
Abstract
More frequent events of drought are predicted to happen in the future, but our ability to predict the effect on the biota may be limited by our partial understanding of extremophiles. Among the few animals that are able to survive in the absence of water for long periods of time are rotifers, tardigrades, and nematodes. Here, we take advantage of lichen collections stored dry at ambient temperature and humidity for years in museums, and through statistical modeling we demonstrate that the survival rates over time do not differ among animal groups but are strongly influenced by the type of substrate (the different lichen species). Our results suggest that desiccated organisms are prone to irreversible damage to biological structures, independently of the different biochemical processes involved in desiccation tolerance by different animals. The influence of the environment overcomes any taxon-specific response to survive extreme droughts. The predicted ability to survive for up to 10 years while desiccated enables these organisms to achieve potential global distributions, endurance against parasites, and even survival when exposed to outer space. Key Words: Anhydrobiosis—Dormancy—Nematoda—Rotifera—Tardigrada. Astrobiology 12, 863–869.
1. Introduction
R
Notwithstanding the interest that these extremophiles are attracting from scientists, especially because of the potential applications of the biochemical mechanisms to survive without water, most questions remain unanswered. For example, it is not clear whether these organisms can survive without water for decennia or centuries, or for shorter times only. The very few available data on their long-term survival are mostly anecdotal, given that experiments on survival can only be performed over timescales of a few years, not for tens of years (Alpert, 2005, 2006). More information is present for nematodes, especially for the plant parasitic ones, given their economic interest; living juveniles have hatched from dormant eggs and cysts kept dry for more than 30 years (Antoniou, 1989; Watanabe, 2006). Little is known about long-term survival of desiccated free-living animals that are of no economic importance; the longest unambiguous survival times for adult animals are those of tardigrades, which are from 4 to 20 years (Bertolani et al., 2004; Rebecchi et al., 2006; Jørgensen et al., 2007), and bdelloid rotifers, from 2.5 to 9 years (Rahm, 1923; Guidetti and Jönsson, 2002).
The aim of this investigation was to provide a further contribution to the evidence for long-term survival in dormancy and verify the ability of direct recovery by adult dormant animals. Understanding long-term survival in these organisms will be important to assessment of the impact of future droughts on ecosystems. We analyzed samples that were kept dry for periods from less than 1 month to ∼150 years and then investigated the rate at which microscopic animals lose their ability to recover after long-term desiccation. We thus obtained statistical models describing survival over time. These models, based on measured and estimated parameters, allow for prediction of the maximum expectations for long-term survival, which, until now, have only been based on anecdotal evidence or single-case reports. The statistical models also allow for prediction of differences in survival rates among different animals that employ different biochemical strategies to withstand lack of water.
2. Materials and Methods
2.1. Sampling long-term desiccated animals
Setting up an experiment to keep animals desiccated for many years would not be realistic, and the results of such experiments would be available only after many years. However, a reliable way to obtain animals kept dry for long periods of time is to consider dry collections of lichens and mosses in museums. Unfortunately, most museums in the past used chemicals to protect samples from parasites, which can potentially kill the desiccated dormant microscopic animals. To avoid this problem, we focused on the historical collection of Swedish lichens of the Swedish Museum of Natural History in Stockholm, where the last protective treatment was performed in 2006 by fumigation with methyl bromide. This particular pesticide is believed not to harm dormant rotifers and tardigrades (Guidetti and Jönsson, 2002), though prolonged desiccation may increase vulnerability of dormant tardigrades to methyl bromide treatments (Jönsson and Guidetti, 2001).
Lichen samples have been kept dry in the museum in closed paper envelopes since the date of collection. We carefully opened the envelopes with the lichen samples, cut an area of 1–2 cm2 from each of them, placed it into a Petri dish, and sprayed Millipore distilled water around the sample but not directly on it before covering the dish. This procedure allows humidity to increase gradually, mimicking natural processes. Moreover, it diminishes the damage to the dormant animals caused by direct immersion in water (Guidetti and Jönsson, 2002). After 3–4 hours, Millipore distilled water was poured into the Petri dish to allow the complete recovery of the dormant animals.
