2000; Edwards et al., 2003a, 2004a, 2004b, 2005; Jorge-Villar et al., 2005b), and only a few studies have been carried out in hot deserts (Jorge-Villar et al., 2005a; Jorge-Villar and Edwards, 2010). It would, therefore, be of great interest to compare the survival strategies of extremophiles in hot deserts with those in cold deserts. Since we do not know whether life exists, or existed, on Mars, nor do we know where or when life might have occurred, we can only speculate as to the environmental conditions under which life might have subsisted. Nevertheless, it is frequently assumed that, during the final period when life may have existed on Mars, conditions were likely to have been cold and dry. It would, therefore, be useful to employ Raman spectroscopy to study as many terrestrial habitats as possible to discern whether different survival strategies can be detected. Obviously, the biomolecular degradation could be different under hot or cold conditions, which would lead to different spectroscopic signatures. The identification of those spectral characteristics is of vital importance for the recognition of life's signals. These studies begin, on Earth, with the identification of different adaptive strategies in extreme environments and their comparison for a deeper understanding and interpretation of Raman spectra. The Tabernas Desert, located in southeast Spain, is a unique environment in Europe for its arid climate, geomorphological dynamics, biodiversity, and endemic species, and it is therefore an ideal scenario for this type of study. Vegetation is sparse or, in places, absent in this region's intensely dissected natural badlands landscape (Bryan and Yair, 1982), which is characterized by a dense drainage network and short steep slopes with narrow watersheds (Scheidegger et al., 1968). Geomorphologically, badlands are like a miniature desert developed on nonconsolidated and relatively impermeable material that is exposed to rapid water erosion (Campbell, 1989). Both lithology and climate determine the geomorphological dynamics of this system, which form an extremely complex topography, and as a result, important microclimatic variations are noticeable within short distances (Cantón et al., 2001). Significant differences in slope, aspect, insolation, and humidity typical of the badlands could induce primary colonizing species, such as lichens and cyanobacteria, to develop specific adaptive strategies in these extreme, dynamic, and active environments.
We present here, for the first time, a Raman spectroscopy study of four extremophile organisms from the Tabernas Desert (Spain): a cyanobacteria colony and three lichen species. Insolation and other topographical environmental parameters of this hostile habitat were analyzed within a Geographic Information System (GIS) in an attempt to relate the biostrategies developed by the four organisms with environmental parameters as well as with their field distribution. The combination of both analytical techniques would be valuable for obtaining complementary information about complex adaptation strategies to environmental factors implemented by different extremophile communities.
2. Methodology
2.1. Experimental area
The study area, the Tabernas Desert in southeastern Spain (Province of Almería), is one of the largest badlands in Europe with an area of 150 km2. It is located in the Sorbas–Tabernas basin and surrounded by several Betic ranges: the Gador, Nevada, Filabres, and Alhamilla Mountains (Fig. 1). Specifically, the study site is a 12.65-hectare gully, located between 37°0′40″N, 2°26′29″W and 37°0′55″N, 2°26′10″W. The altitude ranges from 247.5 to 382.5 m. The landscape is characterized by its complex geomorphology with bare, eroded, and poorly developed soils (Epileptic Regosol) in steeper southwest-facing slopes (gradients up to 70°). In contrast, northeast-facing slopes with gradients less than 30° usually have more developed soils (Endoleptic Regosols) that are covered with biological crusts, including many species of terricolous lichens such as Diploschistes diacapsis, Squamarina lentigera, and Lepraria crassissima as well as cyanobacteria. At the pediments, on gentle slope gradients, soils are thicker (Haplic Calcisol) (Miralles-Mellado et al., 2011) and covered by biological soil crusts in open areas with patches of annual plants (dominated by Stipa capensis) and scattered perennial plants (with predominance of Helianthemun almeriense, Hammada articulata, Artemisia barrelieri, Salsola genistoides).
