
Abstract
Early in its history, Earth's surface developed from an uninhabitable magma ocean to a place where life could emerge. The first organisms, lacking ion transporters, fixed the composition of their cradle environment in their intracellular fluid. Later, though life adapted and spread, it preserved some qualities of its initial environment within. Modern prokaryotes could thus provide insights into the conditions of early Earth and the requirements for the emergence of life. In this work, we constrain Earth's life-forming environment through detailed analysis of prokaryotic intracellular fluid. Rigorous assessment of the constraints placed on the early Earth environment by intracellular liquid will provide insight into the conditions of abiogenesis, with implications not only for our understanding of early Earth but also the formation of life elsewhere in the Universe. Key Words: Astrobiology—Early Earth—Hadean—Origin of life. Astrobiology 13, 294–302.
1. Introduction
Our investigation is based on two assumptions: (i) all contemporary creatures on Earth originated from a Last Universal Common Ancestor (LUCA); (ii) LUCA and its descendants incorporated and preserved in their structures the composition of the prebiotic environment (e.g., Chopra et al., 2010). We admit the possibility that early metabolic processes strongly influenced the composition of the first organisms (e.g., Holm and Baltscheffsky, 2011). If this is the case, our analysis of the elemental composition of present-day cytoplasm does not give any information about the environmental conditions of the origin of life and early evolution.
The aim of this study was to conduct a survey of the chemical composition of living creatures from today's biosphere in an attempt to extract the key properties of the potential environment where life originated. In the present work, we focused on prokaryotes due to their close proximity to LUCA. We compared these with a suite of geochemical fluids, including the most abundant modern terrestrial solutions, and those produced by dint of geochemical modeling of Hadean Earth. This allowed us to speculate about where and in what conditions life originated.
2. The Early Biosphere
Estimates based on the best current paleontological and carbon isotopic data indicate that the first cells appeared as early as 3.0–3.3 Ga (fossils, Brasier et al., 2006) or even 3.85 Ga (isotopes, Mojzsis et al., 1996). Some have suggested that, as early as this time, Archaea and Bacteria had already diverged and begun to diversify (Ueno et al., 2006; Philippot et al., 2007; Boussau and Gouy, 2012). A phylogenetic tree based on studies of molecular biology (Hedges et al., 2001), recent research on Early Archean serpentine mud volcanoes (Pons et al., 2011), biomarker evidence of life (Mojzsis et al., 1996), and models and hypotheses on free energy availability in Hadean hydrothermal systems (Martin et al., 2008; Simoncini et al., 2011) reinforce the possibility that hyperthermophilic microorganisms may have emerged as early as 4 billion years ago.
Genomic analysis provides support for the notion that the three main domains of life—Bacteria, Archaea, and Eukarya—evolved from LUCA (Pace, 1997). LUCA was probably not a single cell but a population or community of replicating microorganisms with uncomplicated morphology, modular structures, simple functions, and gene exchange (Zuckerkandl and Pauling, 1965; Pace, 1997; Nisbet and Sleep, 2001). It likely existed in high-temperature conditions as a chemoautotroph between 3.9 and 3.5 Ga when deep-sea benthic environments or subsurface sites were more stable to the establishment and development of life and the evolution of early primitive ecosystems.
There are at least two interpretations for the hyperthermophilic nature of early life. In the first, the early Archean environment served as a hyperthermophilic Eden that was conducive to the emergence of life from nonlife. In this scheme, life emerged in the high-temperature environment provided by impact events or hydrothermal systems (Oberbeck and Mancinelli, 1994; Nisbet and Sleep, 2001; Kelley et al., 2002). A second option is the Noah hypothesis, which holds that LUCA was the product of an early catastrophe that wiped out all but the most thermophilic organisms. This would require that life had an early origin in an unknown environment, with all its descendants except the thermophiles wiped out. The cataclysm could take the form of impact events, which may have heated the oceans to 100°C (Nisbet and Sleep, 2001). Both the Eden and Noah hypotheses assume that the hyperthermophilic species of the Archean eon had genetic and phenotypic traits very close to those of the ancestor of all extant organisms.
3. Biogeochemical Commonalities
No nucleic acid evolution is possible without a supporting geochemistry, later biogeochemistry, and finally biochemistry to provide a steady flow of adequate concentrations of polymerizable precursors (Martin and Russell, 2002). While the essential elements that form the macromolecules of living cells are C, H, N, O, P, and S, many other elements are needed in variable amounts to stabilize these molecules and to maintain a habitable environment.
