
Abstract
Orbital and in situ analyses establish that aerially extensive deposits of evaporitic sulfates, including gypsum, are present on the surface of Mars. Although comparable gypsiferous sediments on Earth have been largely ignored by paleontologists, we here report the finding of diverse fossil microscopic organisms permineralized in bottom-nucleated gypsums of seven deposits: two from the Permian (∼260 Ma) of New Mexico, USA; one from the Miocene (∼6 Ma) of Italy; and four from Recent lacustrine and saltern deposits of Australia, Mexico, and Peru. In addition to presenting the first report of the widespread occurrence of microscopic fossils in bottom-nucleated primary gypsum, we show the striking morphological similarity of the majority of the benthic filamentous fossils of these units to the microorganisms of a modern sulfuretum biocoenose. Based on such similarity, in morphology as well as habitat, these findings suggest that anaerobic sulfur-metabolizing microbial assemblages have changed relatively little over hundreds of millions of years. Their discovery as fossilized components of the seven gypsiferous units reported suggests that primary bottom-nucleated gypsum represents a promising target in the search for evidence of past life on Mars. Key Words: Confocal laser scanning microscopy—Gypsum fossils—Mars sample return missions—Raman spectroscopy—Sample Analysis at Mars (SAM) instrument—Sulfuretum. Astrobiology 12, 619–633.
1. Introduction
The possible occurrence of fossils preserved in gypsum has been largely ignored by paleontologists. The first such find evidently dates from some 90 years ago (Tillyard, 1922), but such reports are rare, most centering on shelly invertebrate animals, the originally calcitic shells of which have been replaced by gypsum (e.g., Rettger, 1924), or on vertebrate bones encrusted and secondarily infused by gypsum (e.g., Paliwal, 2003). Two studies have reported the occurrence in Tertiary selenite of preserved insect parts—a grasshopper wing (Tillyard, 1922) and dragonfly larvae (Schlüter et al., 2003), documented primarily as useful additions to the notoriously poor entomological fossil record—and two recent studies have reported the occurrence of cyanobacterial fossils (Panieri et al., 2008) and cyanobacterial RNA gene fragments in Tertiary gypsum crystals (Panieri et al., 2010). There has been no concerted effort, however, to investigate the paleontology of primary sedimentary gypsum.
The lack of paleontological interest in gypsiferous deposits is easily explained. On Earth, such units are relatively uncommon, and the majority of paleontologists focus on megascopic animals or plants, organisms generally unable to cope with the hypersaline conditions of gypsum deposition. On Mars, however, such deposits, not only of gypsum but also of other evaporitic sulfate minerals, are widespread (e.g., Gendrin et al., 2005; Langevin et al., 2005). Moreover, several papers have recorded the presence of microbial endoliths, as analogues of a possible extant or past martian biota, in translucent gypsum crystals and soil grains (e.g., Stivaletta et al., 2005; Wierzchos et al., 2006; Dong et al., 2007); and it is well established that diverse communities of microorganisms are able not only to thrive in highly saline gypsum-precipitating settings (e.g., Oren et al., 2009) but also, for microbes such as those of a modern sulfuretum (e.g., Gallardo and Espinoza, 2007a, 2007b), to metabolize biologically available sulfate.
In preparation for studies of gypsiferous samples that may be included among rocks investigated by NASA's Mars Science Laboratory at Gale Crater and eventually included in the cache of rocks acquired for laboratory study by Mars sample return missions, we have therefore sought to
(1) determine whether the diverse microbes present in evaporitic settings on Earth are preserved in precipitated gypsum; and if so,
(2) establish whether such fossil-bearing deposits are of relatively common occurrence and
(3) document techniques that can be used to distinguish gypsum-preserved fossils from pseudofossil “look-alikes.”
To establish the relevance of this work to Mars exploration, we first briefly review, below, orbital and in situ analyses that demonstrate the presence on Mars of widespread gypsiferous deposits, some of which exhibit geological features suggestive of deposition in a lacustrine setting. Following this, we discuss the preservation of microbial fossils, so that the gypsum permineralization of the fossils reported here is placed in the context provided by other comparably preserved microorganisms, and address the problem of establishing their biogenicity, a matter certain to be central to interpretation of any lifelike microscopic forms detected in martian rocks. We then record the results of our studies of seven gypsiferous deposits in which we have detected permineralized microscopic fossils: one from the Miocene (∼6 million years old) of Italy; two from the Permian (∼263 and ∼255 million years old) of New Mexico; two from Recent lacustrine deposits of Australia; and two from Recent salterns of Mexico and Peru (cf. Schopf et al., 2010a; Foster et al., 2012).
We used four techniques in our investigation of these gypsum-embedded fossils: transmitted white light optical microscopy; plane polarized light optical microscopy; confocal laser scanning microscopy (CLSM); and Raman spectroscopy. Taken together, these investigations established both the biogenicity of the fossils reported and their indigenousness to the gypsum samples studied, showing that they are permineralized in primary bottom-deposited gypsum crystals and that the assemblages investigated contain both benthic components (e.g., filamentous microorganisms, some similar to those of a modern sulfuretum) and fossils of planktonic forms (e.g., diatoms and cyanobacterium-like filaments and unicells) that settled into the precipitating gypsum from the overlying water column.
Our further studies of these and similar deposits will present relevant geochemical data, illustrate additional specimens, systematically assess “lifelike” microscopic pseudofossils present in such gypsums (many of which, primarily fluid inclusions, are unreported from similarly permineralized fossil assemblages), and formally describe and name the fossils discovered. For the present, our aim is to document the occurrence of these gypsum-permineralized microfossils and demonstrate the use of non-intrusive, nondestructive analytical techniques that can be used to establish their biogenicity.
