
Abstract
In the space experiment “Molecular adaptation strategies of microorganisms to different space and planetary UV climate conditions” (ADAPT), bacterial endospores of the highly UV-resistant Bacillus subtilis strain MW01 were exposed to low-Earth orbit (LEO) and simulated martian surface conditions for 559 days on board the European Space Agency's exposure facility EXPOSE-E, mounted outside the International Space Station. The survival of B. subtilis MW01 spores from both assays (LEO and simulated martian conditions) was determined by a colony-formation assay after retrieval. It was clearly shown that solar extraterrestrial UV radiation (λ≥110 nm) as well as the martian UV spectrum (λ≥200 nm) was the most deleterious factor applied; in some samples only a few spore survivors were recovered from B. subtilis MW01 spores exposed in monolayers. However, if shielded from solar irradiation, about 8% of MW01 spores survived in LEO conditions, and 100% survived in simulated martian conditions, compared to the laboratory controls. The results demonstrate the effect of shielding against the high inactivation potential of extraterrestrial solar UV radiation, which limits the chances of survival of even the highly UV-resistant strain of B. subtilis MW01 in the harsh environments of outer space and the martian surface. Key Words: Bacterial spores—Radiation resistance—Adaptation—LEO—Mars. Astrobiology 12, 498–507.
1. Introduction
ADAPT I: Bacillus subtilis MW01, a UV pre-adapted derivate of B. subtilis 168 (Wassmann et al., 2010), a ubiquitous spore-forming soil bacterium that is able to withstand desiccation as well as high levels of solar UV radiation when in its dormant stage as a spore;
ADAPT II: Halococcus dombrowskii, an Archaea isolate from a Perm-Triassic Alpine salt deposit;
ADAPT III: an epilithic microbial community that is naturally exposed to high levels of solar UV radiation, which was augmented with akinetes of Anabaena cylindrica and vegetative cells of Nostoc commune and Chroococcidiopsis (Cockell et al., 2011).
In this communication, we exclusively present the results of ADAPT I obtained with spores of Bacillus subtilis MW01, the flight experiment, the parallel mission ground reference (MGR) experiment and preflight experiment verification tests (EVTs), and an experiment sequence test (EST). Spores of the parental strain B. subtilis 168 were used in various space experiments and have been known to survive very long periods of time (up to 6 years) in low-Earth orbit (LEO) (Horneck et al., 1994, 2001, 2010). The environment in LEO is hostile to all life-forms. It contains a concatenation of different stressors, such as ionizing radiation, desiccation, high UV radiation levels, and extreme temperature fluctuations. The response of organisms to these stressors has been the focus of numerous studies for a number of decades (reviewed by Horneck et al., 2010). Among the mentioned factors, solar extraterrestrial UV radiation is the most harmful parameter to the organisms used in those space experiments. It has several deleterious effects at the cellular level that cause a variety of lesions, mainly DNA base damage, DNA strand breaks, DNA-protein cross-links, membrane peroxidation, and enzyme inactivation (Cadet et al., 2005; Goosen and Moolenaar, 2008, and references therein).
The martian UV radiation climate is significantly different from that on Earth and in LEO due to the composition and pressure of the martian atmosphere. Compared to LEO, the energy-rich vacuum-UV range (<200 nm) is absorbed by the martian atmosphere and does not penetrate to the surface of Mars. In the experiment ADAPT I, spores of B. subtilis MW01 were exposed to both conditions, those of outer space in LEO and simulated martian surface conditions. The hypothesis tested experimentally in the experiment ADAPT I and the corresponding precursory studies was whether longer-lasting selective pressure by UV radiation of different quality results in a higher UV resistance as well as in a higher resistance against the simultaneous action of further “extreme” environmental factors (e.g., vacuum or low pressure and cosmic radiation) that exist in space or on other planets like Mars.
2. Materials and Methods
2.1. Bacterial strains
Three Bacillus subtilis 168 strains were used in this work; the ancestral (A), the UV-adapted (MW01), and the identically cultivated UV-unexposed control strain (DE69). Strain MW01 was isolated from a short-term evolution experiment, in which B. subtilis 168 cells were cultivated for 700 generations under periodical polychromatic UV (200–400 nm) applied during the stationary growth phase of serially propagated batch cultures (Wassmann et al., 2010). Details on strain origin and the experimental evolution setup have been previously described in detail by Wassmann et al. (2010). All three strains are isogenic to the B. subtilis trpC2 strain 168 (DSM 402), which was originally obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany).
