
Abstract
Humic substances (HS) are high-molecular-weight complex refractory organics that are ubiquitous in terrestrial and aquatic environments. While resistant to microbial degradation, these compounds nevertheless support microbial metabolism via oxidation or reduction of their (hydro)quinone moieties. As such, they are known to be important electron sinks for respiratory and fermentative bacteria and electron sources for denitrifying and perchlorate-reducing bacteria. HS also strongly promote abiotic reduction of Fe(III) when irradiated with light. Here, we show that HS-enhanced Fe(III) photoreduction can also drive chemolithotrophic microbial respiration by producing Fe(II), which functions as a respiratory electron donor. Due to their molecular complexity, HS absorb most of the electromagnetic spectrum and can act as broad-spectrum antennae converting radiant energy into bioavailable chemical energy. The finding that chemolithotrophic organisms can utilize this energy has important implications for terrestrial, and possibly extraterrestrial, microbial processes and offers an alternative mechanism of radiation-driven primary productivity to that of phototrophy. Key Words: Deep subsurface biosphere—Chemolithotrophic microorganisms—Organic matter—Geochemistry—Iron-oxidizing bacteria. Astrobiology 13, 476–482.
1. Introduction

Proposed model indicating the HA-mediated conversion of light energy into the recyclable electron donor Fe(II). Fe(II) oxidation is known to support respiratory reduction of oxygen, nitrate, and perchlorate, as well as photosynthesis.
2. Materials and Methods
2.1. Media/solution conditions
Fer1T was maintained on Fe(II), glucose, and yeast extract in acidic AMD media (pH=1) described by Dopson et al. (2004). Mineral mix for the media was prepared as described in Lovley et al. (1984). Solutions for illumination, as well as dark-treated controls, were made from double deionized water adjusted to pH 1.1 [for 2,6-anthrahydroquinone disulfonate (AQDS) solutions] or 2.1 (for HA solutions) with sulfuric acid and amended with either AQDS (2 g·L−1) or HA (0.5 g·L−1) as necessary. Select experiments were performed in phosphate-buffered water (pH 7.2). For oxic solutions, oxic double deionized water was amended with AQDS or HA, sealed in serum bottles, and autoclaved prior to experimental use. For anoxic solutions, AQDS or HA were dissolved in 50 mL of double deionized water within a serum bottle. Solutions were then flushed with N2 for 25 min and sealed with thick butyl rubber stoppers prior to autoclaving (121°C for 20 min).
2.2. Analytical techniques
Fe(II) production was assessed via the colorimetric ferrozine assay as previously described (Stookey, 1970). The ferrozine assay was conducted in 50 mM HEPES buffer (pH=7), and all results were normalized for light absorption by AQDS or HA samples in water at the reference wavelength of 562 nm. Growth of Fer1T was measured by direct cell counts in a Hausser Scientific counting chamber.
2.3. Humic materials
Five different HA representing coal extract, lake, swamp, and marine sediments, respectively, were used in experiments. Aldrich brand HA and freshwater IHSS Nordic Reference HA (
2.4. Irradiation procedure
For broad-spectrum light experiments, a light box was constructed with a 15 W General Electric Sunshine F15 bulb (λ>approximately 390 nm). For UV experiments, a light box was constructed with a 15 W EIKO Hg fluorescent bulb (F15T8/BL) (2.6 W, 352 nm peak wavelength). Ambient temperature on the surface of the bulb was between 45°C and 50°C. Oxic or anoxic solutions containing AQDS, HA, or lacking organics were decanted into 13 mm diameter fused quartz tubing, which was capped at either end by thick rubber butyl stoppers. These “reaction tubes” were placed directly on the fluorescent bulb for illumination. Illumination times are noted for each experiment. For oxic illuminations, oxic solutions were placed into quartz tubes that were capped under atmospheric conditions. For anoxic illuminations, anoxic solutions were added to N2-flushed quartz tubes, and solutions were transferred to and from the quartz tubes with standard anoxic techniques. Dark control tubes were wrapped in tin foil prior to illumination. For experiments involving Fer1T, media salts and 0.02 g·L−1 yeast extract were added to irradiated solutions after the UV exposure period from sterile, anoxic stocks. In this manner, yeast extract and salts were omitted from potential photochemical reactions. Irradiation of HA with isotopic decay products was achieved in a gammator utilizing a Cs-137 source (a gamma and beta emitter). Glass bottles containing solutions for irradiation were placed in the gammator for 48 h (4.8 kGy total exposure).