Regarding the origin of the cell, it is hypothesized that the organization and composition of the cytoplasm progressed initially from an unstructured, microscopic hydrogel (precytoplasm) to a more complex cytoplasm. The precytoplasm could trap and retain (via diffusion) concentrations of ions from the surrounding environment (Trevors, 2011). These concentrations provided the background against which life was formed. As life evolved, this basic set of elements may have been modified and supplemented. Some authors speculate that changes in intracellular concentration track changes in the availability of elements at Earth's surface (e.g., Chopra et al., 2010). However, certain inorganic elements and ions (such as hydrogen, sodium, potassium, and magnesium) found in modern life forms are relevant for the homeostasis of life and are expected to be conserved with minimal variation among prokaryotes.
We have compiled from numerous sources the available data on prokaryotic composition (Fig. 1). The proportion of certain elements within the cell, such as C, N, and O, is conserved across microorganisms, whereas other elements fluctuate moderately (Na, K, Cl, Mg) or to a greater extent (Zn).
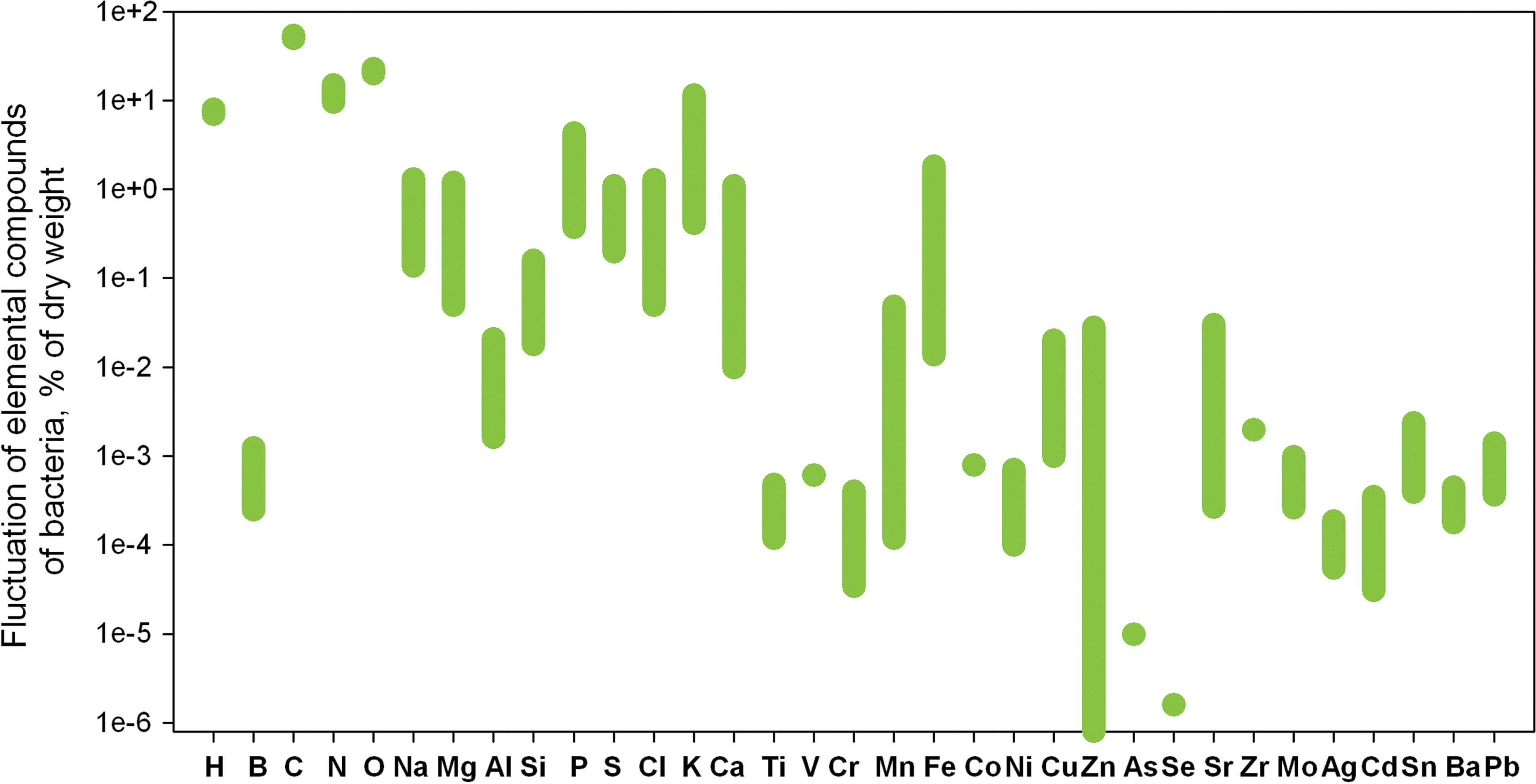
The elemental composition of modern bacteria1 (33 elements).
