
Abstract
Microgravity is one of the most important features in spaceflight. Previous evidence has shown that neurophysiological impairment signs occurred under microgravity. The present study was undertaken to explore the change in protein abundance in human SH-SY5Y neuroblastoma cells that were grown in a microgravity environment. The comparative proteomic method based on the 18O labeling technique was applied to investigate the up-regulated proteins and down-regulated proteins in SH-SY5Y under simulated microgravity. Twenty-two differentially abundant proteins were quantified in human SH-SY5Y neuroblastoma cells. The cell microfilament network was disrupted under simulated microgravity, which was determined by the immunocytochemistry. The concentration of reactive oxygen species, malondialdehyde, and free Ca2+ ion significantly increased, and the level of ATP significantly decreased under simulated microgravity. However, there was no obvious cell apoptosis observed under simulated microgravity. These results provide new molecular evidence for the change in protein abundance in SH-SY5Y cells under simulated microgravity, which might unfold biological mechanisms and the development of effective countermeasures to deal with microgravity-related neurological problems. We believe that the state-of-the-art proteomic assay may be a means by which aerospace scientists will begin to understand the underlying mechanisms of space life activities at the protein level. Key Words: Comparative proteomics—Simulated microgravity—18O labeling—SH-SY5Y. Astrobiology 13, 143–150.
1. Introduction
2. Materials and Methods
2.1. Chemicals and reagents
Dulbecco's modified Eagle's medium (DMEM) was acquired from Invitrogen (Carlsbad, CA, USA). Sequencing-grade modified trypsin was purchased from Promega (Madison, WI, USA). Urea, dithiothreitol (DTT), and thiourea were purchased from Sigma-Aldrich (Steinheim, Germany). The clinostat (Center for Space and Applied Research, Chinese Academy of Science, P.R China) was utilized to simulate microgravity. 18O water (purity>97%) was obtained from Cambridge Isotope Laboratories (Andover, MA, USA).
2.2. Cell culture and treatment
SH-SY5Y cells were cultured in DMEM supplemented with 10% fetal bovine serum and seeded at a density of 1×106 cells/mL in either normal gravity in tissue culture flasks for control cells or in simulated microgravity in clinorotation at 12 rpm for 24 h.
2.3. Sample preparation
The normal gravity group (NG, n=6) and simulated microgravity group (SMG, n=6) were sonicated in chilled buffer (7 M urea, 2 M thiourea, 40 mM Tris, 10 mM DTT, and protease inhibitors) for 30 s by a FS-2 Dounce homogenizer (Jingda Instrument, China). The crude extracts were centrifuged at 17,000g for 30 min, and the resulting supernatants were collected and quantified by Bradford assay.
2.4. Digestion, labeling, and desalting