2.5. Co-irradiations
“Co-irradiations” refer to the irradiation of experimental solutions previously amended with iron. Anoxic, sterile stock solutions of FeCl3 were directly amended to quartz tubes to an approximate final concentration of 1.5 (AQDS) or 3 mM (HA) Fe(III) prior to illumination. After co-illuminations, solutions were amended with yeast extract and salts to supplement missing media components, and Fe(II) production was assessed.
2.6. Photoreductions
“Photoreductions” refer to the illumination of solutions (with AQDS, HA, or without organics) alone, followed by subsequent immediate addition of Fe(III). Fe(II) in this case is generated by dark reactions after the test solution is removed from the irradiation source. For AQDS photoreductions, synthetic ferric (hydr)oxides [Fe(III)-HFO] were added as a Fe(III) source [final Fe(III) concentration=2.0 mM]. For HA photoreductions, 3.0 mM FeCl3 served as an Fe(III) source. In the case of visible light, illuminated HA solutions were exposed to Fe(III)-citrate. In all cases, Fe(III) stock solutions were made anoxic via sparging with N2 or 80:20 N2:CO2 prior to use, and Fe(III) solutions were transferred to reaction tubes by using standard anoxic techniques. After Fe(III) addition, reaction tubes were incubated in the dark at 37°C for approximately 3 h (AQDS) or 1 h (HA) to allow for reaction between reduced media components and Fe(III) under corresponding oxic or anoxic conditions. Production of Fe(II) due to dark reactions was then measured.
2.7. Oxidation of photoproduced Fe(II)
After Fe(II) photoproduction, media was dispensed into 10 mL serum bottles and exposed to atmospheric oxygen. Active cultures of Fer1T grown in AMD media (6–18 mL) were collected and centrifuged for 10 min at 6000 rpm. Cell pellets were resuspended in 1 mL of AMD prepared without Fe(II). One hundred to two hundred microliters (consistent within each experiment) of this concentrated cell solution was added to serum bottles. Negative controls were amended with equivalent volumes of cell-free, Fe(II)-lacking AMD media. Triplicate bottles were aerobically shaken in the dark at 37°C, while Fe(II) and Fe(III) concentrations were monitored.
2.8. Irradiation of HA solutions
Aldrich brand HA (0.5 g·L−1) were dissolved in 10 mL of anoxic 10 mM phosphate buffer (prepared as previously described) prior to autoclaving in sealed, 20 mL serum bottles. Serum bottles containing or lacking HA were exposed to radiation within a gammator as described above. Radiation-free controls were maintained in dark conditions. After irradiation, anoxic Fe(III)-citrate was added to bottles (final concentration=100 mM). Fe(II) production was examined via the ferrozine assay after a 1 h reaction period at room temperature.
3. Results
Using a broad-spectrum light source (according to manufacturer specifications, λ>390 nm, with most emission between approximately 400 and 650 nm), we irradiated anoxic phosphate-buffered (10 mM, pH 7.2) aqueous solutions of commercial Aldrich HA (1.0 g·L−1) in the presence of Fe(III) for 18 h. Irradiation of HA-Fe(III) solutions increased Fe(II) production by 310±10 μM (n=3) over dark-treated controls (Table 1; Supplementary Fig. S1, available online at
Unless otherwise noted, calculated as the difference in Fe(II) concentration between light- and dark-treated samples.
