
Abstract
Carotenoids are common components of many photosynthetic organisms and are well known from the red waters of hypersaline ecosystems where they are produced by halophilic algae and prokaryotes. They are also of great interest as biomarkers in extraterrestrial samples. Few laser Raman spectroscopy studies have examined ancient field samples, where pigments and microscopic life are less defined. Here, we have identified carotenoids in ancient halite brine inclusions, 9 ka to 1.44 Ma in age, from borehole cores taken from Death Valley, Saline Valley, and Searles Lake, California, for the first time with laser Raman spectroscopy. Carotenoids occurred in fluid inclusions as colorless to red-brown amorphous and crystalline masses associated with spheroidal algal cells similar in appearance to the common halophilic alga Dunaliella. Spectra from carotenoid standards, including β-carotene, lycopene, and lutein, were compared to microscopically targeted carotenoids in fluid inclusions. Carotenoids produced characteristic bands in the Raman spectrum, 1000–1020 cm−1 (v 3), 1150–1170 cm−1 (v 2), and 1500–1550 cm−1 (v 1), when exposed to visible laser excitation. Laser Raman analyses confirmed the presence of carotenoids with these characteristic peaks in ancient halite. A number of band sets were repeated at various depths (ages), which suggests the stability of this class of organic molecules. Carotenoids appear well preserved in ancient salt, which supports other observations, for example, preserved DNA and live cells, that fluid inclusions in buried halite deposits preserve intact halophilic microbial ecosystems. This work demonstrates the value of laser Raman spectroscopy and carotenoids in extraterrestrial exploration for remnants of microbial life. Key Words: Raman spectroscopy—Biomarkers—Halite fluid inclusions—Algae—Mars. Astrobiology 13, 1065–1080.
1. Introduction
C
Carotenoids are tetraterpenoids (C40), each with a conjugated polyene chain. The family is divided into two groups: carotenes that are strict hydrocarbons, first discovered in 1831 (Berzelius, 1837), and xanthophylls, which contain oxygen (Britton, 1995). Carotenoids recovered from ancient sediments and sedimentary rocks are of great interest because they are indicators of crude oil maturation (Koopmans et al., 1997). There is now rising interest in using carotenoids as extraterrestrial biomarkers in ancient sediments (Vítek et al., 2009). They are also well known from the red saline waters of hypersaline ecosystems where halophilic algae and prokaryotes, such as Dunaliella and Halobacterium, produce them. Here, we document the widespread preservation of carotenoids associated with pigmented, Dunaliella-like algal cells in tiny pockets of brine, called fluid inclusions, in ancient halite (NaCl) from Saline Valley, Death Valley, and Searles Lake, California, up to 1.44 Ma in age.
1.1. Ancient carotenoids
Cyclic and branched hydrocarbons are valuable biomarkers because of their long-term stability (Simoneit, 2004). Intact carotenoids, β-carotene, and α-carotene, for example, were found in 20 ka sediments from Searles Lake, California (Vallentyne, 1956). Anoxic marine sediments from the Cariaco Trench, offshore Venezuela, 5, 56, and 340 ka in age, all contained unaltered β-carotene (Watts and Maxwell, 1977). β-carotene was identified in a 100 ka interglacial freshwater gyttja (sediment-rich in organic matter common at the bottom of eutrophic lakes) from Rodebaek, Denmark (Andersen and Gundersen, 1955). Isorenieratene was preserved in a Messinian (6 Ma) Italian marl (Keely et al., 1995) and Permian-Triassic sediments from southern China (Grice et al., 2005), and a 20 Ma carotenoid was discovered in clays from a sediment core in the Blake-Bahama Basin (Cardoso et al., 1978).
Although the carotenoid family is stable over long periods of time, chemical changes may cause the transformation of one carotenoid to another; known degradation pathways are shown in Fig. 1. Carotenoids contain numerous C=C double bonds in their structures that are easily altered (Damsté and Koopmans, 1997). The following examples illustrate their molecular lability: (1) Eocene (56–34 Ma) diagenetic products of β-carotene, including reduced forms such as β-carotane and lexane, were identified from Green River shale by gas chromatography–mass spectrometry (Koopmans et al., 1997). (2) Marine sediments from the Cariaco Trench showed carotenoid-specific reduction with increasing depth (Watts and Maxwell, 1977). Zeaxanthin, for example, had two C=C double bonds reduced at the 340 ka sampling depth. Canthaxanthin experienced the same amount of reduction by 56 ka. Spheroidenone and echinenone were absent altogether from the 340 ka, and the 56 and 340 ka intervals, respectively (Watts and Maxwell, 1977). (3) Derivatives of isorenieratene were discovered in Ordovician sedimentary rocks (Koopmans et al., 1996). These studies demonstrate the chemical transformations of individual carotenoids and their overall persistence over geological timescales.

Possible chemical rearrangements of carotenoids via (1) desaturation, (2) cyclization, (3) hydroxylation, (4) epoxidation, and (5) epoxidefuranoxide alteration. Boxes indicate compounds identified in this study. Modified from Rodriguez-Amaya (2001).
Carotenoid degradation depends on burial conditions; they are best preserved in settings with no oxygen or light, and low temperatures (Goodwin, 1980). Sedimentary basins have a slow hydrocarbon transformation rate, whereas hydrothermal conditions promote rapid breakdown (Simoneit, 2004). Carotenoids can oxidize or change their structural arrangement if exposed to high temperature or light (Rodriguez-Amaya, 2001). Ancient intact carotenoids shared one characteristic—anoxia during burial. They were sealed in reduced muds below the surface of Searles Lake (20 ka), oxygen- and light-deprived marine sediments in the Cariaco Trench (5, 56, 340 ka), Denmark's interglacial gyttja (100 ka), Italian marl (6 Ma), and marine clay (20 Ma). Carotenoid-host material from the current study, fluid inclusions in crystals of halite that were buried in the shallow subsurface, also proves to have excellent preservation characteristics, including high salinity and low oxygen. These halites were recovered from burial depths of less than 320 m, so they likely never experienced temperatures above those found in the surface environments in which they precipitated.
