
Abstract
Understanding the behavior of proteins under freezing conditions is vital for detecting and locating extraterrestrial life in cold environments, such as those found on Mars and the icy moons of Jupiter and Saturn. This review highlights the importance of studying psychrophilic “cold-shock” proteins, a topic that has yet to be explored. A strategy for analyzing the psychrophilic RNA helicase protein CsdA (Psyc_1082) from Psychrobacter arcticus 273-4 as a key protein for life under freezing temperatures is proposed. The experimental model presented here was developed based on previous data from investigations of Escherichia coli, P. arcticus 273-4, and RNA helicases. P. arcticus 273-4 is considered a model for life in freezing environments. It is capable of growing in temperatures as cold as −10°C by using physiological strategies to survive not only in freezing temperatures but also under low-water-activity and limited-nutrient-availability conditions. The analyses of its genome, transcriptome, and proteome revealed specific adaptations that allow it to inhabit freezing environments by adopting a slow metabolic strategy rather than a cellular dormancy state. During growth at subzero temperatures, P. arcticus 273-4 genes related to energy metabolism and carbon substrate incorporation are downregulated, and genes for maintenance of membranes, cell walls, and nucleic acid motion are upregulated. At −6°C, P. arcticus 273-4 does not upregulate the expression of either RNA or protein chaperones; however, it upregulates the expression of its
1. Introduction
C
Recent studies have found evidence for water on Mars and on icy moons in the outer solar system (Maltagliati et al., 2011). Water vapor detected in Saturn's moon Enceladus suggests the presence of a liquid ocean beneath its icy surface (McKay et al., 2008). Jupiter's moon Europa is thought to harbor a liquid salty ocean under an icy crust of controversial thickness, with melt pockets within the ice shell itself (Marion et al., 2003; Clark et al., 2011; Schmidt et al., 2011). Jupiter's largest moon Ganymede is thought to have a thicker ice shell than that of Europa, with an interior ocean between ice layers (Patterson et al., 2010). Mars, sometimes referred to as an “astrobiological laboratory,” hosts a permafrost layer, thick water ice deposits at the poles, and may even experience surface water flow as brines (Davila et al., 2010; Glamoclija et al., 2010). The probability of liquid water as well as water ice in polar sheets or contained within permafrost soils on these planets is promising for finding extraterrestrial life and potentially psychrophilic extraterrestrial life.
Organisms that thrive in cold environments are referred to as psychrophiles. Psychrophiles can exist at temperatures ranging from 20°C to below 0°C, with an optimum temperature at 15°C (Morita, 1975). They are the most abundant extremophiles on Earth in terms of biomass, diversity, and distribution (D'Amico et al., 2006). While psychrophilic organisms are widely studied, only a small fraction (0.1%) of catalogued proteins in the Protein Data Bank (PDB) is from psychrophiles, which highlights the necessity to better characterize those proteins. Moreover, the set of proteins that are overexpressed by cells as a response to a drop in temperature, referred to as cold-shock proteins, consist of only 45 individual entries in the PDB, none of which are characterized from psychrophiles. Comparing the amount of research on psychrophiles and their importance for life in cold environments with the numbers of studied proteins that assist life in such environments shows that there is a need to characterize and understand the nature of psychrophilic “cold-shock” protein structures.
Proteins are an important functional component of highly complex surviving systems in specific environments. The environment is one of the factors that determine cell physiology, protein properties, and functionality. This paper presents an experimental model for assessing the psychrophilic protein RNA helicase CsdA - Psyc_1082, from Psychrobacter arcticus 273-4, as a key protein for life under freezing temperatures. The data analyzed to build the experimental model relies on results from previously conducted investigations that are reviewed in this article. Essentially, this paper is designed to highlight the importance of studying psychrophilic “cold-shock” proteins. Testing of mesophilic cells that express psychrophilic proteins for resistance and tolerance to colder and subzero temperatures will help in understanding how psychrophilic cells can exist physiologically in their “extreme” environments. Analyzing psychrophilic “cold-shock” proteins will provide important knowledge of biological sensitivity and tolerance to cold and will give insight into the key cellular workers that sustain life in subzero conditions.
2. Molecular Strategies to Cope with Cold Temperatures
Cold-shock proteins (CSP) are a set of proteins overexpressed by mesophilic cells, which live between 10°C and 42°C, with an optimum of ∼37°C, to adjust the cellular machinery to an abrupt decrease in temperature. Psychrophiles passed through a long-term evolutionary adaptation process, and proteins that were nominated as CSP in mesophiles are expressed as housekeeping proteins in psychrophiles (Hébraud and Potier, 1999; Srinivas and Ray, 2006).