Photoproduced Fe(II) was calculated as the difference in Fe(II) concentration before and after irradiation.
Unless otherwise noted, calculated as the difference in Fe(II) concentration between light- and dark-treated samples.
Photo-produced Fe(II) was calculated as the difference in Fe(II) concentration before and after irradiation.
Although unstable under oxic conditions at circumneutral pH, Fe(II) is nonreactive with oxygen under acidic conditions (pH<5) such as those found naturally in various environments, including volcano crater lakes, terrestrial hot springs, and saline lacustrine or groundwater systems. Acidic, metalliferous environments are also commonly found in areas of active or abandoned mining operations (Dopson et al., 2004). When oxic and anoxic Fe(III)-containing acidic solutions (pH=2.1) of HA from lake, swamp, and marine environments were irradiated with UV light (peak wavelength=352 nm) for 19.5 h, large quantities of Fe(II) (1660±230 to 2340±300 μM, n=3) were produced relative to dark controls (Table 1; Supplementary Fig. S1). In the absence of HA, UV irradiation of Fe(III) solutions produced only 200 and 560 μM Fe(II) under oxic and anoxic conditions, respectively, demonstrating that the HA greatly enhanced UV photoproduction of Fe(II). Even under oxic conditions, HA retain some intrinsic capacity for light-independent Fe(III) reduction (Kappler et al., 2004). As such, HA solutions foregoing light treatment produced some Fe(II) (650±30 to 1400±80 μM, n=3) in response to Fe(III) amendment. However, UV irradiation of HA-Fe(III) solutions increased Fe(II) production by 217–378% compared to dark redox reactions occurring over the same time period. Similarly, UV irradiation of Fe(III) in the presence of the model humic quinone analogue, 2,6-anthrahydroquinone disulfonate (AQDS) (Tratnyek and Macalady, 1989) greatly enhanced photoproduction of Fe(II) under both oxic and anoxic conditions. UV irradiation of AQDS in the presence of Fe(III) for 21.5 h produced substantial amounts of Fe(II) under oxic (2140±60 μM, n=3) and anoxic (2200±30 μM, n=3) conditions, and little to no Fe(III) reduction was observed in identical treatments wrapped in aluminum foil prior to light exposure (Table 1).
Many microorganisms oxidize Fe(II) as an electron donor for the respiration of oxygen, nitrate, or perchlorate (Weber et al., 2006a; Thrash et al., 2011; Carlson et al., 2012). Anabolism during Fe(II) oxidation is accomplished via carbon fixation or assimilation of fixed carbon. HA-enhanced, photoproduced Fe(II) therefore represents a potentially growth-supporting chemical substrate. We investigated biooxidation of HA-mediated photoproduced Fe(II) with the acidophilic archaeon Ferroplasma acidarmanus Fer1T. Isolated from acid mine drainage, Fer1T chemolithotrophically grows at pH 0.2–2.5 via oxidation of Fe(II) with glucose as a carbon source (Dopson et al., 2004). Fer1T is typically cultivated by using FeSO4·7H2O as a source of ferrous iron [20·g·L−1, approximately 72 mM Fe(II)], a significant portion of which is converted to Fe(III) during cultivation. Regardless of the HA source, UV irradiation of HA-Fe(III) solutions produced Fe(II), which was rapidly oxidized by Fer1T in the presence of O2, as is shown for Aurevann Lake HA (Fig. 2). No Fe(II) oxidation occurred in the absence of Fer1T, suggesting that the reaction was biologically mediated.
