
Abstract
The microbiology of the spacecraft assembly process is of paramount importance to planetary exploration, as the biological contamination that can result from remote-enabled spacecraft carries the potential to impact both life-detection experiments and extraterrestrial evolution. Accordingly, insights into the mechanisms and range of extremotolerance of Acinetobacter radioresistens 50v1, a Gram-negative bacterium isolated from the surface of the preflight Mars Odyssey orbiter, were gained by using a combination of microbiological, enzymatic, and proteomic methods. In summary, A. radioresistens 50v1 displayed a remarkable range of survival against hydrogen peroxide and the sequential exposures of desiccation, vapor and plasma phase hydrogen peroxide, and ultraviolet irradiation. The survival is among the highest reported for non-spore-forming and Gram-negative bacteria and is based upon contributions from the enzyme-based degradation of H2O2 (catalase and alkyl hydroperoxide reductase), energy management (ATP synthase and alcohol dehydrogenase), and modulation of the membrane composition. Together, the biochemical and survival features of A. radioresistens 50v1 support a potential persistence on Mars (given an unintended or planned surface landing of the Mars Odyssey orbiter), which in turn may compromise the scientific integrity of future life-detection missions. Key Words: Planetary protection—Spacecraft assembly facility—Extremophiles—Mars—Microbe. Astrobiology 12, 854–862.
1. Introduction
M
Among the differing types of microorganisms, the detection of non-spore-forming and Gram-negative bacteria in spacecraft assembly facilities has not been widely reported in the literature. However, recent reports in which both culture and molecular techniques were used indicate that Gram-negative bacteria are common to spacecraft assembly facilities, with Acinetobacter among the most abundant within this group (Venkateswaran et al., 2001; La Duc et al., 2003, 2004a). The Acinetobacter are broadly associated with soil, water, and clinical environments, and particular species are known for their multi-antibiotic resistance and hydrocarbon degradation properties (Gerischer, 2008; Peleg et al., 2008). Of particular interest, therefore, is the bacterium Acinetobacter radioresistens 50v1, which was isolated from the surfaces of the Mars Odyssey spacecraft (preflight, just prior to encapsulation and launch) (La Duc et al., 2003) and consequently found to be among the first cultivable, extremotolerant (desiccation, ultraviolet radiation, and hydrogen peroxide), non-spore-forming, and Gram-negative bacteria to be isolated from a spacecraft surface. Nevertheless, there have been no studies to date focused upon the molecular underpinnings of the extremotolerance of A. radioresistens 50v1, despite the potential for this microorganism to serve as a contaminant of extraterrestrial environments.
Hence, biochemical insights into the mechanisms and range of extremotolerance of A. radioresistens 50v1 were gained by utilizing a combined approach of proteomics, enzymology, and microbiology. Our studies on A. radioresistens 50v1 show that the mechanisms of tolerance toward oxidative stress include catalase, alkyl hydroperoxide reductase, and ATP synthase, whereas the survival features are consistent with survival under Mars-like degradative conditions, which thereby suggests the potential for persistence of A. radioresistens 50v1 on Mars, given an unintended landing or the final spacecraft disposition of the Mars Odyssey orbiter.
2. Materials and Methods
2.1. Materials
The Gram-negative and control cell lines of Acinetobacter radioresistens ATCC 43998 and Escherichia coli DH5α were obtained from the American Type Culture Collection (Manassas, VA) and Invitrogen (Carlsbad, CA), respectively. Nonstabilized hydrogen peroxide (H2O2) was utilized in the kinetic and survival experiments to eliminate any potential impacts of the stabilizers (e.g., phenol, acetanilide, and sodium stanate); nonstabilized 30% w/w H2O2 (1.11 g/mL) was obtained from Sigma-Aldrich (St. Louis, MO), immediately aliquoted, stored at −20°C, and assayed for consistent H2O2 concentrations prior to use by absorbance spectroscopy (ɛ=43.6 M −1 cm−1 at 234 nm). Other purchased reagents included bovine liver catalase (Sigma-Aldrich), HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) buffer (VWR), NaCl (VWR), phosphate-buffered saline (PBS; 10×PBS: 100 mM potassium phosphate, 100 mM NaCl, pH 7.4) (VWR). Luria-Bertani (LB) medium broth was prepared (per liter) with 5.0 g of yeast extract (Becton Dickinson, Franklin Lakes, NJ), 10.0 g of tryptone (Becton Dickinson), and 10.0 g of NaCl (VWR). Tryptic soy broth (TSB) and tryptone yeast extract glucose broth (TYG-B) were prepared from pre-mixed stocks (Becton Dickinson). All media were autoclaved at 121°C for 30 min, buffers and solutions were sterile filtered (0.22 μm), and pure water (18 MΩ cm−1) was used throughout.
2.2. Survival in aqueous hydrogen peroxide
The survival of the A. radioresistens 50v1 and ATCC 43998 strains were measured before and after exposure to aqueous H2O2. Culture conditions, similar to those typically used when characterizing spacecraft-associated microorganisms, included LB nutrient broth, an incubation temperature of 32°C, and constant agitation at 200 rpm (La Duc et al., 2003, 2007; Ghosh et al., 2010). Exposures were performed on mid-log phase cultures of both strains (0.9–2×109 cfu/mL) with 100 mM and 320 mM H2O2. After a 1 h exposure with constant agitation (which included vigorous gaseous product evolution), 100 μL aliquots were removed, immediately diluted to 1.0 mL with 100 μg/mL bovine liver catalase in PBS (10 mM potassium phosphate, 10 mM NaCl, pH 7.4), and incubated for 5 min at 22°C. This final quenching step in the catalase solution ensured full degradation of all remaining H2O2 prior to plating. Upon completion, suspensions were decimally diluted with LB (10–10,000 fold) and aliquots of 50 and 100 μL spread onto LB media agar plates. Exposures were performed on three independent samples, plating was performed in triplicate for all samples (including the unexposed and ambient controls), and plates bearing 25–300 colonies after incubation (overnight at 32°C) were enumerated to assess survival (U.S. Food and Drug Administration, 1998).