2.2. Cultivation and sporulation
Spores were obtained by cultivation under vigorous aeration in liquid Schaeffer sporulation medium (Schaeffer et al., 1965), and were purified and stored as described in detail by Nicholson and Setlow (1990) and Moeller et al. (2006).
2.3. Sample preparation for the flight and mission ground reference experiment
Spores were air-dried as (arithmetically proved) monolayers on MgF2 or quartz (Suprasil) discs of 11.9 mm diameter and 1 mm thickness. From aqueous spore suspensions, 5×107 spores (except for EVTs where 1×107 spores were used, see below) were transferred to the respective surface such that they spread and air-dried uniformly (as described in detail in Puskeppeleit et al., 1992). Immediately after spotting, all samples were dried under ambient laboratory conditions (temperature 20±2°C and relative humidity 33±5%). The homogeneity of the spore layers was routinely checked by microscopic analysis. The sample discs were finally stored under laboratory conditions until further usage. It has to be mentioned here that, due to experimental constraints and limitation of the EXPOSE-E hardware, only spores of the B. subtilis strain MW01 were tested in the flight and mission ground reference experiment.
Flight and MGR samples were prepared as stacks of sample discs in three layers (Fig. 1A); only the preflight experiments were performed with single sample discs. Depending on the exposure conditions, discs of two different materials were used that differed in their optical transmission characteristics: MgF2 for the exposure of the samples to the full spectrum solar extraterrestrial UV radiation (λ>110 nm) in LEO, and quartz for exposure to simulated martian conditions (λ>200 nm) and all preflight laboratory tests. Two identical sets of samples were provided: one set for the flight experiment and one set for the MGR experiment. Both sets consisted of 32 stacks, which resulted in a total of 96 single samples per set. The stacks were accommodated in two sample trays of the EXPOSE-E facility (for details see Rabbow et al., 2012), depending on the requested exposure conditions:
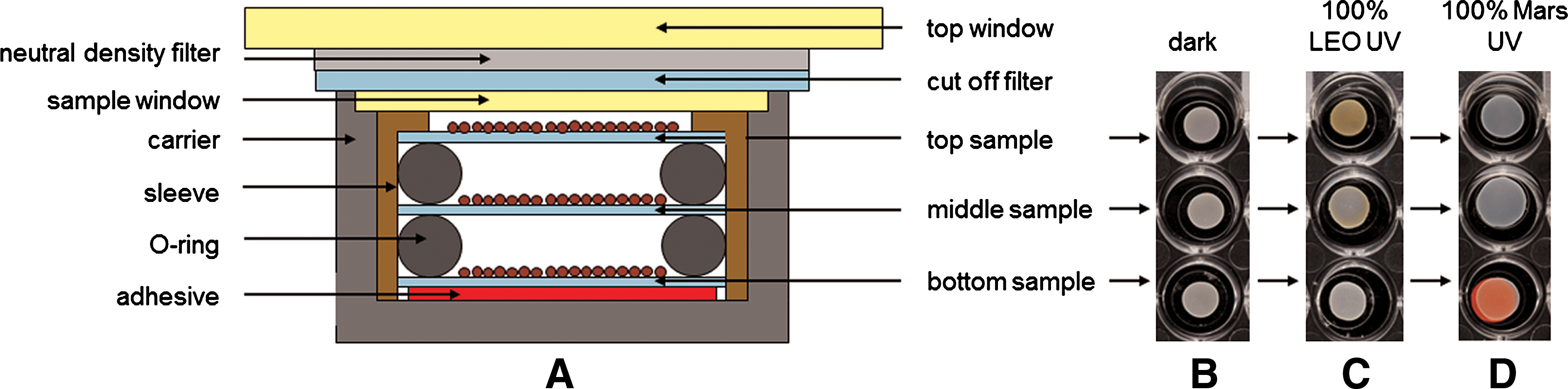
(
• Tray I was vented to space conditions (vacuum) and covered by a MgF2 window that allowed transmission of solar electromagnetic radiation of λ>110 nm. It accommodated 16 sample stacks with MgF2 as sample carrier. Four of these stacks were exposed to non-attenuated UV radiation of λ>110 (100% UV), and four to the same UV range but attenuated by about 3 orders of magnitude by using a 0.1% MgF2 neutral density filter. Eight stacks were dark controls, which were located in corresponding positions underneath the UV-exposed stacks.