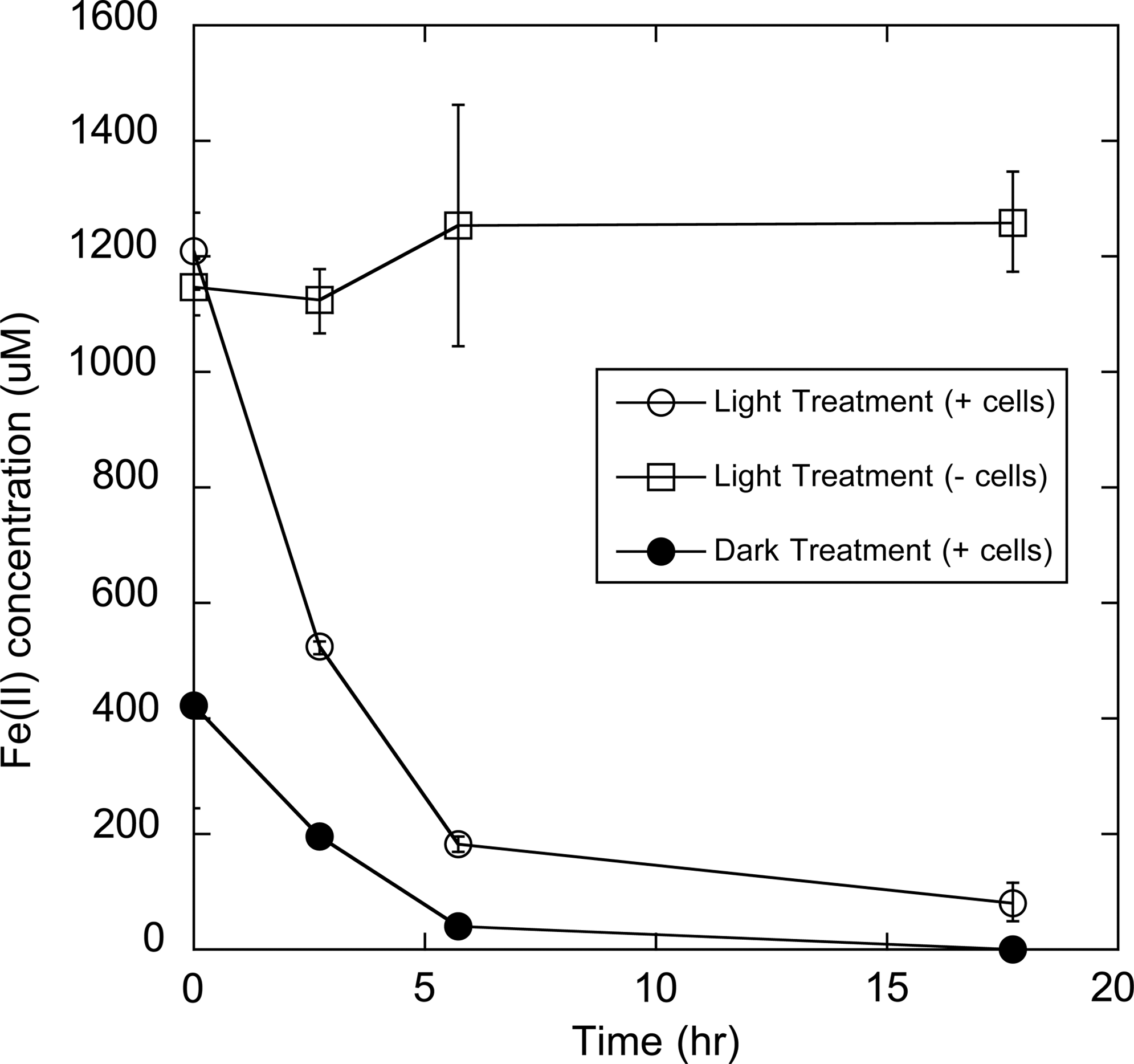
Oxidation of Fe(II) produced from UV irradiation of Aurevann HA-Fe(III) solutions (open circles) and dark reactions (closed circles). Aurevann HA were present at 0.5 g·L−1 at pH=2.1, and the source of Fe(III) was FeCl3. UV irradiation was carried out for 19.5 h under oxic conditions. Open squares represent cell-free controls of photoreduced HA-Fe(III) solutions.
