
Abstract
The search for organic biosignatures is motivated by the hope of understanding the conditions of emergence of life on Earth and the perspective of finding traces of extinct life in martian sediments. Paramagnetic radicals, which exist naturally in amorphous carbonaceous matter fossilized in Precambrian cherts, were used as local structural probes and studied by electron paramagnetic resonance (EPR) spectroscopy. The nuclear magnetic resonance transitions of elements inside and around these radicals were detected by monitoring the nuclear modulations of electron spin echo in pulsed EPR. We found that the carbonaceous matter of fossilized microorganisms with age up to 3.5 billion years gives specific nuclear magnetic signatures of hydrogen (1H), carbon (13C), and phosphorus (31P) nuclei. We observed that these potential biosignatures of extinct life are found neither in the carbonaceous matter of carbonaceous meteorites (4.56 billion years), the most ancient objects of the Solar System, nor in any carbonaceous matter resulting from carbonization of organic and bioorganic precursors. These results indicate that these nuclear signatures are sensitive to thermal episodes and can be used for Archean cherts with metamorphism not higher than the greenschist facies. Key Words: Kerogen—Biosignatures—Origin of life—Archean—EPR spectroscopy. Astrobiology 13, 932–947.
1. Introduction
Even in a chert containing biogenic CM, a part of this material may originate from various contamination processes such as hydrothermalism, endolithic microorganisms, weathering, and anthropogenic contamination (Hoering, 1967; Roedder, 1981; Campbell, 1982; Westall and Folk, 2003). Consequently, the detection method must be able to distinguish primitive biosignatures with the same age as the host rock from more recent biosignatures due to contamination.
In addition, the presence of carbonaceous microstructures in a sedimentary rock is not sufficient to assess that primitive life was present at the epoch of the sediment deposit, because the CM may also have an abiotic origin (Luque et al., 1998; Garcia-Ruiz et al., 2003). As far as very ancient rocks are concerned, a part of this contamination may be of extraterrestrial origin (meteorites, interplanetary dust particles, comets). For example, carbonaceous meteorites, among the most primitive objects of the Solar System (4.5 Gyr), contain a significant amount of CM with chemical composition similar to that of terrestrial kerogens (Hayatsu et al., 1977). Remarkably, this very ancient extraterrestrial CM contains organic molecules of biological importance at the part-per-million level (Cronin et al., 1988). During the early history of the Solar System, Earth and Mars were intensively bombarded by meteorites, micrometeorites, and comets, so that a considerable amount of CM has probably been deposited on these planets (Chyba and Sagan, 1992; Maurette et al., 2006), with the possibility that a fraction was fossilized in sedimentary deposits. This eventuality is of particular concern for the search for traces of organic carbon in the oldest sedimentary rocks on Mars.
The in situ detection of molecular biosignatures in a composite material composed of a silicified rock with embedded amorphous and extremely disordered CM is a real challenge for spectroscopic analysis. For example, the well-known Raman spectroscopy, which gives a typical signature of CM (see, e.g., Quirico et al., 2009; Bower et al., 2013), is not always sufficient to make a clear distinction between biogenic and abiotic CM (Pasteris and Wopenka, 2003; Marshall et al., 2010; Bower et al., 2013). More generally, spectroscopic methods based on electromagnetic radiation in the UV-visible range are limited to the sample surface or subsurface because of the strong optical absorption of most rocks. Spectroscopic methods that work at long wavelengths, such as nuclear magnetic resonance (NMR), are more adapted to a selective detection of molecular biosignatures. In addition to the high resolution of this technique, the weak interaction (dipolar magnetic) of the radiofrequency field with matter allows a deep penetration of the radiation in the sample. Unfortunately, NMR suffers from a low sensitivity and for this reason is not adapted to rocks containing small amounts of CM, sometimes at the part-per-million level. Moreover, NMR spectra are often perturbed by paramagnetic species (transition metal impurities, radicals, etc.) that are always present in natural materials.