The comparative proteomic strategy based on the 18O labeling technique is shown in Fig. 1. Each sample was reduced with 10 mM DTT, alkylated with 50 mM iodoacetamide, and then diluted and digested for 19 h at 37°C by trypsin. After the digestion, the NG peptides and SMG peptides were evaporated to dryness, redissolved in H2 18O or H2 16O, and catalyzed by trypsin in the presence of 50 mM K2HPO4/KH2PO4 buffer at 37°C for 19 h. The residual trypsin activity was quenched by boiling for 10 min and the addition of 3% formic acid (v/v). The 18O labeled peptides (NG) and unlabeled peptides (SMG) were mixed at a 1:1 ratio and desalted by a C18 solid phase extraction column (Supelco, Bellefonte, PA, USA).
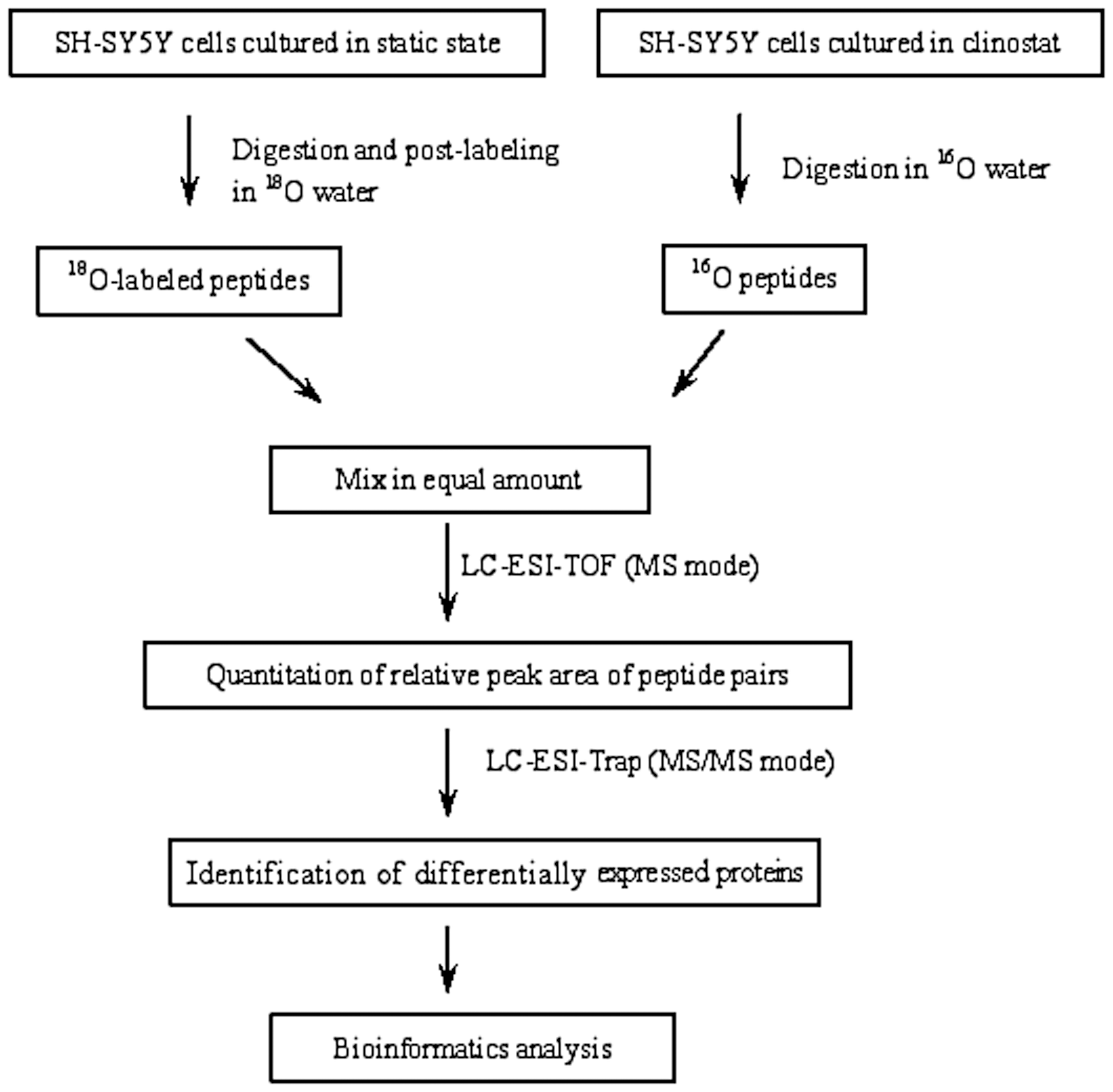
Comparative proteomic analysis workflow based on 18O labeling, LC-ESI-TOF, and LC-ESI-Trap analysis successively.
2.5. Strong cation exchange (SCX) fractionation
Strong cation exchange chromatographic separation of the peptide mixtures was carried out by an Agilent 300-Å SCX 4.6 mm i.d.×250 mm column with a flow rate of 0.7 mL/min. Mobile phase solvents consisted of (A) 10 mM ammonium formate, 25% acetonitrile, pH 3.0 and (B) 500 mM ammonium formate, 25% acetonitrile, pH 6.8. Peptides were separated with a linear gradient from 3% to 40% B over 50 min at 280 nm. The fractions were collected every 2 min and stored at −80°C.
2.6. Capillary LC-ESI-TOF and ESI-Trap analysis
Each SCX fraction was analyzed with a capillary HPLC system coupled to an Agilent 6210 electrospray–time of flight (ESI-TOF) or Agilent electrospray–ion trap (ESI-Trap) mass spectrometer. The 5 μm C18, 150 μm i.d.×150 mm column (Column Technology Inc., Fremont, CA, USA) was eluted with a linear acetonitrile gradient elution from 100% solvent A (water with 0.1% formic acid) to 60% solvent A+40% solvent B (acetonitrile with 0.1% formic acid) over 60 min at a flow rate of 1.8 μL/min, and from 40% to 95% solvent B over 10 min, finally at 100% solvent B for another 10 min. During ESI-TOF and ESI-Trap analysis, the nebulizer pressure was 15 psi. Drying gas flow and temperature were 7 L/min and 325°C, respectively. The mass spectrometer scan range was set between a mass of 100 and 1500.
2.7. Data analysis
Tandem mass spectrometry data generated by data-dependent acquisition via the ion trap were extracted by Mascot Distiller 2.2.The data files of all fractions were merged and then searched against a SwissProt database (updated as of July 1, 2011) by using Mascot (version 2.1; Matrix Science, Boston, MA, USA) with the following parameters: taxonomy, human; fixed modification, carbamidomethyl; enzyme, trypsin; precursor tolerance, 2Da; MS/MS tolerance, 0.8Da; maximum number of missing cleavages, 2. Mascot cutoff scores were determined by using a reverse database strategy producing a false-positive rate of less than 5% for doubly and triply charged peptides. The ratio of chromatographic peak area of 16O2 peptides and 18O2 labeled peptides in MS scans was utilized to quantify the differential peptides.
2.8. Determination of cellular malondialdehyde (MDA)
Malondialdehyde is the final product of lipid peroxidation evaluated by thiobarbituric acid assay. MDA concentration for both the NG and SMG cells was measured at 532 nm by spectrophotometer, and the results were expressed as nmol/100 mg protein.