Ultraviolet irradiation of AQDS solutions alone resulted in the production of reduced chemical species capable of reducing ∼50% (830±80 and 1020±80 μM, n=3, in anoxic and oxic incubations, respectively) of the subsequently added 2.0 mM Fe(III) to Fe(II) in dark reactions (Table 2). Fe(II) produced from dark reactions following the UV irradiation of AQDS was also readily reoxidized by Fer1T, and no Fe(II) reoxidation was observed in the absence of cells (Fig. 3). Notably, Fe(II) concentrations resulting from dark reactions continued to increase over the experimental time period in the absence of Fer1T. This may be due to additional reactions between Fe(III) and multiple redox-active chemical species within the media. These results are consistent with similar observations made on the addition of Fe(III) to anoxic HA after UV irradiation, which also produced electron-donating species capable of Fe(III) reduction (Table 2). Anoxic UV irradiation of Nordic Reference HA increased subsequent, dark Fe(II) production by ∼38%, and this additional Fe(II) was rapidly oxidized in the presence of Fer1T (data not shown). Notably, oxic UV irradiation of HA tended to decrease their overall ability to produce Fe(II), possibly due to oxidative degradation of the HA.
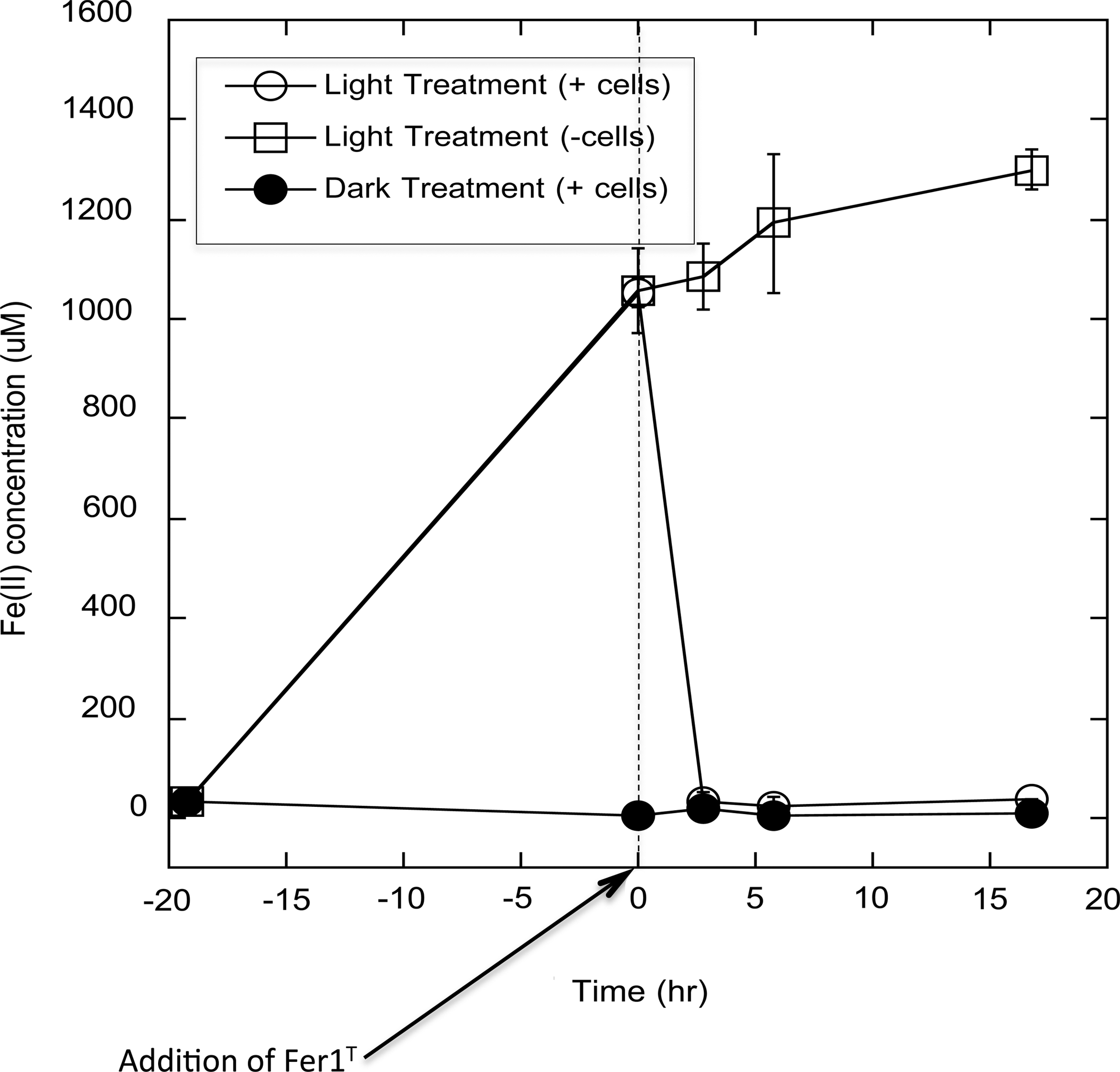