1.2. General information on Raman spectroscopy
Carotenoids in ancient sediments and sedimentary rocks are typically identified by mass spectrometry and liquid or gas chromatography. Here, laser Raman spectroscopy was used to identify carotenoids in cellular materials, most notably Dunaliella-like algae, trapped within fluid inclusions in ancient halite. Raman spectroscopy has become an important area of research that involves pigments as potential biomarkers of life in other parts of the Solar System. Such studies involve analysis of various salts for their carotenoid content. For example, Osterrothová and Jehlička (2011) and Jehlička et al. (2009) analyzed β-carotene through gypsum plates and in a powdered gypsum matrix, respectively. Fendrihan et al. (2009) compared Raman spectra from a β-carotene standard to carotenoids within halophilic archaea sealed in laboratory-grown halite crystals. Vítek et al. (2010) searched for β-carotene, lutein, and zeaxanthin in modern halite crusts from the Atacama Desert in Chile.
Raman spectroscopy is a rapid method of identifying organic compounds (Chadha et al., 1993; Rösch et al., 2003) and is generally considered to be nondestructive (Schrader et al., 1999, Schmitt and Popp, 2006; Jehlička et al., 2009). Direct radiation, however, can degrade organic components if the laser beam is concentrated (Merlin, 1985). Laser Raman analysis determines the wavelength and strength of inelastically dispersed light from molecules and their arrangement (Jenkins et al., 2005) by directing a laser beam of a specific wavelength through a microscope to the sample. Much of the laser light is scattered at the same incoming wavelength, but some light causes molecular vibrations (stretching and deformations) and scattering of light at a different wavelength. The instrument detects this wavelength shift and creates a unique Raman spectrum for the substance (Wilson et al., 1955; Schrader, 1973), which gives information about chemical bonds, molecular arrangement, and the relationship between the sample and its environment (Merlin, 1985). Peak intensities provide knowledge on relative proportions of functional groups (Chadha et al., 1993; Baranski et al., 2005).
Raman spectroscopy is a versatile analytical technique because samples can be solid, liquid, or gas; vary in transparency (Schmitt and Popp, 2006); and contain salts (Jorge Villar and Edwards, 2006). Individual compounds are even identifiable in a complex, heterogeneous matrix (Merlin, 1985). Sample preparation is minimal (Rösch et al., 2003), and costly chemicals are not required (Maquelin et al., 2002). It is possible to analyze a sample as small as a single cell, live or dead (Merlin, 1985; Rösch et al., 2003), for its carotenoid content (Baranski et al., 2005) because of the laser focal point capability of 1–2 microns2 (Xie et al., 2001).
1.3. Recent Raman research in astrobiology with emphasis on evaporites
Raman studies applicable to astrobiology have included examination of controlled systems—various UV-protective pigments, known saline minerals likely to be present on the martian surface, and chosen strains of extremophiles that may potentially resemble extraterrestrial microbes. For example, Raman spectra were obtained from cultures of halophilic archaea (Halobacterium salinarum, Halococcus morrhuae, and Natrinema pallidum) to identify the bacterioruberin and bacteriorhodopsin (Marshall et al., 2007). Jehlička et al. (2013) also used Raman spectroscopy to detect α-bacterioruberin in cultures of Halobacterium salinarum strains NRC-1 and R1, Halorubrum sodomense, and Haloarcula vallismortis. Other halophilic archaea (Halococcus dombrowskii, Halococcus morrhuae, Halobacterium salinarum, Halorubrum sp., Halobacterium noricense, and Haloarcula japonica) were trapped in laboratory-grown halite and then analyzed by laser Raman spectroscopy for β-carotene and additional biomolecular components (Fendrihan et al., 2009). Another study included examination of pigment production (zeaxanthin, β-carotene/canthaxanthin, and decapreno-β-carotene) when microorganisms (Bacillus atrophaeus, Bacillus subtilis, Deinococcus radiodurans, and Natronomonas pharaonis) were exposed to UV radiation (Edwards et al., 2006). Raman spectroscopy was also used to find detection limits for β-carotene in various media. β-carotene was identified in concentrations at or below 10 mg·kg−1 in powdered and crystalline halite, gypsum, and epsomite by using an excitation wavelength of 785 nm (Vítek et al., 2009).
Fewer Raman studies have used natural samples, where pigments and microscopic life are less well defined. Edwards et al. (2003) identified β-carotene in cyanobacteria from an Antarctic rock surface. Lutein, α-carotene, and β-carotene were successfully distinguished in various plants grown locally in Quedlinburg, Germany, in the experimental garden of BAZ (Schulz et al., 2005). Jehlička et al. (2013) found bacterioruberin to be the main pigment in a sample of red cells from a saltern. More work in which natural systems are examined is needed to test the feasibility of using Raman spectroscopy for extraterrestrial research.
1.4. Project objective
The purpose of this study was to identify carotenoids in fluid inclusions in ancient halite and to assess any trends with age. Carotenoid-producing haloarchaea and algae found in modern saline ecosystems (Oren, 2005, 2009) provided information on the types of carotenoids expected in ancient samples.
Common haloarchaeal genera include Halobacterium, Halorubrum, Haloferax, and Haloarcula. Haloarchaea have evolved to survive oxygen-limited or oxygen-deprived conditions (Leuko et al., 2010) as anaerobic fermenters but can also produce energy through respiration and photoheterotrophy (Falb et al., 2008). They produce phytoene, β-carotene, bacterioruberin, and lycopene (Goodwin, 1980; Asker and Ohta, 1999).
