
Abstract

Introduction
C
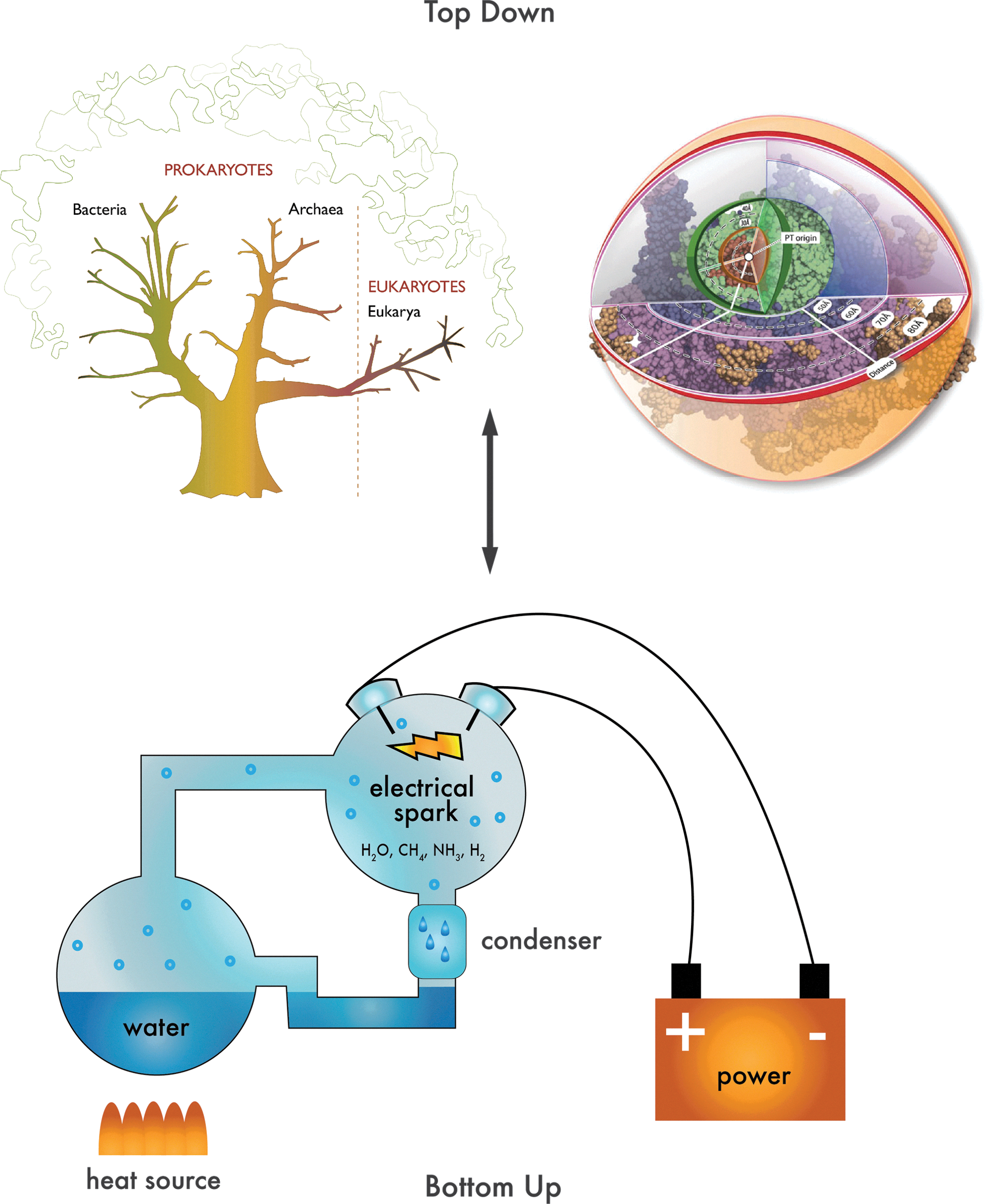
Classic Miller-Urey apparatus (lower panel) used for “bottom up” experiments advanced our understanding of prebiotic chemistry, showing that abiotic process can contribute biomolecule precursors or building blocks. A phylogenetic tree and ribosome onion (upper panel), illustrating phylogenetic and biochemistry contributions to “top down” origin-of-life research.