Fe(II) production by exposing Fe(III)-HFO to previously UV-irradiated oxic AQDS solutions, and Fer1T-dependent reoxidation of photoproduced Fe(II). AQDS (2 g·L−1 at pH=1.0) was UV irradiated for 19.3 h. Fer1T was added to incubations at h=0. Open circles represent Fe(II) concentrations before and after addition of Fer1T. Closed circles represent Fe(II) present in dark-treated controls amended with Fer1T. Open squares correspond to UV-irradiated cell-free controls.
Because of their complex molecular structures, capacity for photolytic degradation into biologically labile organics, and intrinsic redox reactivity, HA could potentially stimulate growth through several alternative mechanisms. We therefore utilized AQDS to determine whether or not growth could be coupled specifically to the HA-assisted photoreduction of Fe(III). An aerobic solution of 2 g·L−1 AQDS (pH=1.1) was irradiated with UV light for 19.5 h in the presence of FeCl3, generating approximately 2080 μM Fe(II). Upon inoculation with an active culture of Fer1T, the photoproduced Fe(II) was rapidly reoxidized over an 18 h incubation (data not shown). Oxidation of this photoproduced Fe(II) occurred with concomitant growth of Fer1T resulting in an almost 4-fold cell density increase from 1.0±0.3×107 to 3.8±0.2×107 cells·mL−1 (n=3) (Fig. 4). No statistically significant growth was observed when Fer1T was introduced to control media foregoing light treatment, indicating that the HA-mediated transformation of light energy was required for cellular replication to occur.
