
Abstract
The space environment contains high-energy charged particles (e.g., protons, neutrons, electrons, α-particles, heavy ions) emitted by the Sun and galactic sources or trapped in the radiation belts. Protons constitute the majority (87%) of high-energy charged particles. Spores of Bacillus species are one of the model systems used for astro- and radiobiological studies. In this study, spores of different Bacillus subtilis strains were used to study the effects of high energetic proton irradiation on spore survival. Spores of the wild-type B. subtilis strain [mutants deficient in the homologous recombination (HR) and non-homologous end joining (NHEJ) DNA repair pathways and mutants deficient in various spore structural components such as dipicolinic acid (DPA), α/β-type small, acid-soluble spore protein (SASP) formation, spore coats, pigmentation, or spore core water content] were irradiated as air-dried multilayers on spacecraft-qualified aluminum coupons with 218 MeV protons [with a linear energy transfer (LET) of 0.4 keV/μm] to various final doses up to 2500 Gy. Spores deficient in NHEJ- and HR-mediated DNA repair were significantly more sensitive to proton radiation than wild-type spores, indicating that both HR and NHEJ DNA repair pathways are needed for spore survival. Spores lacking DPA, α/β-type SASP, or with increased core water content were also significantly more sensitive to proton radiation, whereas the resistance of spores lacking pigmentation or spore coats was essentially identical to that of the wild-type spores. Our results indicate that α/β-type SASP, core water content, and DPA play an important role in spore resistance to high-energy proton irradiation, suggesting their essential function as radioprotectants of the spore interior. Key Words: Bacillus—Spores—DNA repair—Protection—High-energy proton radiation. Astrobiology 12, 1069–1077.
1. Introduction
A
Spores of Bacillus species have been used extensively as biological dosimeters for probing terrestrial and extraterrestrial radiation in outer space or in space simulation facilities (reviewed in Fajardo-Cavazos et al., 2007; Horneck, 1993; Nicholson et al., 2000, 2005, 2009; Horneck et al., 2001, 2010). In contrast to their vegetative state as metabolic active cells, spores of Bacillus species are highly resistant to inactivation by environmental physical stresses such as desiccation, pressure and temperature extremes, and high doses of UV and ionizing radiation (reviewed in Nicholson et al., 2000, 2005; Setlow, 2006, 2007). Historically, several studies have explored the resistance of bacterial spores to ionizing radiation (reviewed in Schmidt, 1955 and references therein); these early studies concentrated mainly on spore survival of wild-type strains of various spore-forming species. More recent experiments have attempted to better understand the underlying molecular factors that lead to spore radiation resistance by use of the model organism Bacillus subtilis, which carries mutations that affect spore structural components, spore properties, or spore DNA repair systems. Use of such mutant spores has resulted in a great increase in our understanding of the mechanisms of spore resistance to both UV radiation (Moeller et al., 2005, 2009, 2011a; reviewed in Nicholson et al., 2000, 2005; Setlow, 2006) and more recently spore resistance to X-rays and high-energy (HZE) particle bombardment (Moeller et al., 2008, 2010, 2011b).
Since sporadic SPE as well as the continuous exposure to GCR, with highly abundant protons of a few hundreds of mega electron volts, are the major hazards that constrain astronaut activities in space, there is a need to reach a better understanding of the effects of proton radiation on biological samples for radiation biological and space medical purposes (Chatterjee and Borak, 2001; Nelson, 2003; Cucinotta and Durante, 2006); however, the cellular factors responsible for the resistance of bacterial spores to high-energy proton radiation have been poorly investigated in detail. Therefore, in this communication we have examined the possible roles in spore resistance to highly energetic proton radiation (218 MeV) offered by various spore structural components, as well as the roles of the non-homologous end joining (NHEJ) and the homologous recombination (HR) DNA repair pathways.
2. Materials and Methods
2.1. Description of B. subtilis strains used
Bacillus subtilis strains used are listed in Table 1. Laboratory strain 168 (WN131) served as the wild-type strain. All mutant strains are congenic with strain WN131. Strain WN661 is a spontaneous mutant of wild-type strain WN131 that produces spores lacking brown pigmentation in their coats, presumably due to a mutation in the cotA-encoded copper-dependent laccase (Hullo et al., 2001). Strain WN469, which carries mutations in the cotE and gerE genes and results in spores lacking spore coat layers (Riesenman and Nicholson, 2000), was a generous gift from Adam Driks. The following strains were generous gifts from Peter Setlow: (1) strain WN553, which produces spores lacking dipicolinic acid (DPA) (Paidhungat et al., 2000); (2) strain WN1273, which lacks α/β-type small, acid-soluble spore proteins (SASP) (Mason and Setlow, 1986); and (3) strain WN1274 with increased spore core water content (Popham et al., 1995). Strain WN463 is deficient in the RecA-mediated HR DNA repair pathway; strain WN1087 is deficient in the NHEJ pathway; strain WN1141 lacks both HR and NHEJ pathways (Moeller et al., 2011a).