2.9. Quantification of cellular reactive oxygen species (ROS)
The cellular total ROS was quantitated with a cell-permeable probe, 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA). Both the NG and SMG cells were harvested, washed with phosphate-buffered saline (PBS), and resuspended in serum-free medium. The cells were loaded with 10 μM DCFH-DA for 30 min at 37°C, washed with PBS, and placed on ice. Fluorescence was measured by a Coulter EPICS XL flow cytometer (Beckman Coulter, Hialeah, FL, USA). The excitation wavelength was 488 nm, and the emission was 525 nm.
2.10. Quantification of intracellular Ca2+ concentration
A Fluo-3 AM ester fluorescent probe (Beijing Biolead Biology Sci & Tech, China) was used to measure intracellular Ca2+ concentration on a flow cytometry following the manufacturer's protocols. Both the NG and SMG cells were washed with PBS twice and replaced with 2 μM Fluo-3 AM solution. Each sample was incubated at 37°C for 45 min and then for 20 min at room temperature. The fluorescence was determined with a Coulter EPICS XL flow cytometer (Beckman Coulter, Hialeah, FL, USA) at 488 nm for excitation and at 525 nm for emission.
2.11. Measurement of mitochondria membrane potential (ΔΨm)
As a cationic fluorescent probe, rhodamine-123 was used for measurement of mitochondrial membrane potential. Both the NG and SMG cells were incubated with 1 μM Rho-123 for 30 min at 37°C, washed twice with PBS. Fluorescence of rhodamine 123 was measured by flow cytometer.
2.12. Analysis of cell apoptosis
After SH-SY5Y cells were cultured in clinostat or flasks for 24 h, the cells were washed with PBS twice, suspended in 500 μL binding buffer, and stained by 5 μL fluorescein isothiocyanate (FITC)–conjugated annexin V and 5 μL propidium iodide. Annexin V-FITC apoptosis detection kit (Beyotime, China) was used to detect apoptosis of NG and SMG SH-SY5Y cells by Coulter EPICS XL flow cytometer (Beckman Coulter, USA). The excitation wavelength was 488 nm, and the emission wavelength was 620 nm.
2.13. Fluorescence cytochemistry
Both the NG and SMG SH-SY5Y cells were incubated with FITC-phalloidin for 30 min to probe the structure of the cytoskeleton and incubated with 4′,6-diamidino-2-phenylindole (DAPI) for 15 min. Fluorescence imaging of cells was visualized on a Leica TCS-SP5 confocal microscope (Leica Microsystems, Mannheim, Germany).
2.14. Western blot
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was performed with a Bio-Rad mini protein III apparatus. The samples under NG and SMG were separated by SDS-PAGE (10% gel). The gels were electroblotted onto 0.45 μm PVDF membrane. The membrane was blocked with 10% nonfat milk in PBS for 1 h at 37°C and then incubated with antibodies raised against S100 calcium-binding protein A11 (S100-A11) or glyceraldehyde-3-phosphate dehydrogenase (GADPH, internal standard) and appropriate secondary antibodies. Detection of immune complex was performed by enhanced chemiluminescence.
2.15. Statistics
The results were assessed by ANOVA followed by Newman–Keuls multiple comparisons. The null hypothesis used for all analyses was that the factor had no influence on the measured variable, and significance level is chosen to be 0.05.
3. Results
Due to tens of thousands of proteins in a cell and the wide dynamic range of protein concentration in the sample, two-dimensional LC-MS analysis was utilized to identify more differentially abundant proteins. The NG and SMG samples were fractionated by SCX. Then, each fraction was subjected successively to LC-ESI-TOF and LC-ESI-Trap analysis, which was based on a quantification and identification-decoupled approach (Wang et al., 2010a,b). Time-of-flight and ion trap mass spectra provided quantitative or qualitative information of proteins, respectively. A comparison between the NG and SMG fractions in this comparative proteomic analysis led to the identification of 22 differentially abundant proteins in human SH-SY5Y neuroblastoma cells. A list of proteins that are differentially abundant between NG and SMG is shown in Table 1, including accession number, protein ID, protein names, isoelectric point, and average ratio. Among the differentially expressed proteins, the level of S100-A11 was elevated through proteomic analysis and was further confirmed by Western blot (Fig. 2). In addition to the 16 up-regulated and 6 down-regulated proteins in SMG, we also found that the ROS level had a significant increase in SMG cells (Fig. 3A). The ROS can attack cell membranes and then lead to the lipid peroxide. MDA is the final product of lipid peroxide. MDA concentration in SMG was also significantly elevated compared with NG (Fig. 3B). Simulated microgravity resulted in a 5.7-fold increase for MDA and 6.3-fold increase for ROS, respectively. The excessive ROS can also attack the membrane of mitochondria and open the permeability transition pore, and induced a 84% decrease in mitochondrial membrane potential (Fig. 4A). The free calcium concentration in SMG significantly increased by 93% compared with NG (Fig. 4B), and ATP level significantly decreased by 63% in SMG. However, there was no obvious cell apoptosis observed under simulated microgravity (Fig. 5). To further assess whether simulated microgravity changed SH-SY5Y cytoskeleton architecture, FITC-phalloidin was used to stain the cytoskeletal actin filaments and DAPI to stain the DNA. Immunofluorescence images showed that the SH-SY5Y cells under simulated microgravity actually were much rounder and spread more than cells in the static culture (Fig. 6). The disruption of cell cytoskeletal actin filaments network was induced under the simulated microgravity.