Origin of Life from the Bottom Up
With Bottom-Up approaches, fundamental principles of chemistry, organic synthesis, transition metal chemistry, and molecular assembly are applied in experimental systems that mimic the chemical conditions of prebiotic Earth. Investigators strive to understand and recapitulate steps in the origin of life, including the formation of precursors, building blocks, polymers, macromolecular assemblies, and catalytic functions.
In the seminal “Bottom-Up” experiments, Miller and Urey (Miller, 1953) simulated the conditions of early Earth and used electric discharge to produce amino acids from simple precursors. Later, Oro showed that adenine, a component of DNA, RNA, and ATP, can be produced from hydrogen cyanide under conditions that mimic early Earth (Oro and Kimball, 1962). In fact, many biological building blocks, including additional nucleic acid bases (Ferris et al., 1968), sugars (Butlerow, 1861; Ricardo et al., 2004), nucleotides (Powner et al., 2009), and a wide variety of amino acids (Johnson et al., 2008; Parker et al., 2011) can be produced under hypothesized prebiotic conditions. Further, many of these biological building blocks are found throughout the abiotic solar system (Schmitt-Kopplin et al., 2010). Life on Earth, seen after approximately 4 billion years of evolution, clearly emerged in an environment with the inherent capacity to produce many of the small molecules required as enzymatic cofactors and for biological polymers (Oparin, 1938). Life continues to synthesize some of the metabolites on which it initially depended, before it exhausted the environmental supply.
Many of the input molecules for biology are readily produced under laboratory conditions that simulate those prior to life's initiation. Much can be learned about pathways to life's origins simply by understanding the features and environment of early Earth, along with principles of chemistry and biochemistry. Advances in analytical chemistry (Johnson et al., 2009), synthetic methods (Beier et al., 1999), molecular recognition (Persil and Hud, 2007), and geology (Anbar, 2008; Hazen and Ferry, 2010) are leading to breakthroughs in Bottom-Up approaches to the origin of life. The impact of new technologies is reflected in the recent reanalysis, by high-performance liquid chromatography and mass spectrometry, of samples from Miller's volcanic spark discharge experiment (Johnson et al., 2008; Parker et al., 2011).
Origin of Life from the Top Down
Properties of extant life are used in “Top-Down” approaches to infer molecules, pathways, structures, and assemblies of ancient life. The Top-Down approach is championed by Woese (2000) and others (Hury et al., 2006; Fox, 2010), including the authors here. The premise is that some extant metabolites (Benner et al., 1989), biochemical systems such as translation (Woese, 2000; Hury et al., 2006; Fox, 2010), and metalloproteins (Martin and Russell, 2003; Wachtershauser, 2006; McGlynn et al., 2009) are linked by sometimes indistinct, but ultimately traceable, threads to early biotic and prebiotic chemical systems. Life's history is maintained in part in small molecule repertoires, chemical transformations, assemblies, polymer backbones and sequences. Extant biological systems are said to contain “fossils” that can be conceptually partitioned between molecular fossils (adenosine, glycine, iron sulfur centers, ribosomal RNA, and some DNA sequences) and process fossils (gluconeogenesis, translation). By mining these fossils, we can understand much about the origin of life.
Molecular Phylogeny
Our view of the origin of life, viewed from the top down, is increasingly penetrating. The rapid growth of genome databases and comparative tools is quickly changing the nature of phylogenetic analysis. The exploding sequence database along with ever more powerful search and alignment algorithms, advances in processing, and data storage is revealing new information about origins, lineage, and biodiversity. The marriage of taxonomic and functional process-based evolutionary analysis is enabling inferences into early and even primordial processes that can in some cases be placed in the context of the geological record. New tools allow development of highly complex models of phylogenetic relationships, along with detailed mapping of evolutionary histories. Ever-larger sequence databases increase the probabilistic power of ancestral reconstruction and resurrection (Benner et al., 2007; Gaucher et al., 2008; Harms and Thornton, 2010). A similar trajectory of the structural database is facilitating development of disciplines of paleo-biochemistry, paleo–inorganic chemistry, and early biotic biophysical chemistry, as first envisioned by Pauling and Zuckerkandl (Pauling and Zuckerkandl, 1963; Zuckerkandl and Pauling, 1965). The ability to look back from extant biology by molecular resurrection provides the basis for linking evolutionary origins and evolutionary paths with models of Earth's early history.