CmR, chloramphenicol (5 μg/mL), ErmR, erythromycin (1 μg/mL), SpcR, spectinomycin (100 μg/mL), TetR, tetracycline (10 μg/mL).
2.2. Production and purification of spores
Spores were obtained by cultivation under vigorous aeration in double-strength liquid Schaeffer sporulation medium (Schaeffer et al., 1965), purified by lysozyme treatment and buffer washing, and stored as described previously (Nicholson and Setlow, 1990). Where appropriate, chloramphenicol (Cm, 5 μg/mL), erythromycin (Erm, 1 μg/mL), spectinomycin (Spec, 100 μg/mL), or tetracycline (Tet, 10 μg/mL) were added to the medium. Spore preparations were free (>98%) of growing cells, germinated spores, and cell debris, as determined by phase-contrast microscopy.
2.3. Sample preparation
Spore suspensions of the different B. subtilis strains were prepared in sterile distilled water to a final concentration of 1×1010 spores/mL. Spacecraft-qualified, chemfilm-treated aluminum 6061 coupons (13 mm in diameter×1 mm thick; Moeller et al., 2011a) were produced (Titusville Tool & Engineering, Titusville, FL) and autoclaved (121°C, 30 min) prior to use. Samples for proton irradiation were prepared by applying 50 μL aliquots of spore suspensions (5×108 spores) onto 13 mm in diameter aluminum coupons so that they spread homogenously on the surface, yielding multilayer samples with a thickness of ∼25 spore layers (Moeller et al., 2011a). In our study, spacecraft-qualified aluminum coupons were chosen to simulate surface materials of a spore-contaminated spacecraft. The non-coverage of the spore load was calculated for the 5×108 spores/aluminum coupon and resulted in a non-coverage rate (i.e., fraction of unshielded spores) of ≤1%. One set of spore samples consisted of three identically prepared coupons. Spore samples were air-dried under ambient laboratory conditions (20°C, 33±5% relative humidity).
2.4. Spore exposure to 218 MeV high-energy protons
Proton irradiations were performed at the University of Florida Proton Therapy Institute (
2.5. Spore recovery and survival assay
Air-dried spore layers were removed from coupons with a polyvinyl alcohol stripping method as described previously (Lindberg and Horneck, 1991). Spores were released from the polyvinyl alcohol stripping film and resuspended in 1 mL sterile distilled water, resulting in >95% recovery of the spores. This procedure does not affect spore viability (Lindberg and Horneck, 1991). Spore survival was determined from appropriate dilutions in distilled water as colony-forming ability after incubation overnight at 37°C on nutrient broth agar plates (Difco, Detroit, USA). To control for contamination, genetic marker tests were performed on chemically defined agar media for the respective amino acid auxotrophy (i.e., tryptophan) or antibiotic resistance (i.e., Cm, Erm, Spec, or Tet).
2.6. Numerical and statistical analysis
The spore surviving fraction was determined from the quotient N/N 0, with N=the number of colony-forming units of the irradiated sample and N 0 that of the non-irradiated controls. To determine the curve parameters, the following relationship was used: ln (N/N 0)=−k i×D+n, with k i=spore inactivation constant (Gy−1), D=dose (Gy) and n=extrapolation number, that is, the intercept with the ordinate of the extrapolated semi-log straight line. The constants k i and n were determined by linear regression. Spore survival was plotted as a function of proton irradiation dose. Data are reported as D 10 values, that is, the dose resulting in a spore inactivation of one order of magnitude, and D 37 values, that is, the dose lethal for 63% of the initial spore population, determined from the linear portion of the semi-logarithmic curve (i.e., the reciprocal of the spore inactivation constant k i; according to Moeller et al., 2008, 2010, 2011b). Each experiment was repeated in triplicate, and the data shown are expressed as averages±standard deviations. The results were compared statistically with a Student t test. Values were analyzed in multigroup pairwise combinations, and differences with P values of ≤0.05 were considered statistically significant.