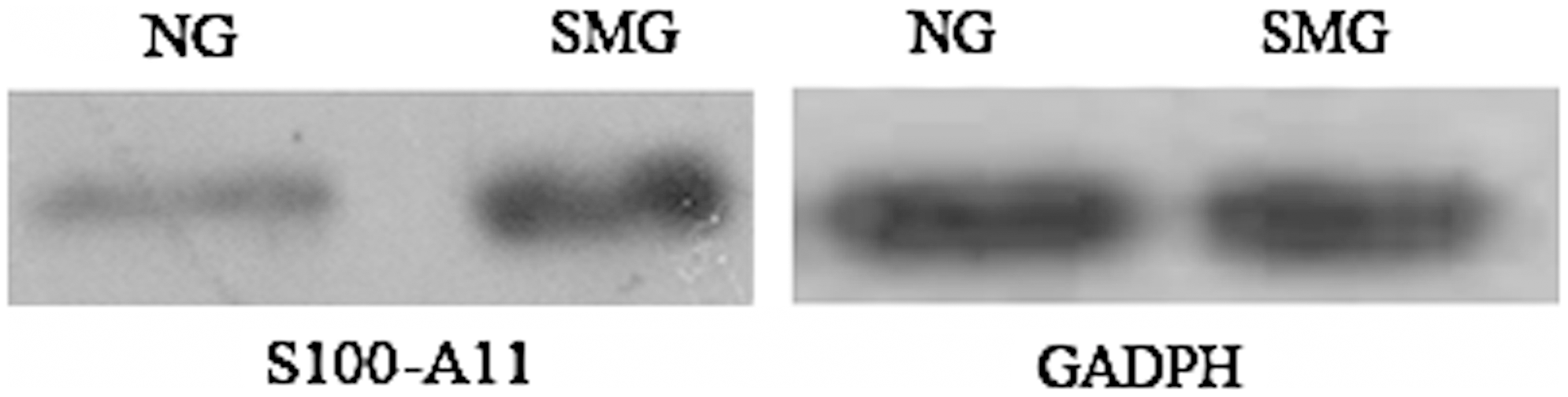
Western blot of S100-A11 and glyceraldehyde-3-phosphate dehydrogenase (GADPH) in NG and SMG (n=3).

Effect on SH-SY5Y cellular MDA concentration (
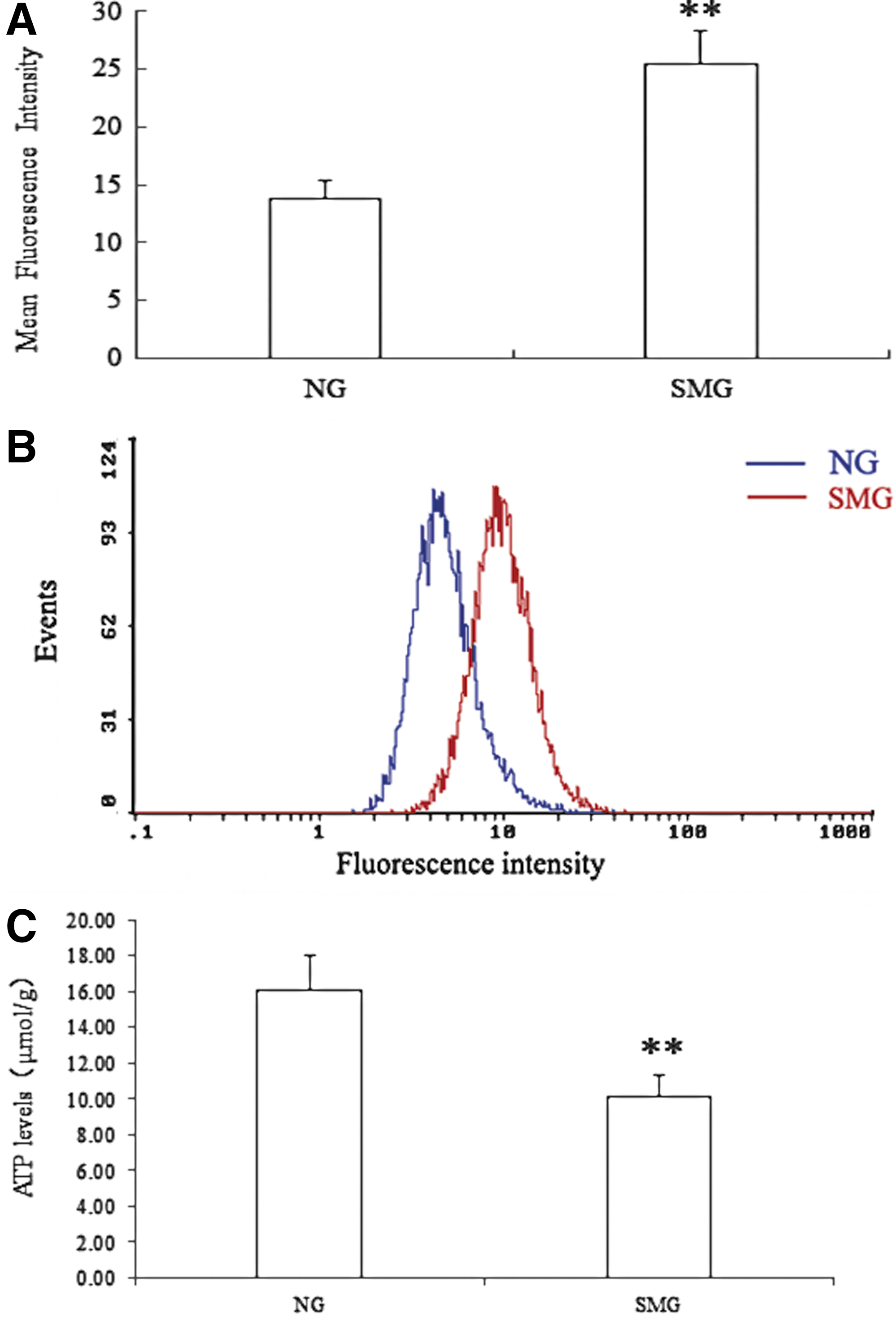
Effect on SH-SY5Y mitochondrial membrane potential (
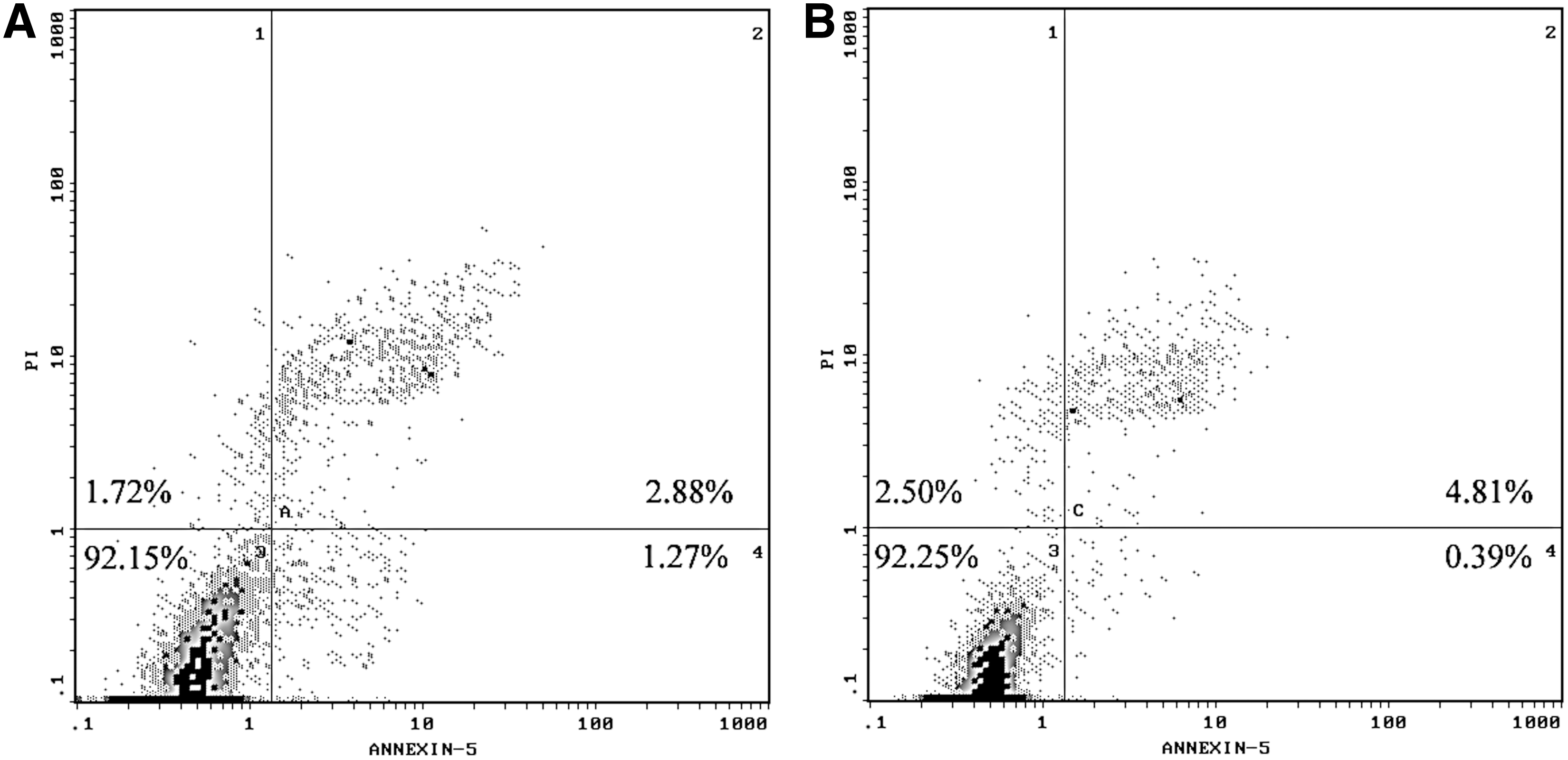
Effect on SH-SY5Y cell apoptosis under simulated microgravity. Apoptosis was quantified with the annexin V-FITC apoptosis detection kit. The percentage of annexin-V-positive cells was determined and shown. There was no significant difference between NG (
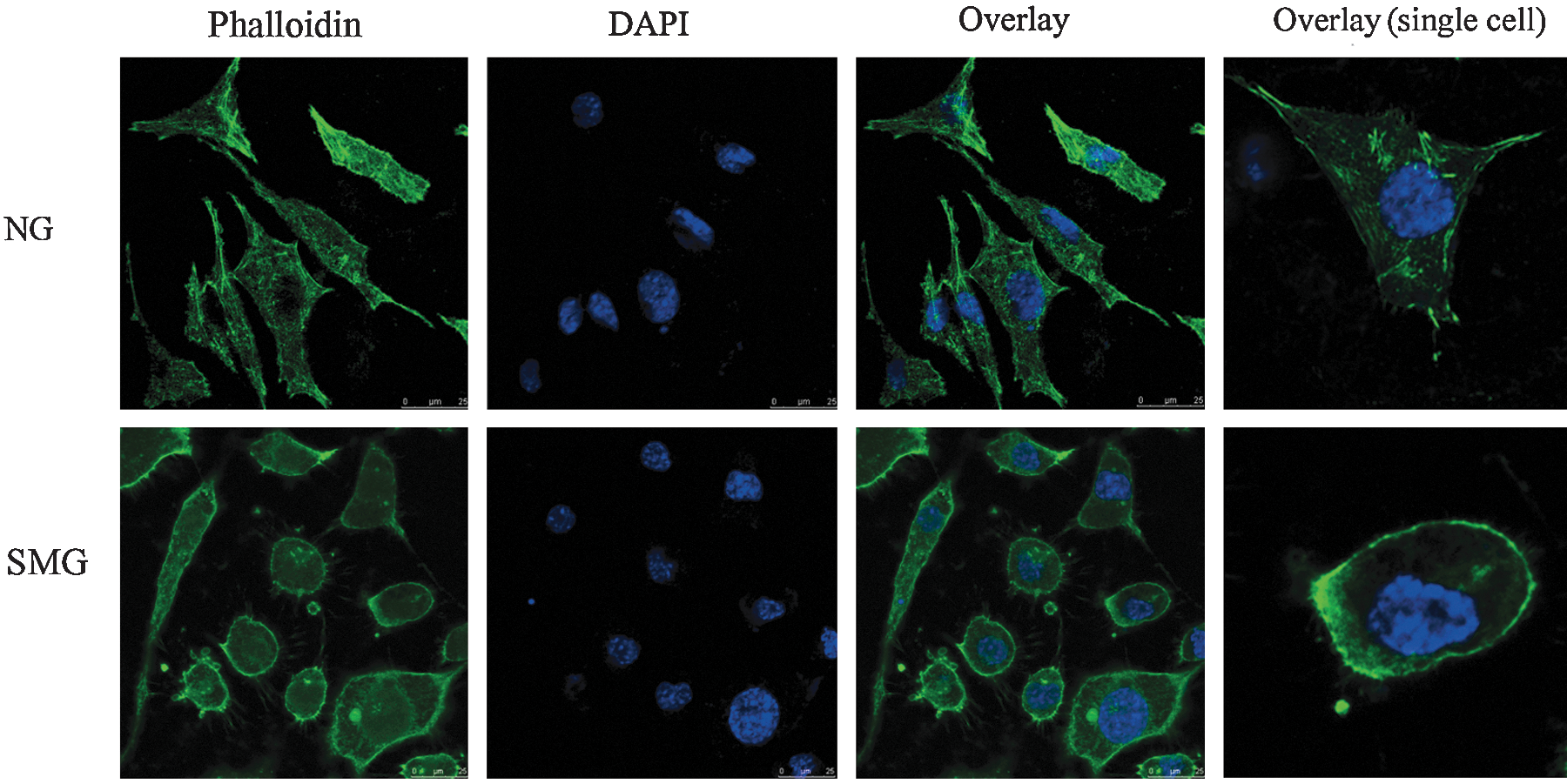