4. Cytoplasmic Concentrations Compared with Geochemical Fluid
Searching for life's cradle environment can be reduced to the search for the natural solution or solutions that maximally correspond with relative and absolute concentrations of chemical elements in living creatures. Prebiotic evolution probably requires long-term invariability of environmental conditions. Another requirement for the emergence of life is that it occur on the boundary of environments with different redox potentials (e.g., Galimov, 2006). Solutions that satisfy these conditions include seawater, fluids of oceanic or terrestrial hydrothermal systems, or solutions formed under subaerial weathering on the surface of the primordial continents.
The elemental composition of modern seawater is different from that of prokaryotes (Fig. 2). Performing the Kolmogorov-Smirnov 2 samples test (KS test)1 with the significance level equal to 0.05, we are able to accept the null hypothesis (that the two elemental concentrations are drawn from the same distribution) only for Mg (p=0.06), Ca (0.06), Sr (0.082), and Mo (0.1). In addition, the bulk salinity of modern seawater (3.5%) and prokaryotes (30%) differs significantly. The extreme nature of these differences suggests that seawater (at least that of modern oceans) is likely not the primary environment for prebiotic evolution.
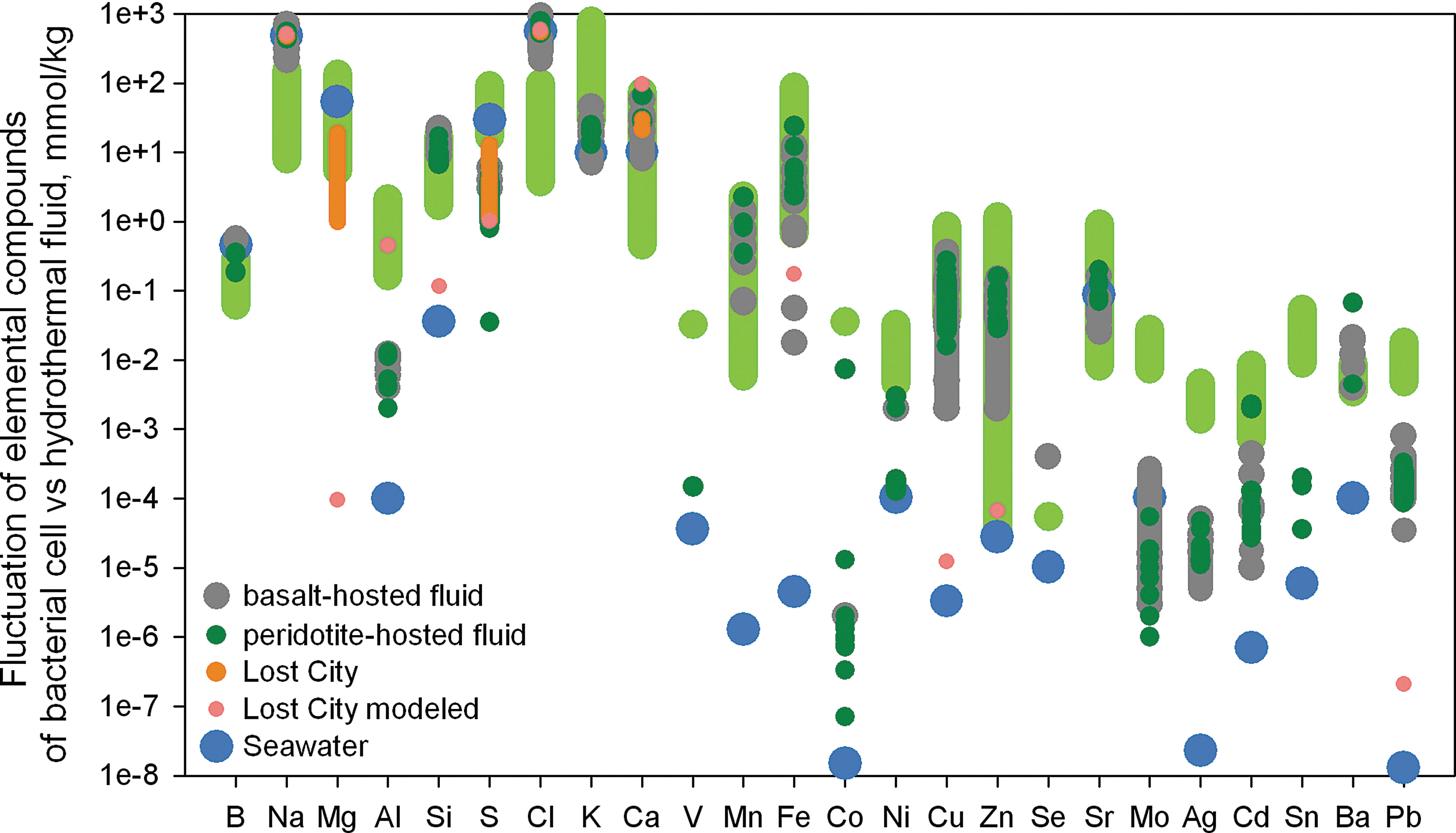
Modern seawater1 and oceanic hydrothermal fluids2 vs. bacterial elementary composition3 (marked by light green, as in Fig. 1).