3. Results
To study the impact on spore viability of high-energy proton radiation, spores of different genotypes of the laboratory model organism B. subtilis were exposed as air-dried multilayers on spacecraft-qualified aluminum coupons to 218 MeV proton radiation. After spore removal from the aluminum coupons, inactivation kinetics of wild-type and mutant B. subtilis spores were determined in response to proton radiation (Figs. 1 and 2). Essentially, first-order exponential survival curves were obtained for spores of all strains tested, and the resulting best-fit curves were used to calculate D 10 values (i.e., proton dose reducing survival to 10%), D 37 values (i.e., proton dose reducing survival to 37%), and spore inactivation constants for statistical comparison (Table 2).
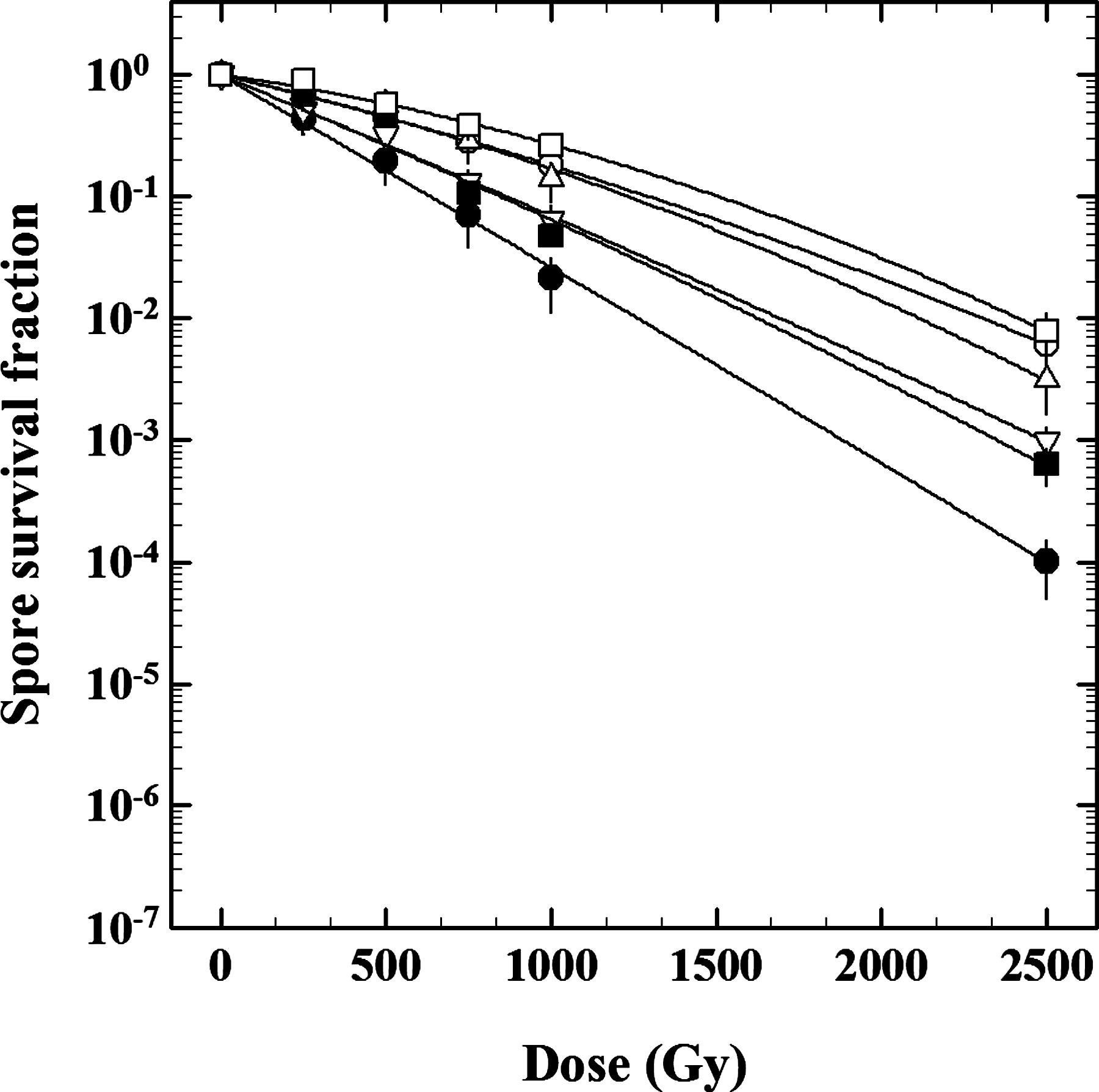
Survival curves of B. subtilis spores carrying various structural mutations exposed to 218 MeV protons. Spores from the following strains were tested: wild-type strain WN131 (open circles), spore coat–deficient strain WN469 (open squares), pigmentation-deficient strain WN661 (upward open triangles), strain WN1274 with increased spore core water content (downward open triangles), DPA-deficient (filled squares), and α/β-type SASP-deficient (filled circles). Data are averages±standard deviation (n=3). Error bars for survival data not shown were smaller than the plot symbol.