2. Gypsum on Mars
The search for preserved signatures of a past martian biosphere is central to current Mars exploration programs. Among the most promising potential targets for such studies are evaporitic chemical sediments (Farmer and Des Marais, 1999), shown both by orbital and in situ analyses to be present on the surface of Mars.
Orbital spectroscopic studies of the martian surface have detected aerially extensive deposits of sulfates interpreted to contain hydrous sulfate minerals (Gendrin et al., 2005; Squyres et al., 2006; Bishop et al., 2007; Roach et al., 2007; Wiseman et al., 2009), among which gypsum is compatible with the spectra acquired and, in some instances, is the primary mineral identified (Catling et al., 2006). Of particular relevance are regions having “bright surface units” that exhibit the spectroscopic signature of gypsum and are therefore regarded as representing significant deposits of fine-grained (e.g., Northern Plains), isolated mound/mantle (Iani Chaos), or layered gypsum units (viz, Juventae Chasma: Catling et al., 2006; Bishop et al., 2007). With the exception of the gypsum dunes of the Northern Plains (currently interpreted as a secondarily geochemically altered gypsiferous deposit; Fishbaugh et al., 2007), the detected units are interpreted as deposits of primary gypsum that formed in localized sites of upwelling acidic groundwater and/or shallow playa-like settings (Glotch and Rogers, 2007; Dobrea et al., 2008; Warner et al., 2011) or by evaporative concentration of an early Noachian sea or extensive lake (Catling et al., 2006).
These orbital studies are corroborated by in situ analyses acquired at the landing sites of NASA's Mars Exploration rovers, Spirit and Opportunity (Spirit, within Gusev Crater; Opportunity, on the plains of Meridiani Planum). In Gusev Crater, the wheels of Spirit were used to “trench” the regolith to a depth of 6–11 cm, and analyses of the material exposed showed it to be composed in part of white, fine-grained, sulfate minerals, which are interpreted to have formed by evaporative concentration of salt-rich groundwater (Arvidson et al., 2006; Hurowitz et al., 2006; Wang et al., 2006; Johnson et al., 2007). On the opposite side of the planet, at the landing site of Opportunity in Meridiani Planum, the Opportunity rover analyzed outcrops that contain hematite concretions—the well-known “martian blueberries” (Clark et al., 2005; Squyres et al., 2006)—and provided data that show rocks of this region to be composed of reworked sulfate-rich sediments (Squyres et al., 2006). Subsequently, along the rim of Endeavour Crater, data provided by Opportunity show that several outcrops are intruded by white mineral veins of nearly pure gypsum, which is interpreted to have been emplaced by percolating groundwater (Showstack, 2011).
Taken together, the orbital and in situ analyses establish that sulfate minerals, including gypsum—in some deposits demonstrably layered, like those of lacustrine deposits—are present on the martian surface.
3. Preservation of Microbial Fossils
Microbial fossils can be preserved in rocks by two principal processes: compression, their carbonaceous remnants flattened along bedding planes during the compaction and lithification of fine-grained clastic sediments, such as shales and siltstones; and permineralization, their cells infused by and three-dimensionally preserved within a precipitated mineral matrix (most commonly quartz, calcite, or apatite). Of all modes of fossil preservation, cellular permineralization provides the most faithful lifelike representation of biological form and cellular anatomy.
The fossils reported here are all preserved by permineralization (known commonly, but incorrectly, as “petrifaction”), a type of preservation first described more than a century ago (White, 1893). The misnomer petrifaction (from petrify, “to convert into stone”) dates from this early study when the process was misinterpreted as reflecting “molecule-by-molecule…replacement” of cellular organic matter by minerals (White, 1893). In contrast with this early account, permineralization is now known to result from the pervasion of mineral-charged solutions into biological tissues during early stages of diagenesis, prior to their decay and cellular disintegration (Schopf, 1975), a process analogous to the epoxy embedding of tissues of modern organisms in preparation for their study by transmission electron microscopy. The organic structures of such fossils, in many specimens preserved in microscopic detail, are not mineral-replaced. Rather, the permeating fluids infill micellar, intercellular, and intracellular spaces—replacing the watery milieu of the biomolecular components, not the molecules of the organic structures themselves—to produce a mineral-infused inorganic-organic mix that in microbes serves to preserve their physically robust organic-rich cell walls. The fossils reported here are permineralized in gypsum, rather than in quartz, calcite, or apatite, as is common for such preservation, but the process of preservation is the same: calcium sulfate–charged solutions infused their cells, prior to their decay and disintegration, and allowed for preservation in three-dimensional microscopic detail.
4. Establishment of the Biogenicity of Microbial Fossils
Establishing the biogenicity of ancient microscopic fossils—of distinguishing bona fide fossil microbes from “fossil-like” mineral grains and other microscopic pseudofossils—posed serious difficulties in the 1960s and 1970s when studies of Precambrian microorganisms, presumably the most likely analogues of past life on Mars, were just beginning (e.g., Cloud, 1973). Since that time, the problem has been addressed repeatedly and effectively: comprehensive tabulations have been compiled that separate authentic fossils from nonfossil “look-alikes” (Schopf and Walter, 1983; Mendelson and Schopf, 1992), and the characteristics expected of bona fide fossils have been tabulated and discussed in detail in a paper that presents side-by-side comparisons of authentic Precambrian microbes and reports pseudofossils with which they might be confused (Schopf et al., 2010b).