Electron paramagnetic resonance (EPR) spectroscopy offers a good compromise between sensitivity (much better than NMR) and resolution (lower than NMR). Its frequency domain in the microwave range allows a deep penetration of the electromagnetic field probe into the sample, so that EPR spectra correspond to the bulk of the material and are much less sensitive to surface alteration and contamination (Binet et al., 2008). All carbonaceous materials, natural or synthetic, contain paramagnetic defects (hereafter referred to as radicals), which can be detected with high sensitivity by EPR spectroscopy. These defects can be described as molecular fragments with unpaired electrons responsible for the EPR response. This is the case for synthetic amorphous carbon (Robertson, 2002), coals (Uebersfeld et al., 1954; Retcofsky et al., 1975; Mrozowski, 1988a, 1988b; Whelan and Thompson-Rizer, 1993), carbonaceous shales (Rigali and Nagy, 1997), cherts (Skrzypczak-Bonduelle et al., 2008; Bourbin et al., 2012), and carbonaceous meteorites (Binet et al., 2002, 2004a; Gourier et al., 2008; Delpoux et al., 2011). However, compared to NMR, standard cw-EPR (continuous wave EPR) suffers from a rather poor resolution about 103 times less than NMR, so that radicals in all kinds of carbonaceous materials always give the same type of EPR signal, mostly in the form of a more or less narrow and symmetrical line characterized by a g-factor (related to the magnetic field value of the line) close to the free electron spin value 2.0023 (Fig. 1) (Uebersfeld et al., 1954; Retcofsky et al., 1975; Mrozowski, 1988a; Whelan and Thompson-Rizer, 1993; Binet et al., 2002; Skrzypczak-Bonduelle et al., 2008). The width of this signal is due to the hyperfine (hf) interactions of the unpaired electron spin of the radical with all neighboring nuclei, giving hundreds of overlapping lines that merge into a single EPR line without apparent structure. This unresolved hf structure, which is expected to contain useful information, can be resolved with pulsed-EPR techniques, whereby nuclear magnetic resonances of various nuclei with nonzero nuclear spin (1H, 13C, 14N, 31P, 29Si,…) located in the vicinity of the unpaired electron can be indirectly detected by recording the nuclear modulation of the electron spin echo (Höfer et al., 1986; Schweiger and Jeschke, 2001). With these pulsed EPR methods, the unpaired electron spins of radicals play the role of internal probes, which transfer the microwave excitation to neighboring nuclear spins at the nanometer scale. The resulting nuclear magnetic excitations are transferred back electron spins and read in the form of modulations of the electron spin echo at nuclear frequencies.
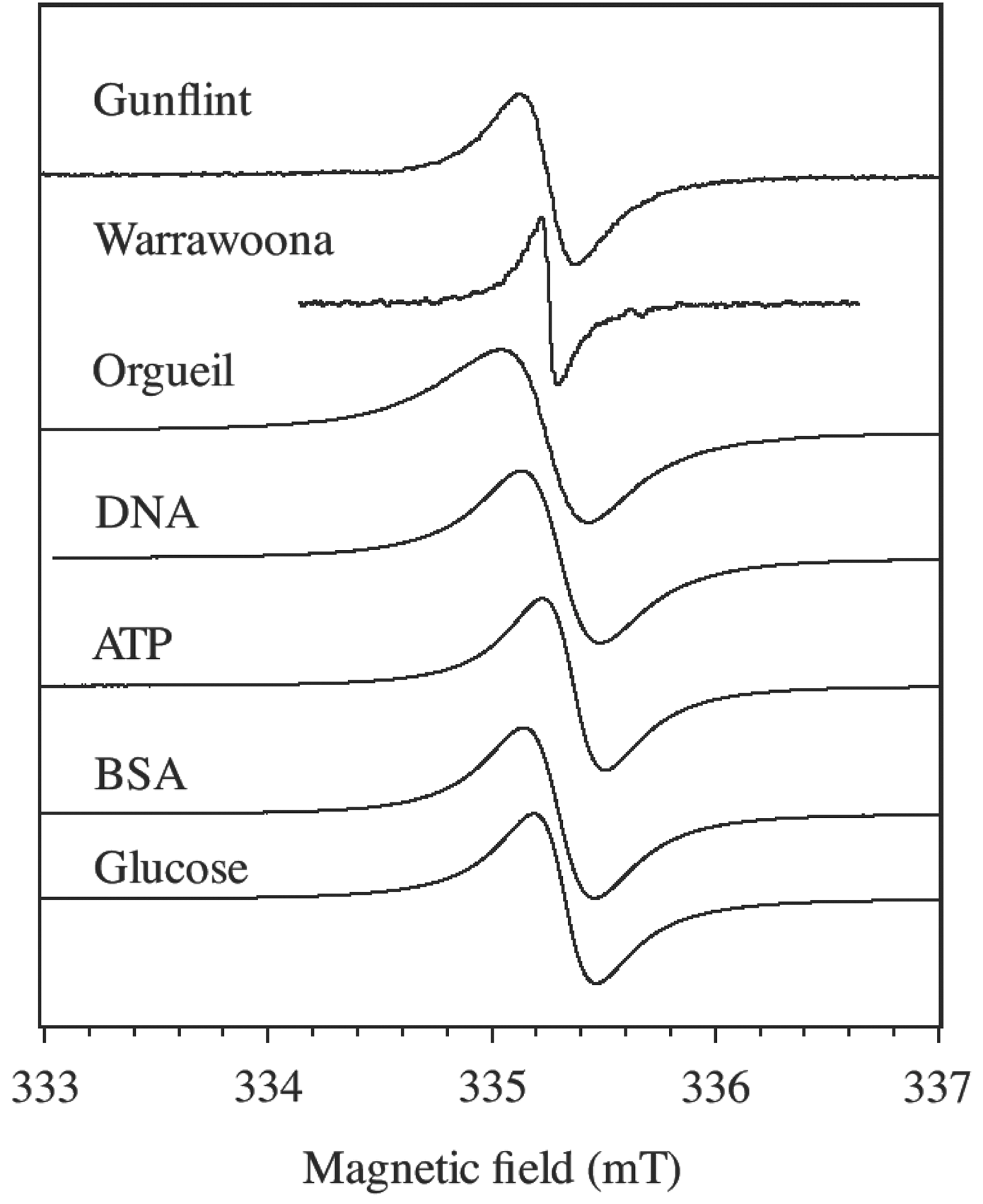
cw-EPR spectra at room temperature of Gunflint (1.9 Gyr) and Warrawoona (3.5 Gyr) cherts, CM extracted from the Orgueil meteorite, and molecules encapsulated in silica and treated at 550°C.
