
Abstract
The hyperarid core of the Atacama Desert is one of the driest and most inhospitable places on Earth, where life is most commonly found in the interior of rocks (i.e., endolithic habitats). Due to the extreme dryness, microbial activity in these habitats is expected to be low; however, the rate of carbon cycling within these microbial communities remains unknown. We address this issue by characterizing the isotopic composition (13C and 14C) of phospholipid fatty acids (PLFA) and glycolipid fatty acids (GLFA) in colonized rocks from four different sites inside the hyperarid core. δ13C results suggest that autotrophy and/or quantitative conversion of organic matter to CO2 are the dominant processes occurring with the rock. Most Δ14C signatures of PLFA and GLFA were consistent with modern atmospheric CO2, indicating that endoliths are using atmospheric carbon as a primary carbon source and are also cycling carbon quickly. However, at one site the PLFA contained 14C from atmospheric nuclear weapons testing that occurred during the 1950s and 1960s, indicating a decadal rate of carbon cycling. At the driest site (Yungay), based on the relative abundance and 14C content of GLFA and PLFA, there was evidence of possible preservation. Hence, in low-moisture conditions, glycolipids may persist while phospholipids are preferentially hydrolyzed. Key Words: Endoliths—Extremophile—Carbon isotopes—Radiocarbon—Lipids. Astrobiology 13, 607–616.
1. Introduction
T

Map of the Atacama Desert region showing the sampling sites at Monturaqui (gypsum and ignimbrite rock) and halite (Yungay and Salar Grande). The dashed lines indicate the locations of evaporitic deposits.
Given the extreme dryness, it is expected that these microbial communities cycle carbon very slowly, similar to endolithic communities found in other hyperarid deserts such as the Antarctic Dry Valleys, where sandstone endoliths are thought to cycle carbon on timescales of several thousand years (Bonani et al., 1988; Sun and Friedmann, 1999). However, the viability and carbon cycling rates of endolithic communities in the hyperarid core remain unknown. Phospholipid fatty acids (PLFA) are an abundant component of the lipid membrane of bacteria and eukarya and are useful for characterizing the rate of carbon cycling within microbial communities. Since PLFA are hydrolyzed within days to weeks of cell death (White et al., 1979; Harvey et al., 1986), PLFA can be used as markers of the viable microbial community, and these compounds are not found in ancient sedimentary rocks. Glycolipid fatty acids (GLFA) are less well studied, but they have been linked to cyanobacteria (Sinninghe Damsté et al., 2001), which are common endolithic microorganisms (Walker and Pace, 2007), including in the Atacama Desert (Wierzchos et al., 2006, 2011, 2013; Dong et al., 2007).
The carbon isotopic composition of PLFA and GLFA can provide information about carbon cycling pathways in viable microbial communities (Fig. 2). The carbon isotopic composition of naturally produced organic matter depends on (1) the isotopic signature of the carbon source, (2) isotope effects associated with the assimilation of carbon, (3) isotope effects associated with the metabolism and biosynthesis of carbon, and (4) cellular carbon budgets (Hayes, 1993). While the 13C content of organic matter is affected by all four factors, mass-dependent fractionation effects are removed in reporting of radiocarbon (14C) data. The isotopic composition of the bulk organic matter does not necessarily reflect the isotopic composition of individual compounds. Due to the factors listed above, the 13C content varies between carbohydrates, proteins, and lipids. For example, PLFA are depleted in 13C relative to other products of biosynthesis, and the magnitude of this depletion depends on whether autotrophic or heterotrophic processes form the lipids (Hayes, 2001; van der Meer et al., 2003).