A principal conclusion of such analyses is that, of the cascade of mutually reinforcing evidence needed to establish biogenicity, two prime traits stand out: (1) authentic fossil microorganisms should exhibit cells and cell lumina, discernible walled compartments that, as typically preserved, are devoid of remnants of their originally water-rich cytoplasmic contents; and (2) the walls of such cells should be demonstrably of carbonaceous (kerogenous) composition. These traits, coupled with the biogenicity-indicating cascade of other evidence discussed by Schopf et al. (2010b), will need to be affirmed before the existence of past (or present) life on Mars can be established. Indeed, together with the absence of other biological characteristics, it was the lack of such carbonaceous cellularity that showed the “possible microfossils” reported from martian meteorite ALH84001 (McKay et al., 1996) to be nonbiological inorganic microstructures (Bradley et al., 1997; Schopf, 1999).
However, and unlike such studies of putative fossils in meteorites, the cost of collecting, amassing, and bringing martian rocks to Earth for detailed investigation—some tens of millions of US dollars per gram of rock—imposes additional major constraints. In particular, it will be imperative that the techniques used for laboratory analyses of these exceedingly precious samples are both (3) non-intrusive and (4) nondestructive (i.e., that they in no way alter the studied samples).
Given the forgoing, the techniques to be used for laboratory-based paleontological studies of Mars rocks will be required, at a minimum, to be capable of establishing (1) the cellularity and (2) carbonaceous composition of any putative “lifelike” forms detected, and to do so using methods that are (3) non-intrusive and (4) nondestructive. All four of the techniques used to document the gypsum-permineralized microbial assemblages reported here—transmitted white light and plane polarized light microscopy, CLSM, and Raman spectroscopy—meet these requirements. A fifth technique, secondary ion mass spectroscopy, can be similarly useful in establishing the carbonaceous (as well as the elemental isotopic) composition of individual cellular microscopic fossils (e.g., House et al., 2000; Oehler et al., 2006, 2009, 2010; Williford et al., personal communication). It remains to be determined, however, whether secondary ion mass spectroscopy (which can be used only on surface-exposed specimens and produces minute surface-altering analytical pits) will meet the highly restrictive requirements that are likely to be set for acceptable studies of samples returned from Mars.
5. Gypsum-Permineralized Microscopic Fossils
5.1. Materials and methods
Fossiliferous samples
The seven microfossiliferous samples studied, shown by their sedimentological setting and predominantly euhedral crystallinity (Fig. 1) to be bottom-nucleated primary gypsums, are from one Cenozoic geological unit, two Paleozoic geological units, two Recent lacustrine gypsiferous deposits, and two Recent salterns:

Microfossiliferous bottom-nucleated primary gypsum from (
(1) the Late Miocene (Messinian, ∼6 million years old) Vena del Gesso Formation at Monte Tondo Quarry, northeastern Italy (Fig. 1A; GPS coordinates: 44°15′10.88″N, 11°40′4.94″E);
(2) the Late Permian, ∼263 million years old, Seven Rivers Formation at Seven Rivers Borrow Quarry, southeastern New Mexico, USA (Fig. 1B; 32°17′41.46″N, 104°31′53.04″W);
(3) the Late Permian, ∼255 million years old, Salado Formation at Crystalline Phoenix Mine, southeastern New Mexico, USA (Fig. 1C; 32°7′24.43″N, 104°24′41.60″W); and Recent gypsum deposits at
(4) Pernatty Lagoon, Mount Gunson, south-central South Australia, Australia (Fig. 1D; 31°30′21.50″S, 137°14′16.00″E);
(5) Chinkapoole Lake, Swan Hill, northeastern Victoria, Australia (Fig. 1E; 35°10′57.78″S, 142°48′6.39″E);
(6) Saltern Pond #9, Exportadora de Sal, Guerrero Negro, northwestern Baja California Sur, Mexico (Fig. 1F; 27°56′35.37″N, 114°2′49.24″W); and
(7) a saltern pond at Salinas de Otomo, Pisco Province, Ica Region, west-central Peru (Fig. 1G; 13°59′52.33″S, 76°14′3.80″W).
Modern sulfur bacteria
The filamentous sulfuretum bacteria illustrated here (Figs. 4V–Y, 5E–G, 5M, 5N) are live specimens obtained from core samples acquired at the sediment-water interface from sublittoral depths (<200 m) off central Chile (∼36°S). Compared with certain of the gypsum-permineralized microfossils, these sulfuretum microbes are “macrobacteria” as defined by Gallardo and Espinoza (2007a, 2007b) and were shown by them to be widespread along the southeastern Pacific continental shelf in oxygen-deficient, sulfide-, sulfate-, and organic-rich sediments below the oxygen minimum zone. The sulfuretum-containing cores were collected with either a custom-built monocorer or a Mini-Multiple Corer MC 600 (Oktopus Gmbh, Kiel, Germany).
Optical microscopy
For studies by optical microscopy at University of California, Los Angeles, the uncovered thin sections were veneered by a thin (∼1 μm thick) layer of fluorescence-free microscopy immersion oil (R.P. Cargille Laboratories Inc., Cedar Grove, NJ, USA). Photomicrographs, both in transmitted white and plane polarized light, were acquired with either a Leitz Orthoplan biological microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA), equipped with an Olympus DP12 digital microscope camera (Olympus America, Melville, NY, USA), or a Leitz Orthoplan 2 biological microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA), equipped with a Nikon DS-5M-L1 digital microscope camera (Nikon Instruments Inc., Melville, NY, USA). At Universidad de Concepción, Chile, transmitted white light and phase contrast photomicrographs of sulfuretum bacteria were acquired by use of a trinocular Zeiss A1 Axio Imager microscope (Carl Ziess Inc./W. Reichman y Cia. Ltda., Santiago, Chile) equipped with a binocular phototube, Zeiss Plan-Neofluar objectives, and a Canon PowerShot G6 digital camera (W. Reichman y Cia. Ltda., Santiago, Chile).