2.3. Survival under multiple stress conditions
The survival of the A. radioresistens 50v1 and ATCC 43998 strains were also measured after exposures to the multiple stress factors of desiccation, vaporous H2O2, a low-temperature H2O2 plasma, and UV radiation. Mid-log phase cultures (1.0–1.2×109 cfu/mL) of each strain were harvested in PBS, transferred into sterile water, and decimally diluted into 96-well plates by using 50 μL aliquots per well. For the desiccations, the 96-well plates were placed slightly ajar in a bio-hood for 10 days, whereupon the dried plates were exposed to H2O2 (vapor and plasma), UV radiation, and a combination of all stress factors [using the sequence of (1) desiccation, (2) vapor and plasma phase H2O2, and (3) UV radiation].
Exposures to the vapor and plasma phases of H2O2 were performed with a STERRAD 100S sterilization system (Advanced Sterilization Products, Irvine, CA). The 96-well plates were introduced into the STERRAD chamber and subjected to 1–4 cycles of sterilization, where each cycle initially involved vaporization (40 mPa) of injected aqueous H2O2 at the final concentration range of 0.33–3.0 mg/L. All samples were incubated for 12 min within the diffusive vapor of H2O2 and then for an additional 2 min in a radio-frequency-powered plasma of the H2O2 mixture (400 W, 13.56 MHz). Ultraviolet irradiations of the 96-well plates were performed at 1 J m−2 s−1 in the dark with a low-pressure mercury vapor lamp, which emitted predominantly at 254 nm (UV254). Samples were exposed for differing time intervals (25–1000 s) to reveal the impacts of cumulative radiation dosage (25–1000 J m−2). Radiant outputs were measured by a UVX radiometer, which was fitted with a UVX-25 filter and calibrated to a traceable standard from the National Institute of Standards and Technology. All plates were inoculated with either TSB or TYG-B media and incubated at 32°C for 72 h. Microbial survivals were assessed with the most probable numbers method (Kempf et al., 2005), where exposures were performed on three independent samples by using a 10-fold dilution series with eight tubes per dilution. Experimental controls included the 10-day desiccated and unexposed (but similarly treated) samples.
2.4. Native catalase specific activities
The specific activities of hydrogen peroxide degradation were measured by using extracts of A. radioresistens 50v1, A. radioresistens ATCC 43998, and E. coli DH5α. Cultures of each strain were grown in LB media at 32°C with constant agitation (200 rpm). Mid-log phase cells were harvested at 5445g for 10 min at 4°C (Beckman Coulter Allegra 21R centrifuge), washed twice, and the final cell pellets (∼0.5 g wet cells) fully resuspended in 10 mL 50 mM HEPES buffer (pH 7.5) containing 100 mM NaCl (50 mL beaker). While placed in an ice bath, the suspensions were then subjected to a Virsonic 600 ultrasonic cell disrupter for two 30 s timed cycles with a power setting of 5 and an incubation of 30 s (on ice) between the two cycles. Upon completion, the samples were transferred to sterile conical tubes, centrifuged, and the supernatants collected, stored on ice, and immediately analyzed.
Kinetic assays of H2O2 degradation (Beers and Sizer, 1952; Yumoto et al., 2000) were performed on three independent samples with six measurements of rates each. Reaction mixtures contained 20 mM H2O2 in 50 mM HEPES (pH 7.5) with 100 mM NaCl and were initiated with the addition of 100 μL of the extract. All reactions were thoroughly but gently mixed, performed at 22°C, and the changes in absorbance at 240 nm measured every 2 s for a minimum of 30 s/sample (Beckman Coulter DU-640 spectrophotometer). Longer reaction times (>60 s) yielded the formation of millimeter-sized gas bubbles (presumably molecular oxygen), which was consistent with the catalase-like degradation of H2O2. The specific activities of H2O2 degradation (or catalase specific activity by inference) were calculated by using the initial linear rates, a molar absorptivity (ɛ) of 43.6 M −1 cm−1, and total protein contents, which were measured by using a Bio-Rad Protein Assay and bovine serum albumin standards (Bio-Rad, Richmond, CA). Catalase specific activities were expressed in Units/mg protein, where a Unit of activity represented the micromoles of substrate converted per minute.
2.5. Stress-induced catalase specific activities
The impact of exogenous H2O2 on the catalase specific activities of the A. radioresistens 50v1 and ATCC 43998 strains was also measured. Cultures of each strain were grown to mid-log phase in LB media at 32°C and exposed to the final concentrations of 1 mM H2O2 and 490 mM H2O2. Suspensions were agitated in an orbital shaker at 200 rpm at 32°C and sampled 15 min after the exposure for both strains and after 30 and 45 min for the 50v1 strain. At each respective time point, 1 mL aliquots of each culture (including control unexposed cultures) were removed, and the catalase activity from the total culture, the cellular component (i.e., cell pellet), and cleared media were measured. Cell pellet samples were prepared by harvesting at 10,000g for 3 min (Beckman Coulter Microfuge 18), and the supernatants were saved for analysis. Cell pellets were additionally washed by full resuspension in 1.0 mL of 50 mM HEPES (pH 7.5) with 100 mM NaCl, followed by reharvesting and discarding of the supernatant wash. All samples (total culture, cell pellet, and cleared media) were diluted to 10 mL with sterile 50 mM HEPES (pH 7.5) with 100 mM NaCl, and the cell pellet (and total culture) samples ultrasonicated as described. Specific activity measurements were performed as described on the final extracts and diluted cleared media by using three independent samples with at least two measurements of rates at each experimental step.