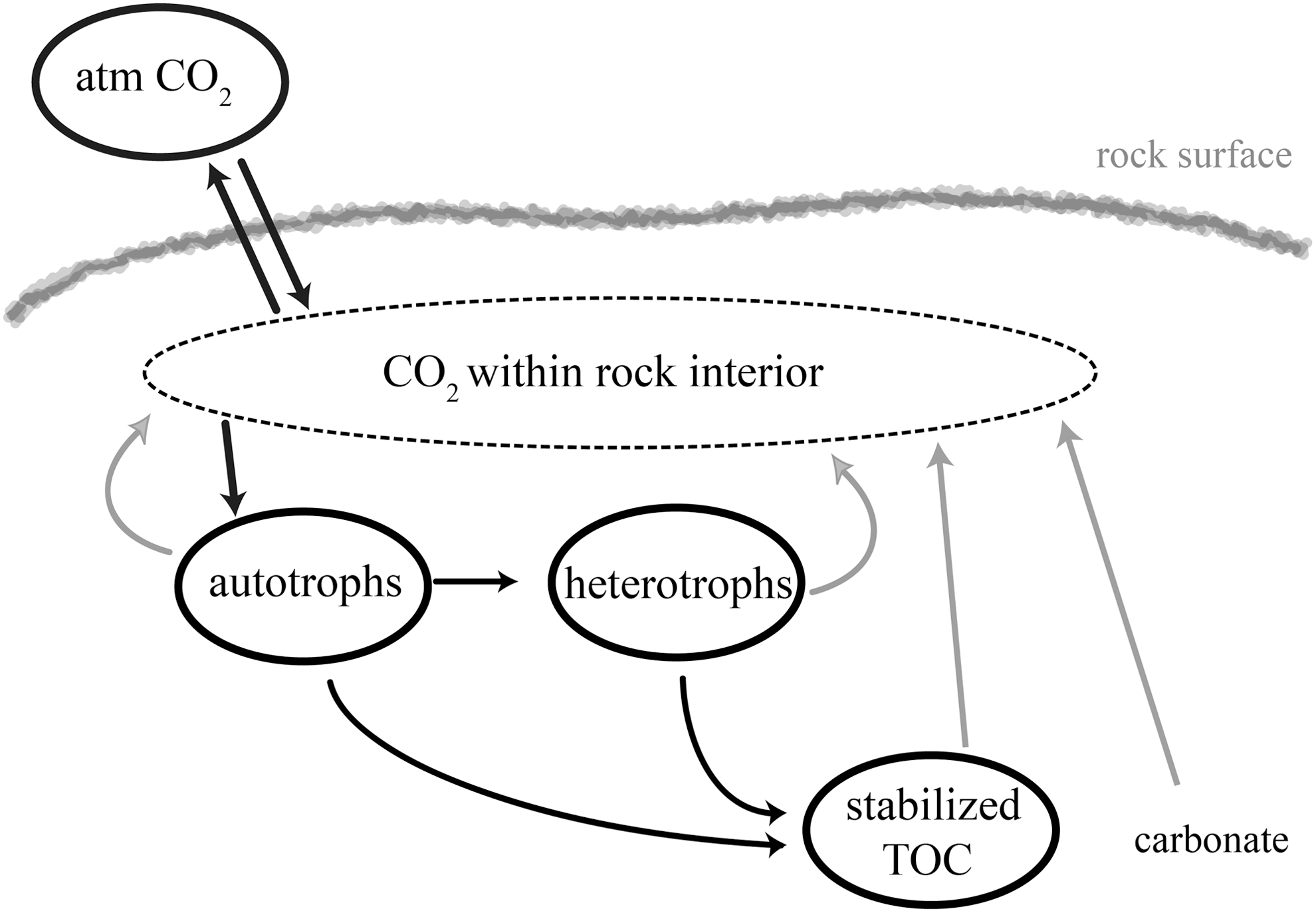
Pathways of carbon input, transformation, and loss from microbes in cryptoendolithic communities. Black arrows indicate pathways of carbon incorporation, while gray arrows indicate carbon recycling pathways. Adapted from Trumbore (2009).
Additionally, natural abundance radiocarbon profiling of PLFA and GLFA can help elucidate sources and cycling rates of carbon in natural systems (Slater et al., 2005). 14C is a naturally occurring isotope of carbon formed in the upper atmosphere by cosmic rays that decays to 15N with a half-life of 5730 years. However, thermonuclear weapons testing during the 1960s nearly doubled the 14C/12C ratio in Northern Hemisphere atmospheric CO2 (Fig. 3) (Nydal and Lövseth, 1983). Since thermonuclear weapons testing was banned in 1963, the bomb-produced 14C serves as a global isotope tracer for carbon exchange. There has been a well-documented steady decrease in the 14C/12C ratio of atmospheric CO2 due to uptake of bomb carbon into the ocean and terrestrial carbon pools and the simultaneous release of pre-bomb carbon from fossil fuel combustion, biosphere respiration, and ocean outgassing (Levin and Hesshaimer, 2000; Randerson et al., 2002; Naegler and Levin, 2006). On the timescale of half a century, the radioactive decay of 14C is comparatively small, and the decrease in atmospheric 14CO2 from year to year is greater than the precision of 14C measurements. Therefore, the bomb carbon record allows us to infer timescales for carbon exchange between the atmosphere and a given reservoir on annual to decadal timescales.
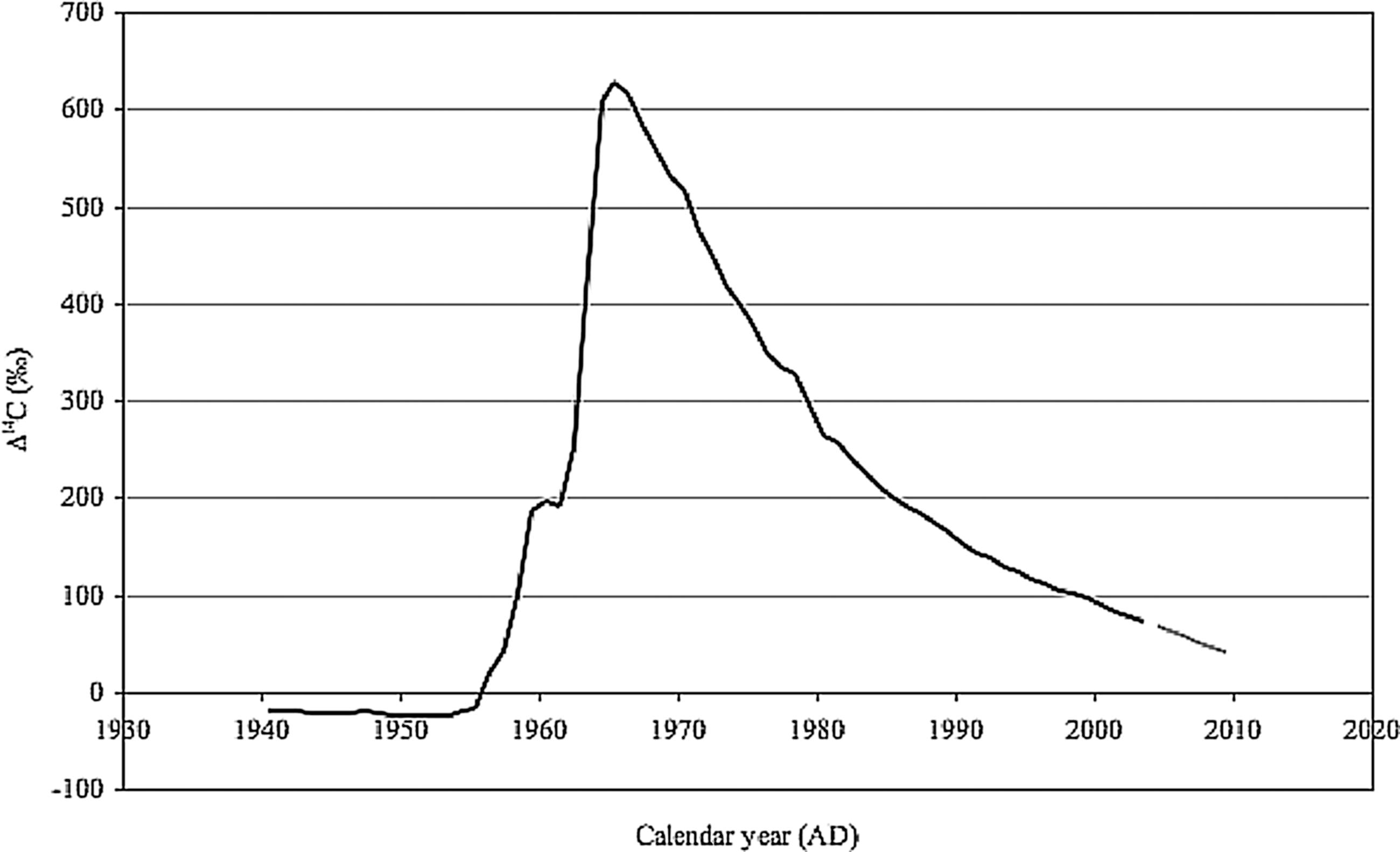
Based on the above, the 14C content of membrane lipids has been used to identify both modern (Brady et al., 2009) and old (i.e., pre-bomb) carbon sources in natural microbial communities (Cowie et al., 2010). In endolithic systems, it has been shown that microbes use atmospheric CO2 as a carbon source (Vestal, 1988), and these microbes are cycling carbon slowly (Bonani et al., 1988). Consequently, measuring the Δ14C, the ratio of 14C/12C measured for a sample in a given year relative to 1950 may provide insight into the rate of carbon cycling (e.g., activity) of an endolithic community. The incorporation of atmospheric CO2 would be expected to have a similar Δ14C as the modern atmosphere (e.g., Δ14C=+40‰). Observation of higher Δ14C values in viable communities (i.e., in PLFA) would be consistent with a bomb-carbon signal and, therefore, carbon cycling in decadal timescales. Alternatively, if the Δ14C of PLFA are lower than present-day atmospheric CO2, we would infer that microbes cycle carbon slowly enough that 14C decay is recorded, and/or the microbial community is recycling very old carbon (pre-bomb).
The objective of this study was to use natural abundance isotope (13C and 14C) analysis of membrane lipids to assess the carbon cycling rates of different endolithic communities within the hyperarid core of the Atacama Desert and to determine the potential for preservation of biosignatures within these communities. Our results show that these endolithic communities cycle carbon quickly, on the order of years to decades. At Yungay, the driest location studied, the GLFA were more abundant and older than the PLFA, indicating a better preservation potential of these compounds.