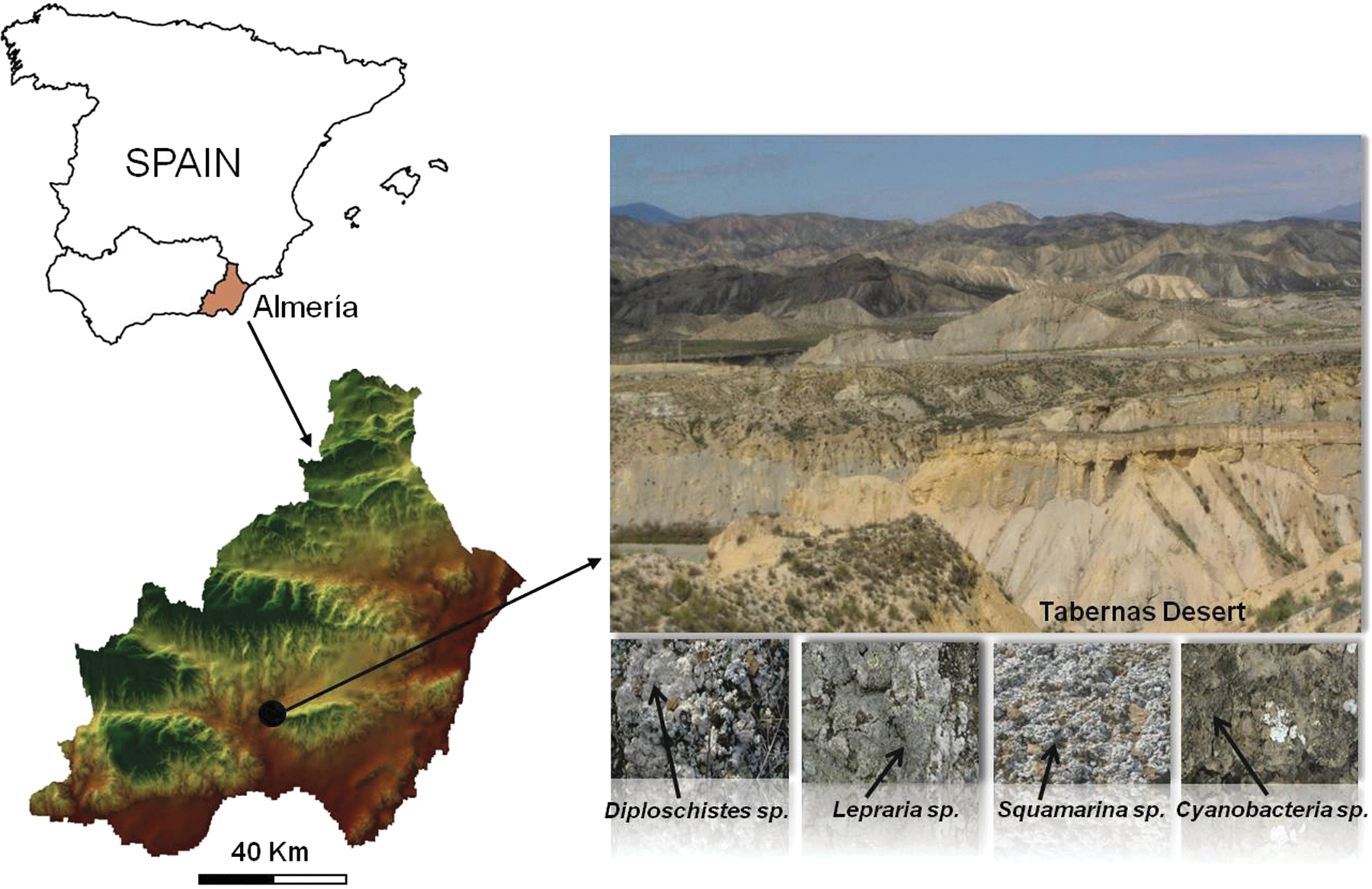
Study area and specimens. Color images available online at
The lithology is mainly a carbonated-gypsiferous Miocene mudstone, and the average mineralogical composition is muscovite 35%, paragonite 10%, chlorite + smectite 3%, quartz 10%, calcite 20%, dolomite 2%, and gypsum 20% (Cantón et al., 2001). The climate is characterized by a particularly strong water deficit during the summer months. The average annual temperature is 17.9°C, with an absolute maximum of 45°C and absolute minimum of −4.5°C, and average annual rainfall is 235 mm (estimated from a 30-year record of the Tabernas weather station), with high inter- and intra-annual variability. The maximum, 24 h rainfall during the study period was 76 mm. Rainfall intensity in the region exceeds 100 mm h−1 only for a 5 min interval, or 150 mm h−1 for intervals under 1 min. Annual potential evapotranspiration is around 1500 mm, which indicates a considerable annual water deficit.
2.2. Specimens
We sampled four biological soil crust (BSC) types that are very common and abundant in the Tabernas Desert. One of them is dominated by cyanobacteria and the others by chlorolichens classified as three different lichen crust types: Lepraria crassissima, Diploschistes diacapsis, and Squamarina lentigera. The crust types were selected based on the work of Lázaro et al. (2008), who classified BSC types in the Tabernas Desert according to their development stage, using crust composition and color as indicators. The main characteristics of these BSCs are as follows:
(1) BSC dominated by cyanobacteria (Fig. 1). The crust is made up of diverse species, with a relatively high proportion of Microcoleus sheath material. This crust seems to be a primary colonizer of bare soils and progressively, when the environmental conditions are suitable, is replaced by more evolved BSCs dominated mainly by lichens (Lázaro et al., 2008; Louise-Bevan, 2008).
(2) BSC dominated by L. crassissima (Fig. 1). This highly diverse crust is associated with small lichen species (Collema spp., Placynthium nigrum, and Toninia sedifolia) and cyanobacteria, and is typically found on steep north-facing slopes, where it is only able to colonize the shadiest spaces among vascular plants (Lázaro et al., 2008; Louise-Bevan, 2008).
(3) BSC dominated by D. diacapsis (Fig. 1). This crust forms large plate-like mats. It is scattered around a variety of microhabitats and covers mostly gentle slopes with any orientation, including completely exposed, flat areas, and pediments (Lázaro et al., 2008; Louise-Bevan, 2008).