Understanding the origin of life requires that we bridge the gap between prebiotic chemistry and extant biochemistry. One must connect the Top-Down and Bottom-Up approaches. We believe one key to understanding the origin of life is a realization that there was no discontinuity in the origin of life. The process was smooth and continuous with no clear demarcation between prebiotic and biotic. Each step was thermodynamically or kinetically favored under the prevailing conditions. Living systems are linked, by traceable threads, to their prebiotic chemical origins. Elements of the nonliving world were co-opted and incorporated into living systems. “Life or protolife learned from the abiotic world.” Many of those prebiotic elements have been retained in extant biology.
Although a clear path to the origin of life on Earth is yet to emerge, recent insights are providing a picture of the evolutionary history and origin of key elements of the major tenants we typically use to define life, information pathways, and metabolism. Significant clues can be gleaned from extant biology and deep phylogenic studies that provide a strong foundation for making meaningful connections to the salient elements of origin-of-life theories like the “RNA World” or “Iron-Sulfur World.” Our own work on ribosome and iron-sulfur enzyme structure, function, and evolution provides examples of the important advances being made and connections that are emerging.
Translation and the Ribosome
The ribosome is a macromolecular machine responsible for the synthesis of coded protein in all living organisms. Koonin (Wolf and Koonin, 2007) and Gogarten (Fournier et al., 2010) showed that a high level of detail of early ribosomal evolution is accessible from analysis of sequence databases. Elements of the ribosome are very highly conserved, in sequence and especially in three-dimensional structure, throughout the tree of extant life, and are very, very old, predating the last universal common ancestor. In fact some elements of the extant ribosome, the peptidyl transferase center (PTC) in particular, are clearly older than coded protein.
We view the PTC as one of our most ancient and informative molecular fossils. The PTC emerged from the RNA World (Rich, 1962; Woese, 1967; Crick, 1968; Orgel, 1968; Gilbert, 1986), participating in and enabling the transformation to current biology of information transduction from nucleic acid to protein. The PTC provides a direct connection to the distant evolutionary past, to early life, and to the origin of life (Woese, 2001; Wolf and Koonin, 2007; Bokov and Steinberg, 2009; Hsiao et al., 2009; Belousoff et al., 2010; Fournier et al., 2010). Understanding the origin of the PTC is key to understanding the origin and early evolution of life on Earth.
Important information about the PTC has been revealed by recent determination of high-resolution, three-dimensional ribosomal structures from disparate regions of the evolutionary tree. The current structural database contains X-ray–determined structures from S. cerevisiae, T. thermophilus, H. marismortui, E. coli, D. radiodurans, T. thermophile (Cate et al., 1999; Ban et al., 2000; Harms et al., 2001; Selmer et al., 2006; Ben-Shem et al., 2010; Rabl et al., 2011). Although the ribosome contains many proteins, RNA is the central player in catalysis and decoding (Noller et al., 1992; Ban et al., 2000; Nissen et al., 2000; Harms et al., 2001; Ogle et al., 2001; Yusupov et al., 2001; Schuwirth et al., 2005; Selmer et al., 2006). These three-dimensional structures allow one to infer how the primitive RNA of the PTC interacted with itself and with ions and proto-protein (Hsiao et al., 2009). Our interpretation of these structures is that the ancestors of modern RNA and protein were short and heterogeneous (in backbone and sequence) (Hsiao et al., 2009). We believe that the combined evidence is consistent with a model in which the ancestral PTC performed primitive catalysis as a nonspecific condensation enzyme. The ancestral PTC was a “sausage maker,” producing a noncoded mixture of short racemic oligomers of peptide, esters, and other products of simple condensation reactions.
In these models of early ribosomal evolution, some of the products of the primitive PTC bound to it, conferring catalytic advantage, and were converted over time to life's earliest coded proteins. The primitive catalytic sausage maker has been retained in the modern ribosome. Unlike modern enzymes, specificity of the ribosome is layered on by factors (synthetases, translation factors, the small subunit) beyond the catalytic center.
The Origins of Replication
Modern DNA replication is a highly complex process in which molecular assembly (base pairing) and chemical transformation (polymerization) are tightly coupled. The antiparallel arrangement of DNA strands and the non-self-complementarity of the four monomer units (G, C, A, T) require two replication cycles for duplication of a given strand of DNA (CGAT gives GCTA gives CGAT).