2. Materials and Methods
2.1. Sampling of endolithic microbial communities
Rock samples (1 kg) containing endolithic microbial communities were obtained from four locations in May 2009 (Table 1, Fig. 1). Colonized halite crusts were collected at two locations (Salar Grande, SG; Yungay, Yu). The halites are evaporitic crusts specific to the hyperarid core of the Atacama Desert. Yungay is the driest of the sampling sites and is the same area where previous studies have demonstrated low microbial abundances in soils and rocks (Navarro-González et al., 2003; Connon et al., 2007; Lester et al., 2007). Salar Grande is farthest north, and while still very dry, this location experiences high humidity and fogs (de los Ríos et al., 2010). At both locations, the halite is colonized by a unique species of cyanobacteria, with associated heterotrophic bacteria and archaea (de los Ríos et al., 2010). Colonized gypsum and ignimbrite samples were collected in the Monturaqui meteorite impact crater region. Fragments of rocks at the Monturaqui sampling site (Fig. 1) were classified as ignimbrite according to Wierzchos et al. (2013). Here, rainfall is higher than at Yungay, but the mean annual relative humidity can be lower (Wierzchos et al., 2012, 2013). At each location, surface rock samples were collected with a rock hammer and sealed in sampling bags with desiccant. Samples were stored at room temperature in the dark until they were returned to the lab 10 days later when they were stored in a freezer at −20°C until analysis. Samples were analyzed within 6 months of collection. Prior to extraction, samples were homogenized with a mortar and pestle.
The reported TOC content of the halites is for the desalted material. PLFA and GLFA concentrations are the total identified fatty acids per gram of dry weight of rock. PLFA and GLFA uncertainties are the standard deviation of three replicate lipid extractions.
2.2. Organic carbon preparation
Inorganic carbon was removed by treating dry samples with 1 M HCl. To ensure enough carbon for δ13CTOC analysis, and to minimize interactions of salt with quartz, halite samples were desalted by soaking the samples in Milli-Q water overnight, centrifuged, decanted, and subsequently dried at 45°C to remove any remaining water. The δ13CTOC of the pretreated samples was determined on an EA-Delta XL at McMaster University, and triplicate analyses gave a precision of ±1‰ for the δ13CTOC. Prior to desalting, halite samples contained 0.2–0.4% total organic carbon (TOC). After desalting, the percent TOC of the halite samples was an order of magnitude higher. δ13C TOC values of the halite samples before and after desalting were the same within 1‰.
2.3. Membrane lipid extraction and identification
Homogenized rock samples (∼100 g) were extracted twice by the modified Bligh and Dyer process (White et al., 1979). The organic phase was collected, and the GLFA and PLFA fractions were separated via silica gel chromatography into the neutral lipids fraction (DCM), glycolipid fraction (acetone), and phospholipid fraction (methanol) (White et al., 1979). The neutral lipid fraction was not subjected to further processing and was stored for future analysis. The glycolipid and phospholipid fractions were retained and processed separately. Each fraction was evaporated to dryness under a stream of nitrogen gas and reacted to fatty acid methyl esters (FAME) via mild alkaline methanolysis (Guckert et al., 1985). FAME were further purified with a second silica gel and eluted with 5 mL of hexane, 5 mL of dichloromethane, and 5 mL of methanol.