After 1 hour from complete hydration, we started isolating all animals, both alive and contracted, and placed them in embryo dishes with Millipore distilled water and a few drops of the water medium from their original sample. Animals were considered “alive” only when their movement was coordinated; small contractions of the body in animals that exhibited no other movement were not considered sufficient for successful recovery. Alive animals were counted and removed from the embryo dish; contracted organisms were kept in the embryo dish for 5 days, rinsing them every 2 days to allow for proper oxygenation of the water. During these 5 days, the Petri dishes with the original samples were monitored at least once a day, and additional animals moved to the embryo dishes for the final count. No food was added to the samples. We counted how many live and dead individuals of tardigrades, nematodes, and bdelloid rotifers were found in each sample. Animals that did not recover after 5 days were considered dead; animals usually recover within a few hours, but to provide additional evidence that the contracted animals were dead, approximately 150 bdelloids, 100 tardigrades, and 50 nematodes were checked at optical high magnification (up to 1000×) to confirm that the internal anatomy was degraded. For the animals that recovered, we identified to species level all the bdelloid rotifers.
We focused on 117 samples of dry specimens of three species of lichens in the collection of the Swedish Museum of Natural History, namely, Hypogymnia physodes, Parmelia saxatilis, and Xanthoria parietina, which were collected in Sweden from 1852 to 2011. We already knew the composition of the species assemblages of bdelloid rotifers living in these lichens throughout Sweden (Fontaneto et al., 2011), and we could thus assess whether only a restricted subset of bdelloid species are capable of long-term survival.
To minimize the possibility of contamination of the old dry samples during the laboratory procedures, all laboratory tools (e.g., pipettes, scissors, tweezers, dishes) and the Millipore distilled water were sterilized under UV light prior to use.
Moreover, to discern whether the capacity for long-term survival of dormant animals in lichens preserved at the Swedish Museum of Natural History is unique or biased by the way samples were kept, we analyzed additional samples preserved in the private collection of Ulf Swenson, Knivsta, Sweden (four samples, from 1987 to 1999), and in the Museum of Evolution in Uppsala, Sweden (five samples, from 2003 to 2008). These additional collections had never been subjected to chemical treatments.
2.2. Statistical analyses
A generalized linear model was used to assess the influence of different variables on the percentage of alive individuals out of the total number of individuals counted in each lichen. The explanatory variables included (1) the age of the sample as a continuous variable, (2) the taxonomic group as a categorical variable with three levels (tardigrades, nematodes, and bdelloid rotifers), and (3) the substrate species of lichen as a categorical variable with three levels for the three species (H. physodes, P. saxatilis, and X. parietina). A logit link and quasibinomial error structure to account for overdispersion were applied. Model comparison was conducted by an analysis of deviance test that follows an F distribution for overdispersed data (Venables and Ripley, 2002). Models were compared with and without the explanatory variables age, taxonomic group, and lichen species; only variables that improved the deviance significantly (with use of an F test) remained in the final model. All analyses were performed in R.14.0 (R Development Core Team, 2011).