The fluids of submarine hydrothermal systems are very promising environments for the origin and evolution of the first cells (e.g., Russell et al., 1998). The prokaryotes that inhabit these environments are metabolically simple and likely closer in evolutionary space to LUCA. The fluids are rich in molecular hydrogen, methane, sulfides, and ammonia, which are the raw materials for chemosynthesis. Compounds produced in hydrothermal systems fluctuate across very wide ranges in concentration and composition. In particular, their salt content is highly variable as a result of phase separation between vaporized solution and brines. Furthermore, not all hydrothermal fields have been explored or even discovered, and the fluids studied so far have always been contaminated with water from the open ocean.
We applied the KS test for the comparison of cellular composition and hydrothermal fluid from peridotite and basalt-hosted vents (Fig. 2). In the first case, similarities have been shown only for Ca (p=0.119), Mn (0.118), Fe (0.771), Cu (0.209), Zn (0.078), and Ba (0.586), and in the second only for B (p=0.052), Cu (0.059), and Zn (0.062). A number of publications dedicated to the unique hydrothermal field Lost City propose it is an analogue for the environment of early Earth where life originated (e.g., Kelley et al., 2002), mainly because hydrocarbons are very abundant in the hydrothermal fluids of this field (Konn et al., 2009). Our analysis as reflected in Fig. 2 (excepting Al and Ca) does not support the conclusion that this is the origin environment of life. This may be due to the paucity of direct observations of fluids of this hydrothermal field, despite the many references to this site in the scientific literature. We did not implement a statistical analysis of the compositions of Lost City fluid and living matter because the available data for Lost City fluid is not robust enough for the KS test.
We conclude that the elemental payload of prokaryotic cells is in good accordance with the modern submarine hydrothermal fluid composition in the case of heavy metals (Mn, Fe, Cu, Zn, Ba). This is promising for the conclusion that hydrothermal fields are the cradle of life, especially given the time stability of these environments. However, the absolute concentrations and especially ratios of elements such as Na, Mg, Al, Cl, and K in living matter do not fit their abundance in hydrothermal fluid (Fig. 3). Analyzing the dataset of Fig. 3, we determined that the KS test for elemental ratios of living cells and hydrothermal fluids from basalt-hosted vents shows a link between the two for Si/Na (p=0.4), Mn/Na (0.15), and Fe/Na (0.059). In the case of the peridotite-hosted vents, we found a link for the Mn/Na ratio (p=0.322) only. If today's prokaryotes have the elemental composition of the environment where they originated, they did not emerge from oceanic hydrothermal systems.
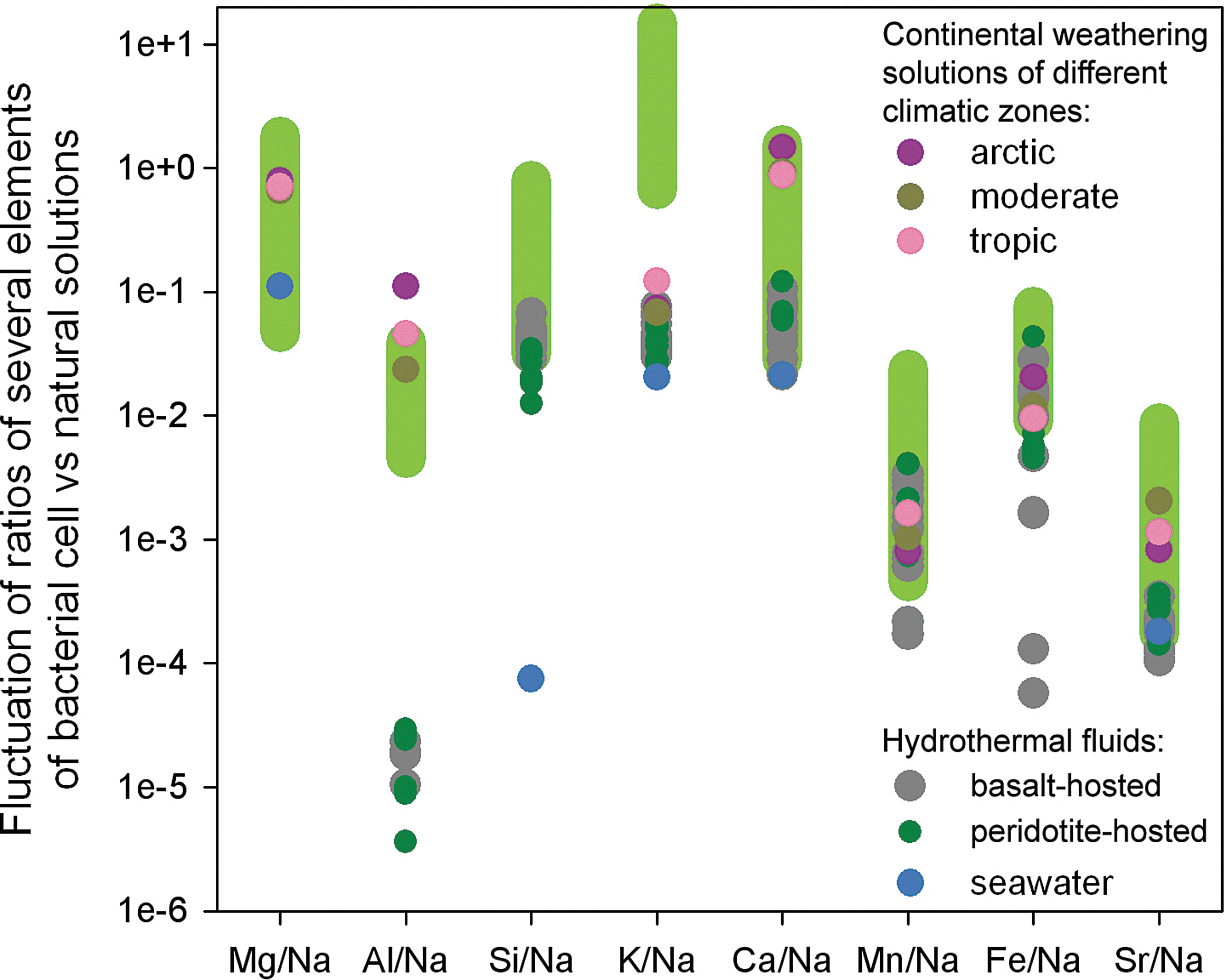