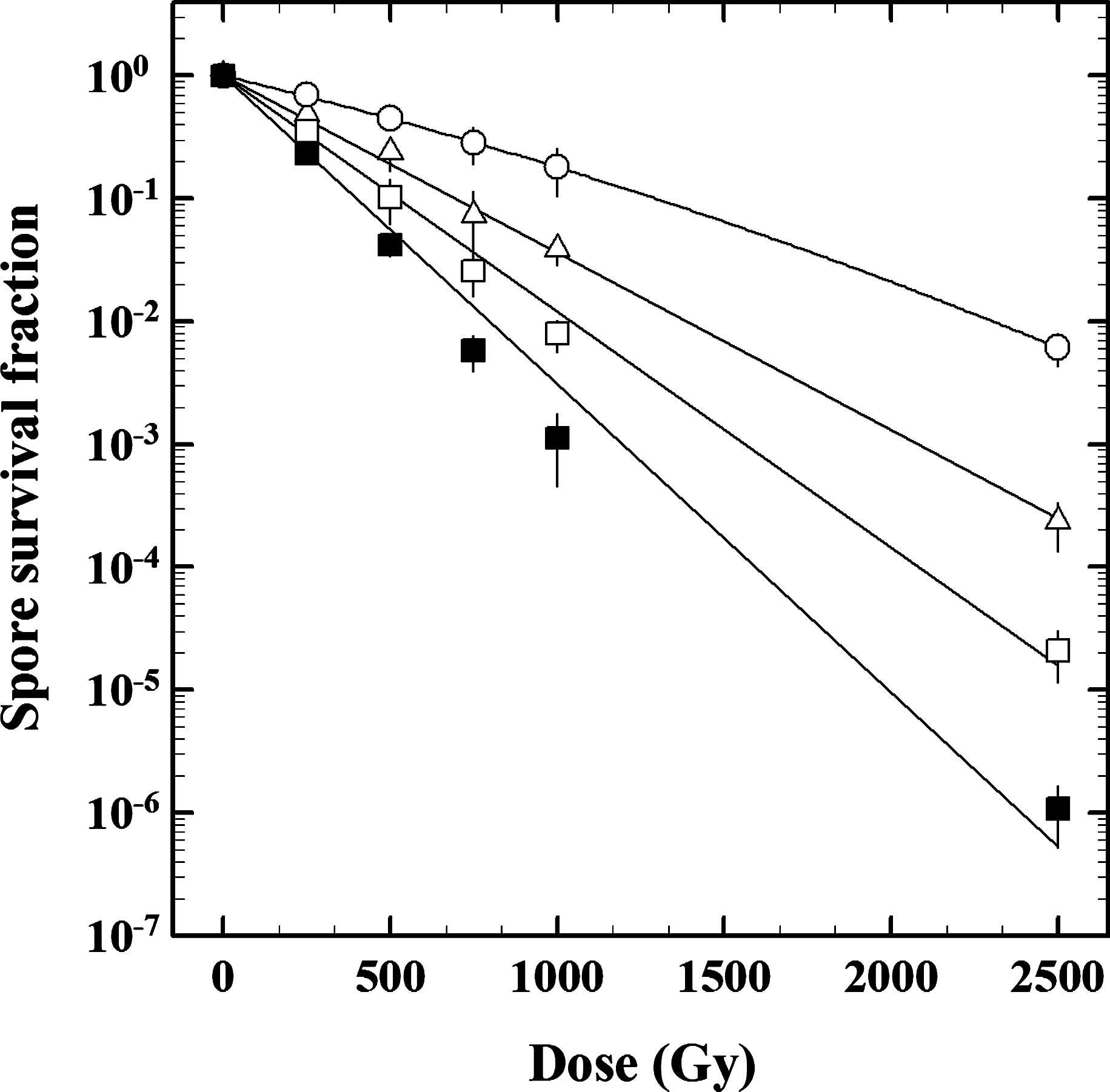
Survival curves of B. subtilis spores deficient in DNA repair exposed to 218 MeV protons. Spores from the following strains were tested: wild-type strain WN131 (open circles), HR-deficient strain WN463 (open triangles), NHEJ-deficient strain WN1087 (open squares), and strain WN1141 deficient in both HR and NHEJ (filled squares). Data are expressed as averages and standard deviations (n=3). Error bars for survival data not shown were smaller than the plot symbol.