Confocal laser scanning microscopy
CLSM studies were carried out by use of an Olympus Fluoview 300 confocal laser scanning biological microscope system equipped with two Melles Griot lasers, a 488 nm 20 mW output argon ion laser, and a 633 nm 10 mW output helium-neon laser (Melles Griot, Carlsbad, CA, USA). Images were acquired with use of the fluorescence-free microscopy immersion oil noted above and a 60× oil-immersion objective (numerical aperture 1.4) with filters in the light path, to remove wavelengths <510 nm (for 488 nm laser excitation) and <660 nm (for 633 nm laser excitation) from the laser-induced fluorescence emitted by the specimens, and the Olympus “Protocol Processor,” to maximize useful data throughout the specimen. To provide maximum spatial information, the images were deconvoluted by use of the computer program Huygens Essential v3.2 (Scientific Volume Imaging Inc., the Netherlands) and subsequently processed by use of the VolView v2.0 three-dimensional-rendering computer program (Kitware Inc., Clifton Park, NY, USA) that permits their manipulation in three dimensions (Schopf et al., 2006).
Raman spectroscopy
Raman spectra and images of the fossils and associated minerals were obtained by use of a T64000 (JY Horiba, Edison, NJ, USA) triple-stage laser-Raman system that has macro-Raman and confocal micro-Raman capabilities. This system permitted acquisition both of point spectra and of Raman images that display the two-dimensional spatial distribution of molecular-structural components of the specimens and their associated matrix, with the varying intensities in such images corresponding to the relative concentrations of the molecular structures detected. Due to the confocal capability of this system, use of a 50× objective (having an extended working distance of 10.6 mm and a numerical aperture of 0.5) provided a horizontal resolution of ∼1.5 μm and a vertical resolution of 2–3 μm, and use of a 100× objective (working distance 3.4 mm, numerical aperture 0.8) provided a horizontal resolution of <1 μm and a vertical resolution of ∼1 μm. A Coherent Innova (Santa Clara, CA, USA) argon ion laser provided excitation at 351.1 and 457.9 nm, which permitted data to be obtained over a range from ∼300 to ∼3000 cm−1 by use of a single spectral window centered at 1800 cm−1. Thus, and despite the strong fluorescence of the geochemically highly immature kerogen analyzed (Fig. 7), Raman spectra could be acquired simultaneously of the major bands (at ∼1365 and ∼1604 cm−1) of the kerogen comprising the fossils (e.g., Figs. 2A–H, 3J–O, 7) as well as of the major band of diatom-comprising quartz (at ∼466 cm−1; e.g., Figs. 3O, 7) and the major (at ∼1006 cm−1) and minor bands (e.g., at ∼492 cm−1) of the fossil-embedding gypsum matrix (e.g., Figs. 3M, 7).

Optical photomicrographs (white light: A, C, D, I, K, N, O; plane polarized light: B, E, F, L) and confocal laser scanning microscope (CLSM) images (G, H, J, M, P

Optical photomicrographs (white light: A, C, E–J, P, Q, T, U; plane polarized light: K, R, V), confocal laser scanning microscope (CLSM) images (B, D, L, S, W), and 2-D Raman images (M–O) of permineralized microfossils in petrographic thin sections of primary gypsum from the Late Miocene Vena del Gesso Formation, Italy, compared, in part, with a modern cyanobacterium (E) and modern diatoms (P and T). (
For two-dimensional Raman images (Fig. 3M–O), the specimen-containing thin section was covered by a thin veneer of the fluorescence-free microscopy immersion oil noted above (the presence of which has been shown to have no discernable effect on the Raman spectra acquired; Schopf et al., 2005), and the fossil was centered in the path of the laser beam projected through the microscope of the system. The laser power used was ∼1–8 mW over a ∼1 μm spot, an instrumental configuration well below the threshold that results in radiation damage to such specimens (Schopf et al., 2005).
6. Results
Our survey of gypsiferous deposits revealed that although carbonaceous particulate debris is abundant in some water column-precipitated sedimentary gypsums (e.g., those in Permian strata of the Delaware Basin of West Texas and southwestern New Mexico, USA), such units are generally devoid of cellularly preserved microfossils. In contrast, bottom-nucleated gypsum, precipitated in quiescent settings at the sediment-water interface, can be abundantly fossiliferous. Figure 1 shows specimens of the seven such bottom-nucleated crystalline gypsums in which we detected permineralized fossil assemblages, all of which include filamentous microbes, most of which we interpret to be benthic components preserved in place, and all but two of which contain fossils of photic-zone microorganisms, such as diatoms and cyanobacterium-like filaments and unicells that have evidently settled into the deposits from the overlying water column.
Discussed in turn, below, are the fossils we detected in gypsiferous units of the Miocene of Italy, the Permian of New Mexico, and Recent lacustrine and saltern deposits of Australia, Mexico, and Peru. Following this, we summarize results of our Raman spectroscopic analyses of these fossils and compare the usefulness of Raman and CLSM for their study.