An Agilent 6890 gas chromatograph coupled to an Agilent 5973 quadrupole mass spectrometer was used for identification and quantification of FAME. Chromatographic separation was performed with either a DB-5 MS column (30 m×0.25 mm, 0.25 μm film thickness) or an HP-88 MS column (100 m×0.25 mm, 0.25 μm film thickness). FAME were quantified by using commercially available (Supelco, Inc) calibration standards methyl tetradecanoate (C14:0), methyl hexadecanoate (C16:0), methyl octadecanoate (C18:0), and methyl eicosanoate (C20:0). FAME were identified with the use of several bacterial reference standards, (Bacterial Acid Methyl Esters Mix from Matreya, Inc., and 37 FAME mix from Supelco), mass-fragmentation patterns, and retention times. Lipid concentrations are reported in moles of FAME normalized to the total mass of rock used prior to desalting (i.e., nmol/g). Since endolithic communities often colonize rock in laminae and we were not able to isolate these laminae, lipid concentrations in the colonized regions may be higher than those reported here. Relative lipid abundances are reported in mole percent of the lipid fraction. The limit of detection was 0.05 nmol/g, an order of magnitude lower than the least-concentrated sample.
2.4. Gas chromatography—isotopic ratio mass spectrometry analysis of FAME
Stable carbon isotope ratios (δ13C) of individual FAME were determined by an Agilent 6890 gas chromatograph coupled to a Thermo Delta Plus XP isotope mass spectrometer via a Conflo III interface. Reproducibility for isotopically characterized hexadecane and octacosane laboratory standards was better than 0.3‰. Precision for microbial FAME was typically 0.5‰ minimum of three analyses, and all analyses were less than 1‰.
2.5. Radiocarbon analysis of FAME and TOC
The 14C content of FAME was determined by using two types of preparation techniques. First, concentrated FAME extracts were purified and concentrated with preparative capillary gas chromatography per Slater et al. (2005). FAME were separated on an Agilent gas chromatograph (60 m×0.53 mm DB-5 column, 0.25 μm film thickness) interfaced with a Gerstel preparative fraction collection system where samples were trapped in prebaked Pyrex U-tubes at <0°C. U-tubes were rinsed with 1 mL of dichloromethane into prebaked gas chromatograph autosampler vials. Samples were rerun via gas chromatograph–mass spectrometer to check for purity. The second method of 14C analysis of FAME measured the Δ14C of whole FAME samples, without any chromatographic separation (i.e., ΣPLFA and ΣGLFA). All FAME and pretreated TOC samples were transferred to prebaked 6 mm quartz tubes with an additional milliliter of dichloromethane, and the solvent was dried under a stream of ultrahigh-purity nitrogen. Cupric oxide and silver wire were added to the tubes, and they were evacuated to 10−6 torr before being flame-sealed under vacuum. Tubes were then heated to 850°C for 2 h. The resulting CO2 was purified, quantified, and reduced to graphite by using standard procedures (Santos et al., 2007). 14C measurements were performed at the KECK Carbon Cycle Accelerator Mass Spectrometry Laboratory at the University of California Irvine. The reported data is in Δ14C notation expressed in per mill, which is the deviation of a sample from the 95% activity in 1950 A.D. of Natural Bureau of Standards oxalic acid 1 normalized to δ13C=−25‰ with respect to Pee Dee Belemnite (Stuvier and Polach, 1977). Both δ13C and Δ14C values of the FAME were corrected during data processing for the addition of methanol carbon during transesterification. The measurement uncertainty for Δ14C of TOC values is the observed variation on three replicates (±4–9‰), which was greater than the accelerator mass spectrometry (AMS) reported error. The uncertainty associated with the Δ14C of PLFA and GLFA values (±20‰) reflects the total measurement uncertainty, which includes the uncertainty from the AMS measurement as well as sample preparation and handling. Since the two preparations of FAME (individual FAME and ΣFAME) were found to be within our 14C analytical error, we report all Δ14C data as ΣPLFA and ΣGLFA.
2.6. FAME nomenclature
The methyl ester derivatives of PLFA and GLFA are designated in terms of the total number of carbon atoms:number of double bonds. The symbol indicates that the first carbon-carbon double bond starts on the n th carbon from the CH3 end. The prefixes i- and a- refer to iso-branched and anteiso-branched, respectively.
3. Results
3.1. Rock TOC, 13C, 14C
The TOC content of the rock samples ranged from 0.1% to 0.4% (Table 1). The δ13CTOC and Δ14CTOC values are given in Table 2, and Δ14CTOC values are plotted in Fig. 4. Among the various rock types, δ13CTOC was alike (varied by 6.7‰, n=4) and similar to the expectation of C3 photosynthesis (Farquhar et al., 1989). The 14C content of TOC ranged from values consistent with the atmosphere after weapons testing in the 1950s through to more depleted radiocarbon values consistent with a 14C age of a few hundred years. The TOC sample that was most enriched in radiocarbon had a Δ14C value of +75‰, and the most depleted had Δ14C value of −34‰.