(4) BSC dominated by S. lentigera (Fig. 1). This squamulose lichen-crust colonizes predominantly intensely rugged microhabitats on slopes, with a preference for the eastern orientation. It is particularly fragile when dry due to its rough morphology (Lázaro et al., 2008; Louise-Bevan, 2008).
2.3. Raman spectroscopy
Micro-Raman spectra were collected with a miniaturized handheld instrument, the Deltanu Inspector Raman spectrometer, with a Nuscope microscope attachment and 100× magnification lens. Laser excitation was at 785 nm. The spot size was 35 μm. Lichens were placed on a manual mobile platine with no optical fiber. The highest laser power available was 120 mW at the source. However, of the power settings available (i.e., very low, low, medium, high, and very high), we chose between the very low and the low intensities to avoid damage to the sample by heating.
To improve the signal-to-noise ratio, we used 1–5 s exposure time and 5–10 accumulations. The spectral resolution was 8, 10, and 12 cm−1, but for compound identification we usually used 8 cm−1. The wavenumber range was 200–2000 cm−1.
Our Raman analyses in a laboratory located at the University of Burgos (Spain) were first performed on dry specimens collected after a long period of drought. Samples were then sprayed with water and, after approximately 10 min, analyzed again. After finding that results could only be acquired from humid samples, field analyses were repeated during three rainy days. The Raman spectra were performed under shelter to protect both spectrometer and computer from water. Nevertheless, experimental parameters, such as temperature, humidity, wind, and altitude did not change from the original field sampling area.
A large number of areas (an average of 30 different spots) were surveyed on each specimen for a more complete knowledge of the biopigments. This procedure also provides a rough idea of the prevalence of biomolecules, since the most abundant pigment will appear in more spectra than scarcer ones.
2.4. Topographical attributes
The locations of the different BSC types (D. diacapsis, S. lentigera, L. crassissima, and cyanobacteria) were recorded at 96 points during the sampling campaigns with a Differential Geo Positioning System. The most important topographical attributes of each point were derived from a 1 m resolution digital elevation model (DEM) with the Idrisi GIS (version Kilimanjaro) and the Solar Analyst 1.0 extension of the ArcGis (version 8.1) GIS software. The terrain attributes extracted were
(i) Altitude;
(ii) Slope gradient;
(iii) Gradient on orientations from least to most exposed to insolation, according to the orientation classes established by Parker (1982) for calculating the Topographic Relative Moisture Index (TRMI);
(iv) Contributing area (ARE), which is the specific area that drains into each site, with use of a single-drainage-direction algorithm (Quinn et al., 1991);
(v) Wetness index (W), developed by Beven and Kirkby (1979), related to the spatial distribution and size of zones of saturation or variable source areas for runoff generation. Moore et al. (1988) found a strong correlation between the distribution of W and the distribution of surface soil water content.
(vi) Slope length factor (LSF), which is an index for the potential sediment transport with use of an algorithm derived from the unit stream power theory (Moore and Burch, 1986)
where n and m are constants (n=0.4, m=1.3).
(vii) Global, direct, and diffuse solar radiation at summer and winter solstices and at equinoxes. Direct solar radiation analysis was carried out with a simple transmission model for each location (List, 1971; Gates, 1980; Pearcy et al., 1989; Rich, 1989, 1990), and the direct radiation received was calculated based on the position of the Sun, atmospheric attenuation, and surface orientation. For diffuse solar radiation, the standard model, which considers that the diffuse radiation varies with zenith angle, was used (Pearcy et al., 1989; Rich, 1989, 1990). Global radiation for each location was found as the sum of the direct and diffuse radiation.
(viii) Hours of insolation received by each sampling point at summer and winter solstices and equinoxes.
For each sampling point, terrain and radiation attributes were calculated from the averages of nine values found from 3×3 m windows (3×3 pixels).
3. Results
3.1. Raman spectroscopy results
3.1.1. Cyanobacteria
There is no evidence of scytonemin, chlorophyll, or carotenoids in any of the Raman spectra collected on cyanobacteria with our portable Deltanu Inspector Raman spectrometer. The absence of these molecules in the spectra could be related to the characteristics of the mini-Raman instrument. It is also possible that it results from low cell densities in the crust or from a masking of the signal by the mucilaginous sheath material. Signatures at 1416, 1408, 1359, and 945 cm−1 suggest a mycosporine-like amino acid (MAA), whereas the 1596, 1540, 1539, 1237, 1187, 1038, 894, 746, 684, 598, 553, and 381 cm−1 bands could be assigned to amino acids, such as L-phenylalanine, L-tyrosine, and L-triptophane (Fig. 2).