Cold-shock response was first studied in mesophiles, with Escherichia coli being the best characterized (Smith et al., 1994; Panoff et al., 1998; Yamanaka, 1999). In mesophiles, a drop in temperature represses the expression of most protein syntheses, and the cell enters a lag growth phase called the “acclimation phase” (Yamanaka, 1999). During this phase, three processes can happen: (1) the cell is not able to recover cellular activity and dies, (2) the cell enters an inactive dormancy state, or (3) the cell induces the expression of a specific group of proteins to recover the cellular physiology. CSP are one of the types of proteins that are mainly expressed by mesophiles in the lag phase to recover the cell physiology. The other type is referred to as cold-acclimation proteins (CAP). In the lag phase, after the cold shock, CSP levels are increased suddenly for a short period, whereas CAP expression increases gradually and does not fall as fast as CSP (Srinivas and Ray, 2006). Additionally, some of the proteins considered CSP in mesophiles are characterized as CAP in psychrophiles.
Psychrophiles developed strategies to overcome the challenges dictated by permanently cold environments, like the reduction of enzyme activity, decrease in membrane fluidity, alteration of nutrient transport, decrease in rates of transcription, translation, and cell division, protein cold-denaturation, inappropriate protein folding, and intracellular ice formation (Thieringer, 1998; Hébraud and Potier, 1999; Yu and Owttrim, 2000; Srinivas and Ray, 2006). Psychrophilic enzymes need to enhance their catalytic efficiency at low temperatures, which therefore allows cells to express more flexible cold-adapted enzymes to counterbalance the decrease in affinity for substrates (Feller and Gerday, 1997; Siddiqui and Cavicchioli, 2006; Chiuri et al., 2009). Structural and molecular analyses of psychrophilic enzymes by homology modeling, nuclear magnetic resonance, crystallography, and the metagen/transcript/proteomic approach have shown that the increase of protein stability and flexibility in cold-adapted proteins is the result of a combination of several processes (Deming, 2002; Gianese et al., 2002; Bae and Phillips, 2004; Watanabe and Takada, 2004; Grzymski et al., 2006; Feller, 2007; Rodrigues and Tiedje, 2008; Rodrigues et al., 2008; Bergholz et al., 2009; Metpally and Reddy, 2009; Ayala-del-Río et al., 2010; Casanueva et al., 2010; Feller, 2010). Such processes include a decrease in the protein core hydrophobicity, which increases cold-temperature reaction rates and enzymatic efficiency to structure-dependent processes such as substrate and DNA/RNA binding; a decrease in strength of intramolecular bonds (fewer hydrogen bonds and salt bridges); a decrease in aliphatic residues; a decrease in salt-bridge-forming residues such as arginine, glutamic acid, and aspartic acid; a decrease in the number of proline and arginine residues (these residues are known to increase the entropy of unfolded proteins) (Linding et al., 2003); an increase in the entropy (thermodynamic disorder) to sustain macromolecular dynamics and function; an increase in the number of hydrophobic side chains that are exposed to the solvent; and an increase in the number of glycine and lysine residues. Not all of these processes are present in the same cold-active enzyme, but this list represents some of the changes observed by comparing psychrophilic enzymes to their mesophilic and thermophilic homologues (Russell, 2000; Ayala-del-Río et al., 2010).
3. Psychrobacter arcticus 273-4: A Model of Life under Subzero Conditions
The genus Psychrobacter is frequently detected in permafrost soil, which constitutes 20–25% of Earth's continental surface (Rodrigues et al., 2009). P. arcticus 273-4, a strain isolated from a continuously frozen permafrost soil 20,000 to 30,000 years old within the Kolyma lowland region of Siberia, is considered a model among microorganisms for life in freezing environments (Bakermans et al., 2006, 2009). Inhabiting permafrost with temperatures ranging from −10°C to −12°C, P. arcticus 273-4 was characterized by its responses to potential stresses imposed by the environment (Ponder et al., 2005). It grows in temperatures from −10°C to 28°C, with an optimum at 17°C, using physiological strategies to survive under freezing temperatures, low water activity, limited nutrient availability, and gamma radiation for long periods of time (Ponder et al., 2005, 2008). The genome of this strain has been sequenced, and its transcriptome and proteome have been analyzed under subzero and positive temperatures (Bergholz et al., 2009; Ayala-del-Río et al., 2010). Together, all studies reveal specific adaptations that allow this strain to inhabit freezing environments by adopting the slow metabolic strategy (Bergholz et al., 2009; Ayala-del-Río et al., 2010). Finally, P. arcticus 273-4, at freezing temperatures, does not upregulate the expression of either RNA or protein chaperones (proteins which help in the folding of other proteins and RNA molecules), but it does upregulate the expression of its CsdA - Psyc_1082 (Bergholz et al., 2009).