2.6. Proteomics of hydrogen peroxide–exposed cells
The impact of H2O2 exposure on the proteomes of the A. radioresistens 50v1 and ATCC 43998 strains were measured on three independently exposed samples by two-dimensional polyacrylamide gel electrophoresis and mass spectroscopy. Cultures of each strain (20 mL) were grown to mid-log phase in LB media at 32°C, whereupon 10 mL aliquots were aseptically removed and transferred to sterile 50 mL conical tubes. For the exposures, a final concentration of 1.0 mM H2O2 was added to the cultures and the suspensions gently agitated by inversion every 4–5 min for a total of 15 min at 22°C. Aliquots of each culture were removed before and after the exposure and accordingly referred to as the control and exposed samples (for the proteomics), and treated as a paired set in the ensuing analyses. Samples were centrifuged at 5445g for 10 min at 4°C, and the pellets completely resuspended and washed in PBS a total of three times. Final cell pellets were stored at −80°C; however, prior to storage, cultures were spread onto LB agar plates, incubated overnight at 32°C, and enumerated for colony-forming units (cfu; ∼2×109 cfu/mL for both strains). No impact on the survival of either strain was observed. Preparation of the frozen cells for analysis involved resuspension of the thawed pellet (∼1 g wet cells) in 5.0 mL Bugbuster Master Mix (EMD4Biosciences, USA), followed by addition of 10 μL of Halt* Protease Inhibitor Cocktail (Thermo Scientific, USA) per 1 mL of the extract. Suspensions were gently mixed, incubated at ∼22°C for 20 min, and centrifuged at 5445g for 20 min at 4°C (to remove the insoluble cell debris). The supernatants were concentrated to a range of 4.3–5.0 mg/mL.
Protein extracts were separated by two-dimensional gel electrophoresis and analyzed by mass spectrometry by the Institute for Integrated Research on Materials, Environment and Society (IIRMES) at the California State University, Long Beach. Experimental conditions included 300 μg protein/gel, isoelectric focusing with a pH 3–10 gradient, separation with 12% polyacrylamide gels, staining with Coomassie blue, and imaging and measurement of the protein volumes (of the control and exposed samples) by densitometry with the Progenesis SameSpots analysis software (Nonlinear Dynamics, Ltd., USA). Target proteins were subjected to trypsin digestion, and peptide mass spectra were obtained on an Applied Biosystems 4800 MALDI/TOF mass spectrometer in the MS and MS/MS modes. Spectra were assigned to bacterial sequences in the MSDB database by using confidence intervals of >95% and the Mascot search engine (Matrix Science, Boston, MA), which typically provided matches to Acinetobacter sp. ADP1 (unless otherwise specified). Statistical analyses on the electrophoretic profiles were performed by comparing the changes in protein volume between the control and exposed samples for each strain with a paired Student t test, where the difference in protein volumes (or expression differences) was considered significant when p≤0.05, and inferred as a trend in the proteome of the strain when p≤0.1 (Ethen et al., 2006; Trost et al., 2009; Jovanovic et al., 2010). Control analyses were additionally performed on stationary phase cultures of both strains (Applied Biomics, Hayward, CA).
3. Results
3.1. Survival in aqueous hydrogen peroxide
This study updates and extends upon the preliminary findings of La Duc et al. (2003) regarding the extremotolerance toward H2O2 for A. radioresistens 50v1. For the sake of simplicity, the cell line of A. radioresistens ATCC 43998 is here on referred to as the type strain. In the survival studies, mid-log phase cultures of each strain were exposed to 100 and 320 mM H2O2 for 1 h followed by plating of the quenched cultures on agar plates. As displayed in Fig. 1 (inset graph), the 100 mM H2O2 exposure resulted in a ∼2-log reduction in surviving colonies for the 50v1 strain, whereas, in sharp contrast, the type strain showed no significant survival after exposure. At the higher concentration of 320 mM H2O2, the 50v1 strain survival was also reduced to insignificant levels by a >8-log decrease in survivability, which thereby demonstrates a very robust disinfection of ∼109 cfu/mL by ∼1% w/w H2O2 (320 mM H2O2). In terms of significance, these results suggest a potential for sterilization by using common disinfectant solutions such as 3% w/w H2O, which is contrary to the interpretation of prior studies (La Duc et al., 2003 and unpublished results) that indicated only ∼2-log and ∼6-log reductions for 50v1 and type strains at 1.5% w/w H2O2 (490 mM; 5% v/v of a 30% w/w solution).