The eukaryotic unicellular photosynthetic green alga, Dunaliella (Oren, 2005), which belongs to the phylum Chlorophyta (Borowitzka and Siva, 2007), is also prevalent in hypersaline environments and is the primary producer for many of the world's extreme saline environments. Carotenoids, such as β-carotene, lutein, violaxanthin, neoxanthin, zeaxanthin, and low quantities of α-carotene (Goodwin, 1980), are produced in Dunaliella chloroplasts when exposed to intense sunlight or nutrient deprivation (Oren, 2005). Carotenoids found in Chlorophyceae are shown in Table 1, and carotenoids documented in Dunaliella are shown in Table 2. Carotenoid production by algae is highly dependent on environmental conditions. Carotenogenesis in Dunaliella salina is controlled by salinity, nitrogen availability, and UVB incidence. Dunaliella produce carotenoids in highly saline, nitrogen-limited, and UVB exposed environments (Abd El-Baky et al., 2004), conditions which commonly occur in hypersaline lakes and marine waters, evaporation ponds, salterns, and salt flats (Ben-Amotz et al., 2009).
Modified from Table 4-1 of Rowan (1989).
Modified from Goodwin (1980).
Schubert et al. (2009a, 2009b, 2010) investigated communities of carotenoid-producing halophilic microorganisms preserved in fluid inclusions trapped inside halite crystals from Death Valley and Saline Valley. These halites came from modern sediment crusts and borehole cores up to 90 m below the surface. Halophilic prokaryotes and eukaryotes, including pigmented algae that resemble modern Dunaliella, were recognized in ancient fluid inclusions with light microscopy (Schubert et al., 2009a, 2010; Lowenstein et al., 2011). Environmental scanning electron microscopy assisted in distinguishing prokaryotic cells from microparticles. Minerals in fluid inclusions were identified from their crystal habits and optical characteristics (Schubert et al., 2009a). Petrographic properties, along with solubility and melt temperatures, allowed the red, pink, and orange crystals and globules associated with these Dunaliella-like cells in fluid inclusions to be tentatively identified as β-carotene (Schubert et al., 2010).
Communities trapped within fluid inclusions in halite (Fig. 2J, 2L–2N) contain prokaryotes and Dunaliella-like algal cells, microparticles, crystals (Schubert et al., 2009a; Lowenstein et al., 2011), pigmented globules (Fig. 2I and 2K), and fluorescent organic matter (Schubert et al., 2010), all of which are ideal for laser Raman analysis. After burial, fluid inclusions in halite are removed from light and essentially free of oxygen; photochemical and oxygen-related degradation do not affect the biomolecules within.

Color photomicrographs showing modern Dunaliella in broth media (
Laser Raman spectroscopy was used in the present study to identify pigmented materials in fluid inclusions in halite from Death Valley, Saline Valley, and Searles Lake, California. Although archaea and algae both generate carotenoids in saline systems, pigments in fluid inclusions were almost always associated with Dunaliella-like algae (Fig. 2A–2D). Dunaliella are spherical to ellipsoid in shape and vary in size from 4–20 microns in length and 2–20 microns in width (Borowitzka and Siva, 2007). Healthy cells have two flagella at the anterior end of the cell, making them motile. Unhealthy cells lose their flagella during the aplanospore stage. Carotenoids accumulate when exposed to intense sunlight or when experiencing nutrient deprivation. These and other cellular constituents are contained within the plasma membrane rather than a firm, protective cell wall, as in most algae (Oren, 2005).
2. Methods
2.1. Sample sources
Closed basins in California, including Death Valley, Saline Valley, and Searles Lake, provided source material for this study (Fig. 3). Natural samples were collected from sediment cores taken from (1) Death Valley (DV93-1); (2) Saline Valley (SV-4A); and (3) Searles Lake (KM-3) (Fig. 4). DV93-1 was drilled to a depth of 186 m; the upper 36 m (30–43 ka) was used here (Lowenstein et al., 1999). The entire SV-4A core, ∼100 m in length (0–150 ka), was sampled (Howe, 1998; Lowenstein et al., 2011). The Death Valley and Saline Valley cores were dated by using the uranium series isochron method (Lowenstein et al., 1999). KM-3 is 930 m long and dated paleomagnetically to 3.18 Ma at a depth of 693 m (Smith et al., 1983); the upper 319 m (0–1.44 Ma) was used here. All three cores were petrographically examined for carotenoids.

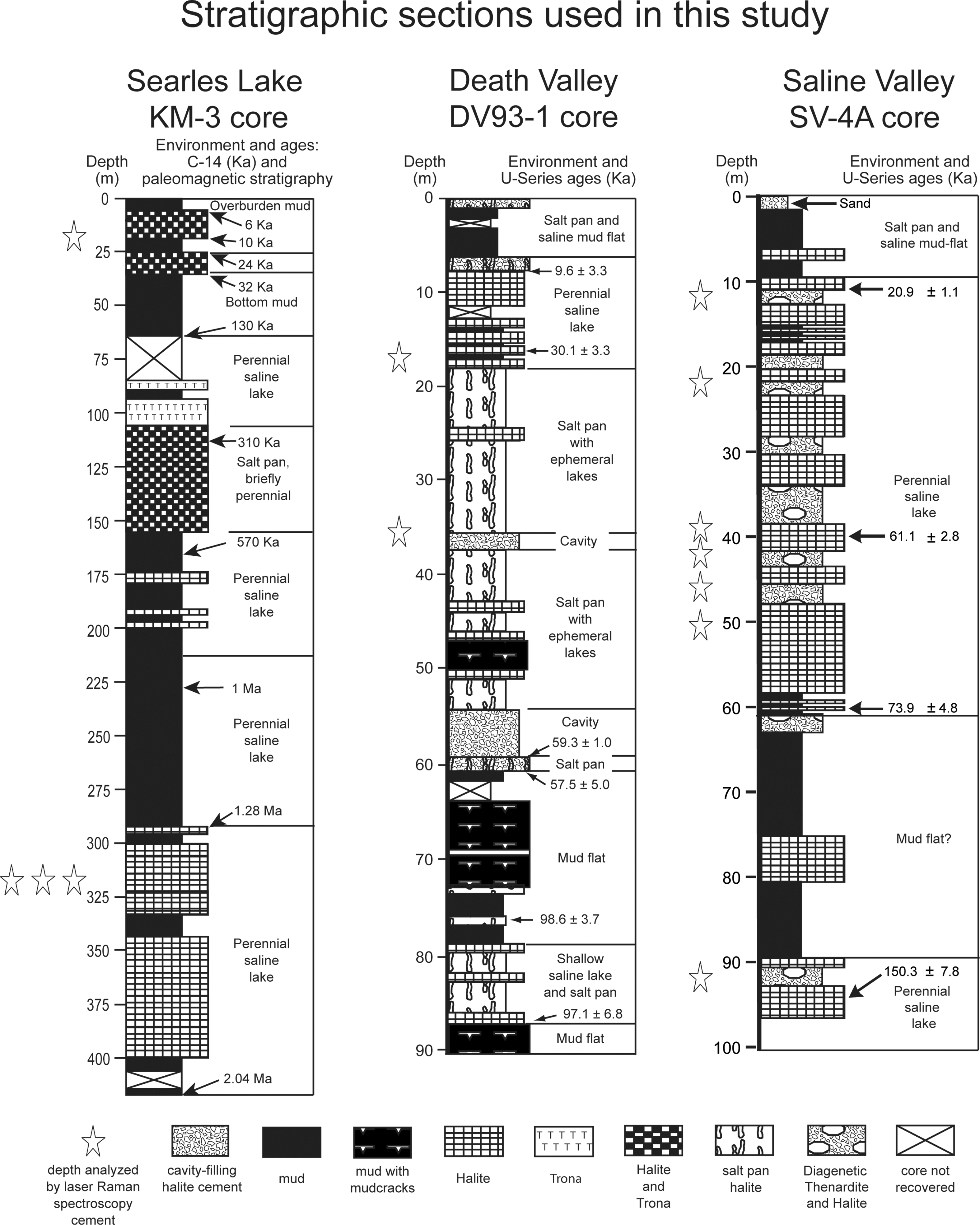