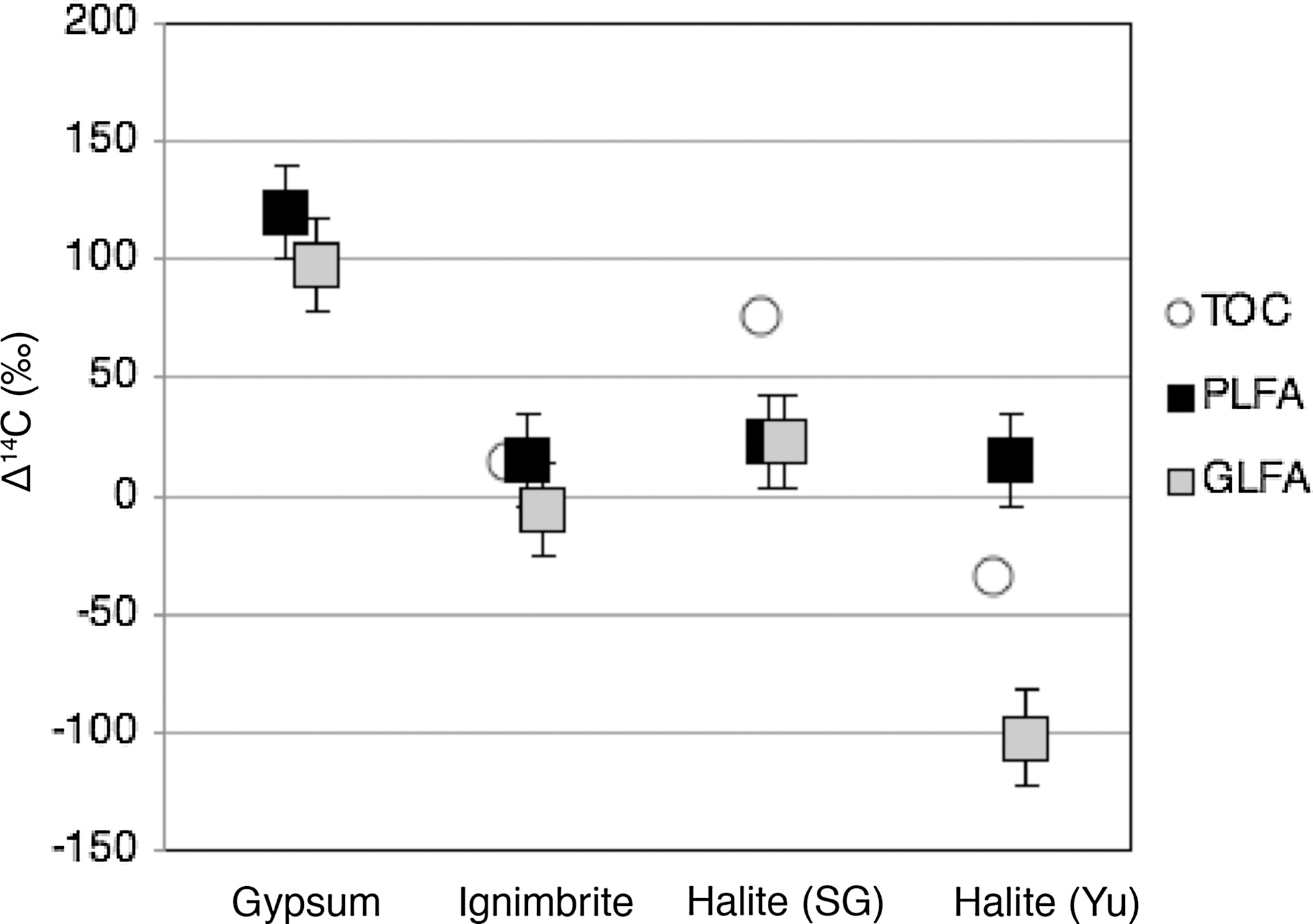
Δ14C values for the TOC, PLFA, and GLFA of endolithic communities in the Atacama Desert. Error bars are shown for all points, and if not visible they are smaller than the symbol. As noted in Table 2, analytical limitations prevented us from determining the TOC Δ14C of the gypsum.
The uncertainties of all δ13 measurements were ±1‰. Uncertainties of Δ14CTOC are the observed variation of replicate analyses. The Δ14C of PLFA and GLFA were measured as a compound class, and replicate Δ14C values are listed. Δ14CPLFA and Δ14CGLFA uncertainties are±20‰, which is larger than the standard deviation of replicate Δ14CPLFA and Δ14CGLFA values of replicate samples with the exception of Yungay, where we observed a 25‰ difference between replicates.
δ13CTOC values of desalted and salty samples were within 1‰. 14CTOC was determined on the desalted samples.
n.d. is not detected. Due to the fact that gypsum is composed of hydrated minerals, we were not able to successfully convert the sample to CO2 for 14C analysis due to the large volume of water.
3.2. Phospholipid- and glycolipid-derived FAME
Phospholipid fatty acid concentrations (nmol PLFA/g rock) varied by almost 2 orders of magnitude, from 0.6 nmol/g in the halite at Yungay to 40 nmol/g in the gypsum (Table 1). Halite samples from Salar Grande had a PLFA abundance of 10 nmol/g, while at Yungay the abundance of PLFA was more than an order of magnitude lower (0.6 nmol/g). The concentration of PLFA normalized to the TOC content was much greater in the nonhalite samples than the halite samples (18 μmol/g TOC vs. 1.5 μmol/g TOC, respectively). Using a generic conversion factor of 2×104 cells pmol−1 (Green and Scow, 2000), we converted the PLFA concentration to cell abundance. The calculated cell abundance ranged from 8×108 cells/g in the gypsum to 1.2×107 cells/g in the halite at Yungay. Endoliths typically colonize rock in thin layers beneath the rock surface; however, to have enough material for isotopic analysis we subsampled sections of rocks that could have included uncolonized zones, and as such estimated cell abundances are likely to be higher than 107 to 108 cells/g in the area of highest colonization. The observed FAME were grouped into four compound classes: saturated, monounsaturated, polyunsaturated, and branched FAME; these are reported in Table 3. The abundance of microbial GLFA (nmol/g) was linearly related to the PLFA (r 2=0.96) and was highest in the gypsum and lowest in the samples from Yungay (Table 1).