• Tray II was hermetically closed, filled with a 103 Pa CO2 atmosphere, and covered by a quartz window with transmission of solar electromagnetic radiation of λ>200 nm. Here, the conditions in LEO were used to simulate martian surface conditions (Rabbow et al., 2012). Tray II accommodated the residual 16 stacks of samples on quartz carriers. Four of these stacks were exposed to 100% UV radiation of λ>200 nm and four to 0.1% UV radiation of λ>200 nm with additional quartz neutral density filters. Again, eight stacks were dark controls located underneath the UV-exposed stacks.
The total duration of exposure was 1.5 years (Rabbow et al., 2012 in this issue).
An identical arrangement of sample stacks in two trays was used for the MGR experiment. During the MGR, the environmental data of the flight unit (e.g., temperature profile, vacuum exposure, UV radiation; Rabbow et al., 2012) were followed with use of the Planetary and Space Simulation facilities (
2.4. Preflight UV resistance tests
Samples were exposed to polychromatic (200–400 nm) UV irradiation emitted by a sunlight simulator (SOL2, Dr. Hönle AG, Berlin, Germany). Irradiation was carried out under conditions as described previously (Wassmann et al., 2010).
2.5. Preflight heavy ion radiation and X-ray resistance tests
Spore exposure to accelerated heavy Ar (500 MeV/n; 90 keV/μm) and Fe (500 MeV/n; 200 keV/μm) ions was performed with the Heavy Ion Medical Accelerator (HIMAC) at the National Institute for Radiological Sciences (Chiba, Japan), and the X-ray irradiation (150 keV/19 mA) was performed at the DLR as described previously (Moeller et al., 2010).
2.6. Ground-based experiment verification tests and experiment sequence test
Several ground-based EVTs and an EST were performed at the DLR. Tests included exposure to single or combined space-relevant parameters under simulated space or martian conditions. The parameters of the exposure facilities have been described elsewhere (Onofri et al., 2008; Rabbow et al., 2009; Billi et al., 2011).
2.7. Survival determination
To recover spores after exposure, samples were transferred into glass vials. Spores were abraded from discs in water with sterile quartz sand. Serial 10-fold dilutions were plated on solid nutrient broth medium. After overnight growth at 37°C the number of colony-forming units (CFU) was counted.
2.8. UV dosimetry and transmission measurements
To determine how much UV radiation was encountered by middle and bottom samples, the transmission of several samples was determined. Wavelength scans (200–400 nm) were performed (Hitachi U-3200 double monochromator spectrophotometer) on spore-covered and blank MgF2 and quartz discs, respectively. Transmission values were offset against fluences based on model calculations performed by RedShift Design and Engineering BVBA, Belgium (T. Beuselinck, C. Van Bavinchove), which were provided to the investigators by ESA.
2.9. Numerical and statistical analysis
The surviving fraction of the spores was determined from the quotient N/N 0, with N=the number of CFU of the irradiated sample and N 0 that of the non-irradiated controls. Data are reported as D10 (for X-rays) and F10 (for UV radiation) values, dose and fluence, respectively, lethal for 90% of the initial population, according to Moeller et al. (2007). Each experiment was repeated at least three times, and the data shown are expressed as averages±standard deviations. The treated cells and spores were compared statistically by using the Student t test. Values were analyzed in multigroup pairwise combinations, and differences with P values of ≤ 0.05 were considered statistically significant (Moeller et al., 2005).
3. Results
3.1. Preflight experiments
3.1.1. Ionizing and UV radiation survival
Inactivation kinetics of air-dried wild-type (168) and UV-adapted (MW01) B. subtilis spores were obtained in response to different qualities of radiation relevant for space exposure. Spores of both strains were almost equally affected by all tested qualities of radiation (Fig. 2). Nearly exponential survival curves were obtained in response to ionizing radiation (Fig. 2A and 2B), provided in the form of heavy ions and X-rays. Best-fit curves were used to calculate D10 values for statistical comparison (Table 1). D10 values for strain MW01 were slightly decreased (max. 5%) relative to the corresponding values of strain 168. F10 values obtained in response to polychromatic UV radiation were 12.4±1.3 kJ/m2 for strain 168 and 14.1±1.2 kJ/m2 for MW01, showing moderate (about 13%) increased resistance for strain MW01 compared to strain 168. In contrast to ionizing radiation, in response to UV, inactivation curves exhibited a downward concave shape (Fig. 2C), which indicates biphasic inactivation kinetics. On average, 0.1% of the dried spores seemed to be at least partly shielded by upper spores, which influenced their accessibility to UV, as reported previously by Horneck et al. (1994, 2001), but not to the traversing beam of ionizing radiation.