Stratigraphic columns of Searles Lake core KM-3, Death Valley core DV93-1, and Saline Valley core SV-4A, California. Stars indicate depths where ancient carotenoids were targeted for laser Raman spectroscopic analysis. Death Valley and Saline Valley columns are modified from Lowenstein et al. (1999, 2011) and Howe (1998), respectively. Data from Smith et al. (1983) were used to create Searles Lake column. Carotenoids identified from core samples are shown in Table 3.
Cores from Death Valley and Saline Valley consist of interbedded salts and muds deposited in shallow saline pans, deep perennial lakes, and dry mudflats (Lowenstein et al., 1999, 2011). Core KM-3 was similar to DV93-1 and SV-4A; it is composed of halite, trona, and mud, with minor amounts of nahcolite, gaylussite, and pirssonite (Barczak and Petticrew, 1969; Smith et al., 1983). Searles Lake sediments were interpreted by Smith et al. (1983) to have been deposited in perennial saline lakes with shallow-lake and dry intervals. When lake levels were high during the late Pleistocene, Searles Lake was connected to other basins including Panamint Basin and Death Valley.
2.2. Sample materials
Thin sections from each basin were examined with a Zeiss Axio Imager A1 compound light microscope at 10–100×magnification. Transmitted, reflected, and epifluorescence microscopy (with HBO 100 mercury lamp and Chroma Technology Corp filters) were used to study regions of interest in fluid inclusions in halite, which included pigmented masses and bodies resembling algal cells.
2.3. Observations
Ancient algae, remarkably similar in appearance to the modern halophilic alga Dunaliella, were visible within fluid inclusions in halite in at least three life stages, including vegetative pyriform (Fig. 2O), aplanospore or cyst (Fig. 2F), and palmelloid (Fig. 2J). Although modern Dunaliella display organelles and structures (pyrenoid, chloroplast, cytoplasm, stigma/eyespot, and flagella, Fig. 2A–2D) when examined in situ, such features were more difficult to recognize in ancient cells. A cell membrane (Fig. 2G), cytoplasm (Fig. 2E), and a chloroplast (Fig. 2F) were, however, identified in algae in ancient fluid inclusions (30–150 ka). Chloroplasts were previously identified in algae trapped in ancient fluid inclusions (22–100 ka) from Death Valley, California (Schubert et al., 2010). These cells and structures, as well as extracellular pigments, were photographed with a Zeiss AxioCam MRm black and white camera under 10×, 50×, 100×, and a 100×oil immersion objective. Photographs were formatted in Zeiss AxioVision software (version 4.8).
β-carotene, lycopene, and lutein (Fig. 5) were purchased in powdered form (Sigma-Aldrich) as standards to compare with Raman spectra from pigments found in ancient halite. These carotenoids were selected as standards because they are widespread among microorganisms that live in hypersaline, sun-soaked environments (Tables 1 and 2; Goodwin, 1980; Rowan, 1989) and are likely to be trapped with brines in fluid inclusions. Laser Raman spectroscopy was used to analyze β-carotene, lycopene, and lutein, as well as the natural biological materials trapped in fluid inclusions in halite, discussed previously.
Chemical formulas and structures of the three standards: β-carotene, lycopene, and xanthophyll/lutein.
2.4. Instrumental
The spectrometer used was a JY Horiba LabRam high-resolution Raman microprobe connected to an Olympus microscope at Virginia Tech. The microprobe was equipped with a laser that had an excitation wavelength of 514.5 nm (600 mm−1 grating). Spectra were acquired over the wavenumber range 400–3500 cm−1 with a spectral resolution of 1.99 cm−1. Laser power was set to 50 mW, and three scans were accumulated for all samples and standards. Spectra were created with LabSpec software. A 100× objective (∼1 micron spot size) was used for all standards with an exposure time of 5 s. Raman data were collected from natural biological materials trapped in fluid inclusions in halite with the use of a 10×, 50×, or 100× objective. The laser was directed at pigmented areas or objects that commonly resembled the halophilic alga Dunaliella (Fig. 2E–2F, 2H, and 2O). Laser spot size and exposure time (1–15 s) were adjusted to optimize the Raman signal without causing thermal degradation to the samples. To avoid data influenced by thermal degradation, that is, peak shifts, each sample was examined before and after laser analysis. If visible damage to the pigment was observed, such as a change in color or structure, the spectrum was not reported. One measurement was obtained per sample because multiple analyses, in some cases, caused the intensity of the spectrum to weaken, indicating damage to the sample. Spectral analyses from samples of various ages and burial depths were compared to examine carotenoid persistence over periods of more than one million years.