Ratio of several elements to sodium from modern geochemical solutions (Shvarcev, 1998)1 and bacterial cells (marked by light green).
Continental solutions (lakes, rivers, groundwaters, soil solutions) are highly variable environments. All liquid compounds on the seabed and in the oceanic crust share their composition with seawater. On land, however, the ratios and concentrations of cations in solution cover a wide range. In the water-weathering rock system, due to a low water-rock ratio, absolute cation concentrations are highly variable and depend directly on the chemical composition of the parent rocks. For comparison of weathering solutions with prokaryotic composition, it is necessary to consider ratios of elements to a standard. We selected sodium (Fig. 3). A statistical analysis of ratios of the most abundant cations in prokaryotes and the ones in weathering solutions showed good agreement between the two. Even the ratios of the commonly named “hydrothermal elements” Mn and Fe to Na equal the ratios in prokaryotes: Mg/Na (p=0.646), Al/Na (0.518), Ca/Na (0.071), Mn/Na (0.29), Fe/Na (0.234), and Sr/Na (0.1). The only case in which the null hypothesis was rejected (indicating that the distribution of elemental concentration ratios is not the same between the two fluids) was K/Na with p value 0.025, which is very close to the critical value (0.05). The importance of the Na/K ratio in the origin-of-life environment was discussed by Natochin et al. (2008). Despite the discrepancy in Na/K, its proportion in continental weathering solutions is the closest to living matter, when all natural solutions are compared.
5. Geochemical Modeling
Despite the remarkable correspondence of these modern subaerial solutions to the prokaryotic values, we must recognize the impact of early Earth's conditions on the solution produced.
We simulated the formation of seawater composition as a result of gradual cooling (from 600°C to 15°C) under a dense atmosphere that consisted mostly of water vapor and CO2 or CH4 and its interaction with a surface of solidified magma ocean. Independently, we calculated the composition of weathering solutions formed under subaerial conditions (1 bar CO2 or CH4 atmosphere, surface temperature of 15°C) and also simulated subaerial weathering under an atmosphere with modern pressure of carbon dioxide (PCO2=4×10−4 bar) as a control calculation. The simulations were implemented with the use of the GEOCHEQ code for thermodynamic calculations (Mironenko et al., 2008). The approach used for the kinetics of mineral dissolution was described by Zolotov and Mironenko (2007). In the course of thermodynamic calculations, we studied interactions in the O-H-K-Mg-Ca-Al-C-S-Si-N-Na-Cl-Fe system, and in the case of subaerial weathering simulations, the O-H-K-Mg-Ca-Al-C-Si-Na-Fe system. The initial substrate composition was basaltic komatiite from the Archean greenstone belt at Munro Township (Canada). More details on the methods and results of this investigation are presented in Novoselov and Silantyev (2012).