6.1. Miocene fossils
Diverse microscopic fossils are abundant in primary sedimentary gypsum of the Late Miocene (Messinian) Vena del Gesso Formation at the Monte Tondo Quarry in northeastern Italy (Figs. 1A, 2, 3), the same locality from which gypsum-embedded fossil cyanobacteria have previously been reported (Panieri et al., 2008) and strata of the same age and geographic region as those containing selenite-embedded dragonfly larvae (Schlüter et al., 2003). Present in the bottom-nucleated gypsum are previously unreported broad filamentous fossils (Fig. 2A–H), smaller-diameter filaments (Fig. 2I–M, 2P–R), coccoidal unicells (Fig. 3A–D, 3F–I), and diatoms (Fig. 3J–O, 3Q–S, 3U–W).
Illustrated in Fig. 2A–H are the most abundant of the fossils detected, broad ∼70 μm diameter filaments that occur in interlaced masses, shown by CLSM to exhibit laser-induced fluorescence (Fig. 2G, 2H) consistent with a carbonaceous composition and by plane polarized light microscopy to be gypsum-embedded (Fig. 2B, 2E, 2F) and to pre-date the emplacement of secondary fracture-filling gypsum veinlets (Fig. 2E). Although Panieri et al. (2008) recorded the presence of somewhat similar “spaghetti-like” filament-aggregates interpreted as masses of cyanobacteria that settled into the bottom-nucleated gypsum from photic zone waters, the largest, “30 to 50 μm diameter” filaments reported by these authors are described as “funnel-shaped structures” that differ morphologically from the larger-diameter straight to gently curved cylindrical filaments reported here.
Of the other filamentous fossils that occur in the deposit, two types are here illustrated: (1) exceedingly narrow sinuous microbial filaments, ≤1 μm in diameter (Fig. 2I, 2J), shown by CLSM to be composed of uniseriate strands of minute bead-shaped cells (Fig 2J, arrow); and (2) broader, ∼15 μm diameter sheath-enclosed cellular trichomes (Fig. 2K–M, 2P–R), similar to those of the extant oscillatoriacean cyanobacterium Lyngbya (Fig. 2N, 2O), shown by plane polarized light microscopy (Fig. 2L) to be gypsum-embedded and, especially well by CLSM, to be sheath-enclosed and comprised of uniseriate discoidal cells (Fig. 2M, 2P–R).
Bottom-nucleated gypsum of the Vena del Gesso Formation also contains permineralized unicells (Fig. 3A–D, 3F–I), some similar to the extant chroococcacean cyanobacterium Gloeocapsa (compare Fig. 3A with 3E), and diatoms (Fig. 3J–O, 3Q–S, 3U–W) similar to members of the modern genus Navicula (compare Fig. 3J–L with 3P, and 3Q–S with 3T) that may have been benthic or may have settled into the bottom-nucleated gypsum from the overlying water column. That such fossils are permineralized in the primary gypsum is shown by plane polarized light microscopy (Fig. 3K, 3R, 3V); the gypsum composition of the embedding matrix (Fig. 3M) and the siliceous composition of a permineralized diatom (Fig. 3O) are shown by two-dimensional Raman imagery and Raman spectra (Fig. 7); and the carbonaceous, kerogenous composition of the fossils is shown by two-dimensional Raman (Fig. 3N) supported by three-dimensional confocal laser scanning microscope images (Fig. 3B, 3D, 3L, 3S, 3W).
6.2. Permian fossils
As studied to date, the samples of bottom-nucleated gypsum from the Late Permian Seven Rivers (Figs. 1B, 4A–C) and Salado Formations (Figs. 1C, 4D–L) of southeastern New Mexico, USA, are less fossiliferous than those of the Messinian of Italy, discussed above. Present in gypsum of the Seven Rivers Formation are ∼10 μm diameter filaments composed of what we interpret to be elongate cells (Fig. 4A–C, arrows) similar to those of a modern sulfuretum bacterium (Fig. 4Y). Primary gypsum of the Salado Formation contains short, narrow, needle-like filaments (Fig. 4D–F), comparable to those of modern sulfuretum bacteria (compare Fig. 4D–F with 4V–X), as well as spheroidal, presumably planktonic unicells <10 μm in size (Fig. 4G–L). That these fossils are permineralized in gypsum is shown by plane polarized light microscopy (Fig. 4B, 4E, 4H, 4K), and their optically identified carbonaceous, kerogenous composition is consistent with their laser-induced fluorescence recorded in confocal laser scanning microscope images (Fig. 4C, 4F, 4I, 4L).

Optical photomicrographs (white light: A, D, G, J, M, P, S, V–Y; plane polarized light: B, E, H, K, N, Q, T) and confocal laser scanning microscope (CLSM) images (C, F, I, L, O, R, U) of permineralized microfossils in petrographic thin sections of primary gypsum from the Late Permian Seven Rivers (
6.3. Recent fossils
The four Recent bottom-nucleated gypsiferous deposits investigated—from the Pernatty Lagoon of south-central South Australia (Fig. 1D); the Chinkapoole Lake of southeastern Australia (Fig. 1E); Saltern Pond #9 at Exportadora de Sal in northwestern Baja, Mexico (Fig. 1F); and a saltern pond at Salinas de Otomo, Pisco Province, west-central Peru (Fig. 1G)—contain both benthic (primarily filamentous) and planktonic (e.g., diatoms) components.
