
Abstract
Earth-based microgravity simulation techniques are required due to space research constraints. Using diamagnetic levitation, we exposed Arabidopsis thaliana in vitro callus cultures to environments with different levels of effective gravity and magnetic field strengths (B) simultaneously. The environments included simulated 0g* at B=10.1 T, an internal 1g* control (B=16.5 T), and hypergravity (2g* at B=10.1 T). Furthermore, samples were also exposed to altered gravity environments that were created with mechanical devices, such as the Random Positioning Machine (simulated μg) and the Large Diameter Centrifuge (2g). We have determined the proteomic signature of cell cultures exposed to these altered-gravity environments by means of the difference gel electrophoresis (DiGE) technique, and we have compared the results with microarray-based transcriptomes from the same samples. The magnetic field itself produced a low number of proteomic alterations, but the combination of gravitational alteration and magnetic field exposure produced synergistic effects on the proteome of plants (the number of significant changes is 3–7 times greater). Tandem mass spectrometry identification of 19 overlapping spots in the different conditions corroborates a major role of abiotic stress and secondary metabolism proteins in the molecular adaptation of plants to unusual environments, including microgravity. Key Words: DiGE—Microgravity simulation—Magnetic levitation—Proteome/transcriptome comparison—Callus cell cultures. Astrobiology 13, 217–224.
We have studied the effect of an exposure of 200 min to an environment of altered gravitational and magnetic forces on the overall proteomic profile of Arabidopsis thaliana semisolid cell cultures (callus). The present study is the first systematic multifacility, high-throughput, environmentally controlled collection of experiments that has been performed with the same setup, almost simultaneously in two mechanical facilities (RPM for simulated μg and LDC for 2g) and in a magnet-based facility, with the use of three different effective gravity (g*) conditions [2g*, 1g*, and 0g*, namely, the calculated effective gravity for pure water in the center of each culture chamber, assuming that variations due to differential magnetic susceptibly of intracellular material are below biological sensitive (Schenck 1992; Valles et al., 1997); see also Herranz et al. (2013) for a discussion about terminology]. This made possible inter-experiment comparisons of the results and a pooled analysis with multiple inner controls at similar magnetic fields. Apart from testing what kind of facility would be most suitable and reliable as an altered gravity simulator, a major achievement of this study was the comparison of the proteomic profiles with the transcriptomic results obtained from the same samples [Agilent two-color 44k whole genome microarray data sets, GEO ID: GSE29787 (Manzano et al., 2012)].
A detailed description of the experimental setup, the levitation magnet (Fig. 1A), and the mechanical simulators (Fig. 1B) was provided in a parallel transcriptomic paper (Manzano et al., 2012). In short, we exposed samples to three different conditions (g* and B fields) within a high field magnet (HFML, Radboud University Nijmegen, the Netherlands) (Perenboom et al., 2004; Wiegers et al., 2010). The magnitude g* denotes the value of the effective gravity (the asterisk reflects the presence of the background magnetic field) in each of the three positions within the magnet (Table 1). In the center of the magnet bore (1g* position), the magnetic field strength is maximal (16.5 T), but the magnetic field gradient is zero. At a distance of 81.6 mm above this position, at B=10.1 T, the diamagnetic force on water counterbalances the force of gravity, which leads to stable levitation; this is the 0g* position (zero gravity is only reached in a single point). At a distance of 81.6 mm below the field center, also at B=10.1 T, the sum of the levitation and gravitational forces produces a 2g* effective force. In both cases, we consider that the effective gravity applied on Arabidopsis culture cells (free of starch statoliths) is similar to the one of water. We base this assumption on the fact that cells' main component is water, and most organic compounds within the cell have similar diamagnetic properties as those of water (Schenck 1992; Valles et al., 1997). Additionally, we exposed samples to a mechanical simulator of microgravity, the Random Positioning Machine (μg-RPM, real random mode), and to the Large Diameter Centrifuge (2g-LDC) for mechanical generation of hypergravity, both facilities located in the European Space Research and Technology Centre [ESA-ESTEC, Noordwijk, the Netherlands, Fig. 1B (van Loon, 2007; van Loon et al., 2008)]. In each facility, samples were exposed for 200 min, at 22°C±0.1°C, in darkness. External controls were kept at room temperature (22°C±0.1°C, 1g ground gravity), away from the magnet, LDC, or RPM. As a model system, we used callus semisolid cultures of Arabidopsis thaliana prepared from MM2d suspension cultures (Menges and Murray, 2006). For all devices and conditions, the suspension cultures were strewn on the agar surface 1 week before the start of the experiment and grown at 22°C to allow the callus to reach maximum density in a 1–2 mm thick layer to minimize variations in the magnetic field and of the effective g-force. The samples were preserved immediately after treatment (less than 2 min) by quick freezing in liquid nitrogen and subsequent storage, first in dry ice for transport and then under deep freezing.