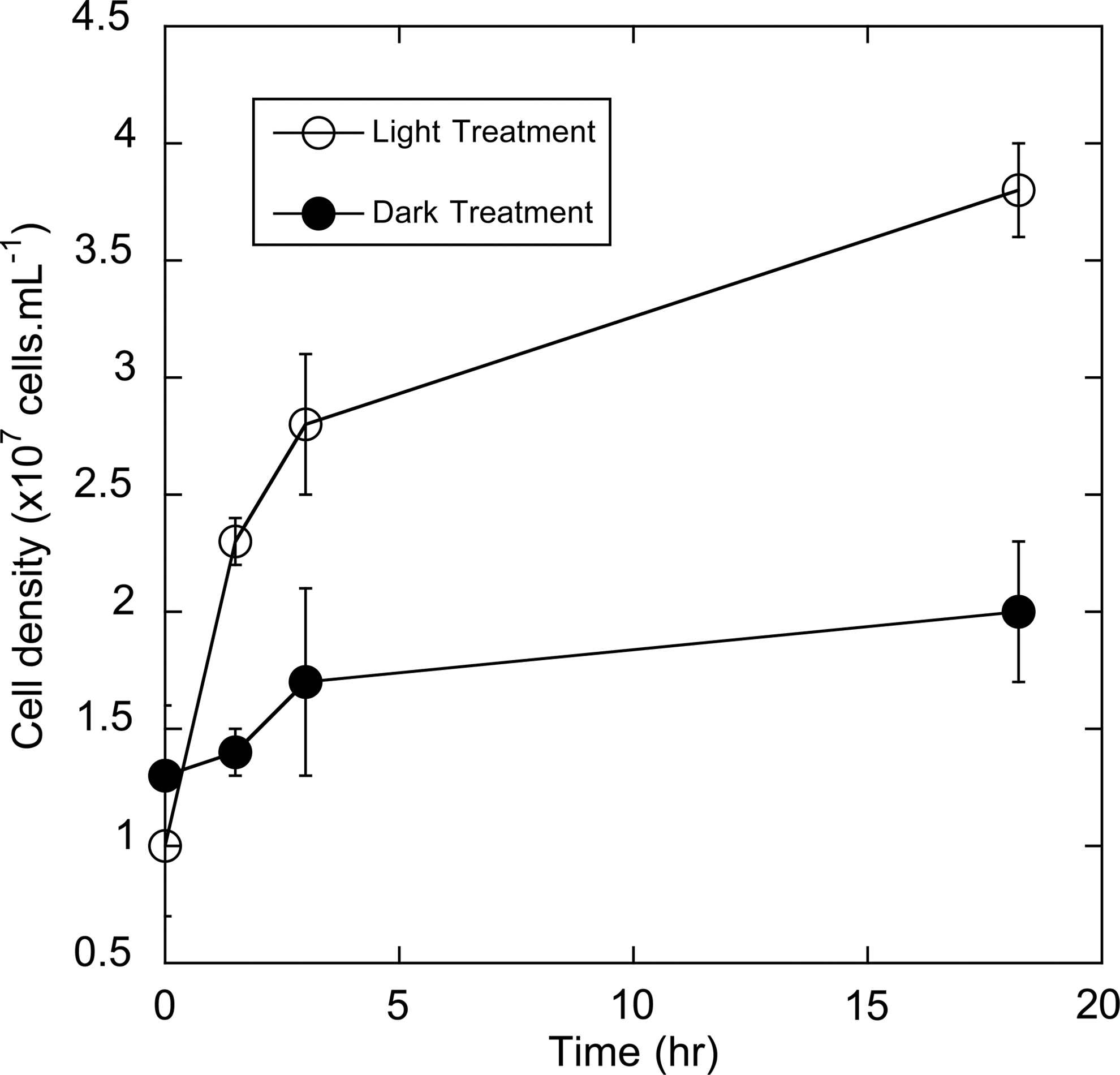
Growth of Fer1T while oxidizing photoproduced Fe(II) (open circles), or in solutions foregoing light treatment (closed circles). Fe(II) was generated in this experiment via the UV irradiation of 2 g·L−1 AQDS (pH=1.1, amended with FeCl3) for a period of 19.3 h under oxic conditions.
4. Discussion
The findings of these studies suggest that HA aid in the conversion of broad spectrum and UV radiation energy into the bioavailable, growth-supporting chemical substrate Fe(II). Although it is known that UV illumination increases the lability of HA to organotrophic metabolism (Wetzel et al., 1995; Van Trump et al., 2006), our data suggest that irradiation of HA can also produce inorganic electron donor substrates. Ultimately, radiation provides the energy for electron transfer to Fe(III) (Spokes and Liss, 1995), thereby converting radiant energy into chemical energy that is exploitable by non-photosynthetic microorganisms. While photoreductive dissolution may increase the bioavailability of iron as a macronutrient (Waite and Morel, 1984; Miller and Kester, 1994), these reactions have not been investigated in terms of biological energy production. Our results suggest that, at least under acidic conditions, HA-mediated photoderived Fe(II) can function as a suitable electron donor for aerobic and microaerophilic respiration by Fe(II)-oxidizing bacteria. Preliminary studies performed with the phylogenetically diverse species Acidothiobacillus ferrooxidans concurred with the presented data (data not shown), indicating that this is not a species-specific metabolic capacity. Our results also imply a role for incident light in (re)generating electron sources for Fe(II)-oxidizing photosynthetic organisms in similar environments.