In the case of a CO2 atmosphere, the resulting seawater composition is very similar to that of modern seawater (Fig. 4a); the dominant cation is Na, and the only significant difference is a low content of Mg. The concentration of this cation is maintained by seabed weathering and continental flow. Despite the high content of Mg in basaltic komatiite, the hydrosphere loses it as a result of serpentinization during hydrothermal circulation. This predicts that the primordial ocean should be poor in Mg before the extended continents formed. The simulated seawater also has elevated Si and Fe content.
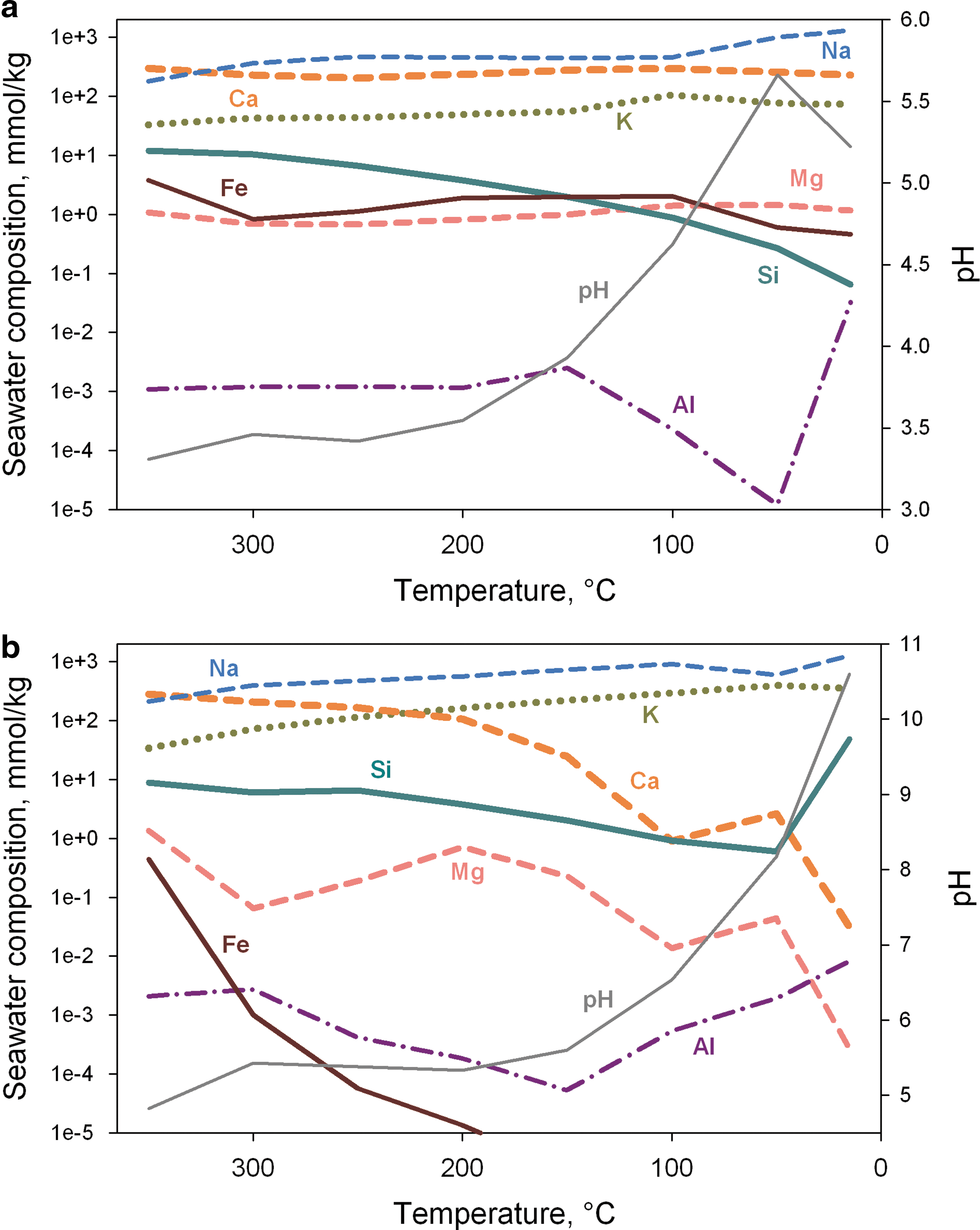
Change in the cation composition of seawater under atmospheres with CO2 (
For a CH4 atmosphere, the composition of the resulting solution was different (Fig. 4b). The main components were Na, K, and Si. The resulting seawater was strongly depleted in Mg, Ca, and Fe. In both modeled cases, the prevalent anion was chloride.