3. Results
Out of the 117 lichen samples from the Swedish Museum of Natural History, 9456 animals were counted (Supplementary File S1; Supplementary Data are available online at
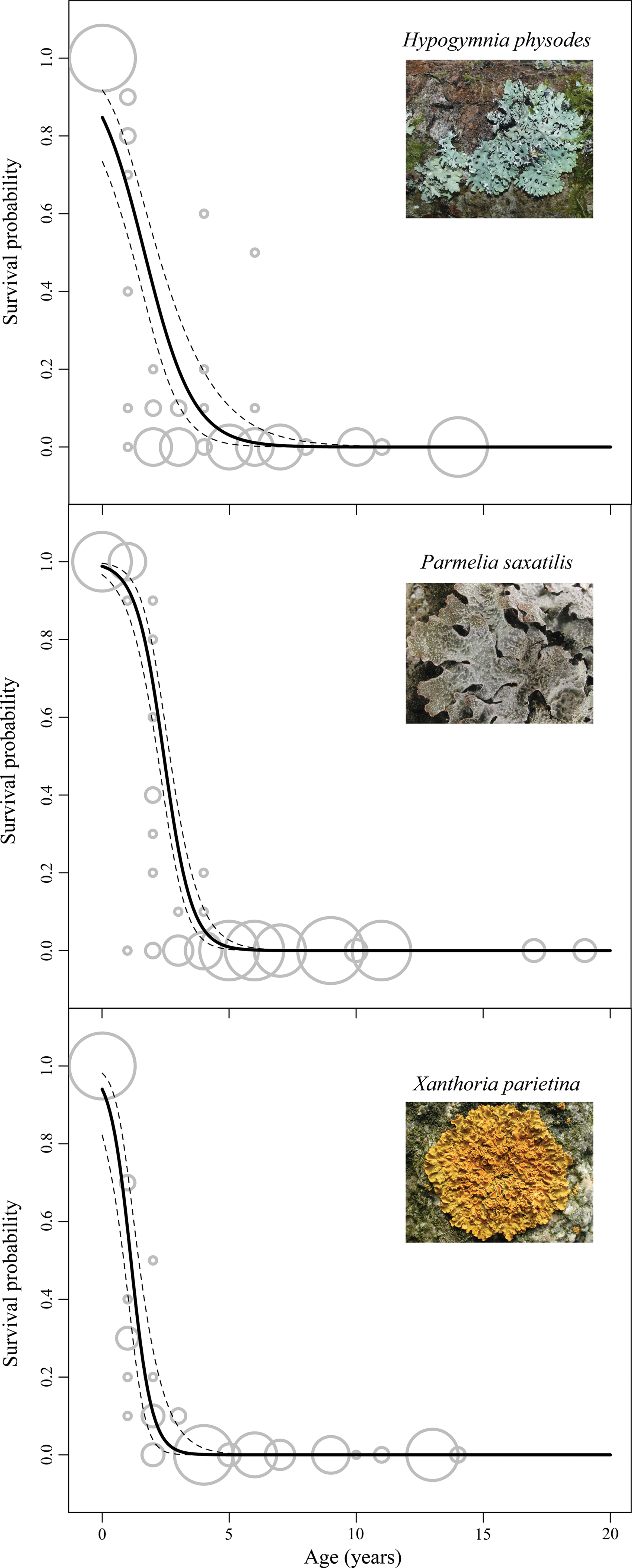
Estimated survival curves of desiccated microscopic animals in the three lichen species. Dashed lines indicate 95% confidence intervals. Curves are limited to the first 20 years of desiccation. Original data on the proportion of animals that recovered are superimposed as gray circles, with diameter proportional to the number of animals. Photos of H. physodes and X. parietina from Wikipedia, freely available under a Creative Commons license [
A significant increase in the proportion of dead animals was found over time (Fig. 1, Table 1), but surprisingly, the analysis provided no evidence of differences in survival rates among the three groups of animals (F 215,2=1.56, p=0.21). On the other hand, the species of lichen had a significant effect on survival, and also its interaction with age was significant (Table 1). The model predicted that, after one year of desiccation, animals survived significantly better in P. saxatilis (98.8%, confidence interval 96.6–99.6) than in H. physodes (84.7%, 73.4–91.7), but there was no significant difference between P. saxatilis and X. parietina (94.0%, 82.2–98.1), On the other hand, the predicted long-term survival of animals was better in H. physodes (below 0.1% of recovery after 10 years) than in P. saxatilis (below 0.1% after 8 years) or X. parietina (below 0.1% after 5 years) (Supplementary File S2).
Generalized linear model with logit link and quasibinomial error structure were used to model the effect of three lichen species (Hypogymnia physodes, Parmelia saxatilis, Xanthoria parietina) and of time remained desiccated (age) on the survival probability of bdelloid rotifers, tardigrades, and nematodes. The survival probability was not significantly different for the three groups (see main text for significance tests).
The other two data sets we analyzed (from collections in Uppsala and Knivsta) were considerably smaller (9 lichens and 1286 animals, Supplementary File S3) but confirmed the same results: in these additional data sets no animals recovered from the lichens kept dry for more than 4 years. These lichen collections have never been fumigated, and this confirms the generality of the trend described from the collection of the museum in Stockholm.
Among the bdelloid rotifers that successfully recovered, 13 species could be identified (Adineta tuberculosa from 2005 and 2007; A. vaga from 2010; Didymodactylos carnosus from 2007; Habrotrocha constricta from 2008, 2009, and 2010; H. pulchra from 2006, 2008, 2009, and 2010; H. spicula from 2007 and 2008; H. sp. from 2007; Macrotrachela ehrenbergi from 2009; M. musculosa from 2007; M. plicata from 2008 and 2010; Mniobia russeola from 2005, 2007, 2008, 2009, and 2010; Mn. scabrosa from 2005 and 2007; and Mn. scarlatina from 2006, 2007, 2008, and 2010), all of which well represent the most common and abundant bdelloid species known to live in Sweden in the three analyzed lichen species.