4. CsdA: A Key Protein for Life under Subzero Conditions
DEAD-box RNA helicases, members of the DEAD-box family, are found in the three domains of life and are known to participate in different processes of RNA metabolism (Rocak and Linder, 2004; Cordin et al., 2006). All DEAD-box helicases present a structurally conserved core element, which contains the nine conserved motifs that are required for enzyme activity in a fragment of ∼400 amino acids (Rocak and Linder, 2004) (Fig. 1). Domains Ia, Ib, IV, and V are believed to bind to the substrate. Q-motif, Motif I, Motif II (the DEAD motive), and Motif VI are known requirements for ATP hydrolysis (for review, Rocak and Linder, 2004).

DEAD-box helicases conserved core element. Alignment of P. arcticus 273-4 Psyc_1082 (CsdA) (YP_264368) with homolog proteins from three psychrophiles [Pcryo_1379: Psychrobacter cryohalolentis K5 (YP_580642); PsycPRwf_1: Psychrobacter sp. PRwf-1(ABQ94096); M. burtoni: Methanococcoides burtonii DSM 6242 (ABE52831)], a mesophile [E. coli K1: E. coli K12 (EFK25707)], and a thermophile [M. jannasc: Methanocaldococcus jannaschii DSM 2661 (AAB98663)]. Black boxes indicate 100% of identity between sequences. Protein domains are identified under alignment (F-Q-I-Ia-Ib-II-III-IV-V-VI). The genome data and gene information of P. arcticus 273-4 was extracted from IMG/JGI (Markowitz et al., 2008). The CsdA - Psyc_1082 was aligned to the GenBank/NCBI database by BLASTp against the non-redundant protein sequences (nr). Two hundred and fifty protein sequences were extracted from the database and aligned by using Clustal W to find conserved regions/motives and for protein domain recognition (Larkin et al., 2007).
DEAD-box RNA helicase has been proposed to participate in a variety of processes such as in the initiation of translation, ribosome biogenesis, stabilization of mRNAs when overexpressed, mRNA processing and decay in a cold-shock response (as a component of degradosomes), unwinding double-stranded RNA, and gene regulation. The specificity of which of these processes it plays a role in is believed to rely on the flanking, less-conserved sequence of the proteins (Rocak and Linder, 2004).
CsdA, a DEAD-box RNA helicase, is recognized as an essential player in regulating gene expressions of certain proteins during microorganism growth at low temperatures because of its helicase activity. CsdA promotes the degradation of double-stranded mRNA stabilized at low temperatures allowing the translation of the mRNA by the ribosomal complex (Rocak and Linder, 2004). In addition, in cold acclimation of mesophilic cells, the primary role of CsdA is in mRNA decay (Awano et al., 2007).
5. The Proposed Model of Life under Subzero Conditions: What We Know about the Components of the Model (E. coli cpn+, P. arcticus 273-4, and CsdA - Psyc_1082)
CsdA - Psyc_1082 is the main component of the model proposed in this study (Fig. 2). Therefore, the next sections contain (1) a review about the mesophilic microorganism to be tested in the model, (2) a review about the psychrophilic microorganism that expresses the protein CsdA - Psyc_1082 in vivo, and (3) in silico structural analyses of CsdA - Psyc_1082 protein by homology modeling.