The immunofluorescence images of SH-SY5Y cells both in NG and SMG. The scale bar represents 25 μm. FITC-phalloidin staining of microfilament probes the structure of cytoskeleton, and DAPI staining of DNA shows cell nuclei. Overlay of the two images and higher magnification image (single cell) are shown on the right.
Isoelectric point (pI) is derived from Swiss-Prot.
16O (SMG) to 18O (NG) ratio from the base peak intensities for the monoisotopic peak for the peptides with and without 18O label, respectively.
Results are the mean±SD obtained from two and more peptides from the same protein.
4. Discussion
During spaceflight, astronauts are exposed to various stresses such as microgravity, radiation, low temperature, and hypoxia. Microgravity can lead to cardiovascular disorders (Karlsson et al., 2009), bone loss (Hwang et al., 2009), space motion sickness (Sa et al., 2009), immune systems disorders (Martinelli et al., 2009), and neurological disorders (Wang et al., 2009). Because spaceflight is costly and difficult to schedule, we used a clinostat to simulate microgravity on the ground. Simulated microgravity is based on the principle of gravity vector averaging. Microgravity simulation is achieved by continuously changing a cell's position in the clinostat relative to Earth's gravity vector direction (Zhang et al., 2012). In the present study, 22 differentially abundant proteins were quantified in human SH-SY5Y neuroblastoma cells exposed to simulated microgravity by way of comparative proteomic technology.