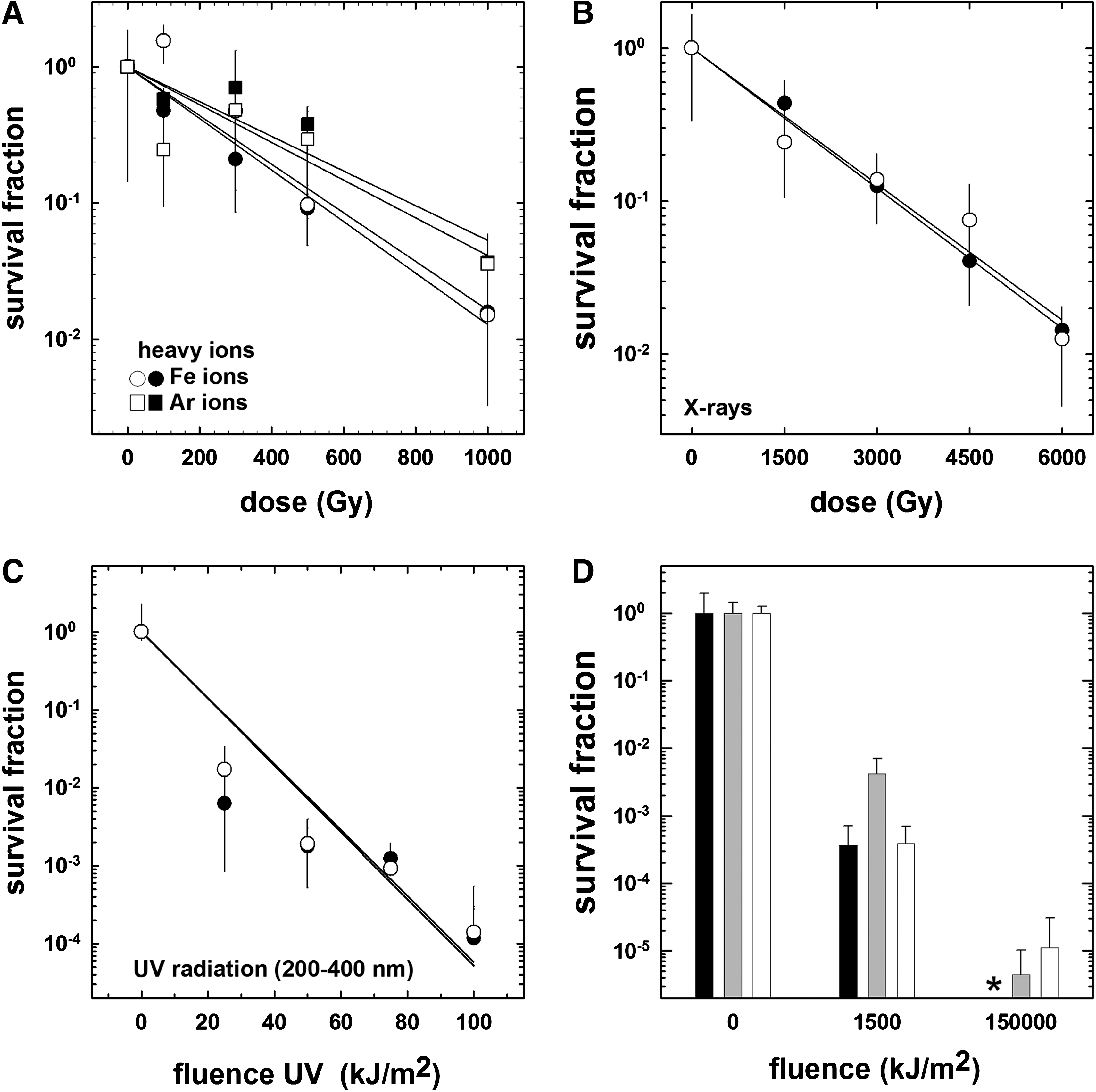
Results of the preflight experiments of spores of Bacillus subtilis strains 168 (filled symbols) and MW01 (open symbols). Spores of both strains were irradiated, and survival curves were obtained in response to (
Data are derived from Fig. 2A–2D.