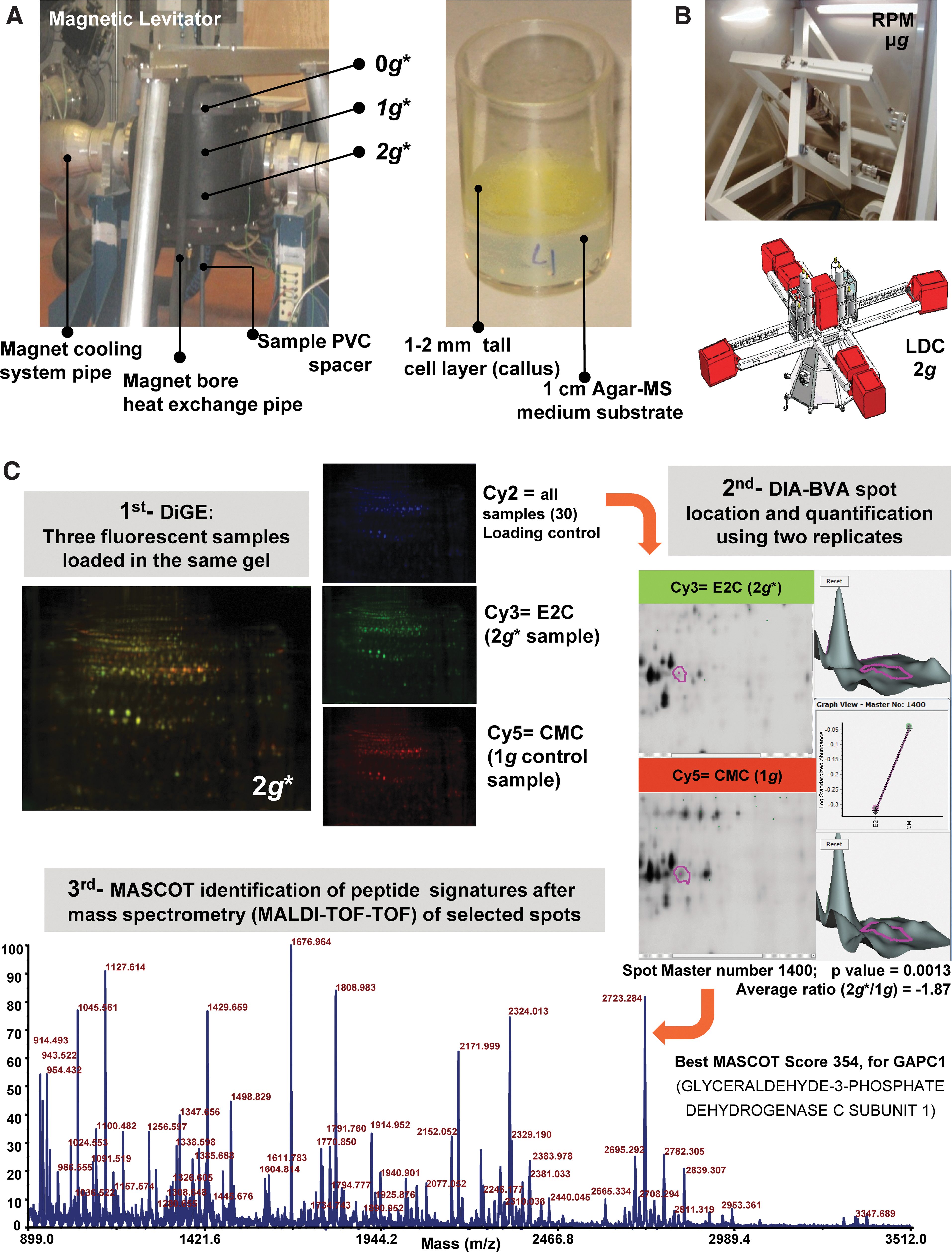
Altered-gravity ground-based facilities and experimental approach. (
— indicated that magnetic/mechanical perturbations are minimal and similar to the control for those samples. Number of significantly altered spots was determined by using a Student t test (p<0.05) with two replicates. The numbers obtained, including a more variable third replicate, are shown between brackets for comparison.