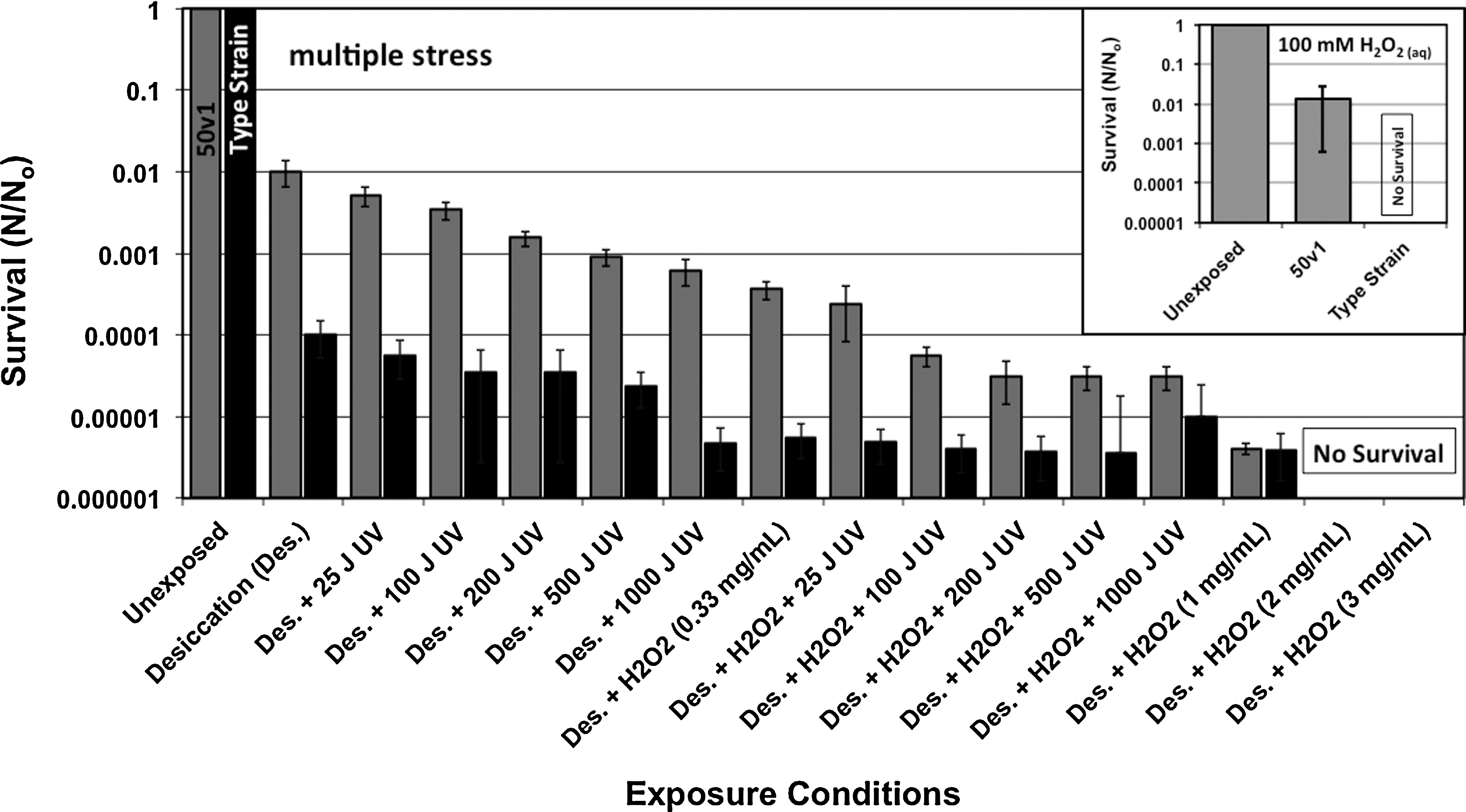
The survival of A. radioresistens 50v1 (gray) and the type strain (black) after exposure to 100 mM H2O2 for 1 h (inset graph), and to the multiple stress factors (primary graph) of desiccation (des, 10 days), vapor and plasma phases of H2O2 (0.33–3.0 mg/mL), and UV254 irradiation (25–1000 J m−2). For the 100 mM H2O2 exposures (N 0 109 cfu/mL), the error bars represent the standard deviation (n=3) of the averaged survival ratios (N/N 0), and the type strain displayed no significant growth after exposure (<25 colonies/plate). For the multiple stress exposures (N 0 108 cfu/mL), the ratio of N/N 0 was derived from the averaged survivals and initial titers for each strain, and the error bars represent the 95% confidence limits for each survival ratio, as calculated from the standard error of the most probable numbers method; for the exposures, the combination and experimental sequence are indicated on the x axis label, and an area of 1 m2 is assumed for simplicity's sake.
3.2. Survival under multiple stress conditions
The impacts of sequential exposures to desiccation, vaporous and plasma phase H2O2, and UV irradiation (UV254) were measured for each of the Acinetobacter strains. As also shown in Fig. 1, the 50v1 and type strains manifested ∼2- and ∼4-log respective reductions (from ∼108 cfu/mL) in survivability after the 10-day desiccations, which decreased to ∼3-log and ∼5.5-log reductions when combined with UV254 irradiation (25–1000 J m−2). For the 50v1 strain, the overall survival after UV254 exposure (from 200 to 1000 J m−2) was higher than previously reported (Newcombe et al., 2005), which thereby suggests that desiccation prior to exposure may increase the degree of survivability. Interestingly, the survivals were also very similar to those obtained from the desiccation and H2O2 exposures (0.33 mg/mL), which yielded total ∼3.5-log and ∼5.5-log reductions for 50v1 and type strain, respectively. The similarities, therefore, revealed that sterilization for which very low H2O2 concentrations were used (in the vapor and plasma phases) was much more effective than UV254 irradiation at high cumulative dosage (1000 J m−2).
When exposed to the sequential stress conditions of desiccation (10 days), vapor/plasma H2O2 (0.33 mg/mL), and UV254 (25–100 J m−2), the survivals of the 50v1 and type strains reduced by ∼4.5-log and ∼5.5-log, respectively. At the higher UV254 dosages (200–1000 J m−2), however, no further significant reduction in survival was observed for either strain; thus indicating that these conditions (UV254 irradiation in conjunction with desiccation and exposure to low concentrations of oxidant) were insufficient in achieving complete deactivation of the Acinetobacter strains. Complete deactivation was only achieved when using higher concentrations of H2O2 (2–3 mg/mL), thereby suggesting a predominant role of chemical degradation by the vapor/plasma phase H2O2 in the sterilization process.