Gypsum of the Pernatty Lagoon contains thin, short, needle-like filaments (Fig. 4M–O) that are similar both to those of the Permian Salado Formation (compare Fig. 4M–O with 4D–F) and similarly narrow modern sulfuretum bacteria (Fig. 4V–X). Such short needle-like filaments occur also in the Chinkapoole Lake gypsum (Fig. 4P–R) together with capitate diatom frustules (Fig. 4S–U) comparable to those of extant Achantinium spp. (compare Fig. 4S–U with 4Z, 4AA), as well as other modern diatoms (e.g., Geissleria decussis, Navicula kotschyi, and Placoneis clementis). Gypsum from Saltern Pond #9 in Baja, Mexico, contains straight to sinuous, narrow, ∼1.5 and <1 μm diameter uniseriate filaments composed of bead-shaped cells (Fig. 5A–D, 5H–L) that are similar to narrow modern filamentous sulfuretum microbes (Fig. 5E–G, 5M, 5N). And gypsum of the saltern pond at Salinas de Otomo, Peru, contains narrow, branched, actinomycete-like filamentous microbes composed of minute bead-shaped cells (Fig. 6). The gypsum permineralization of these fossilized Recent microbes is shown by plane polarized light microscopy (Figs. 4N, 4Q, 4T, 5K, 6B), and their optically identified carbonaceous, kerogenous composition is substantiated by confocal laser scanning microscope images (Figs. 4O, 4R, 4U; 5B–D, 5I, 5L, 6C, 6D).

Optical photomicrographs (white light: A, E–H, J, M, N; plane polarized light: K) and confocal laser scanning microscope (CLSM) images (B–D, I, L) of permineralized microfossils in petrographic thin sections of Recent primary gypsum from Saltern Pond #9 at Guerrero Negro, Baja, Mexico, compared with modern filamentous sulfur bacteria from sublittoral sediments off central Chile (E–G, M, N; cf. Gallardo and Espinoza, 2007a). (

Optical photomicrographs (white light:
6.4. Relative usefulness of Raman spectroscopy and CLSM
As has been established for many quartz-permineralized Precambrian microscopic fossils—ranging in age from ∼650 Ma (Schopf et al., 2005) to ∼3,465 Ma (Schopf et al., 2007)—Raman spectra and, particularly informatively, Raman imagery (Schopf et al., 2005; Schopf, 2009; Schopf and Kudryavtsev, 2010) can provide direct evidence, acquired in situ, of the composition both of permineralized fossils and their embedding mineral matrices (Figs. 3M–O, 7). However, in gypsiferous deposits that contain highly immature kerogenous material, such as the fossiliferous gypsum samples studied here from the Miocene of Italy (Figs. 2, 3) and Recent lacustrine and saltern deposits (Figs. 4 –6), the strong fluorescence of the preserved organics can obscure (Fig. 7A, 7B) or mask completely the very much weaker Raman signal both of the fossils and the minerals that comprise the fossil-embedding matrix. As is shown above, for such samples plane polarized light microscopy can be used to document the permineralization and indigenousness of the fossils detected (Figs. 2B, 2E, 2F, 2L, 3K, 3R, 3V, 4B, 4E, 4H, 4K, 4N, 4Q, 4T, 5K, 6B).
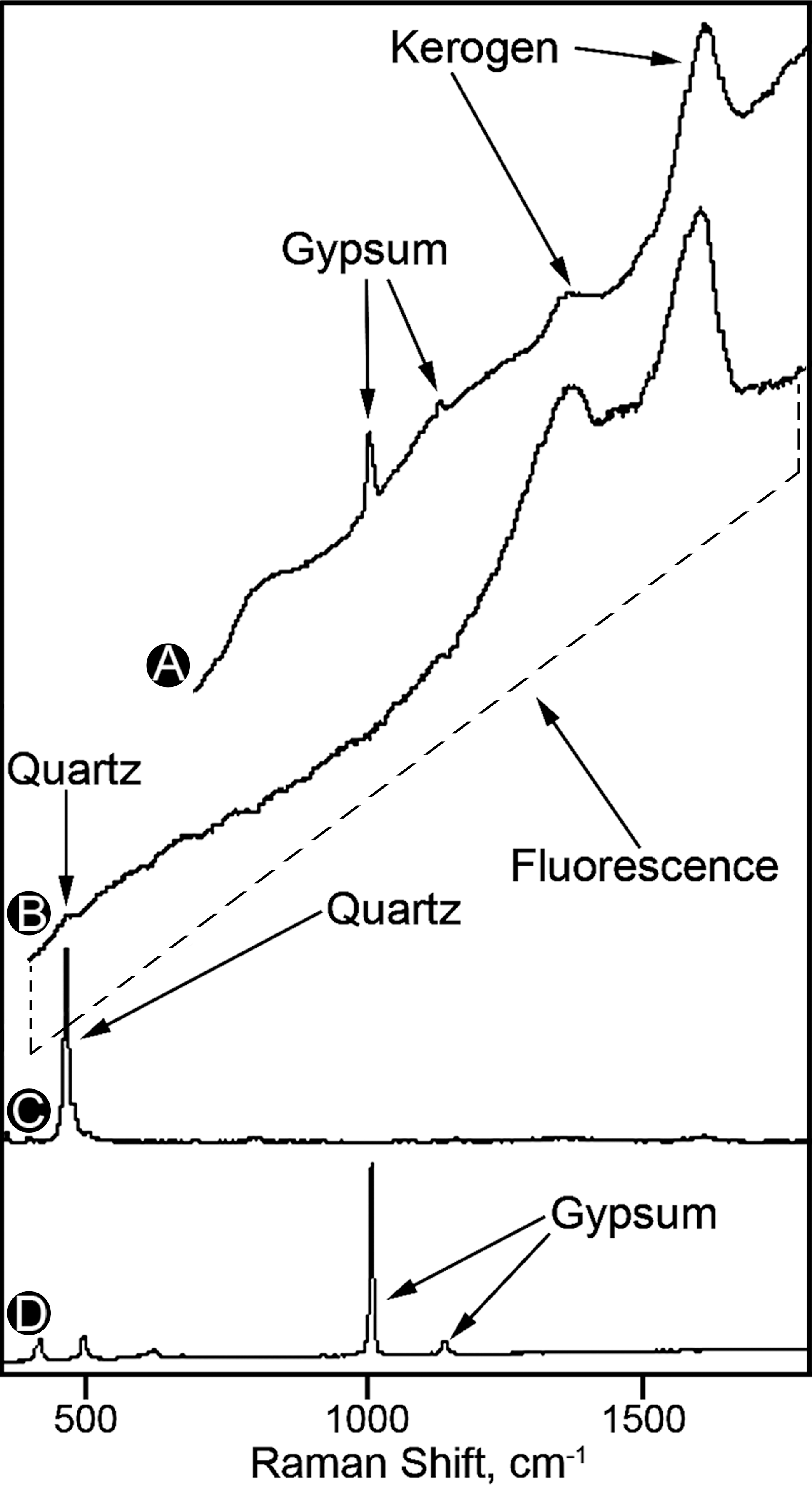
Raman spectra of the kerogen of a broad microbial filament (
It is useful to note that data acquired in studies of microscopic fossils by use of Raman and CLSM are complementary, Raman providing direct evidence of the kerogenous composition of the fossils, and the fluorescence-based images of CLSM supporting such analyses. Moreover, for most organic-walled microscopic fossils, the ranges of kerogen maturity that define the usefulness of the two techniques overlap, which makes it possible to use both techniques to analyze the same individual specimen (e.g., Schopf et al., 2005, 2010c; Schopf and Kudryavtsev, 2010).