Lipid isotopic values: δ13C values
The PLFA and GLFA δ13C values were lower than the TOC (Table 2). Due to the low biomass in these samples, δ13CPLFA and δ13CGLFA were determined only for the most abundant FAME (C16:0 and C18:1). The TOC was 10±2‰ more enriched in 13C than both PLFA and GLFA for samples in the same rock type. The gypsum δ13CPLFA values were −31.6±1‰ and −30.3±1‰ for C16:0 and C18:1, respectively, while the ignimbrite rock δ13CPLFA values were within error (δ13CPLFA=−29±1‰ for C16:0 and C18:1). At Salar Grande, the δ13CPLFA of C16:0 and C18:1 were lower than the nonhalite samples, with values of −34.7±1‰ and −32.9±1‰, while the C16:0 and C18:1 δ13CPLFA values at Yungay were the same within error (δ13CPLFA=−33.0±1‰). For the ignimbrite rock, gypsum, and Yungay samples, the GLFA were within error of the δ13CPLFA values for all samples, while at Salar Grande the δ13CGLFA of C16:0 was lower (−31.2±1‰) than the δ13CPLFA of C16:0 (−34.7±1‰).
Lipid isotopic values: Δ14C values
The 14C content of replicate analysis of lipids is given in Table 2, with the average values plotted in Fig. 4. The PLFA in the gypsum were the most enriched in radiocarbon of all samples (Δ14CPLFA=+120±20‰). Δ14CPLFA values were similar in the ignimbrite (Δ14CPLFA=+15±20‰), Salar Grande halite (Δ14CPLFA=+30±20‰), and Yungay halite (Δ14CPLFA=+15±20‰). The Δ14CGLFA values were within 2σ (40‰) of the Δ14CPLFA for all samples except Yungay. The Δ14C of GLFA at Yungay was the lowest for all lipid or TOC samples at −102±20‰.
4. Discussion
4.1. Endolithic communities use atmospheric CO2: evidence from carbon isotopes
Atmospheric CO2 is the primary carbon source for the endolithic communities examined from the Atacama Desert based on 14C and 13C analyses (Fig. 1). The Δ14C value of atmospheric CO2 is estimated to have been +50‰ in 2007 (Czimczik and Welker, 2010) and decreasing at ∼5‰ per year (Levin et al., 2008). When these samples were collected, the Δ14C of the atmospheric CO2 was estimated to be +40±10‰. The predominant Δ14C signal of the microbial membranes, or PLFA, was positive (i.e., Δ14C>0‰) and within error of the modern atmosphere (Table 2). These results demonstrate that carbon within the cellular membranes is consistent with present-day atmospheric CO2 and that carbon within this system cycles quite quickly.
While the endolithic communities contain 14C that is consistent with the modern atmosphere, some samples were even more enriched in 14C than the modern atmosphere. For example, the gypsum sample had a Δ14CPLFA=+120±20‰, and the biomass, as represented by the TOC, at Salar Grande had a Δ14CTOC of +75±5‰. Since 14C undergoes decay over time and before the 1950s the atmospheric CO2 Δ14C was ∼0‰, the only way these samples could have a positive Δ14C is if the endolithic communities incorporated atmospheric “bomb carbon” into their cells and have retained this carbon within the endolithic system since its incorporation. The implications of this observation with respect to carbon cycling will be discussed further below.
Microbial utilization of modern atmospheric CO2 is also supported by the δ13C of lipids and TOC. The TOC and PLFA δ13C results indicate that Atacama endolithic communities contain autotrophic organisms and are using atmospheric CO2 as their primary source of carbon. In 2009, the atmospheric δ13C of CO2 was −8‰ (Keeling et al., 2010). The δ13C offsets between the atmospheric CO2 and the biomass within the rock are small (11–17‰, Table 2). Observation of reduced isotopic discrimination during autotrophy is generally interpreted as evidence of carbon limitation (Hayes, 2001), and the extent of carbon limitation in these systems is discussed further below. The δ13C offsets between the FAME and the TOC are between 8‰ and 10‰, which are consistent with autotrophy (Hayes, 2001). Such an offset is typical of bacterial communities but does not exclude the presence of eukaryotes (Sakata et al., 1997). The GLFA and PLFA δ13C values were within 2σ and thus indistinguishable from each other. The similarity in the 13C/12C ratio of the two types of lipids may indicate that the same substrate was used to form these lipids (Abraham et al., 1998). These isotopic results, indicating the presence of an autotrophic community, are in agreement with previous studies that identified the presence of cyanobacteria in the microbial communities of the Atacama Desert (Warren-Rhodes et al., 2006; Wierzchos et al., 2006, 2011, 2013; de los Ríos et al., 2010).
4.2. Cycling time of carbon within the endolithic communities
C cycling within the 14C-enriched gypsum sample
Even though the 14C content of PLFA varied by location, all samples contained contemporary carbon. The most radiocarbon-enriched PLFA were in the gypsum. The Δ14CPLFA value of the gypsum, +120±20‰, corresponds to the atmospheric Δ14C CO2 value either in 1960, when the Δ14C was rising due to nuclear weapons testing (Fig. 3), or more recently (e.g., circa 1994), when the atmospheric Δ14C CO2 was decreasing. A simple two end member mixing model between the Δ14C of the present-day atmosphere and the highest Δ14C CO2 of the bomb peak in 1964 indicates that 13% of the C in the microbial membrane lipids derives from the height of the bomb peak in 1964 and the remainder of the lipids has a Δ14C from the present-day atmosphere. However, as it is unlikely that no atmospheric carbon was incorporated into the cellular membranes between 1964 and the present, this estimate is overly simplified.