3.3. Hydrogen peroxide degradation
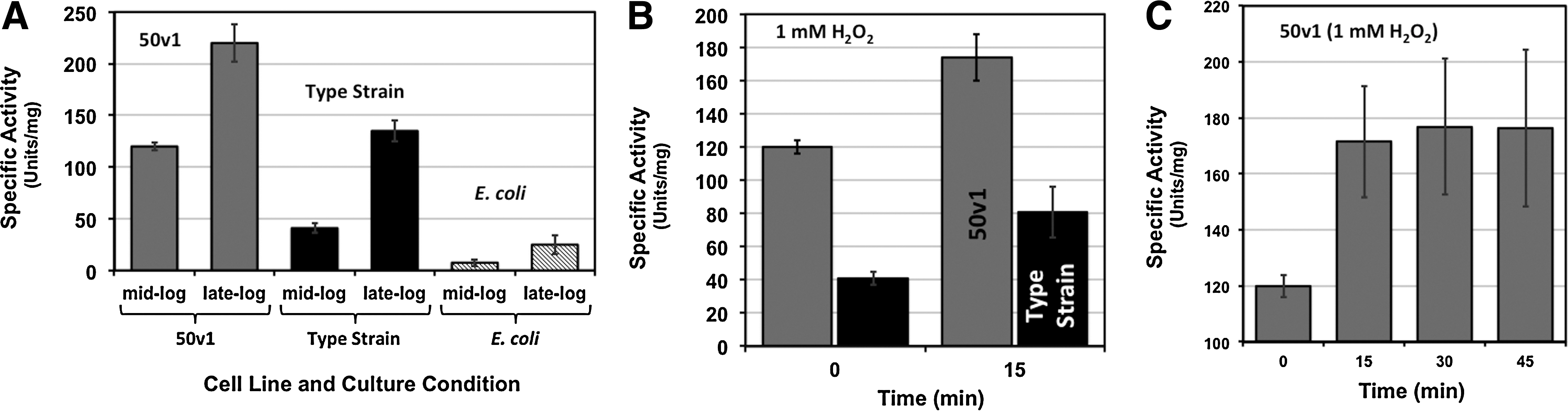
The H2O2-degrading properties of cellular extracts from A. radioresistens 50v1, the type strain, and E. coli were measured by using spectrophotometric assays and considered to be the dominant result of a catalase-like degradation due to the significant evolution of gaseous products in the kinetic and suspension-based survival experiments. Cellular extracts of each strain were prepared with ultrasonication and the specific activities of hydrogen peroxide degradation compared across the strains. As displayed in Fig. 2A, the trend in catalase specific activities was 50v1 (120±4 Units/mg)>type strain (41±4 Units/mg)>E. coli (7.6±3.3 Units/mg), which (for the Acinetobacter strains) matched the overall trend in survivability against H2O2 (50v1>type strain). These results, therefore, supported the role of catalase as a key factor in the survivability. The H2O2-degrading properties of both strains were also dependent upon culture conditions with late-log phase cultures consistently providing higher catalase specific activities than those obtained at mid-log phase.
(
For the Acinetobacter strains, the changes in catalase specific activities were also measured after the addition of exogenous H2O2. Each strain was exposed to 1 and 490 mM H2O2 and the catalase specific activities measured before and after the addition of the oxidant. As shown in Fig. 2B, both the 50v1 and type strains responded to the addition of H2O2 with ∼1.5- and ∼2-fold increases in catalase expression (or regulation), after a 15 min exposure, to yield the final catalase specific activities of 170±13 and 81±15 Units/mg, respectively. For the 50v1 strain, additional measurements taken at 30 and 45 min after the exposure (Fig. 2C) revealed no further increases in the catalase specific activity. Experiments addressing the intra- or extracellular location of the catalase after the exposure showed no significant activity in the media component (or supernatant) of cultures of either strain, thereby indicating that the H2O2 exposure did not result in cell rupture (or cell wall fragmentation) and concomitant release of catalase (or other H2O2-degrading enzymes) into the media. Lastly, no enzymatic activity was detected after exposure to 490 mM H2O2 (1.5% w/w), which was consistent with the lack of observed survival at 320 mM H2O2 and an absence of significant metabolic activity arising from nonviable cells.
3.4. Proteomic analysis of H2O2 exposure
The impact of H2O2 on the proteomes of the A. radioresistens 50v1 and type strains were measured and characterized using two-dimensional polyacrylamide gel electrophoresis and mass spectrometry. Exposures were performed with 1 mM H2O2, which was sublethal for both strains, and an exposure time of 15 min, since longer incubation times yielded no further increases in hydrogen peroxide degradation activity (Fig. 2C). Electrophoretic gels demonstrating the impacts of H2O2 are displayed in Fig. 3; these gels are representative examples of the proteomes and do not reflect the averaged impacts across the independently exposed samples. Statistical comparison of the protein volumes from all replicates, therefore, indicated that 10 proteins from the 50v1 strain changed in expression-fold by ≥1.2 (or 20%) as a result of the 1 mM H2O2 exposures (expression-fold is the ratio between the exposed and control samples). Mass spectrometry revealed that, among this group, only five proteins matched to entries within the NCBI database, which suggested the presence of unknown Acinetobacter proteins. In sharp contrast, the type strain manifested no significant changes or trends in protein expression upon H2O2 exposure.