Data are expressed as averages and standard deviations (n=3). Asterisks indicate D 10, D 37, and IC values that were significantly different (P values of ≤0.05) from the respective values of the B. subtilis wild-type spores.
D 10=dose (Gy) resulting in a spore inactivation of 1 order of magnitude.
D 37=dose (Gy) resulting in 63% spore inactivation.
IC=spore inactivation constant (Gy−1).
3.1. Protective effect of spore structural components
Supramacromolecular structures unique to the spore have been demonstrated to protect spores from inactivation after exposure to a host of environmental extremes (reviewed in Nicholson et al., 2000, 2005; Setlow, 2006, 2007). Spores lacking inner and outer spore coat layers (strain WN469; D 10=1281±76 Gy) and spores lacking brown coat pigmentation (strain WN661; D 10=1065±48 Gy) were inactivated by proton irradiation with kinetics statistically identical to the wild-type strain WN131 (1128±65 Gy) (Fig. 1). In contrast, spores with increased core water content (strain WN1274; D 10=791±45 Gy), spores lacking DPA (strain WN553; D 10=742±41 Gy), and spores deficient in α/β-type SASP (strain WN1273; D 10=607±38 Gy) were significantly more sensitive to proton radiation than the wild-type strain (Table 2). These findings indicated that lethal damage to the spore caused by highly energetic proton radiation occurs mainly interior to the spore coat and that α/β-type SASP, DPA, and spore core dehydration are important determinants of spore resistance to proton radiation exposure.
3.2. Role of HR and NHEJ DNA repair pathways in spore proton resistance
Ionizing radiation is known to cause damage to numerous targets within the cell, including DNA (Goodhead, 1994), due both to (i) direct interaction of the proton with the target molecule and (ii) indirect effects due to the production of reactive species such as oxygen radicals (reviewed in Friedberg et al., 2006). Ionizing radiation induces a large variety of damage to DNA bases, and through interaction with the sugar moiety it can cause formation of DNA strand breaks, either single-strand breaks (SSB) or double-strand breaks (DSB) (Friedberg et al., 2006). We examined the contribution of the two major DNA strand break repair mechanisms, HR (which operates both on SSB and DSB) and NHEJ (which is DSB-specific), in spore resistance to proton radiation (Fig. 2). When exposed to 218 MeV protons, spores of all strains carrying DNA repair deficiencies in HR, NHEJ, or both systems were significantly more sensitive to proton bombardment than the wild-type strain (Table 2). Spores exhibited a hierarchy of resistances (D 10 level), in the following order: wild-type strain WN131 (1128±65 Gy)>HR single mutant strain WN463 (688±38 Gy)>NHEJ single mutant strain WN1087 (477±31 Gy)>HR+NHEJ double mutant strain WN1141 (321±26 Gy) (Table 2). The data suggest that after exposure to 218 MeV protons (i) NHEJ contributes more than does HR to spore resistance, when compared singly; and (ii) together HR and NHEJ appear to contribute additively to spore resistance.
3.3. Relative effectiveness of protons versus X-rays and HZE particles
To determine the sporicidal effectiveness of proton radiation relative to X-rays and heavy ion (HZE) particles, their relative biological effectiveness (RBE) was calculated as the ratio of the spore inactivation constant of protons to the spore inactivation constant of X-rays and five selected heavy ion species (Fig. 3). After exposure to the same doses of proton radiation, it was observed that the wild-type spore survival was slightly higher (but not a statistically significant difference) than the spore survival after X-ray irradiation. Spores of representative proton-sensitive strains WN463 (HR deficient), WN1273 (α/β-SASP deficient), and WN1141 (HR+NHEJ deficient) exhibited shallower slopes than the wild-type spores (Fig. 3), indicating the general sensitivity of these strains to the DNA-damaging effects of protons, X-rays, and HZE particles with a weaker dependency of the LET as the wild-type spores. Here, it should be kept in mind that both low and high LET radiation produce a wide range of damage types (SSB, DSB) but also clustered DNA damage, a variety of different lesions, for example, strand breaks, abasic sites, or oxidized bases in short regions (i.e., one or two helical turns of the DNA); however, not all damage types have the same biological significance (Hutchinson, 1985; Ward, 1985; Goodhead, 1994, 1999; Asaithamby and Chen, 2011). Furthermore, it is known that clustered DNA damage in local regions of the DNA are known to act as critical lesions for increased mutagenicity and high lethality (Sutherland et al., 2000; Lomax et al., 2002; Asaithamby and Chen, 2011), which could be an explanation for the similar RBE ratios of the spores from the proton-radiation–sensitive strains.