To estimate the rate of carbon cycling in the endoliths, we created a yearly integrated mixing model. We assumed that within the rock interior there was a pool of CO2 that originated from either the atmospheric or recycling of relict carbon (Fig. 2). Relict carbon is carbon that has remained within the system from previous years either as preserved intact compounds or as organic matter metabolized to CO2 that is subsequently refixed via autotrophic activity. Therefore, at any given time the total amount of carbon (C
total=1) in the system can be expressed as
where NAC is the fraction of new atmospheric carbon incorporated into the cells and RC is the fraction of relict carbon. If C
total in the system is constant and there is no net accumulation, Eq. 1 can be rewritten as
If the rate of incorporation of NAC is the same through time, that is, that the rate of carbon cycling is constant with the same proportion of new atmospheric carbon and relict carbon being incorporated in any given year, then the radiocarbon signature of the system can be expressed as an isotopic mass balance of Eq. 1:
where in each year (y) there is incorporation of new atmospheric carbon that reflects the atmospheric CO2 Δ14C for that given year (i.e., Fig. 3) and a certain fraction (1 - NAC) that has the Δ14C signature of the remaining carbon. Because the predominant signal was that our Δ14CTOC values are similar to our Δ14CPLFA values, we can assume no significant net accumulation, and the system is in equilibrium. Then the carbon turnover time (tt, years) can be expressed as
where NAC
t
is the annual fraction of new atmospheric carbon (yr−1). So over a given number of years, the yearly resolved resultant isotopic signature of the viable microbial community can be expressed as
Then using the known values of atmospheric CO2 Δ14C over the past 50 years, (Levin and Kromer, 2004; Hua et al., 2006), we iteratively solved Eq. 5 for NAC until the modeled Δ14C y reflects the measured Δ14C of the microbial lipids.
Using the above model, we estimated that carbon in the gypsum samples had a turnover time of 28±5 years. Our model results suggest that each year 3.6±0.6% of the carbon fixed and incorporated into the lipids was due to new autotrophic production in which contemporary atmospheric CO2 was used and the remainder was recycled carbon from respiration of previously fixed organic matter. Notably, since the measured Δ14CTOC in our samples from the hyperarid core of the Atacama were never lower than pre-bomb values (Δ14C=0‰), this suggests the absence of a large pool of pre-bomb nonlipid carbon being recycled within the system (i.e., alkanes). Thus, the above estimate of new atmospheric carbon incorporation may be a lower limit of the community productivity, and it is likely that the endoliths are cycling carbon more quickly than every few decades.
There are several points to consider with respect to this estimate of the rate of carbon turnover. If the system contained pre-bomb carbon that was being recycled even more slowly and undergoing radioactive decay, this would require the estimated rate of new autotrophic production to be faster in order to compensate for the depleted 14C from the recycled carbon. However, there is currently no evidence that old carbon is being incorporated into the lipid membranes of the microbes inhabiting the gypsum. Furthermore, if the system was not in steady state, as assumed in Eq. 4, there would be net accumulation of C within the system, and that would lead to a higher rate of new atmospheric carbon incorporation. Previously, it has been estimated that the net ecosystem productivity of Antarctic endolithic communities is 2 orders of magnitude smaller than the rate of primary productivity (Friedmann et al., 1993). If there was net ecosystem productivity, it would lead to an increase in TOC over time in the rock. While accumulation of TOC over time is certainly possible, we do not have enough data to assess the extent of such an increase. Additionally, the limited space that endolithic communities inhabit precludes unlimited expansion and accumulation of organic matter. In any case, it is not expected that such factors would affect the estimated cycling rate to a large extent. Thus, based on this estimate it appears that the microbial membranes of endolithic communities within the gypsum are enriched in carbon from atmospheric weapons testing during the 1950s and 1960s, and we estimate that the carbon within the viable microbial community is turned over in the system within 30 years.
“Modern” samples
If all the carbon in the lipids of the endoliths was from new atmospheric carbon in 2009 (i.e., atmospheric CO2), then the Δ14CPLFA should be close to +40‰. The 14C content of the PLFA in the ignimbrite and that of the halite samples are within analytical uncertainty of each other and atmospheric CO2 Δ14C. Therefore, the simplest explanation is that the endoliths cycle quickly (i.e., monthly to yearly) and that neither bomb carbon nor more 14C-depleted pre-bomb carbon is present within the viable cells. If the endoliths incorporated a large component of bomb carbon, then the endoliths would be more enriched in 14C (i.e., Δ14CPLFA >>+50‰). Conversely, if older pre-bomb carbon (i.e., stored TOC or carbonate) was being recycled to CO2 and then incorporated as a major component into the lipids, the 14C content would be much lower than what we observed (i.e., Δ14CPLFA <<+50‰).
The observed Δ14C values could also be possible due to a balance of small inputs of bomb at the same time as inputs of older carbon. To have PLFA with a Δ14C close to the present-day atmospheric value, while also turning over carbon every few decades as in the gypsum, the endoliths at the other sites must also be incorporating older carbon into their cellular membranes that is balancing out the bomb carbon inputs exactly. Such a balance of old and young 14C inputs could mask the rate of carbon cycling within the system. However, due to the presence of the bomb spike, and the reasonable assumption that recently produced organic matter will cycle more quickly than older carbon, it is unlikely that such older carbon inputs can be a major contributor to the carbon pools. While the data cannot resolve this possibility definitively, there is evidence of some preservation of older carbon.