4. Discussion
The most important implications of our results are that (1) microscopic animals have the ability to withstand long-term desiccation for years while kept desiccated at room temperature and room relative humidity; (2) nematodes, tardigrades, and bdelloid rotifers do not differ in their abilities; but (3) the substrate (lichen species) influences their survival.
4.1. Long-term survival
Microscopic animals were able to recover after up to 6 years in a desiccated state in our data set and plausibly up to more than 10 years, according to the predictions of the model. Such estimates are in accordance with the longest survival recorded, 9 years (Guidetti and Jönsson, 2002; Rebecchi et al., 2009b). Survival in a desiccated state up to 10 years is a long time in comparison to the life span of these organisms, which is measured in weeks or months (Ricci et al., 2007; Hengherr et al., 2008; Lv et al., 2010). Nevertheless, 10 years for dormant adults is a much shorter time than the decades that resting eggs of monogonont rotifers and cladoceran crustaceans can withstand (Cáceres, 1998; Piscia et al., 2012). Similar to cysts in protists (Foissner, 2011), such resting stages are embryos in a state of arrested development (Boschetti et al., 2011a), which is specifically produced to withstand adverse conditions for long periods of time, and these embryos are equipped with thick external layers that keep them isolated from oxidation and other external processes that could interfere with the metabolism (Wallace et al., 2006). In our study, the organisms that recovered from dry lichens did not come from specifically produced resting eggs or cysts with thick envelopes, but the same organisms that entered the desiccated dormant stage were able to recover and continue their life. For these anhydrobiotic animals that do not possess the thick external layer of the resting eggs to protect them (Marotta et al., 2010), oxidation or other metabolic processes likely influenced their dormant cells and led to shorter survival.
One alternative explanation for the relatively shorter survival time in the analyzed samples, compared to, for example, resting eggs and cysts, could be the potential damage by the fumigation against fungal parasites in the collection of the Swedish Museum of Natural History. Nevertheless, animals from lichens that were already in the collection during the fumigation in 2006 were able to recover, and no threshold effect was seen in the recovery of animals from lichens corresponding to the fumigation in 2006 in our data set. The statistical model is based mostly on data from lichens in which animals did recover, and had been collected and stored from 2006 to 2011, a time during which no fumigation took place. Moreover, the other two smaller collections that we analyzed, which were never exposed to any chemical treatments, confirmed the pattern, with animals recovering from lichens kept dry for less than 4 years and no animals alive from lichens kept dry for longer periods.
4.2. Differences among organisms
The biochemical mechanisms used to survive by anhydrobiotic organisms are not fully understood (Clegg, 2001; Oliver et al., 2005; Tunnacliffe and Wise, 2007), but the main actors are thought to be specific molecules that protect the intra- and extracellular environment against the various damages induced by the lack of water (Crowe et al., 1998; Rebecchi et al., 2007). Sugars such as trehalose and sucrose were, for a time, considered the main molecules involved in the process in plants and animals (Alpert, 2006), but today it is recognized that other molecules are important as well. These include heat-shock proteins, late embryogenesis abundant proteins, chaperones, antioxidants, and others (Goyal et al., 2005; Boschetti et al., 2011b; Grohme et al., 2011; Schokraie et al., 2011; Clark et al., 2012). Trehalose, which is present in nematodes and tardigrades, is absent in bdelloid rotifers (Lapinski and Tunnacliffe, 2003; Caprioli et al., 2004). Moreover, different animals survive desiccation with different mechanisms: bdelloid rotifers and tardigrades seem to stop aging while desiccated [the “Sleeping Beauty” mechanism (Ricci and Covino, 2005; Hengherr et al., 2008)], whereas nematodes seem to age [the “picture of Dorian Gray” mechanism (Ricci and Pagani, 1997)]. It is thus possible that the different processes and metabolic activities during anhydrobiosis will influence long-term survival. Our results show that, in spite of biochemical differences, tardigrades, nematodes, and bdelloid rotifers do not have different long-term survival rates when desiccated.
Within bdelloid rotifers, all the species that recovered are known to be common and widespread in the three lichen species across Sweden (Fontaneto et al., 2011), and no single species continuously recovered from the three lichens. This scenario suggests that differences between organisms do not produce differences in recovery rates in the long term, even at the finer taxonomic scale.