3. Results
Carotenoids produce three distinct peaks in the Raman spectrum when exposed to visible laser excitation (Gill et al., 1970; Rimai et al., 1973; Merlin, 1985; Schulz et al., 2005; Vítek et al., 2009). These bands appear between 1000–1020 cm−1 (v 3), 1150–1170 cm−1 (v 2), and 1500–1550 cm−1 (v 1) (Fig. 6). The v 3 band is of medium strength, created by in-phase rocking movement of methyl groups connected to the polyene chain backbone. v 1 and v 2 wavenumber positions are strong bands caused by in-plane stretching vibrations of C=C and C–C bonds, respectively. The excitation wavelength (514.5 nm) used in this study coincides with the resonance region for carotenoids, which produces an enhanced Raman signal.
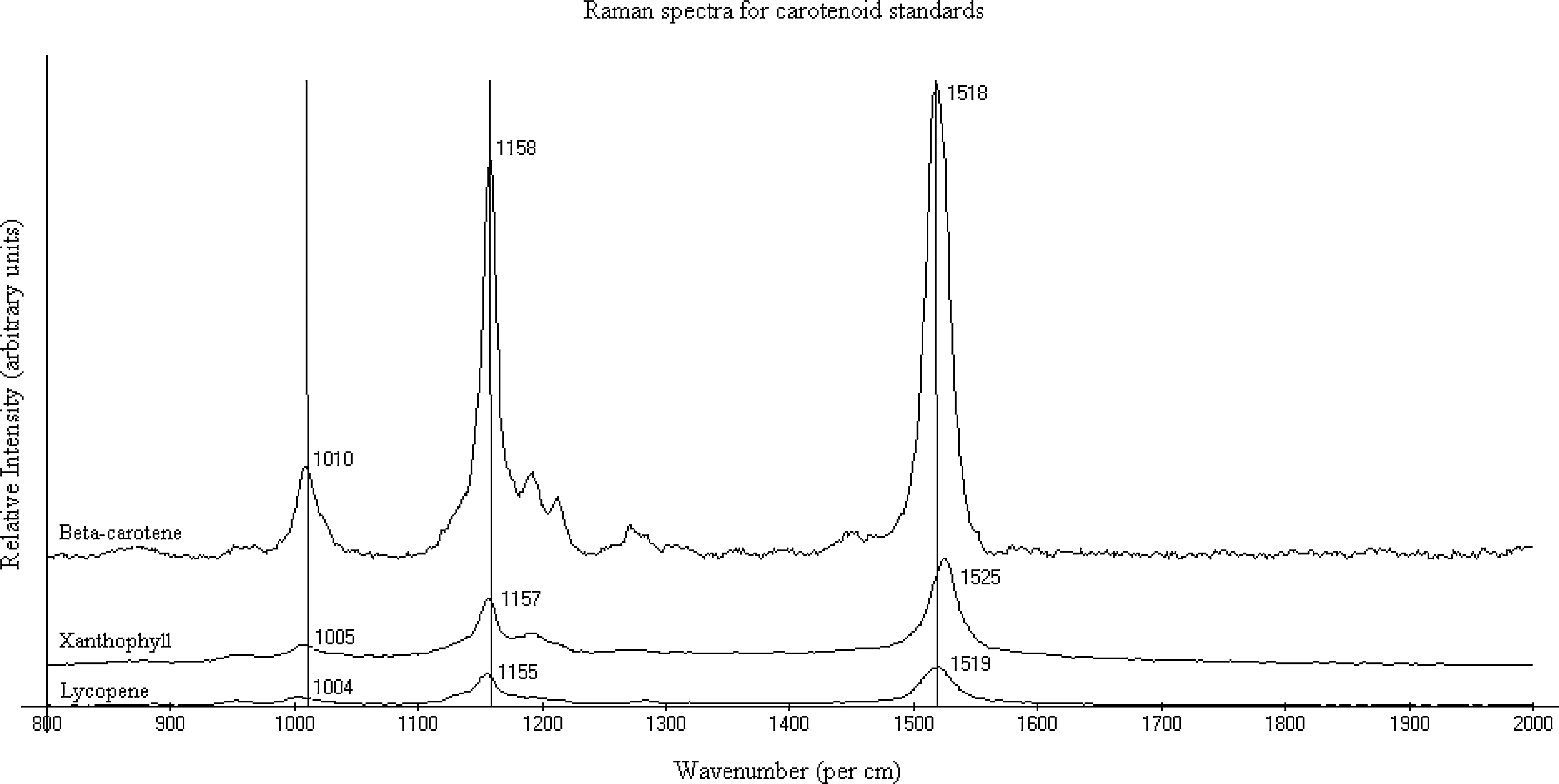
Stacked Raman spectra of carotenoid standards acquired with a 514.5 nm excitation wavelength. Instrument specifications included 50 mW of power, 5 s of exposure time, and a 100×objective.
The spectral region, 1000–1550 cm−1, was studied for lycopene, xanthophyll/lutein, and β-carotene standards. Standards were chosen because of their abundance in the microbial communities of hypersaline environments (Tables 1 and 2). Lycopene produced peaks at 1004, 1155, and 1519 cm−1; xanthophyll/lutein at 1005, 1157, and 1525 cm−1; and β-carotene at 1010, 1158, and 1518 cm−1 (Fig. 6, Table 3). Raman spectra of ancient samples were examined and compared to the characteristic carotenoid peaks for lycopene, xanthophyll/lutein, and β-carotene.