Samples were analyzed as to their proteomic profile as summarized in the example in Fig. 1C by the two-dimensional difference gel electrophoresis (2-D-DiGE) technique. Protein was extracted from three biological replicates as described by Barjaktarovic et al. (2007) and purified by methanol/chloroform precipitation. An amount of 40 μg of control or experimental protein samples was stained with 400 pmol of Cy3 or Cy5 (CyDryes manufacturer, GE Healthcare). Standard control for normalization between gels consisted of a pool of the samples at the same concentration stained with Cy2. DiGE first dimension of electrophoresis was performed in an IPGphor IEF System with a linear 3–11 pH (GE Healthcare). DiGE second dimension was performed on an Ettan Dalt Six by using 12.5% SDS-PAGE at 20°C. Gels were scanned at 100 μm resolution in a Typhoon 9400 scanner (GE Healthcare). Data analysis was performed by using two or three replicates with DeCyder software v7.0, Differential In-Gel Analysis (DIA) and Biological Variation Analysis (BVA) for which, as filter, a fold change≥1.2 or≤−1.2 was used with a Student t test to extract significant results between replicates (p<0.05).
The results show that a 200 min treatment in mechanical facilities did not produce any significant effect at the proteomic scale (Table 1). In fact, we were able to detect one spot significantly altered in the RPM and two in the LDC only when removing the most variable replicate, which suggests that we were only detecting highly significant variations. The effect of a high magnetic field is moderate, producing more decreased than increased spots; a clearly stronger effect is observed in the 0g* and 2g* positions, especially detected in the 2g* position. These effects are quite similar to the synergic effect found at the transcriptomic level with the same samples (Manzano et al., 2012), reinforcing the idea of an enhanced impact of altered gravity on biological systems under suboptimal environments (Herranz et al., 2010, 2012).
To analyze more deeply the proteomic response, we decided to identify the more representative altered spots in several pair comparisons. We chose 19 spots (Table 2), namely, (a) those altered in the RPM/LDC samples, (b) two spots especially easy to isolate with high fold changes in the 0g* position, (c) those spots appearing simultaneously altered in any g* level after comparing its proteomic profile with both the internal (1g*) and the external (1g) controls, (d) two common spots of the comparisons of 0g* and 2g* with 1g, and (e) two outstanding spots revealed by the comparison of 0g* with the external 1g control. Once the spots of interest were identified and located, trypsin-digested spots were analyzed by a 4800 MALDI-TOF-TOF facility (Maccarrone et al., 2010). MASCOT searches with nrNCBI protein database (viridiplantae only) were used to identify the candidate proteins that better fit with the mass spectrum obtained for each spot tryptic peptides, up to eight fragmentation peptides analyzed per spectrum (score higher than 72 to have a p<0.05).
Spots appearing in more than one gravitational condition (comparison based on master and spot numbers) are shown, including statistical significance (t test) and average fold change values (in Average ratio, in italics when reduced, in bold when increased in concentration). Mascot values are also shown for the significant candidates only (it is indicated by dashed lines when more than one candidate is found for the same spot). # highlights candidates identified in another spot. See Supplementary Material for additional method details.
Candidate proteins that appeared during the analysis were easily identified as involved in cellular response to stress conditions, like chaperonin (4.91-fold increase in 2g*), β-glucosidase-15 (almost 10-fold increase in 2g*), and Hsp70 (slightly but significantly decreased in both 0g* and 2g*), together with ATP synthase VHA-A and oxidoreductase Sks5, which appeared in two different spots. Primary and secondary metabolism enzymes usually appear to be affected by environmental stress conditions (Liu et al., 2011), and some of them were also found to be modified in our study (Table 2). Previously performed proteomic analyses with the use of 2-D electrophoresis cross comparisons identified similar types of proteins that were affected by mechanically altered gravity conditions, among which fructose-bisphosphate aldolase in particular was mentioned (Wang et al., 2006; Barjaktarovic et al., 2007, 2009). In our work, in which spots were quantified in the same gel (DiGE), only a few variations were found to be significant in the mechanical facilities, whereas the synergic action of two stressing conditions (magnetic field and altered gravity) revealed proteomic changes similar to those reported in the literature.