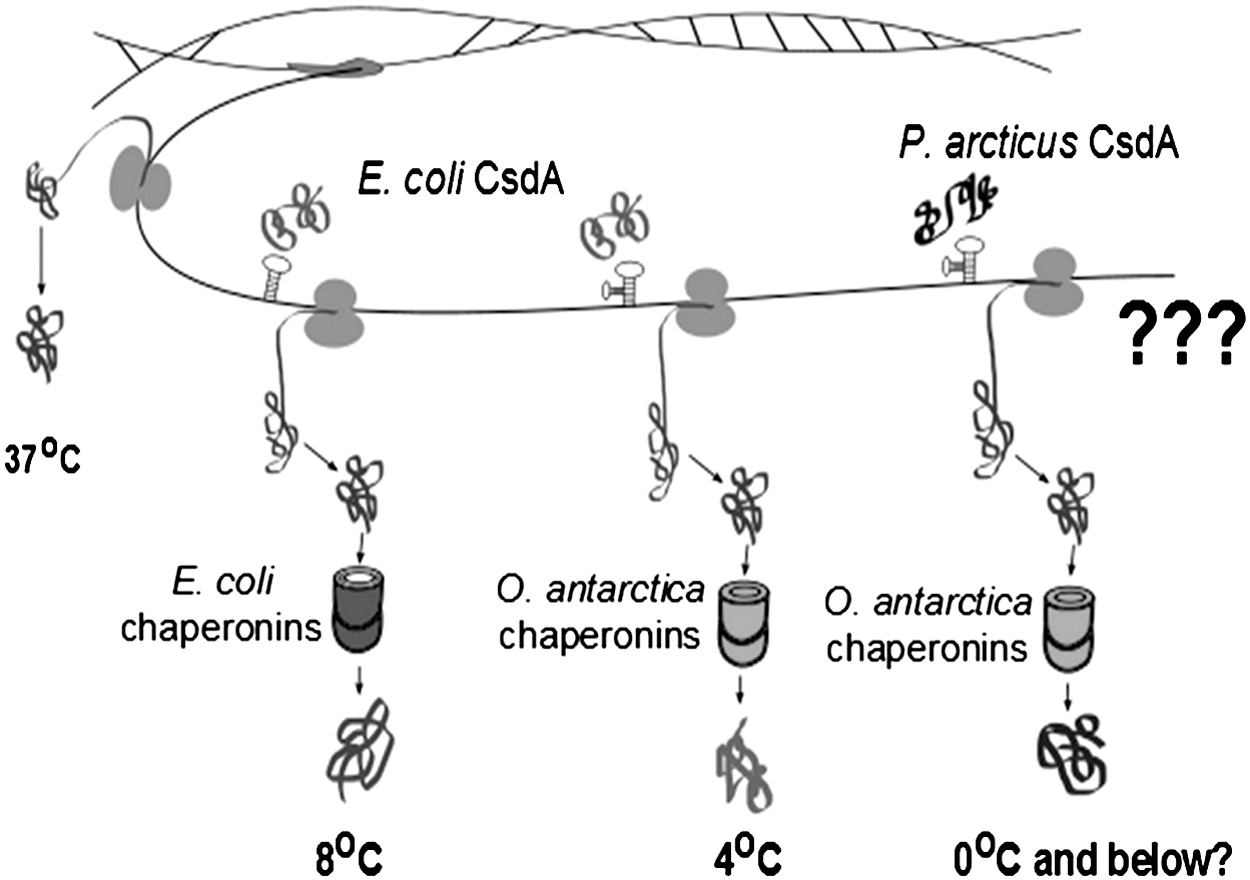
Schematic representation of the model proposed for the advance of psychrophilic protein expression in mesophiles and extension of the minimum growth temperature. At 37°C, mesophilic cells (in this case, E. coli) do not require CsdA, chaperonin, or related helper proteins for gene expression; at 8°C, in response to cold shock, mesophilic cells (E. coli) express CsdA and chaperonins to keep cell functionality; at 4°C, transformed E. coli cells expressing psychrophilic chaperonins from O. antarctica (cpn+ ) increased the expression level of 19 housekeeping proteins and were able to grow, lowering their limit of cell survival by 4°C. Could the transformed mesophile E. coli cpn+ be able to grow at temperatures at or below 0°C by expressing another set of helper proteins which work in unwinding and stabilizing RNA secondary structure? The expression modeling suggested by the “???” relies on the expression of P. arcticus 273-4 CsdA - Psyc_1082 by the transgenic E. coli cpn+ constructed by Strocchi et al. (2006) to test for resistance/tolerance to colder and subzero temperatures.
“Omics” analyses
The range of growth for any given parameter is determined by key cellular functions or structures that fail at the parameter limit. At a specific low temperature, housekeeping gene expression and membrane integrity fail in mesophilic cells. In wild-type E. coli cells grown at 15°C, it was found that at least 15 proteins were induced during the beginning of the lag phase to repair the cell from the damages caused by the drop in temperature and to recover the cellular physiology (Panoff et al., 1998). Chaperonins, proteins responsible for (re)folding of other proteins acting in the final process of protein expression, are some of the CSP produced in the beginning of lag phase, and they are a key determinant for E. coli growth at low temperatures (Ferrer et al., 2003).