Yellow, orange, red, and brown pigments, which are commonly associated with Dunaliella-like cells in fluid inclusions, all yielded carotenoid signatures from Raman analyses. Carotenoids were detected in all 46 samples from 13 depths in ancient halite from Death Valley, Saline Valley, and Searles Lake, ranging in age from ∼9 ka to 1.44 Ma. (Fig. 4, Table 3). It should be noted that cells pigmented with green chlorophyll created very low signal-to-noise ratios due to strong fluorescence produced by the laser during analyses; this fluorescence overwhelmed peaks needed for biomarker identification.
The number of double bonds in the main chain of each carotenoid was determined by the v 1 peak position; more double bonds result in a lower wavenumber (Withnall et al., 2003). Ancient samples were dominated by 9 or 11 conjugated double bonds in the main chains. These could include β-carotene (9), α-carotene (9), zeaxanthin (9), xanthophyll (9), and lycopene (11). Hydrocarbons with fewer conjugated double bonds, such as phytoene (3), phytofluene (5), and ζ-carotene (7), were not present.
3.1. Standards
Lycopene (1004, 1155, and 1519 cm−1), xanthophyll (1005, 1157, and 1525 cm−1), and β-carotene (1010, 1158, and 1518 cm−1) have similar peak positions (Fig. 6). Given a spectral resolution of 1.99 cm−1, the following band ranges overlap: v 3 for lycopene and xanthophyll; v 2 for lycopene, xanthophyll, and β-carotene; v 1 for lycopene and β-carotene. These shared band values complicate unambiguous carotenoid assignments for unknown compounds.
3.2. Death Valley
Nine samples from two depths, 17 m (30 ka) and 36 m (43 ka), were examined for carotenoids in the Death Valley core (Table 3). All five samples from 17 m produced identical v 1 (1523 cm−1) and v 2 (1160 cm−1) peak values, whereas the v 3 band was 1010, 1011, or 1013 cm−1. The four samples from 36 m had consistent v 3 (1010 cm−1) peak values and narrow peak ranges for v 2 (1158–1160 cm−1) and v 1 (1523–1525 cm−1) bands (Fig. 7). With a spectral resolution of 1.99 cm−1, it is likely that the instrument detected the same carotenoid in all samples from 36 m. It is also possible that all the carotenoids identified from Death Valley, 30 and 43 ka, are the same.
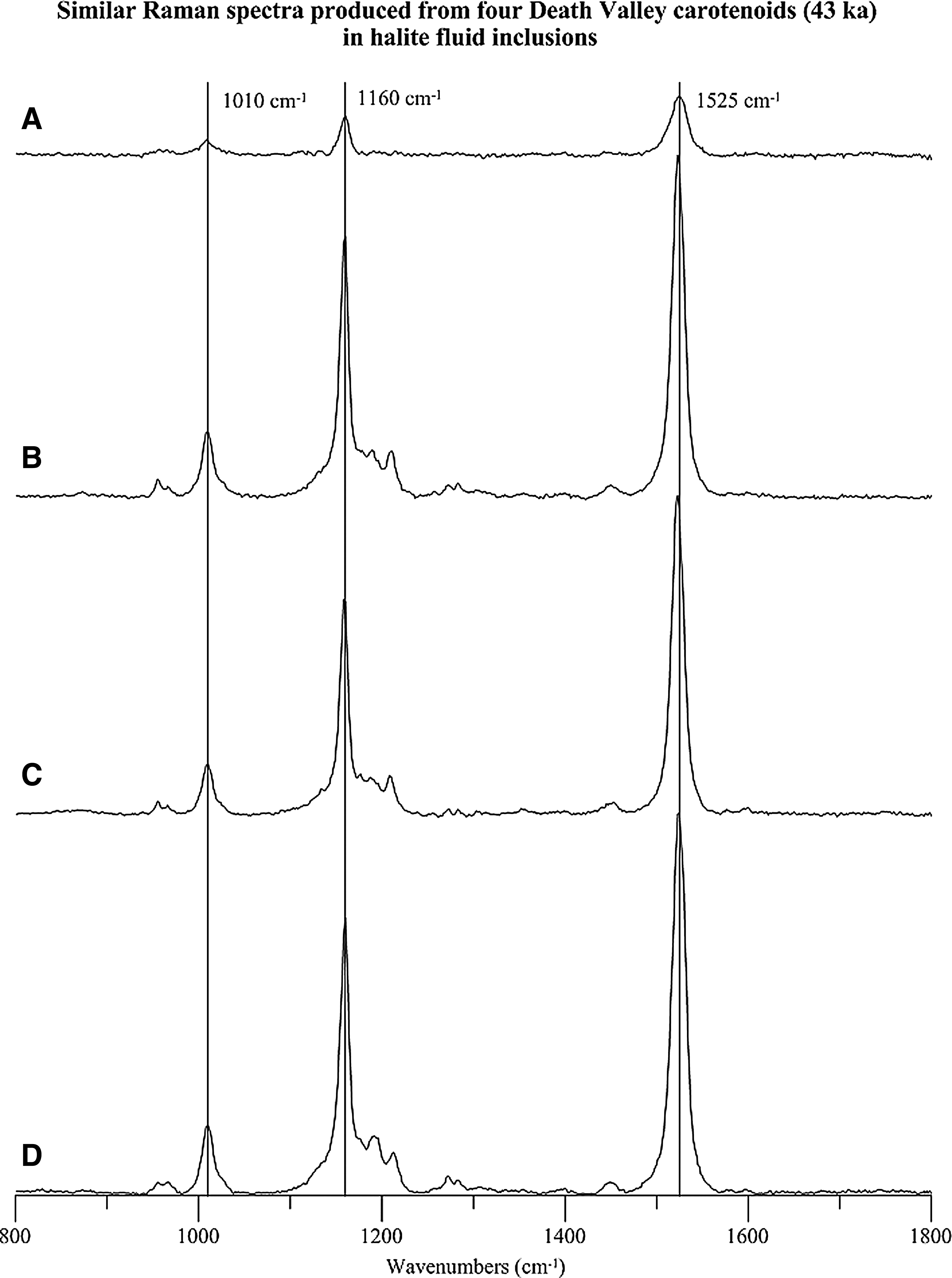
Stacked Raman spectra of ancient carotenoids from Death Valley, California, acquired with a 514.5 nm excitation wavelength. All spectra were produced from carotenoids (43 ka) in fluid inclusions 36 m below the surface. (
Despite the similarities in peak positions, the physical appearances of the Death Valley carotenoids were quite variable. For instance, sample E (Fig. 2G) has orange globules attached to an algal membrane, whereas poorly preserved cell boundaries occur in sample D (17 m).