In this work, we used the bi-dimensional version of this electron spin echo technique, known as hyperfine sublevel correlation (HYSCORE) spectroscopy (Höfer et al., 1986), to investigate the nuclear transitions of 1H, 13C, and 31P in the vicinity of organic radicals in cherts with different degrees of evolution that contain fossil CM of biological origin. The magnetic resonance signals were compared with those of the abiotic CM in carbonaceous meteorites. The influence of metamorphism was experimentally studied by monitoring the effect of carbonization on biological molecules encapsulated in silica.
2. Materials and Methods
Three kinds of materials were studied in this work: lightly metamorphosed cherts containing microorganisms fossilized in the form of carbonaceous microstructures (referred to as F-type), CM extracted from two carbonaceous meteorites (referred to as M-type), and CM originating from carbonization of bioorganic molecules encapsulated in silica (referred to as C-type).
2.1. Cherts and meteorites
Three chert samples (silicified sediments) containing carbonaceous remains of microorganisms (F-type) were provided by the Museum National d'Histoire Naturelle (Paris, France). A 3.49-billion-year-old (Archean) chert (PPRG 006 from the Precambrian Paleobiology Research Group Collection, courtesy of W. Schopf) was collected in the Dresser Formation of the Warrawoona group, Pilbara Craton, Western Australia. It hosts among the oldest putative microfossils (Schopf, 1993). A 1.88-billion-year-old (Paleoproterozoic) chert, referenced as GF 74-1-11 from the Awramik collection, comes from the Schreiber Beach locality of the Gunflint Formation (Ontario, Canada) and contains well-preserved prokaryotic microorganisms (Barghoorn and Tyler, 1965). These two cherts are metamorphosed in the greenschist facies. A non-metamorphized 45-million-year-old chert (Eocene) referenced as PPRG 456 from the Schopf collection comes from the Clarno Formation, John Day basin in Oregon (USA), and contains well-preserved remains of vascular plants.
Carbonaceous matter of meteorites (M-type) was extracted from the mineral host of two meteorites by the usual HF/HCl treatment (Durand and Nicaise, 1980), which does not affect the structure and the concentration of radicals (Binet et al., 2002). Samples of Orgueil (France, fallen in 1864) and Tagish Lake (Canada, fallen in 2000) meteorites were provided by the Museum National d'Histoire Naturelle (Paris, France) and the NASA Johnson Space Center (Houston, USA) with the permission of J. Brook (the finder), respectively. The bulk elemental compositions of the insoluble residues of the HF/HCl treatments (the amorphous CM) were determined by elemental analysis at Wolf Laboratories (Rueil-Malmaison, France) for Orgueil and by Atlantic Microlab, Inc. (Norcross, Georgia, USA) for Tagish Lake. The resulting compositions of the CM are C100H72O18N3S2 for Orgueil and C100H46O15N10 for Tagish Lake (Binet et al., 2002, 2004b). The samples still contain inorganic residues that amount to 8.9% and 7.2%, respectively.