Finally, we compared these specific proteins with the genes we previously found to be affected with the use of an Agilent array on the same samples. Most of the identified matches affected similarly both hypergravity environments (2g* in the magnet and LDC) (Table 3). It must be kept in mind that the decrease of one spot could mean that the affected protein has been postranslationally modified (transformed into a new spot), becoming even more active; this would explain apparent inconsistencies between proteomic and transcriptomic data sets. In fact, we found additional partial matches [e.g., protein GAPC-1, used as example in Fig. 1C, match gene is not affected; but a similar gene, GAPA-2 (AT1G12900), is] that would cross-validate both proteomic and transcriptomic data sets.
Changes in gene expression or protein levels are indicated in bold (increase) or italics (decrease).
Summarizing, we conclude that a 200 min exposure to mechanically altered gravity produces very few alterations in our environmentally controlled system in comparison with other studies in which Arabidopsis callus was exposed to clinorotation or centrifugation with other designs (Wang et al., 2006; Barjaktarovic et al., 2007, 2009). This result can be explained by the different sensitivities of other proteomic approaches (DiGE technology produces more statistically reliable results than independent 2-D gels but provides less number of spots detected) and also by the different duration of the treatment and source of the biological material (for instance, when using seedlings, cellular plus tissular proteomic responses will be observed). Nevertheless, our results are consistent with those of previous studies, which reinforces the idea that the alteration of gravity, together with other physical forces, promotes an abiotic stress response that provides the cells some adaptation benefits in the context of a new environmental situation at genomic and proteomic levels (Barjaktarovic et al., 2007, 2009; Manzano et al., 2012). In fact, several plant species have the capacity to cope with extreme environmental conditions by modifying secondary metabolism, stress, and repair pathways in a concerted way (Van Cutsem et al., 2011; Forster et al., 2012; Payyavula et al., 2012).
Using magnetic forces seems to increase the system susceptibility to altered gravity and affects similar proteins as those that have been described in other studies in which mechanical simulators were used. The same gene ontology functions have been detected by transcriptomic analyses (Manzano et al., 2012). Similar enhanced effects of microgravity on the transcriptomic profile have been observed before when suboptimal environmental conditions were added to the altered gravity stimulus (Herranz et al., 2010, 2012; Manzano et al., 2012). It is possible that the observed effects at the 0g* and 2g* positions could be related to forces generated by differences in magnetic susceptibility between different components of the cell. Further experiments are required to assess this possibility.
Investigation of “pure” microgravity effects should be performed in space, but mechanical and magnetic simulators could be used to study similar phenomena if we are able to distinguish the mechanical/magnetic effects from the gravitational effects in our systems. In addition, magnetic levitation can be an alternative to other ground-based methodologies and allow us to test the biological effects of altered gravitational forces in an unusual environment.
Supplementary Material
All raw and identification data is available as Supplementary Material (available online at
Footnotes
Acknowledgments
The authors acknowledge funding from the Spanish Space Program in the “Plan Nacional de Investigacion Cientifica y Desarrollo Tecnologico” AYA2009-07952, AYA2010-11834-E to F.J.M. and AYA2009-07792-E to R.H. RPM/LDC access was possible due to the Dutch Space Research Organization NWO-ALW-SRON grant MG-057 to J.vL. and the SEGMGSPE_Ph1 access to ESA GBF project contract 4200022650 to R.H., F.J.M., and J.vL. Magnetic levitation at the High Field Magnet Laboratory in Nijmegen was granted by EuroMagNET II under the EU contract n° 228043 and by the Stichting voor Fundamenteel Onderzoek der Materie (FOM), financially supported by the Nederlandse Organisatie voor Wetenschappelijk Onderzoek (NWO). PRIDE files have been generated in the CSIC CNB computational proteomics unit from CSIC & ProteoRed: Carlos III Networked Proteomics Platform. We want to acknowledge also Dr. Mateos and two anonymous referees for their critical review of the manuscript.
Author Disclosure Statement
The authors have declared no conflict of interest.
Abbreviations
BVA, Biological Variation Analysis; DIA, Differential In-Gel Analysis; DiGE, difference gel electrophoresis; LDC, Large Diameter Centrifuge; RPM, Random Positioning Machine.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.