Escherichia coli grows best between 21°C and 49°C, with slower growth below 21°C and no growth below 7.5°C (Strocchi et al., 2006). A transgenic E. coli cpn+ that received the chaperonin Cpn60 and the co-chaperonin Cpn10 from the Antarctic seawater psychrophilic bacterium Oleispira antarctica strain RB8 grew much faster than the wild type in a range of temperatures from 18°C to 8°C, and grew at 4°C (Strocchi et al., 2006). Cpn60 and Cpn10 from O. antarctica present high protein refolding activities in vitro from 4°C to 12°C. The addition of these genes to the expression system of a mesophilic bacterium, which otherwise would have stopped growing at the limit of 7.5°C, gave to the cells the capacity to grow at 4°C. The expression of these two genes in the mesophilic cell increased the expression level of 19 housekeeping proteins by helping to stabilize the housekeeping proteins against cold-mediated inactivation by increasing physical interactions (Strocchi et al., 2006).
Psychrobacter arcticus 273-4, on the other hand, evolved in a freezing environment, and its response to low temperatures is completely different from that of E. coli. Siberian permafrost imposes continuous stress on microbial cells, including temperatures of ∼−10°C, desiccation, decrease in nutrient availability, and low but continuous levels of gamma radiation from soil minerals (Gilichinsky, 2001). P. arcticus 273-4 overcame each of these extreme conditions and developed a cellular physiology to inhabit this “severe” environment. Furthermore, it is not the only psychrophile capable of growth at freezing temperatures. Bacterial metabolisms were detected at −15°C in Psychrobacter sp. and Arthrobacter sp. cells, isolated from Lake Vostok accretion ice, and at −10°C in different isolates from permafrost (Christner, 2002; Bakermans et al., 2003). Metabolic activity was also detected in a permafrost community at −20°C, with a cellular doubling time of 160 days (Rivkina et al., 2000).
Genomic analysis showed that between 38% and 84% of P. arcticus 273-4 amino acid sequences exhibit cold adaptation indicators like less hydrophobicity, reduction in proline content and acidic residues, or higher Arg/Lys ratio (Ayala-del-Río et al., 2010). Its transcriptome was the first transcriptome reported at temperatures below 0°C (Bergholz et al., 2009). Bergholz et al. (2009) analyzed and compared growth rate measurements and transcriptomes at −6°C, 0°C, 4°C, 17°C, and 22°C in acetate medium. Interestingly, P. arcticus at 0°C and −6°C downregulated peptidyl-prolyl cis-trans isomerases, trigger factor, and the major heat shock–associated chaperones. Only chaperones associated with iron-sulfur cluster biosynthesis, oxidative protein damage, and clpB chaperone homologues were upregulated (Bergholz et al., 2009). A correlation between transcriptome and proteome in which 20 proteins were used that had an increase in relative abundance during growth at −6°C identified proteins related to translation, amino acid synthesis, cell wall degradation, and other unknown functions (Bergholz et al., 2009).
Both mesophilic and psychrophilic microorganisms, E. coli and P. arcticus 273-4, respectively, harbor DEAD-box RNA helicase genes (Fig. 3). E. coli contains five DEAD-box helicase (csdA, rhlB, rhlE, dbpA, and srmB), and P. arcticus 273-4 contains two [csdA (Psyc_1082), and rhlB (Psyc_0943)] (Iost and Dreyfus, 2006; Bergholz et al., 2009). Because DEAD-box RNA helicase participates in many cellular processes such as processing, transport, and degradation of RNA or ribosome biogenesis, it is considered a multifunctional enzyme (Prud'homme-Généreux et al., 2004; Phadtare, 2011). In psychrophiles, CsdA is expressed as a housekeeping protein, or it can be considered a CAP. However, CsdA expression in mesophilic microorganisms occurs and is essential only for cold-shock response as a CSP (Jones et al., 1996; Panoff et al., 1998; Hébraud and Potier, 1999; Charollais et al., 2004; Srinivas and Ray, 2006). Experimental investigation found two possible activities of CsdA related to low-temperature adaptations: (1) mRNA and ribosome biogenesis (unwinding activity is important for both functions) and (2) mRNA decay, where its helicase activity is considered crucial for promoting degradation of mRNA stabilized at 15°C in E. coli (Phadtare, 2011). Research has also suggested that CsdA may help 50S assembly by modulating RNA structures (Iost and Dreyfus, 2006). Its unwinding activity may be required to facilitate structural transitions within the RNA and may also allow proper binding of r-proteins (Iost and Dreyfus, 2006). Finally, it is suggested that CsdA prevents and/or resolves rRNA misfolding caused by lowering the temperature, providing assistance to rRNA to reach its active conformation (Phadtare, 2011).