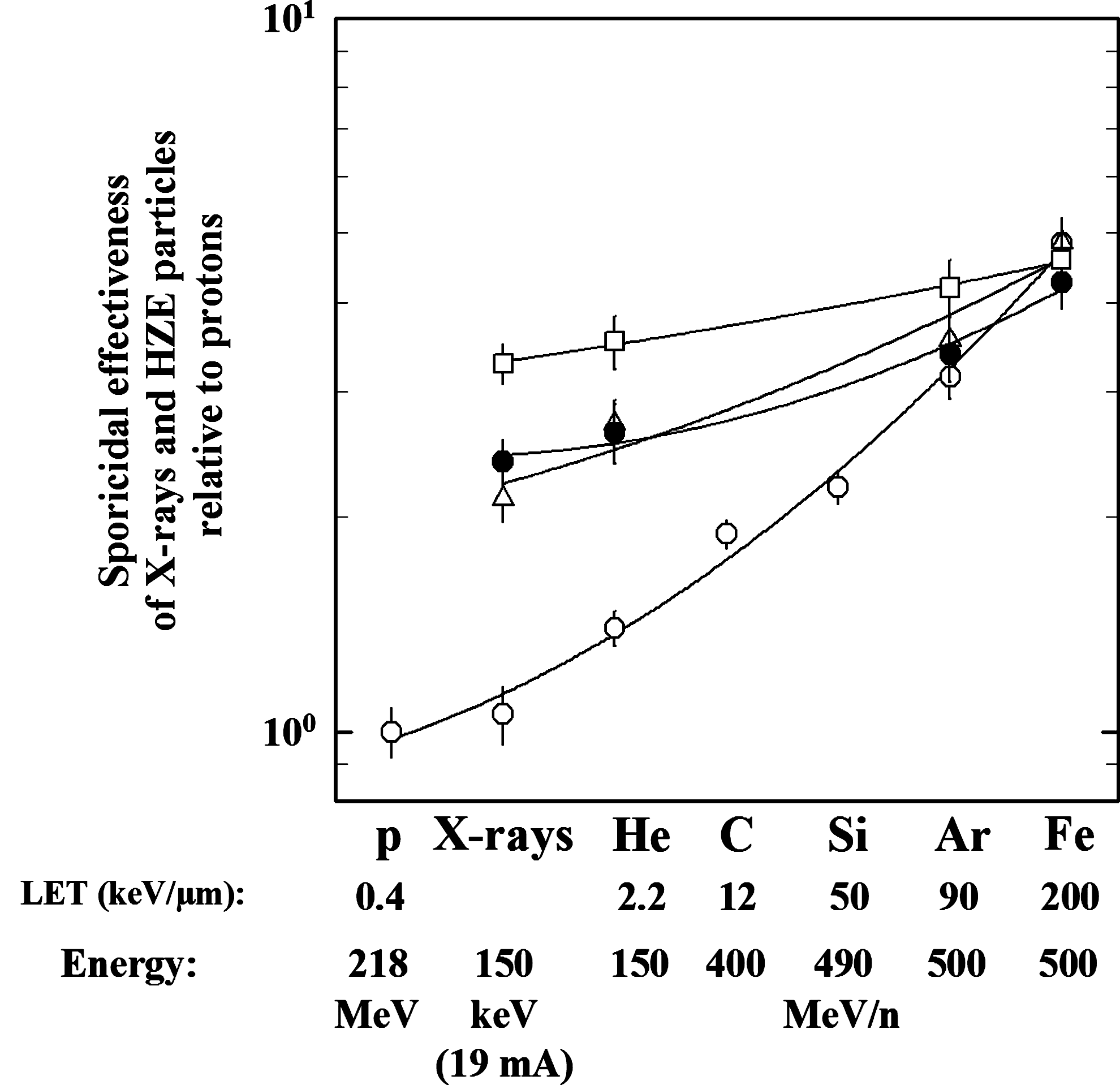
Sporicidal effectiveness (based on inactivation rate constants) of X-rays and HZE particles relative to 218 MeV proton radiation in wild-type and mutant B. subtilis spores. Strains are wild-type strain WN131 (open circles), HR-deficient strain WN463 (open triangles), strain WN1273 lacking α/β-type SASP (filled circles), and HR+NHEJ-deficient strain WN1141 (open squares). Data for X-rays and HZE particles are modified from Moeller et al. (2008, 2010). Error bars for survival data not shown were smaller than the plot symbol.