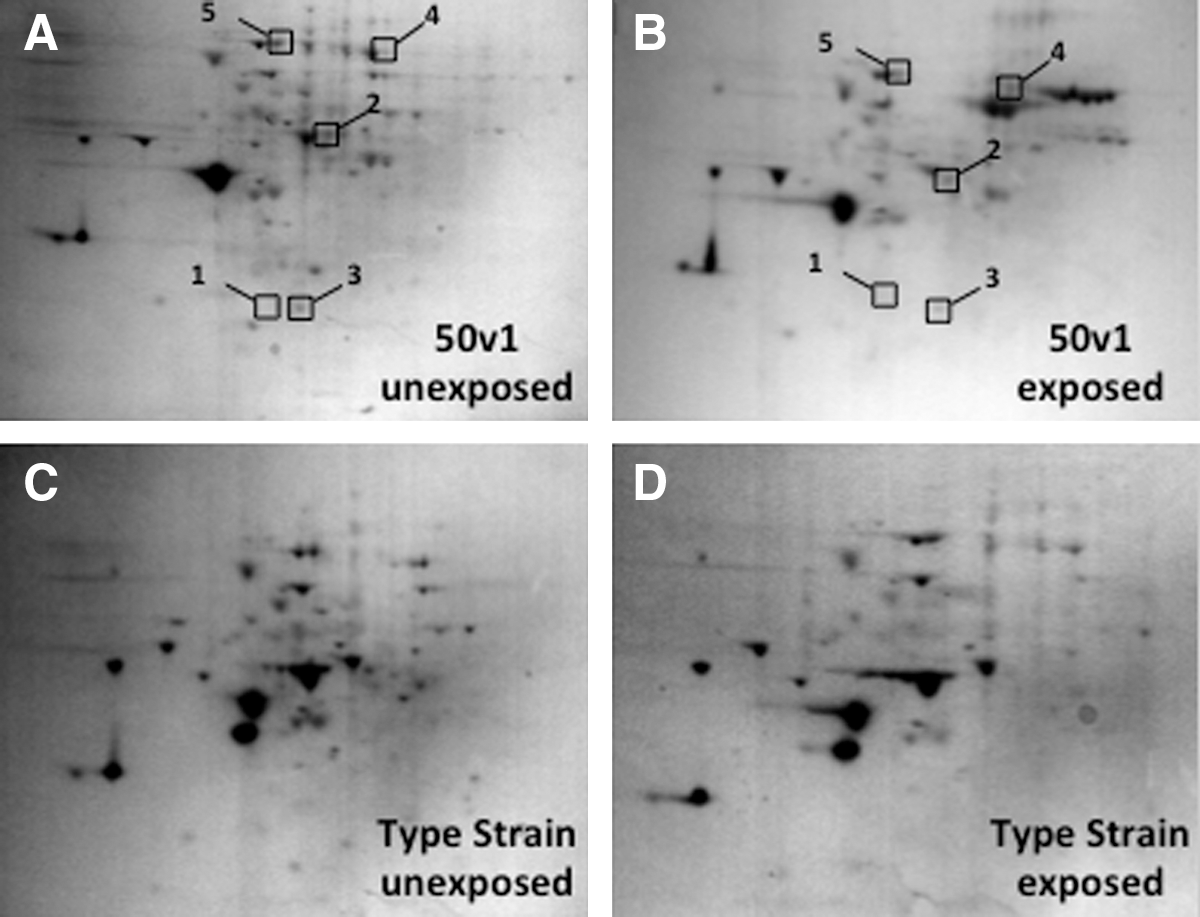
Representative two-dimensional electrophoretic gels demonstrating the impact of 1 mM H2O2 (15 min) on the proteomes of A. radioresistens 50v1 and the type strain, where (
As displayed in Fig. 4, the most noteworthy impact of the H2O2 exposure was the statistically significant downward expression of ATP synthase, which decreased in expression-fold by 40%. More specifically, mass spectral studies indicated a downward expression of the beta subunit of the F1 domain of ATP synthase (Q6FFK0), which supported a modulation of ATP synthesis by the 50v1 strain as a result of the oxidative stress. The proteomics also demonstrated an upward trend in alkyl hydroperoxide reductase (Q6FAK2), which increased in expression-fold by 50%, thus illuminating a potential secondary peroxide degradation pathway, as this enzyme degrades alkyl and hydroperoxides. Upward trends in the 50v1 strain were also observed for the expression of EF-G (Q6FDS6), which increased by 20% and is involved in ribosome translocation and protein folding, and for a hypothetical protein (Q7V264, Prochlorococcus marinus subsp. pastoris), which increased by 50% and possessed a 97% sequence identity (e=1×10−30) to an uncharacterized membrane protein from Synechococcus sp. WH7803 (CAK23819.1). Additionally, a 20% downward trend in the expression of peptidyl-prolyl isomerase (Q6FB14) was observed, which serves differing roles in protein folding. Initial studies were also performed on unexposed stationary phase cultures, which revealed that the 50v1 strain possessed higher abundances of alcohol dehydrogenase (NP_805994), OmpA-like protein precursor (AY033946), EF-Tu (YP_045082), and NADH-dependent enoyl-ACP reductase (YP_047630). In contrast, the type strain possessed higher abundances of the putative ring oxidizing protein (CAC10606), enoyl-CoA isomerase (CAA66096), succinylornithine transaminase (YP_045979), and dihydrodipicolinate reductase (YP_048077).
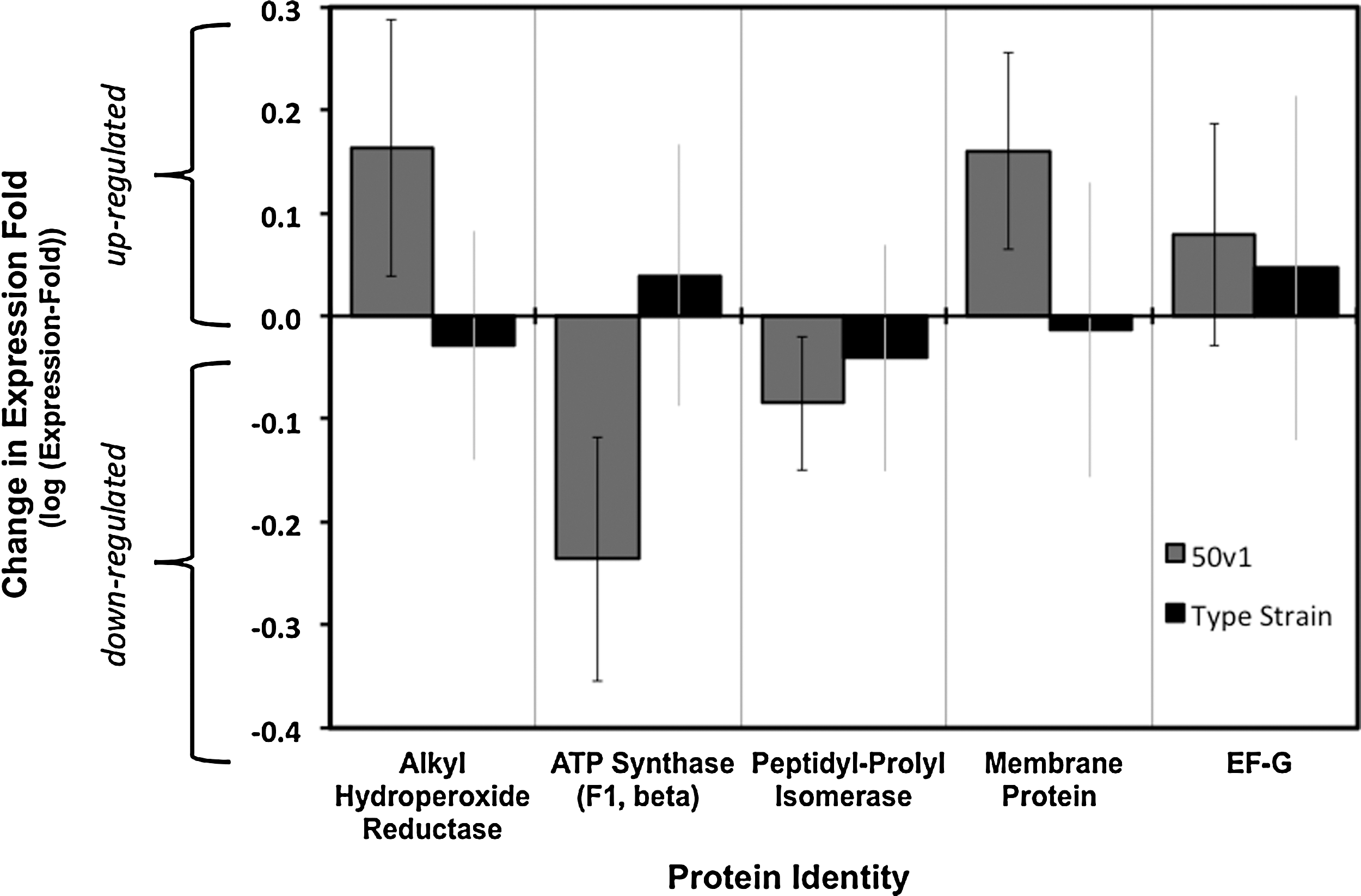
Change in protein expression for A. radioresistens 50v1 and the type strain after a 15 min exposure to 1 mM H2O2 (n=3, error bars display the standard deviation), where the upward and downward changes in protein expression are displayed as positive and negative values, respectively, due to the log transformation of the expression-fold value (y axis); for each of the proteins, the untransformed expression-fold values, standard deviations, and the statistical significances determined from Student t tests are as follows: alkyl hydroperoxide reductase (1.5±0.4, p=0.07, t=3.52); ATP synthase (0.6±0.2, p=0.009, t=10.3); peptidyl-prolyl isomerase (0.8±0.1, p=0.100, t=2.79); membrane protein (1.5±0.3, p=0.100, t=2.73); and EF-G (1.2±0.3, p=0.06, t=3.87).
4. Discussion