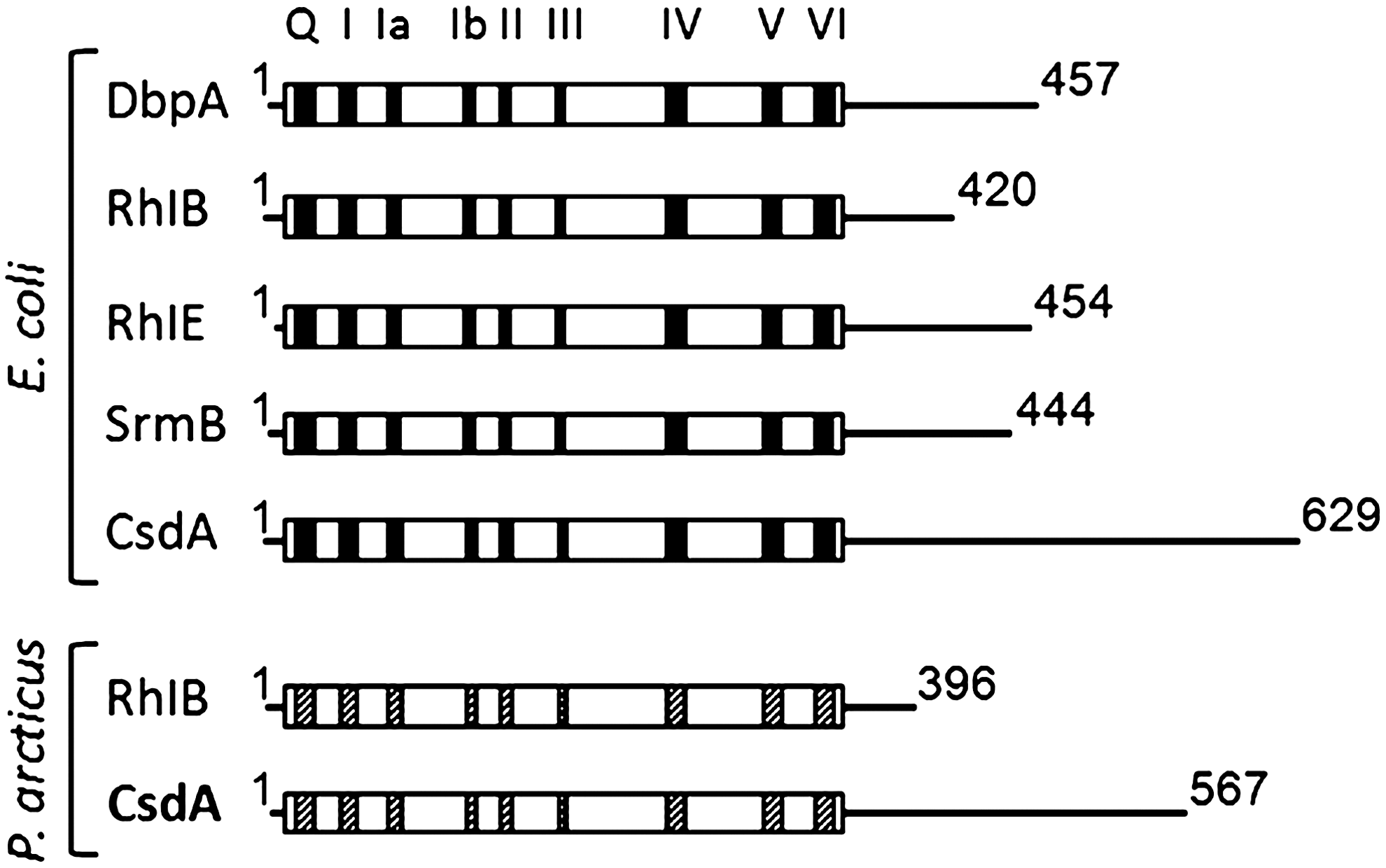
E. coli and P. arcticus 273-4 DEAD-box RNA helicases: conserved motives and sizes. An extension of ∼150 amino acids is observed in the C-terminal region of the P. arcticus CsdA protein. The nine conserved motives that make up the DEAD-box core are represented by black (E. coli) and gray (P. arcticus 273-4) boxes. The number in the C-terminal represents the total size of the proteins.