Despite the similarities of Raman and CLSM, however, the two techniques have different limitations. The geochemical maturation of the biochemically produced organic matter of living systems to the carbonaceous kerogen preserved in the rock record involves the alteration of the organics into an increasingly planar array of interlinked polycyclic aromatic hydrocarbon–dominated macromolecules, as the kerogen progressively matures along a geochemical pathway toward graphite (Vandenbroucke and Largeau, 2007). Because of the fluorescence inherent in geochemically highly immature organics, Raman is not nearly as useful for analyses of such kerogens as it is for geologically older, somewhat more altered carbonaceous matter, a deficiency that can be partly offset by the use of UV rather than visible Raman spectroscopy (Czaja et al., 2009). In contrast, due to the dependence of CLSM on detection of the laser-induced fluorescence emitted by the fossil-comprising organic matter and the diminution of such fluorescence that accompanies the geochemical maturation of kerogen, this technique yields best results for relatively better preserved, geochemically less than fully mature, kerogenous materials (such as those of the Miocene and Recent units studied here).
In sum, (1) Raman, shown to provide useful data for even partially graphitized individual microscopic fossils (Schopf et al., 2005), is best applicable to fossils composed of kerogen that is sufficiently mature to exhibit a relatively low level of laser-induced fluorescence; and (2) CLSM can provide excellent images of microbial fossils composed of geochemically immature organics (e.g., Figs. 2G, 2H, 2J, 2M, 2P–R, 3B, 3D, 3L, 3S, 3W, 4C, 4F, 4I, 4L, 4O, 4R, 4U, 5B–D, 5I, 5L, 6C, 6D) but is not applicable to carbonaceous fossils preserved in relatively highly metamorphosed deposits, those of middle-greenschist or more severely altered terrains, the kerogen of which typically has an H/C ratio ≤0.25 (Schopf et al., 2005).
7. Discussion
The affinities of the broad, ∼70 μm diameter microbial filaments recorded here from the Miocene Vena del Gesso Formation (Fig. 2A–H) are uncertain. Although the vast majority of modern cyanobacterial filaments are of much smaller diameter (for 447 species and strains of extant oscillatoriaceans, a more than 10 times smaller average diameter of 6.7 μm; Schopf, 1992), large-diameter ecophenes of one modern oscillatoriacean cyanobacterium Lyngbya majuscula, including specimens up to 100 μm broad, have been reported (Demoulin and Janssen, 1981). Thus, if the aggregated masses of broad filaments present in the deposit represent photic-zone cyanobacteria that settled into the bottom-nucleated gypsum, as suggested by Panieri et al. (2008), these forms may represent remnants of L. majuscula–dominated cyanobacterial blooms.
Nevertheless, given the abundance of the extensive interlaced masses of especially broad (∼70 μm diameter) filaments in the bottom-nucleated gypsum we have studied, and their preservation in this highly saline, sulfate-rich and presumably sub-photic-zone setting, we regard it as more likely that they represent remnants of a microbial biocoenose permineralized in place. This interpretation is supported by the occurrence in centimeter-sized samples of the Miocene gypsum of loosely intermeshed subparallel groups of these broad filaments oriented more or less perpendicular to the bedding, a distribution that suggests their preservation in growth position and is similar to that of the modern filamentous sulfur bacterium Thioploca in comparably sulfate-rich benthic habitats (Huettel et al., 1996; cf. Gallardo and Espinoza, 2007a, Fig. 10). We therefore regard it as probable that these fossils are allied with such modern non-photosynthetic sulfuretum bacteria as Thioploca, which includes morphotypes of similarly large diameter (Gallardo and Espinoza, 2007b), and Beggiatoa, which includes strains as broad as 120 μm (Nelson et al., 1989).