4. Discussion
Protons are the most abundant type of ionizing radiation in space (Badhwar and O'Neill, 1994; Benton and Benton, 2001; McKenna-Lawlor et al., 2011); therefore, it is of astrobiological relevance to understand the interaction of protons with living organisms. One of the main features of charged particles is their LET value, which is related to the mass of the particle. The density of ionization events along a particle track is much higher for particles with high LET, such as Fe (200 keV/μm), than for low-LET particles such as protons (0.4 keV/μm) (Goodhead, 1999). Of all cellular components, DNA is considered to be the most important radiosensitive target in biological systems (Hutchinson, 1985; Sutherland et al., 2000; Hada and Sutherland, 2006; Yokoya et al., 2008). Depending on the species analyzed, spores are up to 10 times more resistant than growing cells to ionizing radiation, whereas the magnitude of the difference in ionizing radiation resistance between spores and growing cells can be different at different types of ionizing radiation (Moeller et al., 2007; Horneck et al., 2010; Wassmann et al., 2010). DNA strand breaks are major lethal lesions in the genome of spores exposed to ionizing radiation (Micke et al., 1994). Such damages are caused either by interaction of DNA directly with the highly energetic particles or indirectly via interaction with radiation-induced free radicals such as reactive oxygen species (Hutchinson, 1985; Cadet et al., 1999; Dianov et al., 2001).
4.1. Spore protection from proton radiation damage
Cellular damage to dormant bacterial spores poses a unique problem because spores are metabolically inactive, and dormancy can persist for extremely long time periods (reviewed in Nicholson et al., 2000). Therefore, dormant spores can accumulate substantial amounts of DNA damage in an unrepaired state. This cumulative DNA damage must be repaired rapidly during the process of germination, before spores can reactivate gene expression and return to vegetative growth (Setlow and Setlow, 1996). Spores have evolved several mechanisms both for protection of their DNA from damage and for DNA repair during germination (Setlow, 2006, 2007). Spore protective strategies include production of two thick and highly cross-linked proteinaceous spore coat layers (Driks, 2002); production and deposition of a melanin-like UV-protective pigment in the spore coat layer (Hullo et al., 2001); dehydration of the spore core via an unknown mechanism involving the spore cortex protein DacB (Popham et al., 1995; Moeller et al., 2009); production and storage of large quantities of the calcium chelate of dipicolinic acid (Ca-DPA) in the spore core (Paidhungat et al., 2000; Slieman and Nicholson, 2001); and saturation of spore DNA with α/β-type SASP (reviewed in Nicholson et al., 2000; Setlow, 2006, 2007).
The results from the present experiments indicated that pigmentation of the spore coat layers, and indeed the spore coats themselves, did not significantly protect spores from the lethal effects of high-energy proton irradiation. In contrast, spores deficient in various spore core components (core dehydration, Ca-DPA, and α/β-type SASP) were significantly more sensitive than spores of the wild-type strain to high-energy protons. The α/β-type SASP bind to DNA and have also been shown to play a major role in spore DNA protection from several extreme treatments such as UV radiation, oxidizing agents, heat (Setlow, 2007), and ionizing radiation such as X-rays and HZE particle bombardment (Moeller et al., 2008). The water content of vegetative B. subtilis cells is ∼80%, and the core of wild-type spores is relatively dehydrated (∼40%) (Nicholson et al., 2000; Moeller et al., 2009). Spores of mutant strains deficient in the DacB spore cortex protein contain a higher core water content (∼65%), leading to decreased resistance to wet heat, environmental UV radiation (>280 nm) and hydrogen peroxide (Popham et al., 1995; Moeller et al., 2009). In this study, we observed that spores with higher water content were also significantly more sensitive to proton radiation than wild-type spores, likely due to increased production of ionized species such as reactive oxygen species (Lomax et al., 2002). DPA is a major component of the spore core, and studies in B. subtilis have shown that spores lacking Ca-DPA also have significantly increased spore core water and are more sensitive to heat, H2O2 (Paidhungat et al., 2000), and UV radiation (Slieman and Nicholson, 2001). In our experiments, air-dried DPA-lacking spores were significantly more sensitive to proton radiation than the DPA-containing wild-type spores, indicating that DPA protects spore DNA, and perhaps other biomolecules, in the spore interior.
4.2. Contribution of HR and NHEJ DNA repair to spore proton resistance
Despite the protective mechanisms described above, potentially lethal or mutagenic damage eventually accumulates in the DNA of spores during exposure to environmental stresses. One of the aims of this work was to assess the contribution of the HR and NHEJ DNA repair pathways in spore resistance to proton radiation. It was observed that spores lacking either HR or NHEJ singly, as well as the HR+NHEJ-deficient double mutant, were all significantly more sensitive to proton exposure than wild-type spores. Analysis of their relative sensitivities by comparing D 10 values (Table 2) showed that NHEJ-deficient and HR-deficient spores were 2.36- and 1.64-fold more sensitive to protons than wild-type spores, respectively. The HR+NHEJ double mutant spores were 3.51-fold more sensitive than wild-type spores, indicating (i) that NHEJ contributes more than does HR to spore proton resistance and (ii) that the NHEJ and HR pathways act additively to repair proton radiation damage during spore germination.