If the ancestral replicative systems were simple, then modern DNA synthesis has drifted far from its roots. A requirement for simplicity implies that the processes were spontaneous and noncatalytic, and achieved duplication in a single cycle. This requirement also implies that molecular assembly and chemical transformation were decoupled.
Is it possible to construct experimentally testable, molecular-level models of simple replication that meet the requirements above? Yes. In fact “replication” is essentially unavoidable in an environment of random synthesis and molecular assembly. As shown in Fig. 2, random monomer synthesis, random oligomerization, and random degradation will directly amplify a subset of the oligomerized molecules, if that subset forms molecular assemblies. Amplification arises because assembled oligomers are more resistant to degradation than non-assembled oligomers. This assembly-linked resistance to degradation is observed for modern biological polymers, including DNA, RNA, peptides, proteins, carbohydrates, and membranes. The resistance of assemblies to degradation is a readily accessible process that achieves the result of replication for essentially any oligomeric molecules capable of selective assembly.
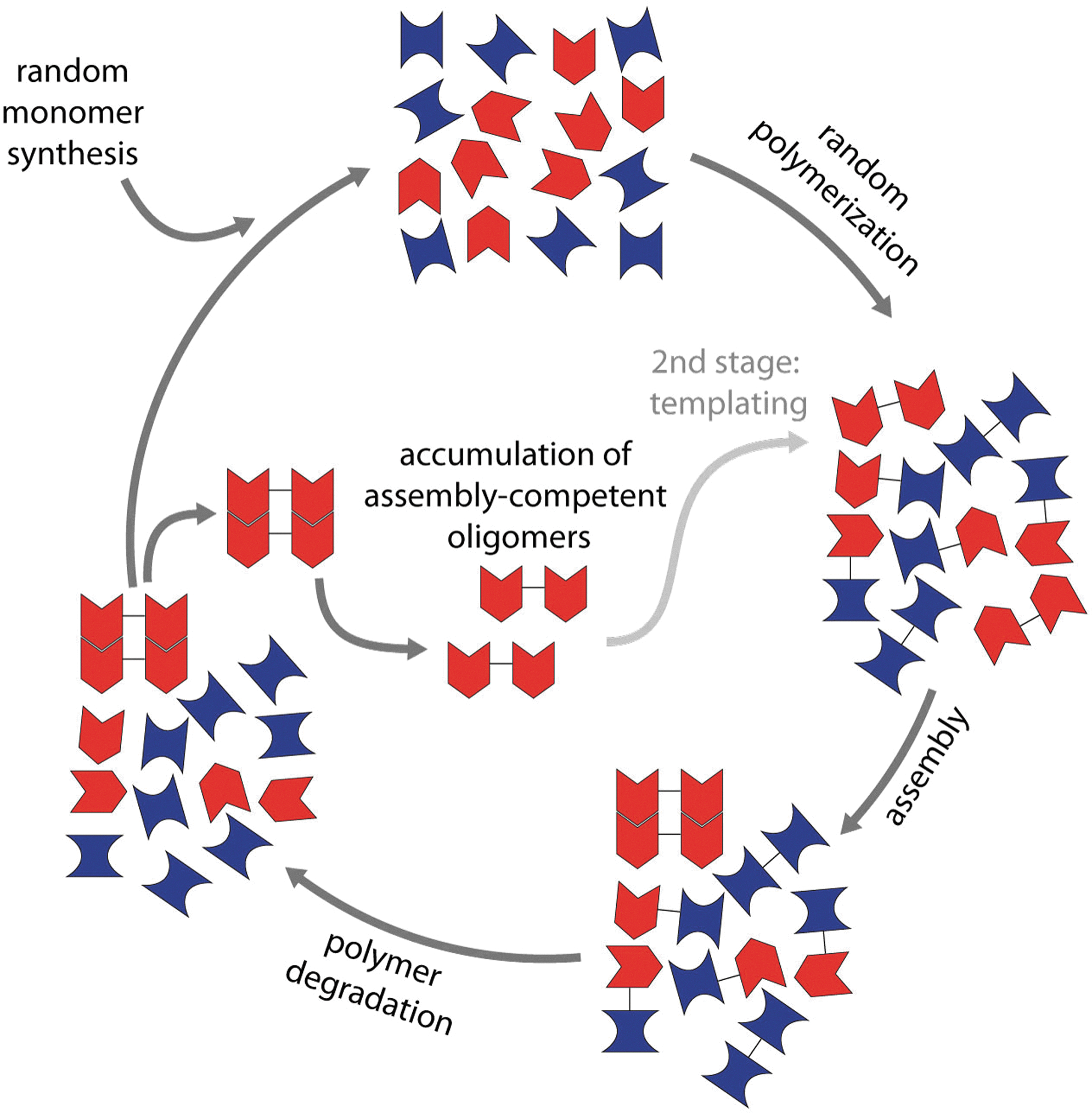
A chemical model of simple “replication.” In this schematic representation, two monomeric species (red and blue) are synthesized and oligomerized in stochastic processes. A subset of randomly synthesized oligomers is capable of forming molecular assemblies, stabilized by noncovalent interactions. Assembled oligomers degrade more slowly than the other oligomers; thus the assembled oligomers accumulate. In later stages, some assemblies dissociate and form templates that promote synthesis of other oligomers. This simplified view shows only two chemical species, which form two-residue oligomers, and separates processes in time and space. In fact, many chemical species can form many oligomer types and lengths, some of which assemble at the same time and in the same space.
Iron and Sulfur
The relationship of prebiotic to biotic is suggested by the structures and reactivities of iron-sulfur motifs within extant proteins. Iron-sulfur proteins are ubiquitous enzymes in life (Holm et al., 1996; Beinert et al., 1997; Johnson, 1998; Beinert, 2000; Noodleman et al., 2002; Johnson et al., 2005) and are integral to energy metabolism. Many iron-sulfur enzymes share critical features with abiotic iron-sulfur minerals (Martin and Russell, 2003; Wachtershauser, 2006; McGlynn et al., 2009). Complex iron-sulfur