Mutant analyses
In E. coli, the deletion of the csdA gene leads to growth defects only at low temperatures, near 15°C (Awano et al., 2007). The deletion of the rhlB gene in the mesophilic cell does not cause any defect in growth at the optimum temperature of 37°C (Awano et al., 2007). However, in P. arcticus 273-4, the deletion of Psyc_1082 (csdA) resulted in decreased cellular growth rates above 4°C (Bergholz et al., 2009). P. arcticus 273-4 with the Psyc_0943 (rhlB) gene deleted did not result in growth at 17°C or 4°C, suggesting that this gene plays an essential role in P. arcticus 273-4 cell physiology at its optimum temperature (Bergholz et al., 2009). It has been shown in E. coli that the deletion of csdA leads to a severe deficit of free 50S subunits and accumulation of 40S particles that correspond to incomplete assembly of ribosomal large subunits (Charollais et al., 2004).
Structural analyses
Protein synthesis and folding are the crucial problems to overcome for life in cold and freezing environments; the secondary structure of RNA is stabilized via H bonds, making translation difficult. Substrate recognition and docking are also affected because the interaction between enzyme-substrate becomes weaker (electrostatic interactions are stabilized by a decrease in temperature, whereas the hydrophobic effect is weak) and ribosome activities slow down (Piette et al., 2011). RNA helicases are overexpressed at low temperatures in many psychrophilic microorganisms such as Methanococcoides burtonii (Lim et al., 2000), Exiguobacterium sibiricum (Rodrigues et al., 2008), Sphingopyxis alaskensis (Ting et al., 2010), and Pseudoalteromonas haloplanktis (Piette et al., 2010). The studies agree that these helicases may help unwind the RNA secondary structures and rearrange them for efficient translation in the cold; however, there have been no studies of structure or homology modeling in any of the psychrophilic helicases.
For enzymes, increased flexibility and decreased stability translate into greater entropy. The thermodynamic effect of cold adaptation is a reduction in the temperature dependence of the maximum catalytic rate (Feller and Gerday, 1997, 2003). This can be achieved through structural plasticity and decreased stability during the activation of an enzyme-substrate complex, resulting in a reduction of the activation enthalpy and an increase in the activation entropy of the reaction. Analyses of the amino acid sequences and structures of psychrophilic enzymes have given rise to the flexibility concept, that is, a psychrophilic enzyme can exhibit increased catalytic activity at low temperatures with limited loss of thermostability through adaptation for reduced numbers of stabilizing interactions between key amino acid residues (Grzymski et al., 2006). The genome of P. arcticus 273-4 shows a statistically significant shift in amino acids compared with mesophiles, to those known to favor flexibility at low temperatures for most cell functions, but particularly for those involved in growth and reproduction (Ayala-del-Río et al., 2010).
To explore the structure of protein Psyc_1082, a tertiary structure prediction was conducted by homology modeling with the program RaptorX (Peng and Xu, 2011). The secondary structure of Psyc_1082 was aligned against four distantly related DEAD-box RNA helicase proteins with tertiary structures characterized by X-ray diffraction of the protein crystal. The DEAD-box RNA helicases from an archaeon, Methanococcus jannaschii (1HV8), and three eukaryotes—Drosophila melanogaster (2DB3), Saccharomyces cerevisiae (3I62), and Homo sapiens (3EX7)—were extracted from and are available at the PDB.
The homology modeling shows that the Psyc_1082 monomer consists of two α/β domains, found in all known monomeric helicase structures (Fig. 4). An extension of ∼150 amino acids is observed in the C-terminal region of the protein (Figs. 3 and 4). DEAD-box terminal extensions are non-conserved regions that are thought to mediate the interaction of the protein with specific partners or RNA substrates, and it is where the protein specificity resides to act in different cellular processes (Silverman et al., 2003). The NEFF score of the model is 9.0. NEFF can be interpreted as the entropy of a sequence profile derived from multiple alignment, and its value ranges from 1 to 20. A small NEFF, such as 9.0, means low homology because of the lack of homologous information for the structure prediction in protein databases (Peng and Xu, 2010).

Structural analysis of CsdA - Psyc_1082. The tertiary structure was predicted by homology modeling comparing the secondary structure of CsdA - Psyc_1082 to distantly related and characterized DEAD-box RNA helicases from Methanococcus jannaschii (1HV8), Drosophila melanogaster (2DB3), Saccharomyces cerevisiae (3I62), and Homo sapiens (3EX7) as templates. The box indicates CsdA - Psyc_1082 disordered C-terminal extension rich in glycine, serine, and arginine residues.