The levels of heat shock proteins (HSPs) HSP70 and HSP90 were up-regulated in SMG compared to NG cells (Table 1). On one hand, increased HSP levels can be indicative of increased cellular stress (Kozeko and Kordyum, 2006). Our results show that the level of MDA and ROS was significantly elevated in SMG, which are important markers of oxidative stress. It was also previously reported that simulated microgravity induced a significant increase in the level of MDA and ROS in affected cells or tissues (Qu et al., 2006; Chowdhury et al., 2009; Wang et al., 2009). Up-regulation of HSP70 and HSP90 might protect cells against stress and maintain their proliferative potential under simulated microgravity (Cotrupi and Maier, 2004). This might, therefore, represent an important adaptive response of human SH-SY5Y cells to simulated microgravity (Mariotti and Maier, 2009). On the other hand, HSPs play a more important role in regulating cell apoptosis. Not only can they promote cell apoptosis to maintain a population of cells with normal activities, but they also act as the key signal protein to inhibit cell apoptosis in order to protect the cell. In our case, there is no cell apoptosis observed in SMG, although ROS and MDA are elevated in SMG. In a previous study, simulated microgravity inhibits the proliferation of K562 erythroleukemia cells but does not result in apoptosis either (Yi et al., 2009). Thus, up-regulation of HSP70 and HSP90 might inhibit cell apoptosis in SMG.
Interestingly, the abundance of cytoskeleton-related proteins in the present study underwent change in SMG cells. The cytoskeleton is a complex network of fibers that is sensitive to gravity (Schatten et al., 2001). Cytoskeletal integrity is important for cellular functions and influences the cell division, neuronal outgrowth, and apoptosis. It has been reported that microgravity affects microtubules, microfilaments, and intermediate cytoskeletal filaments (Infanger et al., 2006; Rösner et al., 2006). These modifications may be a main reason for previously observed changes in cell architecture. Our results also suggest that the cytoskeletal architecture of neuronal SH-SY5Y cells is sensitive to gravity changes in that these cells underwent change due to microgravity. Moreover, immunofluorescence images also reveal that the shape of cells became round and the cytoskeletal actin filaments were in a state of “chaos” in SMG (Fig. 6).
Compared to NG cells, 14-3-3 protein zeta/delta was up-regulated in SMG cells, which possibly correlates with the oxidative stress under microgravity. An elevated level of 14-3-3 proteins may help cells become resistant to microgravity stress. The cells in SMG might suspend the proliferative and metabolic machinery. It has been previously reported that the expression of 14-3-3 protein also increases in vascular smooth muscle cells and breast cancer cells under modeled microgravity (Coinu et al., 2006).
S100 calcium-binding protein A11 is a member of the S100 family of calcium-binding proteins that is localized in the cytoplasm and/or nucleus of a wide range of cells. This protein may function in motility, invasion, and tubulin polymerization. Our results show that the free Ca2+ ion concentration was significantly elevated in SMG. The elevation of the intracellular Ca2+ concentration may have resulted from increased ROS, which occurs in the cells in simulated microgravity. S100-A11 protein with two EF hands binds two calcium ions per molecule, responds to the elevation of Ca2+ concentration, and functions as Ca2+ sensors. Therefore, the up-regulation of S100-A11 in SMG was closely correlated with the increment in the intracellular Ca2+ concentration (Sui et al., 2008). S100-A11 was also shown to interact with cytoskeletal proteins, which might be correlated with the disruption of cytoskeletal architecture under microgravity (Zhao et al., 2000). Based on the preliminary results, we propose a potential mechanism of SH-SY5Y cell impairment that may occur under simulated microgravity (Fig. 7).