3.1.2. EVTs and EST
MW01 spore viability after EVTs and EST was determined by cultural methods. EVTs and EST were performed at the Planetary and Space Simulation facilities at the DLR, Cologne. Figure 2D shows the results of exposure of dried spores of strain MW01 to polychromatic UV in vacuum and in simulated martian atmosphere, compared to corresponding dark controls. To prevent shielding by overlaying spores—an issue described above—and to increase the homogeneity of spore distribution within the samples, the number of spores per disc for the EVTs was reduced to 1×107. Irradiation of the samples in vacuum with 1.5×103 kJ/m2 UV led on average to an increased inactivation by 3.5 orders of magnitude, whereas no survival was detected in response to the maximum applied fluence of 1.5×105 kJ/m2. For the EST, the amount of spores was increased to 5×107 spores per disc, to determine the limit of detection even when paying the costs of using imperfect monolayers. Consequently, irradiation with 1.5×103 kJ/m2 UV in vacuum led to less inactivation, namely, by 2.5 orders of magnitude compared to EVTs, and we were able to detect some survivors even after exposure to 1.5×105 kJ/m2. Survival was reduced by 5.5 orders of magnitude, which is very close to our threshold of detection (6 orders of magnitude). Irradiation in simulated martian atmosphere led to a relatively high inactivation of 3.5 orders of magnitude in response to 1.5×103 kJ/m2 and of 5 orders of magnitude after exposure to 1.5×105 kJ/m2.
3.2. ADAPT I: flight and mission ground reference experiment
3.2.1. Spore recovery and sample analyses
Visual inspection of the samples after retrieval revealed obvious alterations in color (Fig. 1B–1D). Figure 1C shows that the color of top samples, exposed to 100% LEO UV radiation, turned from off-white to brownish. Middle samples showed still-visible color changes toward brownish, whereas bottom samples' color was visually not altered. This effect was exclusively observed for samples exposed to the unattenuated complete range of extraterrestrial UV in LEO of wavelengths λ≥110 nm. A second alteration was recognized during the analysis of the samples. After approximately two and a half years in the dry state, spores adhered to each other and to the sample carriers (which has never been reported in previous B. subtilis spore space experiments). Hence, spore recovery was extremely impeded. This effect was observed for samples exposed to all conditions tested, even for the laboratory controls, which ruled out a space-mediated effect. Furthermore, adherence was observed for both types of sample carriers, MgF2 and quartz discs. With the recovery method described above, spores could be resuspended, but clusters in the suspensions were still visible by microscopic observations.
3.2.2. Loss of transmission in the sample stacks and determination of the shadowing effect
Fluences provided by ESA (RedShift Report 2, 2011) were modeled for a position in the center of top sample sites. To calculate the fluences as to which middle and bottom samples were exposed during the mission, the transmission in the range of 200–400 nm wavelengths of several spore-covered discs of both materials was determined. Measurements showed that the irradiance was reduced by approximately 30±2% per passage through a spore-covered MgF2 disc and by 45±5% through an equally coated quartz disc. The observed loss of transmission was constant over the monitored spectrum of wavelengths. The resulting fluences for samples accommodated at different positions in the stacks are listed in Tables 2 and 3. The above-mentioned color changes of some of the samples had no influence on the individual transmission values.
If survival value was “0,” it was below the threshold level of detection (of >5×107 CFU).
If survival value was “0,” it was below the threshold level of detection (of >5×107 CFU).
3.2.3. Laboratory control samples
Spores from laboratory control samples also exhibited the above-mentioned adherence to the discs and clumped when they were resuspended. Consequently, the recovery, as determined via CFU counts, was significantly decreased. The average numbers of CFU obtained were 1.2±0.4×106 spores per MgF2 discs and 8.2±1.6×105 spores per quartz disc. With respect to the number of spores originally spread on discs of both materials, recovery rates were at 2.4±0.8% (MgF2) and 1.7±0.6% (quartz), respectively. Recovery controls performed directly after sample preparation and on untreated references from preflight experiments, stored in the dry state for up to 1 month, revealed recovery rates above 90% (data not shown). The respective laboratory controls were used as reference (N 0) for survival determination of flight and MGR samples.