Similarly, because metabolically diverse modern microbes of essentially identical morphologies can exhibit markedly differing bioenergetic capabilities—an example of “microbial mimicry” (Schopf, 1994, 1999)—the affinities and, thus, the metabolism of the other filamentous benthic components of these assemblages is difficult to establish on the basis of morphology alone. The ∼10 μm diameter, elongate celled-filaments of the Permian Seven Rivers Formation (e.g., Fig. 4A–C) may represent an exception to this generalization, since the elongate, rather than discoidal, cells exhibited by these fossils are very rare in modern photosynthetic microbes of comparable diameter (Demoulin and Janssen, 1981; Schopf, 1992) but are well established to occur in similarly ∼10 μm diameter sulfuretum-inhabiting non-photosynthetic filamentous microorganisms (compare Fig. 4A–C with 4Y).
Perhaps the most striking result of this study—other than discovery of the widespread presence of permineralized microscopic fossils in bottom-nucleated gypsum, established here for the first time—is the morphological similarity of the majority of the filamentous fossils detected in each of the investigated seven gypsiferous units to components of the extant sulfuretum biocoenose recently described by Gallardo and Espinoza (2007a, 2007b). Despite the problems posed by microbial mimicry, this notable similarity—in organismal and cellular morphology, as well as the presence both of the fossil assemblages and the modern sulfuretum biota in quiescent, sublittoral, sulfate-rich sediment-water interface habitats—indicates that the benthic filamentous components of the fossilized assemblages are plausibly interpreted as bacteria like those of the modern sulfuretum. Given this, it seems probable that, like the metabolically complex modern sulfuretum, the bioenergetics of the bottom-inhabiting components of the gypsum-preserved assemblages involved an interplay of multiple redox processes driven by sulfate-reducing bacteria that generated H2S used by the benthic filamentous sulfur-oxidizing members of these microbiotas as the electron donor for anaerobic oxidative phosphorylation.
The evidence presented thus suggests that the filamentous benthic microorganisms fossilized in these bottom-nucleated gypsums are members of a sulfuretum consortium that has remained largely unchanged over hundreds of millions of years, an example of the evolutionary stasis (“hypobradytely”) of microbial prokaryotes first documented for cyanobacteria (Schopf, 1968, 1994). However, in comparison with cyanobacteria—aerobic photoautotrophs that over the Precambrian adapted to a gradually evolving photic-zone environment—non-photosynthetic sulfuretum communities inhabit an oxygen-deficient, sulfide-rich, quiescent deep-water setting that has remained unchanged over billions of years. We therefore imagine that such communities may have been extant since early in the Precambrian, as suggested by Gallardo and Espinoza (2007a; cf. Van Kranendonk et al., 2012), and that this evidently evolutionarily stable, early-evolved, non-photosynthetic biota therefore represents a promising target in the search for past (or present) life on other planets.
8. Conclusions
The results reported here of studies of seven gypsiferous units—from the Miocene of Italy; the Permian of New Mexico, USA; and Recent deposits from Australia, Mexico and Peru—establish that diverse microscopic organisms are permineralized in bottom-nucleated gypsum deposits. These assemblages include both benthic components, primarily filaments preserved in situ, and planktonic microbes, such as diatoms and cyanobacterium-like filaments and unicells, that have settled into the precipitating gypsum from overlying photic-zone waters. Similar deposits of evaporitic salts, including gypsum, are known to be abundant on Mars. The findings reported here, of obvious relevance to the search for evidence of past life on Mars, lead us to suggest that the non-intrusive, nondestructive analytical techniques used here should also be applied to investigate samples acquired by future Mars sample return missions.
Finally, we note that NASA's Mars Science Laboratory, equipped with the Sample Analysis at Mars (SAM) suite of instruments, is scheduled to land on Mars at Gale Crater in August 2012. Following analyses by the Chemistry and Mineralogy (CheMin) instrument, powdered samples of surface-collected rocks are to be transferred to SAM for, among other investigations, mass spectrometric analysis of organic components. SAM is not designed to identify fossil microorganisms and thus “prove” the existence of past life on the planet—a finding that we believe, if based on the detection of fossilized microorganisms, would require thorough laboratory-based studies of rocks acquired by Mars sample return missions. But, as we report here, preserved carbonaceous matter can be abundant in deposits of sedimentary gypsum, including those that lack discernible microscopic fossils. Because biology is the only known efficient producer of copious amounts of carbonaceous matter, we are confident that the identification by SAM of organic matter in martian sulfate deposits (cf. Aubrey et al., 2005) would serve as a “smoking gun” and provide compelling impetus for further detailed astrobiological investigations of Mars.
Footnotes
Acknowledgments
We thank J. Shen-Miller, Malcolm Walter, and two anonymous reviewers of our manuscript for helpful suggestions. The work of J.W.S., I.S.F., and A.B.K. was supported at UCLA by the Center for the Study of Evolution and the Origin of Life. Additional support was provided, for J.W.S., by the Australian Centre for Astrobiology at the University of New South Wales; for J.D.F., by grants from the NASA Astrobiology and Mars Exploration programs; for I.S.F., by the UCLA Department of Earth and Space Sciences and the Natural Sciences and Engineering Research Council of Canada; for A.B.K., by the Penn State NAI Astrobiology Research Center; and for V.A.G. and C.E., by Chile's FONDECYT Project 1110786, the University of Concepción DIUC Project 210.112.103-1.0, and the Census of Marine Life Program (
Author Disclosure Statement
No competing financial interests exist for any of the authors.
Abbreviations
CLSM, confocal laser scanning microscopy; SAM, Sample Analysis at Mars.