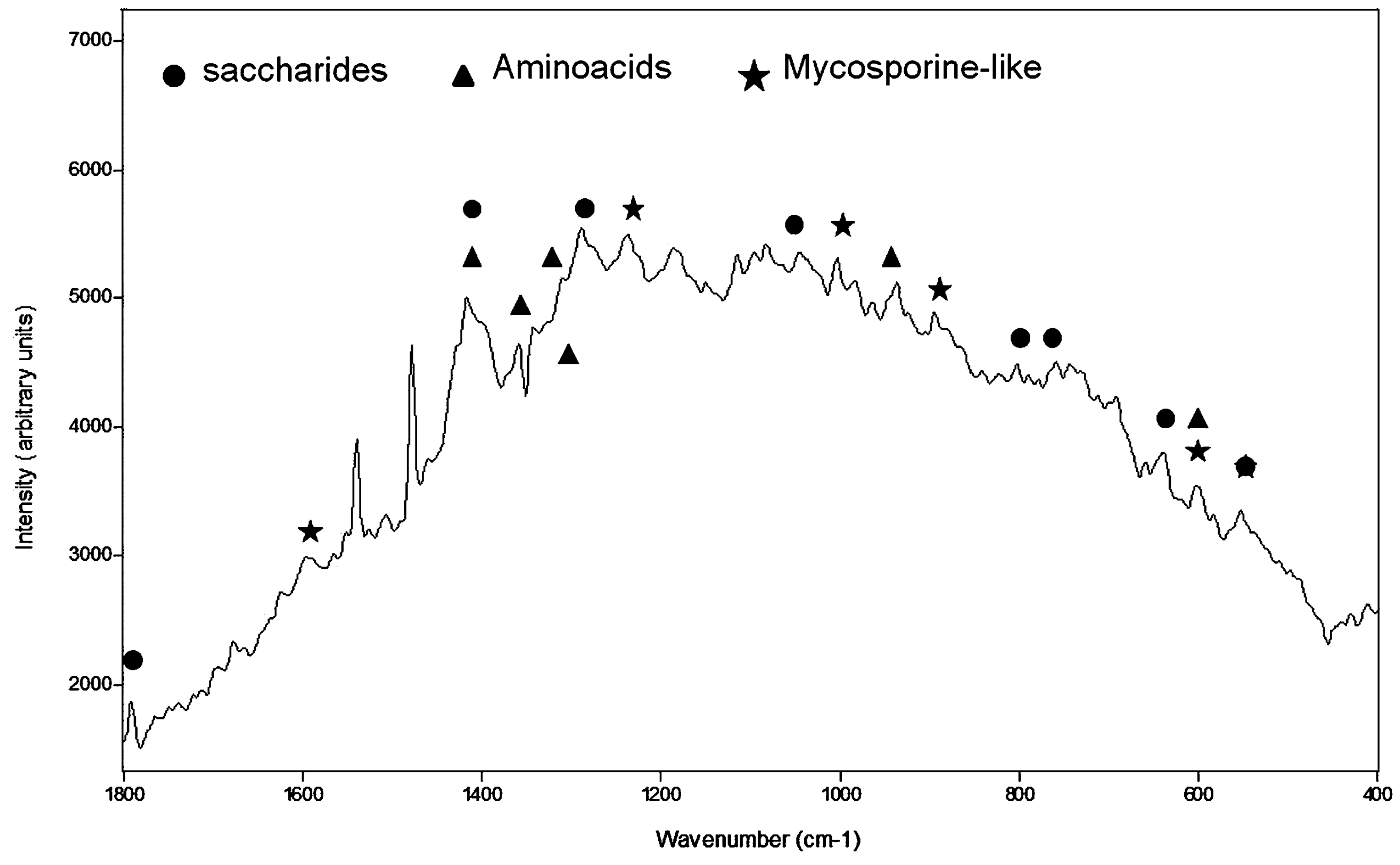
Raman spectrum achieved on cyanobacteria colony showing bands assigned to mycosporine-like amino acids, saccharides, and amino acids.
3.1.2. Lichens
In the analysis of dried lichens, spectra show strong fluorescence, and no Raman signatures appear in any of the spectra acquired. However, Raman spectra of moist lichens show broad bands (Table 1). Biomolecules usually display complex Raman spectra, with numerous signatures, particularly in the wavenumber range 1600–900 cm−1. Because of the size of the spot analyzed (35 μm) while using our handheld Raman spectrometer, signatures of more than one pigment may contribute to the same Raman feature; this is reflected as a broad band in the spectra. The low spectral resolution also contributes to this phenomenon. As a result, the new signature would neither be centered at any position of the characteristic molecular signatures of any compound nor at the average place for the specific wavenumber of each compound. Instead, it would have shifted unknown wavenumbers to higher or lower regions of the Raman spectrum, making correct biomolecular assignment very difficult.
CAR, carotenoid; OX, oxalate.
3.1.2.A. Squamarina lentigera
The Raman bands at 1492, 1388, 1325, 1074, 980, and 914 cm−1 were assigned to chlorophyll, whereas signatures at 1521, 1153 cm−1 are characteristic of a carotenoid. Both bands are related to the stretching vibration of the C&z.dbnd;C and C–C carotenoid backbone chains, respectively, and suggest the presence of zeaxanthine. Parietin could also be identified because of the bands at 1587, 1540, 1466, 1388, 1286, 1190, 980, 842, 620, 582, 431, 383, and 323 cm−1; signatures at 1760, 1587, 521, 1487, 1466, 1340, 1286, 1250, 1190, 997, 701, 646, 620, 504, and 431 cm−1 could indicate that rhizocarpic acid was also produced by this lichen, and bands at 1629, 1604, 1530, 1480, 1462, 1323, 1287, 1184, 1152, 1142, 1070, and 992 cm−1 could be related to usnic acid. Calcium oxalate monohydrate (whewellite) was identified by the Raman signatures at 1488, 1466, and 504 cm−1.