enzymes perform key redox interconversions of small molecules thought to have been of paramount importance in prebiotic and early biotic chemistry. The parallels between the structure and reactivity of iron-sulfur enzyme and iron-sulfur minerals appear much too strong to be coincidental, suggesting that life learned how to make iron-sulfur compounds by mimicking the features of iron-sulfur minerals present in rock formations in the nonliving Earth (Wachtershauser, 1992). It is with regard to transition metals and especially iron-sulfur minerals that parallels between life and what we know to be common features of early Earth are arguably the strongest. Since the parallels appear to be too strong to be coincidental, we can conclude that life learned how to make iron-sulfur compounds by mimicking the features of iron-sulfur minerals present in rock formations in the nonliving Earth.
Ligand-Accelerated Catalysis and Organic Nesting
It has been previously suggested that the path to metal enzymes important for both early and contemporary biology occurred stepwise (McGlynn et al., 2009). Conceptually, ligand-accelerated catalysis is the modification of mineral surfaces and metal clusters by organics, which can occur iteratively to evolve and tune the diversity and efficacy of minerals and enhance various high-barrier reactions (Fig. 3). “Nesting” represents the first step whereby life or protolife generated mobile forms of the reactive elements of minerals and enhanced the reactivity by providing a water-soluble ligand environment. In our recent work, we have made significant advances in understanding how iron-sulfur clusters, specifically the complex iron-sulfur clusters that catalyze reactions important for early life, are synthesized in biology (Shepard et al., 2011; Peters and Broderick, 2012). We have learned that, for some systems, those that are involved in certain forms of hydrogen metabolism and nitrogen fixation, hydrogenase, and nitrogenase, the active site metal clusters are synthesized stepwise. Key structural determinants are modifications introduced through the decomposition of simple organics such as amino acids in unique organic radical-based reactions in the vein as what has been rationalized for prebiotic ligand-accelerated catalysis (McGlynn et al., 2009). The final step of complex iron-sulfur maturation is the insertion and nesting of the synthesized cluster in a structural protein specifically evolved for maximal activity. These observations bring us significantly closer to defining a continuum between the nonliving and living Earth.