4.3. Implications for lithopanspermia and planetary protection
Approximately 1% of wild-type B. subtilis spores survived after exposure to high doses (up to 2.5 kGy) of highly energetic (218 MeV) protons (Fig. 1). Therefore, a substantial fraction of spores is capable of surviving exposure to one of the major components of space radiation encountered during the second stage of lithopanspermia, transit through interplanetary space (Nicholson, 2009; Horneck et al., 2010). Concerning shielding against radiation in space, a few micrometers of meteorite material are sufficient to give efficient protection against solar UV radiation if the material is without cracks (Horneck et al., 2001); however, higher shielding requirements are necessary to protect against the more penetrating lethal effects of high-energy particles such as protons (reviewed in Nicholson et al., 2000; Nicholson, 2009; Horneck et al., 2010). In the open space environment, assuming an average total dose of high-energy protons of 0.2–0.5 Gy per year (depending on various physical parameters, e.g., solar activity, contribution of high- and low-energy charged particles) (Cucinotta and Durante, 2006; Held, 2009), spores on a spacecraft with an aluminum covering (shielded from solar UV) would require approximately 2256–5640 years (for 218 MeV protons) to reach the D 10 value determined in this communication. Taking into account that secondary radiation (e.g., electron-induced bremsstrahlung radiation) is created by the transport and interaction of primary particles through the spacecraft materials, spore survival inside a meteorite could vary due to the mineralogical composition and its shielding ability in a lithopanspermia-relevant interplanetary journey. A time span of approximately 6000 years complies with estimates of the time required for boulder-sized rocks to travel from one planet of our solar system to another, for example, from Mars to Earth (Melosh, 1988; Nicholson, 2009). In addition, a small fraction of meteorites ejected from the surface of Earth or Mars would be placed on trajectories favorable for transit between the two planets in a matter of months, similar to spaceflight transit times (Melosh, 1988; Horneck et al., 2010). Based on these considerations, proton irradiation in interplanetary space would likely not be sufficient to completely sterilize all rocks transferred between Earth and Mars.
In the context of planetary protection, bacterial spores have been identified as major spacecraft contaminants of concern in the prevention of forward contamination of astrobiologically interesting bodies such as Mars (Venkateswaran et al., 2004; Bruckner et al., 2008; Nicholson et al., 2009). The data reported in this communication also suggest that bacterial spores contaminating UV-shielded or interior portions of spacecraft would be exposed to insufficient proton radiation for their complete inactivation during a typical Earth-to-Mars voyage of 6–8 months' duration.
A future goal of our work is to compile a complete catalogue of all types of DNA damages incurred in spores exposed to various forms of ionizing radiation and to elucidate the roles of various DNA repair systems (e.g., direct reversal, base excision repair, nucleotide excision repair) utilized during spore germination to repair these lesions, in order to gain deeper understanding into the physiological responses of microbes exposed to extraterrestrial solar and galactic radiation.
Footnotes
Acknowledgments
The authors are very grateful to Roelf Slopsema, Jeff Rexford, Thomas Berger, and Daniel Matthiä for consulting, providing technical data, and discussion. We express gratitude to Adam Driks and Peter Setlow for their generous donation of Bacillus subtilis strains. We express thanks to the two anonymous reviewers for insightful comments. R.M. and G.R. were supported by the DLR grant DLR-FuE-Projekt ISS-Nutzung in der Biodiagnostik, Programm RF-FuW, Teilprogramm 475. W.L.N. was supported by grants from the NASA programs Planetary Protection (NNA06CB58G) and Astrobiology: Exobiology and Evolutionary Biology (NNX08AO15G).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
Ca-DPA, calcium chelate of dipicolinic acid; Cm, chloramphenicol; DPA, dipicolinic acid; DSB, double-strand breaks; Erm, erythromycin; GCR, galactic cosmic radiation; HR, homologous recombination; HZE, high-charge and high-energy particles; LET, linear energy transfer; NHEJ, non-homologous end joining; RBE, relative biological effectiveness; SASP, small, acid-soluble spore proteins; SPE, solar particle events; Spec, spectinomycin; SSB, single-strand breaks; Tet, tetracycline.