3.1.2.B. Diploschistes diacapsis
A weak signature centered at 1503 cm−1 could be associated with decaprene-beta-carotene, a long-chain carotenoid, and the band at 1505 cm−1 to astaxanthine, whereas the peak at 1520 cm−1 could be related to zeaxanthine. The broad signature at 1322 cm−1 was related to chlorophyll.
Bands at 1689, 1594, 1460, 1351, 1284, 1184, 1127, 985, 692, 601, and 537 cm−1 and signatures at 1786, 1735, 1680, 1594, 1503, 1351, 1284, 1184, 1033, 979, 780, 711, 780, 692, and 601 cm−1 could be assigned to usnic acid and rhizocarpic acid, respectively. We could infer the presence of emodin from 1700, 1665, 1589, 1416, 1290, 1136, 1096, 939, 495, and 468 cm−1 peaks, although some of its characteristic signatures do not appear in the spectrum. Strong broad bands centered at 1290 and 1416 cm−1 along with 1096 cm−1 could be related with a saturated fatty acid, but this assignment is doubtful.
3.1.2.C. Lepraria crassissima
Raman bands at 1753, 1713, 1676, 1380, 1315, 1287, 1135, 1082, and 1029 cm−1 could be related to the presence of fumarprotocetraric acid, and the signatures at 1604, 1592, 1483, 1378, 1348, 1315, 1242, 1111, 1029, 1002, and 943 cm−1 could be assigned to calycin. Lutein, a carotenoid, could be identified because of the 1525 and 1154 cm−1 bands. Some Raman spectra show signatures of calcium oxalate dihydrate (weddellite) at 1473 and 910 cm−1 with shoulders of calcium oxalate monohydrate (whewellite), centered at 1468 and 1486 cm−1. A very weak broad feature centered at 1320 cm−1 was related to chlorophyll.
3.2. Topographical attributes
Although there is little difference in the altitude and slope range of the biological crust types analyzed (Diploschistes diacapsis, Squamarina lentigera, Lepraria crassissima, and cyanobacteria) (Table 2), there are differences between L. crassissima and the other lichen species (D. diacapsis and S. lentigera), and cyanobacteria as well, in the results for the W and LSF topographical indices and for the ARE (Table 2). In general, the L. crassissima ARE, LSF, and W indices were higher than D. diacapsis, S. lentigera, and cyanobacteria (Table 2). L. crassissima, D. diacapsis, S. lentigera, and cyanobacteria were also observed to differ in the TRMI. Eighty percent of L. crassissima sampled was facing north, while 56.6% of D. diacapsis and S. lentigera lichens sampled faced preferentially east and west and 10.7% south. Forty percent of the cyanobacteria faced east-west and 23.3% south.
Av±SD, average±standard deviation.
Some differences were also observed in the direct, diffuse, and global incident solar radiation during both the equinoxes and solstices among L. crassissima, D. diacapsis, S. lentigera, and cyanobacteria. L. crassissima had lower global, direct, and diffuse incident solar radiation at the equinoxes and solstices than the other lichens and cyanobacteria (Table 2). However, it is noteworthy that, in the winter solstice, diffuse incident solar radiation was even higher than direct solar radiation (diffuse=402 W m−2) for this lichen. Cyanobacteria showed slightly higher incident solar radiation (diffuse, direct, and global) at the winter solstice and equinoxes, and lower at the summer solstice (Table 2), than D. diacapsis and S. lentigera. The number of hours of incident sunlight is very similar for all species, although somewhat lower for L. crassissima (Table 2).
4. Discussion
4.1. Raman spectroscopy of pigments and biomolecules in extremophile organisms and their ecophysiological functions
The fact that analysis of dry lichens generated no recognizable Raman signatures in the spectra and analysis of wet specimens generated bands that are clearly distinguishable from fluorescence (Table 1) is strikingly different from analyses performed in a laboratory of the University of Bradford (UK) on other lichen species (Edwards et al., 2003a, 2003b, 2003c, 2005; Jorge-Villar et al., 2005a; Jorge-Villar and Edwards, 2010). In those analyses, there was no problem in acquiring and identifying biopigments from the Raman spectra. This could be explained, on the one hand, by the characteristics of the handheld spectrometers, which may have technical limitations that restrict their capacity to detect weak signals. On the other hand, the lack of recognizable Raman signatures could be due to the physical status of desiccated cells and molecules of terriculous lichens living in the extreme environment of the Tabernas Desert. Heber et al. (2007, 2010) documented a reversible inactivation of photosystem II upon desiccation in cyanolichens, presumably as a photoprotective mechanism. They concluded that the desiccation induced conformational changes of the pigment–protein complex, which resulted in the fast radiationless dissipation of absorbed light energy. Energy dissipation is inactivated and structural changes are reversed when water becomes available again (Heber, 2008). The increase in the reflectivity of dry versus wet specimens could interact with the instrument and contribute to decrease its sensitivity even more.