Insights into the biochemical mechanisms and range of extremotolerance for A. radioresistens 50v1 were gained through a combination of microbiological, proteomic, and enzymatic studies. As demonstrated in the survival experiments, the extremotolerance is broader than previously reported, with A. radioresistens 50v1 manifesting remarkable resistances toward high concentrations of oxidant (100 mM H2O2), irradiations with UV254 (1000 J m−2), and to the sequential exposures to desiccation, vapor and plasma phase H2O2, and UV254 irradiation. These results are significant, as the measured survivals under 100 mM H2O2 are among the highest reported for non-spore-forming and Gram-negative bacteria (Miller, 1969; Ichise et al., 1999; Buckova et al., 2010; Tondo et al., 2010). In comparison, and as expected, the survivals of certain spore-forming and Gram-positive bacteria are significantly higher, for instance the spacecraft-associated Bacillus odysseyi and strains of Bacillus pumilus display only ∼1–2-log reductions in 1.6 M H2O2 (La Duc et al., 2004b; Newcombe et al., 2005).
Enzymatic contributions to the extremotolerance of A. radioresistens 50v1 were supported by comparisons of the specific activities of hydrogen peroxide degradation, which revealed that the 50v1 strain possessed ∼3-fold and ∼16-fold respective excesses over the type strain and E. coli, which served as Gram-negative and non-extremotolerant controls. The specific activity of hydrogen peroxide degradation (or catalase specific activity) was also H2O2 concentration dependent, as the addition of 1 mM H2O2 induced substantial increases in catalase activity, which supported an OxyR-like response to H2O2 exposure (Zheng et al., 2001; Barbe et al., 2004). Nevertheless, the ∼3-fold difference in catalase specific activities between the 50v1 and type strains was perhaps insufficient in providing a rationale to the vastly different survivals in 100 mM H2O2 (∼2-log reduction for the 50v1 strain versus no survival for the type strain). In a similar comparison, both E. coli and Vibrio rumoiensis S-1, a Gram-negative bacterium isolated from fish egg bleaching plants (bleaching agent was H2O2), display limited and respective survivals of ∼7% and ∼3% in 0.4 mM H2O2, despite a catalase specific activity of 4000 Units/mg for V. rumoiensis S-1 (Ichise et al., 1999; Yumoto et al., 1999), thus further suggesting that catalase activity is necessary but not sufficient in conferring an extremotolerance toward H2O2. Indeed, the extreme survival of V. rumoiensis S-1 during the bleaching process is additionally related to use of cell surface and/or extracellular catalase as a means of degrading the bleaching agent. For the 50v1 strain, however, the role of extracellular catalase was eliminated as a protection strategy due to an observed lack of catalase specific activity in the cell free extracts of H2O2-exposed cultures.