3.3. Saline Valley
Carotenoids in fluid inclusions from Saline Valley came from halite deposited in perennial saline lakes, ranging in age from ∼21 to 150 ka. v 1 and v 2 bands in 17 of the 19 samples from Saline Valley closely resemble those in Death Valley (Table 3). However, the v 3 band position was shifted slightly from 1010–1013 cm−1 in Death Valley samples to a lower wavelength range of 1007–1010 cm−1 in Saline Valley. Nearly all spectral differences in carotenoids analyzed from Saline Valley halite (21–150 ka) were below the spectral resolution of the instrument, except for Raman band positions in samples I and K from 21 m (35 ka) (Fig. 8). Those samples had lower peak positions (1004, 1155, and 1509–1511 cm−1) for all three bands than any other carotenoid from Death Valley or Saline Valley (Table 3); it is likely that the Raman signal from samples I and K reflects a different compound.

Stacked Raman spectra of ancient carotenoids from Saline Valley, California, acquired with a 514.5 nm excitation wavelength. (
3.4. Searles Lake
Carotenoid peak positions were most variable in Searles Lake samples (Fig. 9). Band values ranged from 1000 to 1015 cm−1 (v 3), 1153 to 1160 cm−1 (v 2), and 1509 to 1523 cm−1 (v 1). Similar to Saline Valley samples I and K, the 10 youngest (9 ka) Searles Lake samples had bands with low wavenumbers, 1000–1011 cm−1 (v 3), 1153–1160 cm−1 (v 2), and 1509–1515 cm−1 (v 1). The low wavenumber values of the v 1 bands indicate that these carotenoids had 11 conjugated double bonds (Table 3). Nearly all other carotenoids from Death Valley, Saline Valley, and Searles Lake had structures with nine conjugated double bonds.

Stacked Raman spectra of ancient carotenoids from Searles Lake, California, acquired with a 514.5 nm excitation wavelength. These spectra show some of the variability in band positions found in Searles Lake carotenoids, which was not as prevalent in Death Valley or Saline Valley. (
Older carotenoids from Searles Lake (1.42–1.44 Ma) had higher band values, 1008–1015 (v 3), 1158–1160 (v 2), and 1517–1523 cm−1 (v 1), which more closely resembled peaks from (21–150 ka) Death Valley and Saline Valley samples. The carotenoids identified from Searles Lake were well preserved and visibly associated with algae in several life stages (Fig. 2I–2L and 2O). Green and red-toned spherical shapes, 10–20 microns in diameter, resemble bodies of the alga Dunaliella in various life stages. For instance, Fig. 2J shows mucilage-covered green algae in the palmelloid stage, whereas Fig. 2O shows a clear/yellowish vacant algal membrane whose shape is similar to the pyriform stage, and Fig. 2K displays orange carotenoids with flecks of green in cystlike bodies.
Carotenoids were identified in core KM-3 up to 1.44 Ma, the oldest confirmed carotenoids found in fluid inclusions in halite to date. They were identified by carotenoid signatures 1015, 1158, 1523 cm−1 and 1011, 1160, 1522 cm−1 in samples A (a) and A (b), respectively (Table 3).
4. Discussion
4.1. Carotenoids are associated with algal cells in ancient fluid inclusions
Examination by laser Raman spectroscopy identified carotenoids, which are known products of halophilic algae, in halite from Death Valley, Saline Valley, and Searles Lake, California. Carotenoids appeared in a variety of forms in modern and ancient algal cells; some occurred within algal-shaped bodies (Fig. 2K), whereas others were external to algal bodies, that is, expelled from, but still attached to, the membrane, or independent of an algal membrane (Fig. 2H). Carotenoids commonly filled entire cells with pigment, whereas extracellular carotenoids appeared as globular (Fig. 2G), crystalline (Fig. 2L) or noncrystalline (Fig. 2N) masses. There was no correlation observed between Raman band sets (bands v 1, v 2, and v 3), physical characteristics, and the age of carotenoids preserved in these fluid inclusions.
Other studies have reported that Raman bands are influenced by molecules surrounding carotenoids. Certain biological material, for example, proteins and membrane lipids, affect the chemical characteristics of carotenoids and therefore shift Raman peaks (de Oliveira et al., 2010; Jehlička et al., 2013). Microscopic examination of samples showed that carotenoids from Death Valley, Saline Valley, and Searles Lake were often attached to algal bodies or unknown biomolecules. This may explain why peak positions from the reference standards lycopene, xanthophyll, and β-carotene were commonly not the same as the Raman bands from many natural carotenoids.
4.2. Identification of specific carotenoids proved challenging
Specific carotenoids were difficult to identify with certainty. The band set (1011, 1160, 1520 cm−1) from Saline Valley sample A (60 ka), for example, matched the band set (1010, 1158, 1518 cm−1) for β-carotene, given a spectral resolution of 2 cm−1. But the v 1 band of sample A also overlaps the v 1 band range of lycopene (1517–1521 cm−1). This overlap did not allow the unambiguous assignment of sample A as β-carotene. The same complication exists for samples G, I, and J (1.42 Ma) from Searles Lake.