The results of the subaerial weathering simulations show remarkably different geochemical trends. The bulk composition of rocks weathered under CO2 atmosphere is characterized by accumulation of Si and Al; under the CH4 atmospheric conditions, Si, Al, Fe, and Mg. In both cases, the Na and K primarily accumulate in clays, which are later dissolved. K-containing illite is more resistant than Na-containing montmorillonite, which causes the final weathered solution to be more enriched in potassium than sodium. Thus, the resulting weathering solutions are characterized by high Mg/Na and K/Na ratios (Fig. 5). This phenomenon was first explored, with examples of Archean weathering profiles, by Alfimova and Matrenichev (2006).
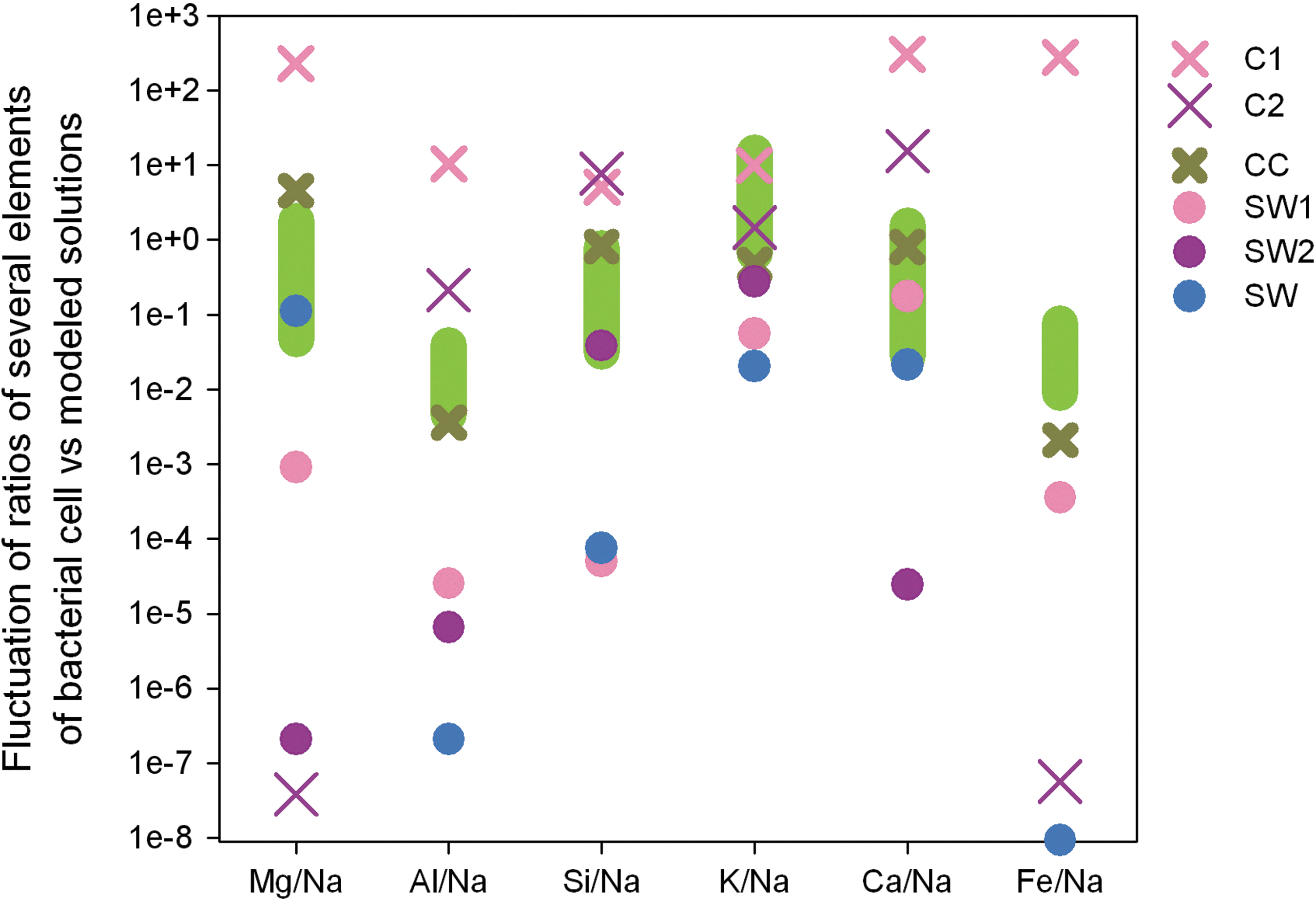
Comparison of calculation results with elemental ratios of bacterial cells (marked by light green). C1, the solution produced as a result of subaerial weathering under carbon dioxide atmosphere (PCO2=1 bar) after 2000 model years1; C2, results of subaerial weathering under methane atmosphere (PCH4=1 bar) after 20 model years1; CC, results of subaerial weathering under carbon dioxide atmosphere (PCO2=4E-4 bar) after 325 model years1; SW1, seawater compound produced in the system with CO2 atmosphere; SW2, seawater produced in the system with CH4 atmosphere; SW, modern seawater. All calculated data are for 15°C.