3.2.4. ADAPT I—Flight experiment
Survival data from the flight experiment are summarized in Table 2. With respect to the corresponding laboratory controls, UV-shielded samples, which were located directly beneath the corresponding UV-exposed samples and subjected to LEO vacuum during the flight experiment, were inactivated by one order of magnitude. The combined exposure to 100% LEO UV irradiance and vacuum led to a total inactivation of all spores in the top (3.6×105 kJ/m2) and middle (2.5×105 kJ/m2) layers. Survival in the bottom layer (1.7×105 kJ/m2) was reduced by 4.5 orders of magnitude. The samples beneath the 0.1% transmission filters (max. fluence 3.5×102 kJ/m2) were less affected. Spore survival was reduced to 1–5% in those stacks. MW01 spores exposed to simulated martian conditions survived better than those under LEO conditions. Dark samples were almost not affected compared to the laboratory controls. Exposure to 100% UV λ>200 nm (3.0×105 kJ/m2) at the top sample site resulted in an inactivation by 1–2 orders of magnitude. Similar inactivation was observed for samples in the middle layer (1.6×105 kJ/m2). Paradoxically, samples in the bottom layer, which were exposed to the lowest fluence (9.0×104 kJ/m2), showed the highest inactivation. Survival was reduced by almost 3 orders of magnitude. Samples beneath the 0.1% transmission filters and kept in simulated martian atmosphere showed the highest survival among the UV-exposed samples in this experiment. Top samples (2.5×102 kJ/m2) were inactivated to about 40%, survival, whereas middle (1.4×102 kJ/m2) and bottom samples (7.5×101 kJ/m2) were inactivated to approximately 10%. Curiously again, more survivors were recovered from top samples than from the shadowed samples below, a finding that should be due to problems in spore recovery described above and has to be discussed.
3.2.5. ADAPT I—mission ground reference
With respect to the corresponding laboratory controls, simulated space vacuum and martian atmosphere, together with the temperature simulation, had minor effects on spore survival in the MGR. Survival of dark samples was reduced to (34±16)% for samples kept in simulated space vacuum and showed a 3 times higher survival as compared to the flight experiment. In the simulated martian atmosphere, (82±28)% of the spores survived. In Table 3, the survival of the spores of the MGR experiment is summarized. No spores survived the exposure to simulated LEO conditions with 100% UV (wavelengths λ>200 nm) radiation (fluences of 4.4×105 kJ/m2 for the top layer), neither in the top nor in the middle or bottom layer. Survivors were only found in the bottom layer of the stack covered by 0.1% transmission filters, which were exposed to approximately 3.0×102 kJ/m2 UV (Table 3). Samples at this position were inactivated by 2[[h]] orders of magnitude. No survivors were detected at the top and middle layers of those stacks. Similar to the flight experiment, exposure to simulated martian conditions was less harmful to MW01 spores (Table 3). Survivors were found at the bottom layer of stacks exposed to 100% UV λ>200 nm, which resulted in an accumulated fluence of 1.1×105 kJ/m2. Survival there was reduced by 3[[h]] orders of magnitude. Under protection of 0.1% transmission filters, top-layer samples were exposed to an accumulated fluence of 4.8×102 kJ/m2, which led to an inactivation by a factor of four orders of magnitude. Samples in the middle (2.6×102 kJ/m2) and bottom layers (1.4×102 kJ/m2) showed survival of 5±3% and 16±9%, respectively.