Raman spectra of cyanobacteria in the Tabernas Desert do not show evidence of scytonemin, chlorophyll, or carotenoids, although those pigments were detected in cyanobacteria specimens from other areas analyzed by our group in other projects (Chu et al., 1998; Edwards et al., 1999, 2004c, 2005; Wynn-Williams et al., 2000; Jorge-Villar et al., 2005b, 2006). Nevertheless, these organisms did show evidence of compounds such as saccharides (maltose, glucose, or both), amino acids (L-phenylalanine, L-tyrosine, and L-triptophane), and MAA (Fig. 2). The exopolysaccharides excreted could be carrying out an important protective role against desiccation and UV irradiation in the cyanobacteria (Knowles and Castenholz, 2008; Chen et al., 2009).
Regarding the protective pigments in the lichen samples found by Raman spectroscopy, the spectra show that chlorophyll, a photosynthetic pigment, was present in all lichens. The miniaturized Raman spectrometer was unable to discern different types of chlorophyll from the spectra; however, we can propose assignment of carotenoid types (Table 1). In S. lentigera and L. crassissima, carotenoids were assigned to zeaxanthine or lutein, respectively, whereas in D. diacapsis astaxanthine, zeaxanthine, and decapreno-beta-carotene were identified. The dual role of carotenoids as efficient UV photoprotective molecules (Cockell and Knowland, 1999) and auxiliary DNA repair agents (Edwards et al., 2006; Edwards, 2007) has been recognized in extremophile colonies. Carotenoid pigments also quench the excess energy not used photosynthetically (especially UVB), which would otherwise generate highly toxic singlet oxygen (Cockell and Knowland, 1999).
Based on the large number of Raman spectra acquired for each lichen specimen, we can suggest that there are differences, not only in the type of oxalate produced by these organisms but also in the relative proportion in which those biomolecules appear (Fig. 3). L. crassissima showed mono- and dihydrate calcium oxalates (whewellite and weddellite, respectively), from which weddellite was significantly more abundant. In S. lentigera, calcium oxalate monohydrate (whewellite) was observed, and no bands of weddellite appeared. Only one spectrum on D. diacapsis shows signatures of calcium oxalates, in this case whewellite. The ability of lichens to exude calcium oxalates is associated with a significant antidesiccative water-storage strategy for their survival in extremely dry habitats (Edwards et al., 1997, 2003d; Seaward and Edwards, 1997; Holder et al., 2000; Jorge-Villar et al., 2004, 2005a, 2006; Jorge-Villar and Edwards, 2010) and also as a strategy for removing oxalic acid as waste from the Krebs metabolic cycle (Edwards et al., 1997; Seaward and Edwards, 1997; Jorge-Villar et al., 2005a, 2005b; Edwards, 2007).
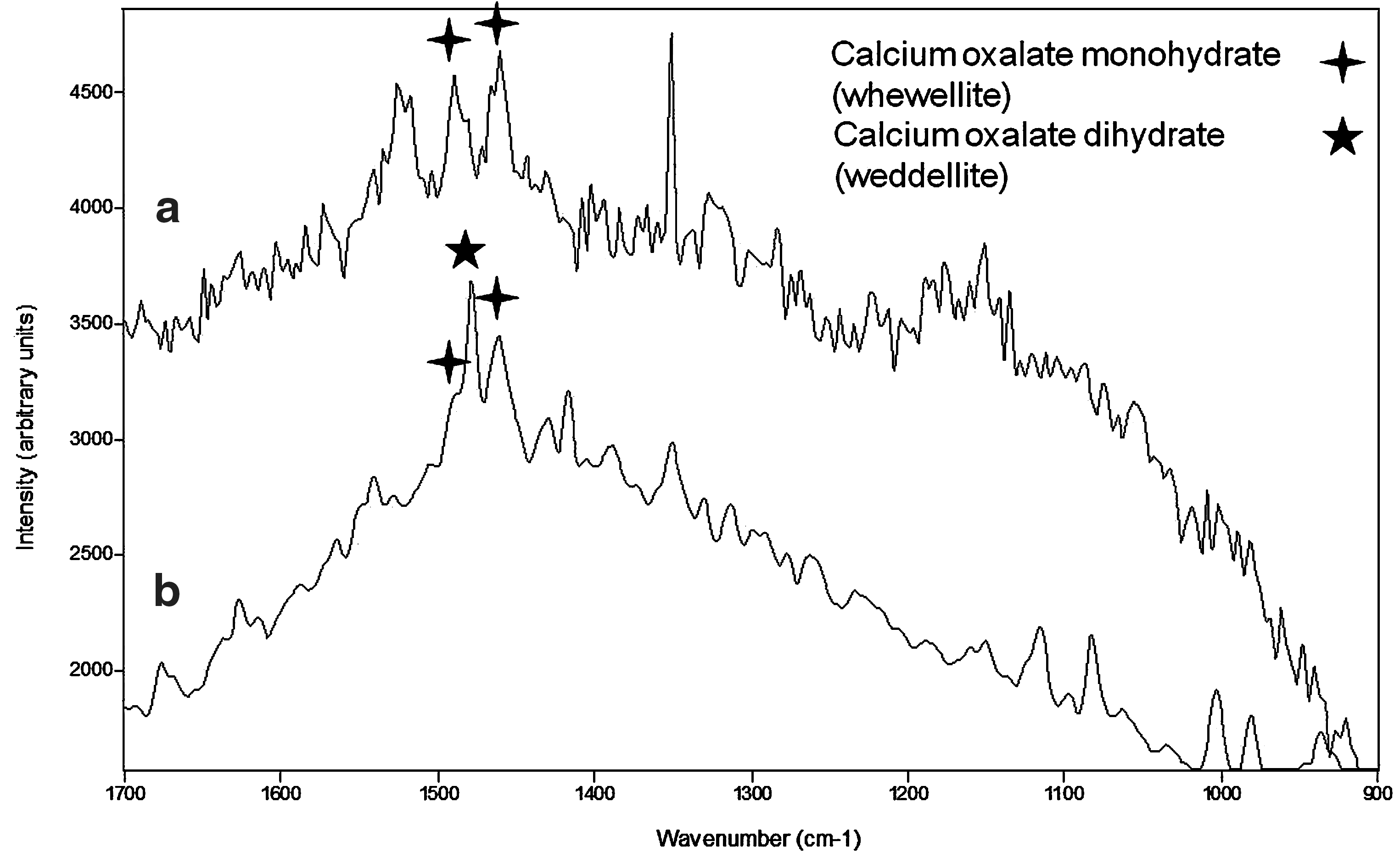
Raman bands identifying calcium oxalates whewellite and weddellite in Squamarina lentigera (Spectrum a) and Lepraria crassissima (Spectrum b).
Other pigments with an important protective role against photostress and oxidation were also found in the lichen communities analyzed in this work (Table 1). Specifically, rhizocarpic acid (antioxidant pigment that acts as a free energy quenching molecule; Wynn-Williams and Edwards, 2000; Jorge-Villar et al., 2006) and usnic acid (UV-radiation screening pigment; Jorge-Villar et al., 2006) could be present in both S. lentigera and D. diacapsis. S. lentigera may also yield parietin, whereas D. diacapsis could produce emodin, both compounds acting as UV-radiation screening.
Lepraria crassissima seems to show the most varied protective strategies, not only with regard to the quantity and variety of oxalates but also in the type of biopigments, since calycin and fumarprotocetraric acid (UV-radiation screening pigments) were identified by Raman spectroscopy.