Although we demonstrate the HA-enhanced photoreduction of Fe(III) under anoxic conditions, anoxic environments receiving sufficient light to produce significant amounts of Fe(II) may be limited on Earth (although such conditions could conceivably exist on other planetary bodies). Therefore, other types of electromagnetic radiation, such as isotopic decay, may be more relevant for HA-enhanced Fe(II) production in anaerobic environments. To test this possibility, we exposed anoxic, phosphate-buffered (pH=7.2) solutions of 0.5 g·L−1Aldrich HA to a Cs-137 radiation source (4.8 kGy total exposure) in the absence of Fe(III). Subsequent amendment of the irradiated samples with Fe(III) led to the production of 300±90 μM Fe(II) (n=3), while non-irradiated samples only produced 40±10 μM Fe(II) (n=3), indicating that radiation exposure increased the production of Fe(II) by 260 μM (Table 2). No Fe(III) was reduced upon exposure to irradiated media lacking HA, demonstrating that HA were necessary for the observed increase in reducing capacity and emphasizing their potential role in mediating transformation of a broad range of electromagnetic radiation into chemical energy. Many microorganisms existing in the nutrient-depleted deep subsurface may take advantage of this reaction. Previous reports have found that isotopic decay produces H2 gas from irradiated water (Lin et al., 2005; Blair et al., 2007). This H2 is a potential electron donor for a number of respiratory processes. Results presented here suggest another mechanism, dependent on natural organic matter and iron, whereby radioactive decay products may support primary production in the subsurface.
5. Significance
Photochemical reactions have been implicated in the geochemical cycling of iron in surface waters. Photochemical reactions with iron have also been cited as important for mediating the bioavailability of iron nutrients (Waite and Morel, 1984; Miller and Kester, 1994). However, the potential role of radiation and humic materials in producing Fe(II) as a growth-supporting chemolithotrophic electron donor has not been previously described. Our results indicate that microbial oxidation of Fe(II) produced in this manner may play a role in both biochemical energy production and the biogeochemical cycling of iron.
Humic materials are ubiquitous in the environment, and Fe is abundant throughout much of the planet (Van Trump et al., 2006; Weber et al., 2006a). Likewise, Earth is constantly subjected to irradiation, not only from the Sun on Earth's exterior but also from isotopic decay kilometers beneath the surface. Data presented here suggest that such radiation, combined with natural organic matter and iron, can come together and contribute to primary productivity on Earth. These results also suggest a mechanism by which bioavailable chemical energy could be produced on other planets with similar radiation sources, thereby expanding our understanding of how primary energy production could possibly occur on other planetary systems.
Footnotes
Acknowledgments
Research on the microbial interactions with humic substances and microbial Fe(II) oxidation in the laboratory of J.D.C. is supported by grant funding from the Natural Research Initiative of the USDA through grant number 2005-35107-16237 and the US Department of Energy, Environmental Remediation Sciences Program, through grant #DE-FG02-98ER63592. The authors wish to acknowledge S. O'Connor and Dr. E. Gjessing for their provision of several HA for this project and Dr. J. Banfield and Dr. C. Belnap for their provision of an active culture of strain Fer1T.
Abbreviations
AQDS, 2,6-anthrahydroquinone disulfonate; HA, humic acids; HS, humic substances.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.