4.3. Effect of lichen species
Interestingly, the species of lichen that acted as a physical substrate for the desiccated animals was a significant predictor of recovery rates. All animals survived longer in H. physodes, and with a wider confidence interval in the estimates for this lichen. This is consistent with the fact that this lichen forms thicker talli than the other two, which may help animals during the process of entering anhydrobiosis. Of course, lichens also have a heightened tolerance for desiccation and may have the capacity to recover metabolic activity after long periods of dehydration (Kappen and Valladares, 2007). No information on recovery rates, however, has been made available for the three species of lichens we used. The delicate process of entering anhydrobiosis involves biochemical, metabolic, and morphological changes, and the rate of water evaporation in time and the lichen species are key elements that influence efficient recovery (Ricci et al., 2003). Thus, the differences in the type of substrate (lichen) may interfere with the differences in the mechanisms allowing desiccation tolerance in different animals. The longest surviving animals (bdelloids and tardigrades) recovered after 9 years and came from a sample of moss and not from our extensive survey of lichens (Guidetti and Jönsson, 2002); mosses may be a better substrate than lichens for long-term desiccation. Such a conclusion strengthens the need for community-based research into interactions between species to predict their responses to climate change (Gilman et al., 2010).
4.4. Eco-evolutionary implications of long-term survival
We could speculate that efficient desiccation tolerance may have surprising effects: if desiccation does induce DNA damage (Neumann et al., 2009; Rebecchi et al., 2009b), the same mechanisms for repairing DNA damage during recovery after desiccation may also be utilized by bdelloid rotifers to successfully repair their DNA after exposure to ionizing radiation (Gladyshev and Meselson, 2008). Moreover, the organisms that survive in anhydrobiosis are the most suitable candidate metazoans to survive under the extreme conditions of outer space: such organisms have been used in experiments on air balloons and on the International Space Station (Ricci et al., 2005; Jönsson et al., 2008; Rebecchi et al., 2009a). Obviously, animals did not adapt to survive ionizing radiation and travel in outer space, and these abilities are a secondary effect of their desiccation tolerance. Surviving drastic climate changes may be another such by-product.
Desiccation tolerance has other consequences for these organisms: desiccated stages can be passively dispersed. Biogeography and macroecology of microscopic animals are debated topics, as there is evidence of cosmopolitan and very widespread patterns of distribution, with no limiting geographic barriers (Artois et al., 2011). For most microscopic animals such as rotifers, tardigrades, and others, latitudinal gradients in diversity seem not to exist (Kotwicki et al., 2005; Fontaneto et al., 2012), and analyses of patterns of diversity reveal significant differences from larger above-ground organisms (Fontaneto et al., 2006; McInnes and Pugh, 2007; Guil et al., 2009; Decaens, 2010). These unusual macroecological properties may be triggered similarly by the long-term survival of desiccated animals, acting as propagules for long-distance dispersal. Given these peculiarities, strictly linked with anhydrobiosis, microscopic animals have been suggested as a model system for experimental biogeography and community ecology (Hortal, 2011).
The fact that desiccated animals can be moved continuously across different spatial scales also allows them to potentially escape parasites. This process, called the hide-and-seek hypothesis, has recently been proposed to explain the fact that asexual bdelloid rotifers can cope with the arms race against parasites even if they cannot evolve quickly (Wilson and Sherman, 2010; Wilson, 2011). Thus, the ability of bdelloid rotifers to withstand desiccation has a far-reaching effect, which allows them of course to survive desiccation but also to win the arms race against parasites, colonize distant areas, survive exposed in outer space, and exist potentially unaffected by global environmental changes.
Footnotes
Acknowledgments
This work was performed thanks to a EU Synthesis grant to D.F. and M.W. Additional sources were used from a NERC-UK grant to D.F. and a Swedish Taxonomy Initiative grant to M.W. We wish to thank Irene Bisang for her logistic help in Stockholm; the Swedish Museum of Natural History for letting us use the lichen collection, Anders Nordin for the samples from the Uppsala collection, and Ulf Swenson for the samples from the Knivsta collection; Chiara Boschetti, Isobel Eyres, Lukasz Szydlowski, and two anonymous reviewers for comments on the manuscript. We thank Anita Stridvall for letting us use the photo of P. saxatilis from the Leif and Anita Stridvall Home Page (
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.