Hence, the impacts of H2O2 on the proteomes of 50v1 and type strains were studied by using two-dimensional gel electrophoresis and mass spectrometry. Comparative analyses indicated that the extremotolerance of A. radioresistens 50v1 was additionally related to lowered energy requirements (down-regulation of ATP synthase), the use of alternative H2O2 degradation pathways (up-regulation of alkyl hydroperoxide reductase), differences in protein synthesis and folding (up-regulation of EF-G and down-regulation of prolyl isomerase), and modifications to the cell membrane (up-regulation of membrane protein of unknown function). Suggestive roles for the cell wall in the extremotolerance were additionally inferred through comparative analyses on stationary phase cultures that indicated a higher relative abundance of membrane surfactant proteins (up-regulation of Omp-A-like protein), increase in biosynthesis of membrane-specific fatty acids (up-regulation of NADH-dependent enoyl-ACP reductase), and decrease in catabolism of unsaturated fatty acids (down-regulation of enoyl-CoA isomerase) for the 50v1 strain. The proteomics, however, have yet to corroborate the observed increases in catalase activity (upon addition of 1 mM H2O2), which supported post-translational regulation of catalase and/or enzymatic contributions from (an OxyR-related) alkyl hydroperoxide reductase (Barbe et al., 2004).
Interestingly, the multiple stress resistances and proteomic profiles of A. radioresistens 50v1 were additionally suggestive of adaptations toward the selective pressures of the assembly facilities and cleaning regime for spacecraft surfaces (isopropanol and ethanol wipes). For instance, adaptations toward the low-humidity conditions of the spacecraft assembly facilities could very well confer the resistance toward oxidative and radiation damage, as desiccation resistance has been shown to relate to both oxidative and radiation tolerance (Fredrickson et al., 2008; Slade and Radman, 2011). Moreover, the observed up-regulated trend of alkyl hydroperoxide reductase, while consistent with oxidative stress, may also be the result of adaptations toward the presence of organic peroxides, which can be formed from isopropanol upon exposure to air or after long-term storage (Kelly, 1996; Clark, 2001). Furthermore, the observed higher abundance of alcohol dehydrogenase in the 50v1 strain is consistent with utilization of ethanol as an alternative energy or carbon source (Abbott et al., 1973).
When taken together, therefore, the combined analyses clearly suggest that the protection strategy for A. radioresistens 50v1, under sublethal oxidative stress, includes the enzyme-based degradation of H2O2 (catalase and alkyl hydroperoxide reductase), energy management (ATP synthase and alcohol dehydrogenase), and modulation of the membrane composition. The observed slight proteomic responses to 1 mM H2O2, nevertheless, indicated that the native metabolic states of A. radioresistens 50v1 were sufficient in handling the impacts of exogenous low-concentration oxidants. Conversely, the lack of measured response in the type strain proteome was perhaps indicative of an inability to appropriately adjust to oxidative stress, which is consistent with the lack of survival for this strain at the higher H2O2 concentrations. Moreover, the proteomic differences served to highlight the unique biochemistry of the A. radioresistens 50v1 strain.
As a result, we assume a probability (albeit low) for Mars contamination given the biochemical and survival features of A. radioresistens 50v1 and the potential for orbiter failure and/or final spacecraft disposition for the Mars Odyssey spacecraft. Consequently, any persistence on Mars would be necessarily dependent upon the final conditions of the spacecraft [i.e., heat generated, degree of breakup, shielding (or shadowing) arising from spacecraft materials, and subsurface burial] (Space Studies Board, 2000, 2006) and the functioning of metabolic pathways under anaerobic environments, such as the phosphate and carbon metabolic pathways in varying Acinetobacter (Rustrian et al., 1997; Boswell et al., 2001). In turn, any proliferation on Mars would be contingent upon (and thereby extremely limited to) the contamination of niche environments, such as sediment-water systems, capable of sustaining aerobic respiration and heterotrophic metabolism (Boström et al., 1988; Muyima and Cloete, 1995).
5. Conclusion
In conclusion, our analyses on A. radioresistens 50v1 provide key insights into the biochemical strategies that confer extremotolerance toward H2O2. Of particular importance is the role of both catalase and alkyl hydroperoxide reductase, thus indicating an interplay between these enzymes in the oxidative stress response for Acinetobacter and the intriguing role of ATP synthase, which has not been commonly reported as a factor in peroxide-related stress. Furthermore, our studies suggest a tolerance toward Mars-like degradative conditions due to the survival of A. radioresistens 50v1 against conditions which served as first-order approximations for the dry (desiccation), radiation (UV254 irradiation), and oxidizing (aqueous, vapor, and plasma phase H2O2) environments of Mars.
Together, therefore, these studies suggest that A. radioresistens 50v1 may persist on Mars and potentially interfere with future life- and biochemical-detection missions (as false positives), which thereby emphasizes the need for robust spacecraft cleaning protocols that chemically degrade and remove biological matter yet retain compatibility with spacecraft materials. This is especially significant when considering that multiple Acinetobacter were discovered in the assembly facility for the Mars Phoenix lander (Ghosh et al., 2010) and that the abundance of these Acinetobacter increased as a result of the spacecraft assembly process (Vaishampayan et al., 2010). Thus, our analyses on A. radioresistens 50v1 support the possible adaptation or resistance toward the clean room and cleaning regime conditions for spacecraft, which, while still speculative, could have long-term impacts on the microbial ecology of the assembly facilities.
Footnotes
Acknowledgments
These studies were supported by the NASA Astrobiology Institute Minority Institutional Research Support program (R. Mogul) and in part by a NASA ROSES 2003 award (K. Venkateswaran). The authors extend their gratitude to Freida Dallal and Charlie Seto for biochemical support, Claire Waggoner (IIRMES) for the proteomics work and discussions, and to the Cal Poly Pomona new investigator funds and Science Educational Enhancement Services program (SEES) for initial support.
Abbreviations
cfu, colony-forming units; LB, Luria-Bertani; PBS, phosphate-buffered saline; TSB, tryptic soy broth; TYG-B, tryptone yeast extract glucose broth.