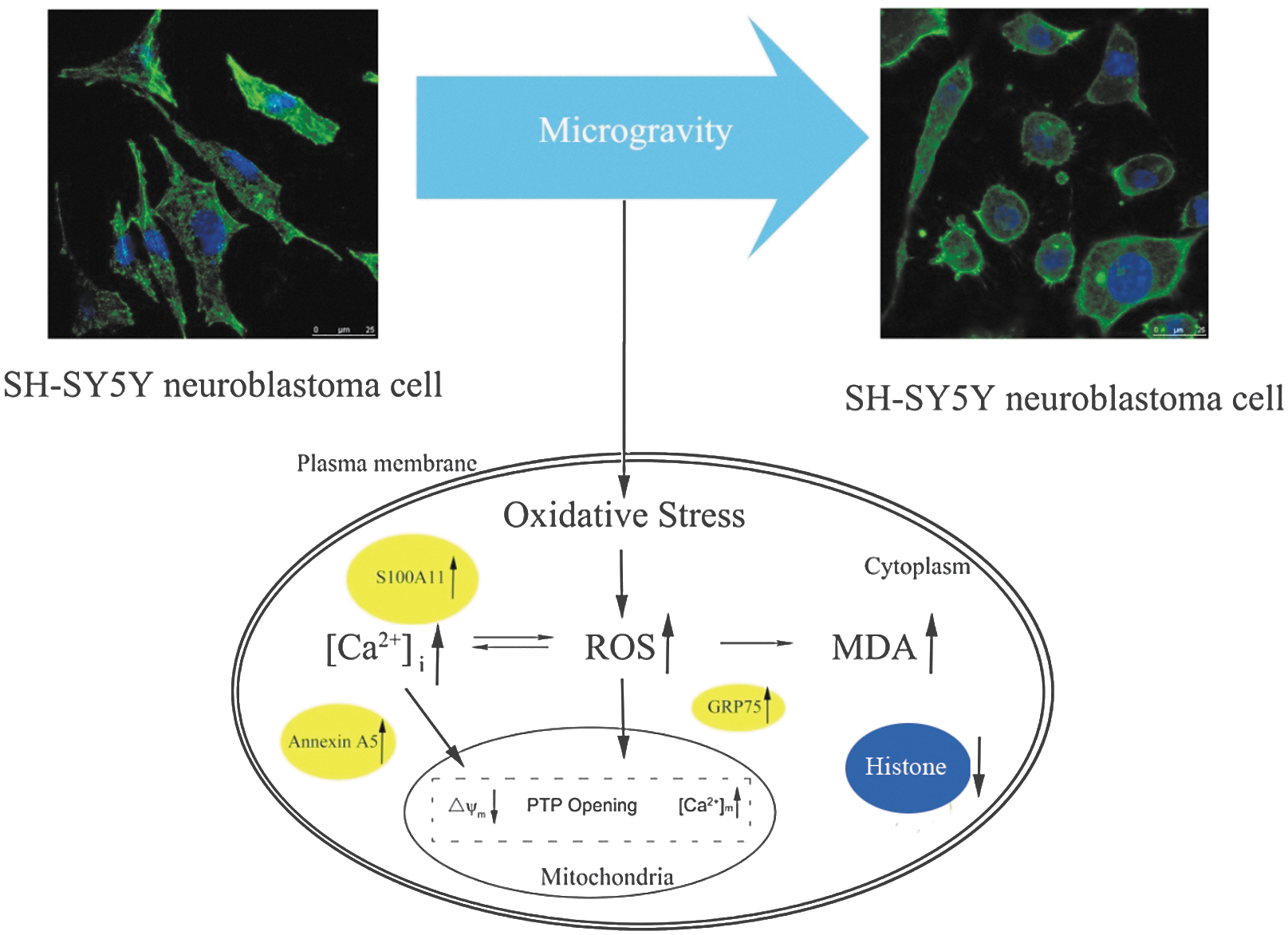
The potential mechanism of SH-SY5Y cell impairment under simulated microgravity.
In the present study, the ground-based simulated microgravity experiments revealed a high degree of changes in protein abundance in neuronal SH-SY5Y cells, which might have an effect on the health and cognitive abilities of astronauts during space travel. We propose that the differentially expressed proteins we identified could be used as potential drug targets for space adaptation syndrome, space motion sickness, and potential neurological diseases induced by microgravity. They could also be used to elucidate possible molecular mechanisms to develop effective countermeasures. It is worth mentioning that, due to its high sensitivity, the proteomic assay was suitable for the detection of proteins in rare and low amounts of aerospace samples. This state-of-the-art proteomic assay may become a key resource for aerospace scientists in understanding the underlying mechanisms of space life activities at the protein level. Because we only analyzed changes in protein abundance under ground-based simulated microgravity, we suggest that in future studies a combination of microgravity and other stresses, such as various radiations, should be used in simulated environments on the ground or in space in an effort to protect astronauts in space and upon their return to Earth.
5. Conclusions
Microgravity is one of the more significant stresses astronauts endure during spaceflight. By way of a comparative proteomic analysis, 22 differentially abundant proteins were discovered in neuronal SH-SY5Y cells under simulated microgravity. The potential mechanism of SH-SY5Y cell impairment under the simulated microgravity is proposed based on our preliminary results. The differentially abundant proteins will serve as markers for future microgravity studies and might provide some beneficial references for developing countermeasures to improve or maintain the health and safety of astronauts during space travel.
Footnotes
Acknowledgments
We thank Dr. Michael Schläppi (Marquette University, Milwaukee, Wisconsin) for language editing of this manuscript. This work was partially supported by the Ministry of Science and Technology of China (2012YQ040140) and National Key Program for Basic Research of China (2012CB910603).
Author Disclosure Statement
The authors declare that no competing financial interests exist.
Abbreviations
DAPI, 4′,6-diamidino-2-phenylindole; DCFH-DA, 2′,7′-dichlorodihydrofluorescein diacetate; DMEM, Dulbecco's modified Eagle's medium; DTT, dithiothreitol; ESI, electrospray ionization; ESI-TOF, electrospray–time of flight; ESI-Trap, electrospray–ion trap; FITC, fluorescein isothiocyanate; HSP, heat shock protein; LC, liquid chromatography; MDA, malondialdehyde; MS, mass spectrometry; MS/MS, tandem mass spectrometry; NG, normal gravity group; PBS, phosphate-buffered saline; ROS, reactive oxygen species; S100-A11, S100 calcium-binding protein A11; SCX, strong cation exchange; SMG, simulated microgravity group.