6. Discussion
The modeling results show that Mg/Na and K/Na of modern prokaryotes are similar to those of weathering solutions formed under the atmosphere with low CO2 content and clement environmental conditions (Fig. 5). Thus, the Na, Mg, Al, Si, K, Ca, Fe proportion in bacteria may reflect the ratio of these elements in solutions in contact with clay-containing sedimentary rocks formed as a result of basalt weathering of primordial Hadean protocontinents.
Clays are traditionally discussed as one of the most favorable environments for prebiotic evolution (e.g., Ponnamperuma et al., 1982; Wächtershäuser, 1988; Sami and Tewari, 2009; Ito et al., 2009). Clays have a relatively high ratio of surface area to volume, which favors adsorption, catalytic properties, and a layer structure that could potentially provide a suitable microenvironment that simulates the cell wall and acts as a matrix for a primitive genetic system.
One intriguing possibility is that primitive cells started to use the chemical elements gradually, depending on their abundance in the environment (e.g., Chopra et al., 2010). If this were true, the particularities of the chemical composition of prokaryotes would have recorded the sequence of events on early Earth (Fig. 6). The formation of the first cell-like structures probably took place in contact with sedimentary rocks that were possibly formed as a result of subaerial weathering on the surface of the earliest continents more than 4 billion years ago. Later, these primitive structures may have spread and reached into other environments where they could further change in response to the different available chemical conditions. Submarine hydrothermal systems may have been one of the most favorable niches. There, conditions would have enriched the proto-cytoplasm composition with heavy elements, due to their high natural abundance, but simultaneously conserved the early imprints of the water/sedimentary rock system. Strong selection may have occurred and produced a bottleneck effect, possibly during the Late Heavy Bombardment (3.9 Ga). The only surviving group of living creatures may have dwelt deep in Earth's crust (Oberbeck and Fogelman, 1990; Woese et al., 1990; Baross, 1998; Abramov and Mojzsis, 2009) and was probably made up of anaerobic hyperthermophiles. As soon as environmental conditions became suitable for life in the oceans and at the surface again, other niches would have been occupied. As most of these environments on early Earth probably resembled the hydrothermal ones, it is not difficult to conceive that hyperthermophiles could easily have adapted to them and later diversified to all modern living groups. Throughout this process, chemical evidence of the cradle environment of life was preserved. This evidence is visible even today and may help to constrain the environment for the origin of life. Here, we have attempted to constrain the scenario of the origin of life based on the elemental composition of life. The evidence, amazingly, points to an origin in a small warm Darwinian pond. But much work remains to be done.

Early events on Earth and study concept. The sketch illustrates the main events during early history of Earth, together with a schematic illustration of the dispersion of life into new environments. Light green lines trace the evolution pathways of primitive life as it occupied different environmental niches. LHB, Late Heavy Bombardment, LUCA, Last Universal Common Ancestor. (Color graphics available online at
Footnotes
Acknowledgments
The authors wish to acknowledge FAPESP (São Paulo Research Foundation) and Universidade de São Paulo for financial support to attend the São Paulo Advanced School of Astrobiology—SPASA 2011. Moreover, we would like to commend the excellent work of the SPASA 2011 Local Organizing Committee and especially that of Dr. Douglas Galante for his help and guidance during the preparation of this manuscript.
We also wish to thank Dr. M.V. Mironenko (Vernadsky Institute, Russia) for providing the modeling code and Aditya Chopra (Australian National University) for sharing prokaryotic composition information and for much helpful advice.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
KS test, Kolmogorov-Smirnov 2 samples test; LUCA, Last Universal Common Ancestor.
1
We did not analyze the V, Co, Se, Cd, because the number of available estimations of these elements in living cells is less than 3—the minimal number of estimations for the KS test application.