4. Discussion
The Bacillus subtilis strain MW01 used in the ADAPT I experiment was isolated from a short-term laboratory evolution experiment (Wassmann et al., 2010) that was carried out in the framework of, and as a precursory study to, this space exposure experiment. MW01 is a UV-adapted descendant of the Gram-positive model organism B. subtilis 168. Spores of this ancestral strain have a long history as models in astrobiology. Several space exposure experiments and ground-based studies have certified its potential to survive a sojourn in space (Nicholson et al., 2000; Horneck et al., 2008, 2010; Nicholson, 2009). Spores of B. subtilis hold the record for space survival after spending 6 years on NASA's Long Duration Exposure Facility (LDEF) (Horneck et al., 1994). The B. subtilis strain MW01 is a very new astrobiological test system (with a significantly higher UV resistance in the vegetative form), which was characterized to some extent before it was used in the ADAPT space experiment (Wassmann et al., 2010). For this purpose, multiple preflight irradiation experiments were performed, always in parallel with the space-proved and well-characterized ancestor. The data presented in Fig. 2 show no dramatic difference in response to different radiation qualities, including heavy ions, X-rays, and polychromatic UV. In summary, spores of strain MW01 have no clear advantages or disadvantages concerning survival in response to solar UV radiation, space vacuum, or martian atmosphere; if a small fraction of the MW01 spores were to sustain the challenges of an extraterrestrial environment and prove be of astrobiological interest in the Solar System (e.g., Mars), their superiority in the vegetative state compared to those of the “space veteran” B. subtilis 168 would be proved (Wassmann et al., 2011). In this regard, MW01 offers a high potential as a candidate for future astrobiological studies on the adaptability toward potential active microbial life and survival on Mars (Wassmann et al., 2010). When comparing the resistance of the B. subtilis MW01 spores (tested on board the EXPOSE-E platform) and spores of B. subtilis flown on NASA's LDEF, the “Exobiological Unit” of the Exobiology Radiation Assembly on board the European Retrievable Carrier, and the Biopan facility of ESA on board the Russian Earth-orbiting Foton satellite, spores of the MW01 strain (exposed as monolayers) were statistically not significantly more or less resistant to space conditions than the respective test wild-type spores of B. subtilis (Horneck et al., 1994, 1995, 2001, 2010), which is in good agreement with the preflight tests.
Within the preflight experiments, the environmental and technical requirements were defined for the space experiment. Concerning radiation, 1.5×105 kJ/m2 was the maximum fluence of polychromatic UV applied during EVTs and EST, since it corresponds to the upper limit of UV irradiance, which was expected by ESA to accumulate during the planned space exposure. All sample preparations and filter configurations, which covered some of the samples (Fig. 1A; Rabbow et al., 2012), were based on this assumption. Unfortunately, the actual accumulated fluence of 3.6×105 kJ/m2 was more than 2 times higher than expected. Among other reasons, this was due to the fact that the exposure time in space was extended to 1.5 times the originally conceived time of exposure. As a consequence, samples exposed to the highest fluences were completely inactivated (Table 2). Originally, it was planned to use the setup of stacked samples to model the shielding effect of the upper (mono-) layers of spores on the spores in the layers below. Since the stacks provided no or just one data point for three out of four tested UV exposure conditions, this goal could not be achieved for the MGR samples, nor could it be achieved, due to clustering, for the flight samples. However, the stacks and the associated shielding effect enabled us to get some data points from those high fluences.
Significant differences in inactivation between the MGR and the flight experiment (Table 2) were observed. The inactivation was orders of magnitude higher for most samples in the MGR compared to the respective samples in the flight experiment. However, the contrary was observed for other experiments on EXPOSE-E. In ADAPT III, MGR samples showed higher survival compared to flight samples. For ADAPT I, the differences in inactivation were due to different fluences encountered by the respective samples. Samples in the MGR were irradiated with higher fluences than those accumulated during the 18-month space exposure (Tables 2 and 3). Local circumstances led to a significant reduced irradiance at ADAPT I sample sites (RedShift Report 2, 2011), compared to a virtual sample site in the middle of the respective compartments (RedShift Report 1, 2010). Among all samples accommodated in trays I and II of the EXPOSE-E facility, ADAPT I samples were situated closest to the EuTEF platform and because of this were partly shadowed (RedShift Report 2, 2011). Actually, the lowest fluence irradiance over the EXPOSE-E surface was achieved at compartment 2-1 where our “Mars samples” were accommodated (Rabbow et al., 2012). Since the sample site–specific fluences were not yet available when the MGR was performed, fluences as calculated in the 2010 report were applied on ADAPT I samples.
The higher survival of spores in dark controls compared to UV exposure, both in ground-simulation and space-exposure experiments, is consistent with data obtained from previous studies (Horneck et al., 1994; Rettberg et al., 2002; Schuerger et al., 2003, 2006a, 2006b). The greater survival of spores exposed to simulated martian conditions compared to LEO parameters could be explained by the different UV spectra acting on samples in the flight experiment (Newcombe et al., 2005; Osman et al., 2008; Horneck et al., 2010). Samples exposed to LEO encountered a much broader spectrum of UV radiation, including the particularly mutagenic and lethal short-wave regions of UVC and vacuum UV. For the MGR, this explanation is insufficient, since for technical reasons only wavelengths of λ>200 nm were applied. However, to compensate for the loss observed at shorter wavelengths, simulated LEO samples were exposed to a higher flux in the range of λ from 200 to 400 nm, relative to the flight samples, which resulted in the same total fluence. Another possible explanation for greater survival under simulated martian conditions is given by the selection history of strain MW01. The results obtained from the experiment presented here show that 200–400 nm UV radiation was the major survival-limiting factor in the chosen combination of Mars-attributed parameters, and MW01 evolved under the selective pressure of this particular UV spectrum (Wassmann et al., 2010).