Linding et al. (2003) presented a comparative study of the primary disorder structure of Psyc_1082 and Psyc_0943, using DisEMBL 1.5. The program predicted two coils in amino acid positions 272 to 336 of the Psyc_1082 sequence not detected in the Psyc_0943 sequence. Hot loop predictions revealed a larger difference in protein disorder over the aligned sequences, with the Psyc_1082 sequence containing a predicted 120 hot loop amino acid positions, compared with only 70 such positions in the Psyc_0943 sequence. The authors detected that P. arcticus 273-4 CsdA has a highly disordered C-terminal extension rich in glycine, serine, and arginine similar to that observed in the psychrophilic M. burtonii (Lim et al., 2000). In the present study, the highly disordered C-terminal extension was also identified in the Psyc_1082 tertiary structure (box, Fig. 4). The result suggests that the disordered C-terminal extension may contribute strongly to the enzyme activity in P. arcticus 273-4 cell system at freezing temperatures.
6. Does Psyc_1082 Have the Capacity to Confer Cellular Growth to Mesophiles at Freezing Temperatures?
If P. arcticus's csdA gene becomes expressed in a transgenic mesophile, such as E. coli that encodes the genes chaperonin Cpn60 and co-chaperonin Cpn10 (cpn+ ) from the psychrophile O. antarctica, will the CsdA protein confer cellular growth capacity of the mesophilic cell at subzero temperatures? (Expression modeling is represented in the right side of Fig. 2).
The model proposed for the extension of the minimum growth temperature presented here is based on experimental results developed through research on two microorganisms: E. coli and P. arcticus 273-4. Studies of E. coli cells that express psychrophilic chaperonins (Ferrer et al., 2003; Strocchi et al., 2006) are used as a model for gene expression and for testing the capacity of CsdA - Psyc_1082 protein in conferring cell adaptations to growth in lower temperatures. In addition, data and results published from experiments conducted on P. arcticus 273-4 to determine its genomic, proteomic, metabolic activity, gene expression, molecular, physiological, and morphological response to cold and subzero temperatures were analyzed to choose the protein to be modeled: Psyc_1082 (Ponder et al., 2008; Bakermans et al., 2009; Bergholz et al., 2009; Ayala-del-Río et al., 2010).
Whether the addition of the Psyc_1082 protein to the cells with the cpn+ genes will allow mesophilic cells to grow at lower or even freezing temperatures—a capacity that the E. coli cpn+ does not have, only growing above 4°C—remains to be tested (Fig. 2).
7. Final Considerations
It is known that cold-inducible DEAD-box RNA helicases are important for unwinding RNA secondary structures during a microorganism's growth at low temperatures, but no research has been carried out on these proteins for any psychrophilic organism. The development of protein crystal and/or protein structure analyses through computer modeling with a structural behavioral modeling prediction of the protein at low and subzero temperatures can provide key information to further investigate the mechanisms that underlie cold-induced biological systems in icy worlds. The protein in focus here is CsdA-Psyc_1082. However, Psyc_1082 is only one example of a psychrophilic CSP that needs to be better understood. Cold adaptations tend to be environment or organism (phylogeny) specific (Hinsa-Leasure et al., 2010); therefore, it is suggested here that studies be conducted on multiple psychrophilic CSP. These studies will allow us to understand adaptations to cold environments and provide insight into, and extend, the boundaries of what we know about life in extreme environments.
CsdA DEAD-box RNA helicase is known to be a key CSP expressed by microorganisms at cold temperatures. Therefore, understanding protein behavior in subzero conditions is essential to understanding the biochemical mechanism that could be used for long-term survival of microorganisms in cold and freezing environments. It has already been shown that the expression of two psychrophilic chaperonins in a mesophilic cell improves the capacity of the cell to grow at lower temperatures (Strocchi et al., 2006). The expression of a psychrophilic protein involved in transcript stability during translation could further improve the efficiency of growth at low temperatures and extend the minimum growth temperature of a mesophilic cell such as an E. coli strain (Fig. 2). Testing mesophilic cells that express key psychrophilic proteins for resistance/tolerance to colder and subzero temperatures is important because it improves our understanding of how psychrophilic cells exist in their extreme environments. Studying and modeling psychrophilic “cold-shock” proteins will provide important new insights into biological sensitivity and tolerance to cold. This can help us gain better comprehension of biochemical mechanisms of the key protein workers that sustain life in subzero conditions, increase our knowledge about habitability in cold environments, and assist efforts for detection of life on Mars, icy moons, and elsewhere in our solar system.
Footnotes
Acknowledgments
This research and E.K. were supported by Fulbright/USA–CAPES/Brazil (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil). Special thanks to Kristien C. King and Christian Klimczak for reviewing the manuscript and providing extremely valuable feedback.
Abbreviations
CAP, cold-acclimation proteins; CSP, cold-shock proteins; PDB, Protein Data Bank.