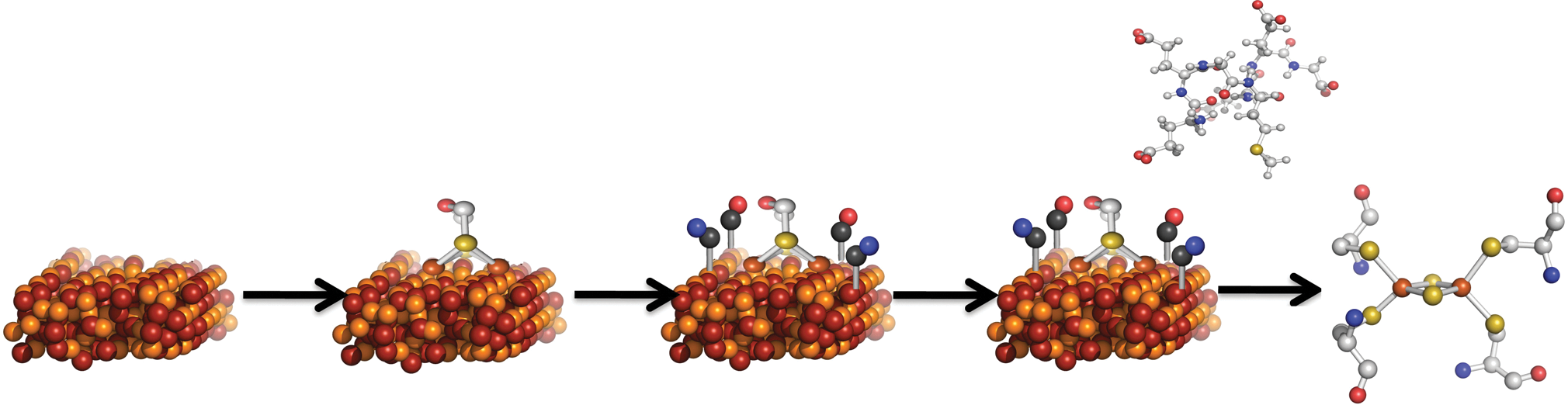
Hypothetical path from reactive mineral surfaces to metal clusters by the modification of mineral surfaces by organic-based ligands followed by the nesting of iron-sulfur moieties within organic groups to form water-soluble clusters.
A Match Made in the Hadean
Throughout extant biology, we see strong partnerships between elements that we might say link back to an RNA World (nucleotides) and the Iron-Sulfur World (iron-sulfur clusters), as illustrated in Fig. 4. For the aforementioned hydrogenases and nitrogenases, key elements of their catalytic sites are unique nonprotein ligands (Shepard et al., 2011; Duffus et al., 2012; Peters and Broderick, 2012). These ligands are introduced into the metal site by the activities of enzymes that utilize nucleotide–amino acid conjugate (S-adenosyl methionine). This conjugate reacts at an iron-sulfur cluster site to form an adenosyl radical that is a key intermediate in catalyzing a variety of biochemical reactions (Sofia et al., 2001). Members of this enzyme class can be rationally linked to many biological activities thought to date back to early and simple life. Indeed, a member of this enzyme class (anaerobic ribonucleotide reductase) is believed to be the most ancient version of a biocatalyst involved in converting ribose to deoxyribose for the first biologically synthesized DNA (Nordlund and Reichard, 2006). Perhaps more significant to the Astrobiologist is the shared role of nucleotides and iron-sulfur clusters in accomplishing biological oxidation-reduction reactions.

The RNA World and Iron-Sulfur World are models based on biology, biochemistry, and geochemistry. These models form a basis for understanding early biotic systems and the origin of life.
Energy metabolism for extant biology is universally driven by redox transformations. Both nucleotides and iron-sulfur clusters facilitate these redox transformations. Modern metabolism exists as a sequential bucket brigade in which electrons are passed between redox centers resulting in the conversion of reducing potential to more unifying biotic energy currency. It is becoming increasingly clear that, for even the simplest and arguably the most ancient forms of life, nucleotide and iron-sulfur based cofactors and coenzymes work hand in hand in an integrated manner in energy metabolism.
As we probe the salient features of the profound contributions of the first generation of origin-of-life theories, we can clearly see elements that can be supported both in experimentation and from observations of the detailed features common to extant biology. It is clear that, for many processes that we consider of paramount importance for early life, features of different theories or worlds (RNA World, Iron-Sulfur World) developed partnerships very early and likely prior to life's beginnings.
Footnotes
Abbreviation
PTC, peptidyl transferase center.