4.2. Relationships between topographic attributes and protective pigment in extremophile organisms
The topographic attributes extracted from the DEM show that lichens and cyanobacteria communities were not randomly distributed but seem to have been associated with some topographic features and microclimates, which is in agreement with findings of other authors (Sancho and Valladares, 1993; Green et al., 1994; Palmqvist and Sundberg, 2000; Gaio-Oliveira et al., 2004; Lázaro et al., 2008). At the same time, the results obtained with the handheld Deltanu Inspector Raman spectrometer suggest that such communities could have adopted different stress-avoidance strategies by producing biological metabolites that would have allowed them to adapt successfully to these microclimates and dominate within specific microniches. The differences among pigments and biomolecules found by Raman spectroscopy in the organisms studied could be related to different metabolic responses to some parameters, such as incident radiation and humidity, as well as their varied ability to achieve hydration.
The cyanobacteria were predominantly located in areas with the highest global and direct radiation levels (Table 2). Their Raman spectra show evidence of effective UV-screening compounds, such as MAAs with maximal UV absorption in the 310–360 nm range (Edwards et al., 2005). Increased MAA concentrations associated with increases in UV flux have been observed in organisms directly in the field (Cockell and Knowland, 1999). This type of screening compound over other response data demonstrates the importance of elucidating the complete range of UV responses in a given organism. Therefore, although wide surveys of UV screening compounds in nature can be of initial value, interpretation of the data in the ecological context must be undertaken with great care. On the other hand, MAAs may have other physiological functions as well as UV screening; for example, cyanobacteria in some high-salt environments, where MAAs with an internal cell concentration of 98 mmol L−1 have been found to be produced in response to increasing salt concentrations. In this case, they are presumed to have a role in balancing the osmotic pressure inside the cells to counteract water loss (Oren, 1997).
Diploschistes diacapsis and S. lentigera lichens can also exist in a variety of microhabitats and are able to colonize sites with comparatively high solar radiation (Table 2). In contrast, L. crassissima is found mainly on north-facing slopes, where solar radiation is at its lowest (Table 2). Despite these differences, all lichen species analyzed developed pigments with a photoprotective role. Nevertheless, it is worth mentioning that D. diacapsis and S. lentigera produced the highest number of pigments with UVB (e.g., usnic acid, emodin, parietin, beta-carotene) as well as with UVC radiation protection (e.g., beta-carotene, rhizocarpic acid, usnic acid, parietin), the latter the most energetic and most harmful radiation. This could suggest that, although individually these pigments could not protect the lichens from the complete range of UV radiation, together they may act as an effective UV radiation screen, since the range that each one covers overlaps with that of the others. Since the microhabitats colonized by the specific lichens are more exposed to direct solar radiation, it is possible that this mechanism has been adopted as a survival strategy. This way they have succeeded in developing a comparatively better adaptive capacity over other lichen species, such as L. crassissima. Nevertheless, it is interesting to note that although L. crassissima is still only able to colonize those areas with little solar radiation, it also showed UV-radiation screening pigments, which could be explained by the relatively high rates of diffuse radiation achieved in areas usually colonized by this community (Table 2).