Older carbon
If endolithic communities are turning over carbon very slowly, such as has been previously suggested (Bonani et al., 1988; Sun and Friedmann, 1999), or if the microbes are incorporating fossil carbon such as carbonate-derived CO2 into their cells, these processes would be reflected in our observed Δ14C values for TOC, PLFA, and GLFA. To reach our observed Δ14CTOC values if 10% of the TOC originated from fossil sources (i.e., Δ14C=−1000‰), the remaining 90% of the C would have to be much more enriched (i.e., more bomb C and less C from the present day). For example, at Salar Grande, to obtain observed Δ14CTOC=+75‰ and incorporate 10% of the TOC from fossil sources, the remaining, nonfossil TOC would have to have a Δ14C=+194‰. Such an enriched 14C value is possible if carbon was fixed either in the early 1960s or 1987 and not since then, when the Δ14C CO2 has been lower. It is unlikely that the microbes have not fixed any carbon in the last few decades. An alternative scenario could be that the microbes contain at least three different pools of carbon: fossil carbon from carbonate, carbon from the peak of the bomb curve, and carbon from the contemporary atmosphere. However, this issue cannot be resolved in more detail with the present data, and it is unlikely that carbonate or other older sources of carbon are contributing a significant fraction of carbon to these endolithic communities.
δ13C systematics
The Δ14C model results indicate that the majority of the carbon being utilized by the microbial communities is recycled carbon. While more difficult to constrain due to the potential for variations in both carbon source δ13C and isotopic discrimination during photosynthesis, the Δ14C model must be reconciled with the δ13C observations. In the case of the gypsum, the Δ14C model indicated that 96.4% of the internal CO2 pool is derived from respiration of organic matter. Respiration-derived CO2 making up the majority of this pool would be expected to have a δ13C equivalent to the organic matter from which it was derived (δ13C=−19.6‰), and the entire pool would be expected to have a slightly higher δ13C due to inputs from the atmosphere (δ13C=−8‰). If this is the case, the isotopic discrimination between the internal CO2 pool and the organic matter is in fact even smaller than the estimate based on the atmospheric CO2 δ13C. This implies that CO2 conversion is approaching quantitative utilization of all available CO2 within the gypsum endolithic community. Such an explanation is entirely consistent with the Δ14C model results. The results for the other three sites are likewise consistent with the explanation that the microbial communities utilize almost all available CO2 during photosynthesis. The slightly greater δ13C depletions in TOC at the halite sites may indicate that there is a slightly greater isotopic discrimination being expressed during photosynthesis; thus these systems are slightly more open. However, due to the potential for variation in both δ13C of the carbon source and isotopic discrimination during photosynthesis, it is not possible to state this categorically.
Older carbon at Yungay
While our Δ14CPLFA findings indicate that these microbes cycle quickly within the viable cellular membranes, the Δ14CTOC and Δ14CGLFA at Yungay suggest the potential for preservation of specific compounds within these communities. The Δ14CGLFA at Yungay was −102±20‰, which equates to carbon that was last in equilibrium with the atmosphere a thousand years ago. One possible explanation for this old carbon in microbial membrane lipids is that the microbes that are producing the GLFA are incorporating older carbon into their membrane lipids. However, since microbes that produce both PLFA and GLFA have been observed to use the same substrates to produce each type of lipid (Abraham et al., 1998), and the observed Δ14CPLFA and Δ14CGLFA are very different, it is possible that the GLFA at Yungay are not from viable cells. This was the only location where the PLFA to GLFA ratio was less than one. It is possible that the GLFA at Yungay are being preserved, sealed in halite fluid inclusions, or both. It has been shown that many microbial cells and their biomolecules could be preserved within halite liquid inclusions (e.g., Schubert et al., 2009; Lowenstein et al., 2011). The greater amount of GLFA may be due to two processes: (1) the PLFA are preferentially hydrolyzed and (2) the GLFA are a result of larger molecules, such as PLFA, being hydrolyzed to form GLFA (Lechevalier and Moss, 1977). These results suggest that viable microbial community cycles carbon quickly and there may be preservation of the GLFA.
5. Implications for the Dry Limit of Life
The hyperarid zone of the Atacama Desert has long been considered one of the most inhospitable places on Earth. This is best exemplified by the total absence of higher organisms and by the extreme scarcity of soil microorganisms (Navarro-González et al., 2003). Similar to other extreme deserts, such as the Dry Valleys of Antarctica, life in the Atacama Desert seeks refuge within rocks as conditions become increasingly dry. We would expect that carbon cycling within the lithic communities in the hyperarid core would be very slow and similar to the Dry Valleys of Antarctica, where sandstone endolithic communities have been estimated to grow on the timescales of 103 to 104 years (Sun and Friedmann, 1999). Our results show that even halite samples from Yungay, the driest part of the desert, contain carbon from modern atmospheric CO2 and confirm that active microbial life is still possible in this extremely dry environment, even when soils cease to be a habitat for life (Navarro-González et al., 2003). It has also been assumed that different lithologies would create different environmental conditions within the endolithic habitat that would influence the rate of microbial activity; however, the fact that the samples from different lithologies cycled carbon at similar rates suggests that the lithology may not be as important as previously thought.
This work is particularly relevant for the search for life on Mars, where extremely dry conditions have existed for at least the last 3 billion years. Our results indicate that, if life ever existed on Mars, the interior of hygroscopic salts would have likely been a place on the surface of the planet where microorganisms could have been active (Davila et al., 2010). Therefore, future efforts to search for evidence of life on Mars ought to target these types of lithologies to maximize the chances of success.
Footnotes
Acknowledgments
Jennie Kirby, Martin Knyf, and Sheila Griffin for technical assistance. Funding to L.A.Z. from the Canadian Astrobiology Training Program and McMaster University Origins Institute. Funding to G.F.S. from Natural Science and Engineering Council grant 288309-09. Funding to A.F.D. from the NASA Exobiology Program. Funding to J.W. from grant CGL2010-16004 from the Spanish Ministry of Science and Innovation. Two anonymous reviewers and Dr. Sherry Cady for providing helpful suggestions that improved this manuscript.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
AMS, accelerator mass spectrometry; FAME, fatty acid methyl esters; GLFA, glycolipid fatty acids; PLFA, phospholipid fatty acids; SG, Salar Grande, TOC, total organic carbon; Yu, Yungay.