Particularly surprising were the results concerning the inactivation in different layers of the stacked samples from the flight experiment. In many cases, the ranking of survival did not fit with the corresponding fluences. Here, survival was defined as the ability of a spore to germinate and form colonies on solid media. Since spores were clustered, the enumeration of these survivors was hardly affected. Concerning general survival determination, ionizing radiation had only minor impact on the spores during the exposure in LEO. The average radiation dose in LEO amounts to 100–800 mGy/year, depending on the inclination, altitude, and number of passages through the South Atlantic Anomaly (Berger, 2003). The accumulated dose over the 1.5-year mission was at the lower end of this range and resulted in about 0.18 Gy as determined by the evaluation of ionizing radiation dosimeters accommodated below dark sample sides in the EXPOSE-E facility (Berger et al., 2012). In relation to the D10-values determined from preflight experiments (Table 1), the dose was negligible. Nevertheless, single spores could have been affected by direct hits of heavy ions (Woese, 1958; Moeller et al., 2010, and references therein). Also negligible was the effect of the mission temperature that fluctuated between +43°C as the upper temperature and −22°C as the lowest temperature (Rabbow et al., 2012). Spores of B. subtilis should neither be affected by temperature fluctuations in this range nor by the one-time high-temperature peak (61°C for less than 1 hour; see Rabbow et al., 2012).
The space experiment ADAPT and its precursory studies were performed to test the hypothesis that longer-lasting selective pressure by UV radiation of different quality results in a higher UV resistance and a higher resistance against the simultaneous effects of further “extreme” environmental factors that exist in space or on other planets, such as vacuum or cosmic radiation. Within the precursory ground-based studies (Wassmann et al., 2010, 2011), this hypothesis was generally confirmed for vegetative cells of our strain B. subtilis MW01. A selection against reoccurring polychromatic UV irradiation led to an increased UV resistance as well as an enhanced tolerance toward other environmental stressors (Wassmann et al., 2010). Within the precursory ground-based studies to ADAPT I (Wassmann et al., 2010, 2011), this hypothesis was generally confirmed for vegetative cells of strain B. subtilis MW01. That MW01 spores have the potential to survive a voyage through open space was shown in this experiment. The survival data of UV-protected MW01 spores (∼8%) fit very well with the collective survival data (∼10–50%) of the other spore experiments on the EXPOSE-E mission (Horneck et al., 2012; Moeller et al., 2012; Nicholson et al., 2012). And, a UV-shielded stay on Mars would potentially not affect the spores to a significant degree. However, even if the spores of strain MW01 have no clear advantages compared to the parental strain 168 (Horneck et al., 2012), in the event that it can be demonstrated that some of them are able to sustain the challenges of an extraterrestrial environment and, thus, potentially come to reside in a habitable place in the Solar System, their superiority in the vegetative state will have been proved.
Footnotes
Acknowledgments
The authors thank the European Space Agency for the flight opportunity and would like to acknowledge the support of ESA (especially René Demets) and of the MUSC team. This work was conducted as part of the ADAPT experiment on EXPOSE-E, and the results will be included in the Ph.D. thesis of Marko Wassmann. The authors also thank Thomas Berger, Ryuichi Okayasu, and Hisashi Kitamura for their superb support and continual readiness to help during the heavy ion irradiations. We are grateful to Takeshi Murakami and the HIMAC operators for their technical assistance and beam monitoring. This study was supported by the DLR grant DLR-FuE Projekt ISS-Nutzung in der Biodiagnostik, Programm RF-FuW, Teilprogramm 475 to M.W., E.R., R.M., G.R., and P.R.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
DLR, German Aerospace Center; EST, experiment sequence test; EVTs, experiment verification tests; HIMAC, Heavy Ion Medical Accelerator; LDEF, Long Duration Exposure Facility; LEO, low-Earth orbit; MGR, mission ground reference.