The different lichen specimens studied here seem to colonize microniches with different water availability (different contributing areas and wetness; Table 2), which suggests that they could have different moisture requirements and different hydration strategies. In this regard, notice that Raman spectra of L. crassissima, which colonize habitats with the largest water-contributing areas and lowest insolation rates (Table 2), showed the highest concentration of mono- and especially dihydrate calcium oxalate (Table 1, Fig. 3). Previous studies of lichen colonization in different environments have shown that the presence of sufficient available moisture encourages the formation of dihydrate calcium oxalate (Holder et al., 2000; Edwards, 2007). Therefore, the preferential formation of weddellite in L. crassissima may be related with the higher availability of moisture in the microhabitats usually occupied by this species. The use of weddellite by L. crassissima communities in the Tabernas Desert could be associated with water storage strategies and tolerance in this desiccated habitat, as also reported by other authors (Edwards et al., 1997, 2003d; Seaward and Edwards, 1997; Holder et al., 2000; Jorge-Villar et al., 2004, 2005a, 2006; Jorge-Villar and Edwards, 2010). In contrast, D. diacapsis and S. Lentigera, which cover locations that receive higher global and direct solar radiation and lower moisture indices than for L. crassissima, showed comparatively lower concentrations of calcium oxalate monohydrate in their Raman spectra (Table 2) and could not, therefore, be contributing to the formation of calcium oxalate dihydrate. In this case, both lichens may have developed other hydration mechanisms.
5. Conclusions
We have undertaken a study of the distribution of three extremophile lichens and a bacterial community in the Tabernas Desert, southeastern Spain, and analyzed topography-related environmental factors of their habitats, such as direct and diffuse incident solar radiation and water availability. We employed a handheld Raman spectrometer to identify oxalates and protective pigments produced by those organisms, and GIS methods to extract the DEM-related parameters.
The evidence from this study suggests that the specific field distribution of lichens and cyanobacteria is related to the topography. Each community is adapted to specific environmental conditions of insolation and wetness, as reflected in their adaptive strategies, such as the protective pigment production or the presence of calcium oxalates. Raman spectroscopy data show that D. diacapsis and S. lentigera lichen communities, which are exposed to the highest rates of incident solar radiation, produce a high number of protective pigments that could play complementary roles. However, L. crassissima, which colonizes areas with lower insolation rates, generates fewer protective pigments. The cyanobacterial community has developed very different strategies from those of the lichens, which could be related to the severe environmental parameters, since they are the first colonizers on soils and withstand the most hazardous conditions.
Our data also reflect agreement between humidity parameters and the presence of calcium oxalate monohydrate and/or calcium oxalate dihydrate. Where humidity was higher, lichens produced both oxalates, whereas with less humidity only calcium oxalate monohydrate was produced. Although it looks like lichens had enough water for survival under the wettest conditions, L. crassissima might require extra water storage and, therefore, produces weddellite to supply it.
Surprisingly, there were no clear Raman signals when the analyses were performed with our portable Raman spectrometer on the four extremophile organisms sampled during a long drought. It is obvious that this lack of results cannot be interpreted as absence of life. There could be several reasons for this phenomenon. First, the design and construction of the miniaturized instrument could have a negative impact on the resolution of the spectrometer and therefore on its capacity to obtain useful results. The increase in reflectivity of dry versus wet specimens could reduce the sensitivity of the instrument even further. It is also possible that reversible changes in the chemical makeup of the organisms under desiccation, for example, desiccation-induced conformational changes in a pigment-protein complex, could be responsible for the lack of signals. This issue is of particular relevance to extraplanetary exploration, where miniaturized analytical instruments could be utilized for the study of extremophile organisms living under severe conditions. A better understanding of Raman spectrometer limitations is, therefore, a prerequisite for employing such instruments in extraplanetary missions. Although this is already being taken into account, our results stress the need for employing a sample manipulation protocol so that the selected specimens are analyzed under optimal environmental conditions as well as handling parameters, namely, humidity, temperature, or light.
Footnotes
Acknowledgments
The authors wish to thank the GEOCARBO (Ref. RNM-3721) and the COSTRAS (Ref. RNM-3614) projects funded by the regional government of Andalucía (Spain); the BACARCOS project (Ref. CGL2011-29429) and CARBORAD (CGL2011-27493) projects funded by the Spanish Ministry of Science and Innovation, the European Regional Development Fund (ERDF), and the Juan de la Cierva Fellowship (Ref. 2008-39669).
Abbreviations
ARE, contributing area; BSC, biological soil crust; DEM, digital elevation model; GIS, Geographic Information System; LSF, slope length factor; MAA, mycosporine-like amino acid; TRMI, Topographic Relative Moisture Index; W, wetness index.