Overlapping band ranges were also observed in Saline Valley. The peak positions (1007, 1158, 1523–1525 cm−1) for Saline Valley samples E (150 ka) and B (a) (67 ka) are consistent with xanthophyll (1005, 1157, 1525 cm−1), but the v 2 band also coincided with β-carotene (1156–1160 cm−1). Therefore, samples E and B (a) cannot be identified as xanthophyll with absolute certainty. Although each carotenoid has a unique physical structure, they are all modified from a parent structure (Britton, 1995). Raman spectroscopy allows for detection of structural subtleties, but the spectral resolution of an instrument may be larger than band value differences between structurally similar carotenoids. This can cause ambiguity in identifying specific carotenoids. Increasing the instrument's precision allows detection of narrower band ranges, which decreases the likelihood of overlapping bands.
4.3. Diverse carotenoids should exist in ancient fluid inclusions
Halotolerant and halophilic algae contain a variety of carotenoids. For instance, one algal species, Dunaliella sp. (Euchlorophyceae, Volvocales), produces seven carotenoids (Table 2); and the class Chlorophyceae, of which Dunaliella salina is a member, produces 17 carotenoids (Table 1). Algae generate carotenoids in response to changing light, temperature, and seasonal stress (Abd El-Baky et al., 2004). Carotenogenesis can increase when cells are deprived of nitrogen as the surrounding brine becomes more concentrated (salt stress), or if cells are exposed to UVB radiation (high light intensity). A community of algae may create a variety of carotenoids, depending on their taxon and environmental history.
Although overlapping band values made the unambiguous assignment of specific carotenoids impossible, at least two carotenoid structures (9 and 11 conjugated double bond lengths) existed among the 46 samples. Carotenoids with nine or more conjugated double bonds are more effective at suppressing damaging singlet oxygen (Rodriguez-Amaya, 2001). Since closed-basin saline lakes receive intense sunlight, microbes there are expected to produce carotenoids with nine or more double bonds for protection from UV radiation.
We speculate that more than one type of alga was common in these ancient saline basins; however, it was not possible to prove that with Raman spectroscopy because carotenoids from different samples could have been produced from different algae or the same algal species in two stress states.
In addition, when analyzing algae such as Dunaliella that produce multiple carotenoids in varying concentrations, a Raman spectrum may only represent the carotenoid that is most effective at scattering light at that particular excitation wavelength (de Oliveira et al., 2010). Therefore, we could not conclude here that the Raman spectrum bands from a given sample (algal cell) identified the most concentrated carotenoid, nor could we determine whether the bands represented one or more carotenoids. A thorough database with information about the type and concentration of carotenoids in algae and other microorganisms, as well as the Raman spectra at all appropriate excitation wavelengths, would be invaluable to resolve some of these questions about Raman spectroscopy in natural biological systems.
4.4. Laser Raman spectroscopy and extraterrestrial life
Fluid inclusions in evaporites preserve microscopic ecosystems from saline environments (Schubert et al., 2009b; Lowenstein et al., 2011). The oxygen-free, light-free environment of buried salt is an ideal setting for preservation of biomarkers, such as DNA and pigments. The same techniques described here to identify carotenoids in halite may be used to search for microscopic life in evaporites on Mars (Squyres et al., 2004; Wang et al., 2006). Carotenoids are ubiquitous in halophilic communities on Earth, and they may also have been produced for UV protection by life on Mars (Vítek et al., 2009). Carotenoids are indisputable evidence of life. β-carotene was selected for the ExoMars mission of the Aurora program as an indicator compound in the search for ancient life on Mars (Vago et al., 2006). This study showed that carotenoid detection by way of laser Raman spectroscopy should be applicable to ancient evaporites on Mars. The longevity of carotenoid preservation in fluid inclusions on Earth is unknown, but results presented here indicate >106 years.
The search for extraterrestrial life should target chloride-bearing salts, for example, halite found on Mars (Osterloo et al., 2008) and, in particular, layered crystalline salts with preserved growth textures and fine-scale layering. Carotenoids are easily detectable in such salt samples with Raman spectroscopy. As demonstrated here, carotenoids produce strong Raman signals, even in complex, heterogeneous ecosystems. Halite provides an excellent host matrix because it does not interfere with Raman signals; it provides a zero-background for spectra (Pasteris et al., 1988). Gypsum (CaSO4·2H2O) may also be a suitable salt to analyze for carotenoids. The v 1 band in gypsum, which is caused by the SO4 symmetric stretching mode, is observed in the same region as the medium v 3 band in carotenoids (White, 2009), but the two stronger carotenoid bands (v 1 and v 2) are most important for carotenoid identification. Laser Raman spectroscopy could provide invaluable information about microscopic life on Mars as exploration progresses.
5. Conclusions
Laser Raman spectroscopy is an important technique for studying fluid inclusion ecosystems in ancient halite from Death Valley, Saline Valley, and Searles Lake, California. Colorful pigments associated with single-celled algae were identified as carotenoids; however, specific carotenoids could not be definitively identified. This research showed that carotenoids are reliable biomarkers; they appear to be stable in the O2-free, light-free, saline environment of a fluid inclusion in the subsurface. The intensity of Raman spectra from carotenoids in fluid inclusions did not decrease with age (21 ka to 1.44 Ma). This research suggests that fluid inclusions in extraterrestrial evaporites would be a possible habitat to explore for microbial life.
Footnotes
Acknowledgments
We thank Dr. Robert Bodnar and Charles Farley for their Raman expertise and for accommodating the use of their Vibrational Spectroscopy Laboratory in the Geosciences Department of Virginia Polytechnic Institute and State University. We greatly appreciate the advice of Dr. Jan Jehlička on our manuscript and the thoughtful reviews of Jan Jehlička, Kathy Benison, and an anonymous reviewer. Funding for this research came from NSF Geobiology and Low Temperature Geochemistry Award EAR-1024692.
Author Disclosure Statement
No competing financial interests exist for Yaicha Winters, Tim Lowenstein, and Michael Timofeeff.