2.2. Encapsulation of organic and bioorganic molecules
C-type CM (carbonization of organic molecules) was prepared to study the effect of thermal metamorphism on biological material. Several types of molecules and biomolecules were encapsulated in silica by a three-step sol-gel method (Livage et al., 2001) in order to reproduce the chemical environment in a chert: glucose, adenosine triphosphate (ATP), deoxyribonucleic acids (DNA), and a protein (bovine serum albumin, BSA). The first step of the synthesis is the preparation of aqueous solutions of about 10 mg·mL−1 of biomolecules from Sigma-Aldrich. Two kinds of macrobiomolecules were used: DNA from salmon sperm and BSA in the form of lyophilized powders. For DNA, it was necessary to add a few milliliters of a 0.1 mol·L−1 NaOH solution to the solution of DNA to obtain a neutral pH, which does not hinder the gel formation. The other molecules are anhydrous α-D-glucose (powder) and a solution of the disodium salt of adenosine-5′-triphosphate (ATP). To allow for the gel formation, a 100 mM aqueous solution was adjusted to pH 7 with a Trizma base buffer. The second step consisted of the preparation of the 0.75 mL·L−1 sodium silicate solution with 3.3 g of Na2Si3O7 (27% SiO2,10% NaOH) in 20 mL of H2O. Then 700 μL of HCl 3 mol·L−1 were poured under magnetic stirring in 5 mL of solution of silicate of sodium to reach pH ∼7. Finally, 5 mL of solution containing the solvated biomolecule was poured at once. The gel appeared in a couple of minutes. The last step was the sample dehydration by freeze-drying. The gels were immersed into liquid nitrogen in order to freeze them very quickly and solidify the water contained in these samples. They were then subjected to a pressure of several millibar to sublimate the water, for approximately 50–60 h. This process allowed a progressive dehydration of the samples without altering their structure, giving a final product in the form of white powders.
2.3. EPR experiments
All the experiments were performed at X-band (9.5 GHz). Continuous wave–EPR spectra were recorded at room temperature with a Bruker ELEXSYS E500 spectrometer equipped with a high-sensitivity 4122SHQE/0111 microwave cavity. Pulsed-EPR experiments were performed at temperatures in the range from 4 to 30 K with a Bruker ELEXSYS E580 spectrometer equipped with a helium flow cryostat. The HYSCORE method was used for the detection of 1H, 13C, 31P, and 29Si nuclei. A spin echo is generated by the pulse sequence π/2-τ-π/2-t 1-π-t 2-π/2-τ-echo at the time τ after the last π/2 pulse; π/2 and π represent the flip angles of the electron magnetization. The echo intensity is measured by varying stepwise the times t 1 and t 2 at constant τ. The pulse lengths were 16 and 32 ns for π/2 and π pulses, respectively. For both t 1 and t 2 domains, 256×256 data points were measured. A second-order polynomial background subtraction was used to remove the unmodulated part of the echo. The magnitude spectrum was obtained after a 2-D Fourier transform of the spectra by using a Hamming apodization window function. The final 2-D spectrum represents the correlations between nuclear frequencies corresponding to the two spin states (up and down) of the electron and gives the nuclear frequencies and the hyperfine interactions of nuclei in the vicinity of the unpaired electron spin (Höfer et al., 1986). We always checked that the HYSCORE spectra correspond to the probed cw-EPR lines by recording three-pulse echo-detected EPR spectra (π/2-τ-π/2-T-π/2-τ-echo), which represent the echo intensity versus the magnetic field strength.
The samples were prepared by introducing several hundreds of milligrams of material in 4 mm quartz tubes. In the case of bioorganic molecules encapsulated in silica (C-type), the tubes were then maintained in a vacuum (10−2 bar) for at least 4 h and sealed to avoid oxidation of powders during thermal treatments and the effect of the paramagnetism of molecular oxygen on the EPR signal of radicals. This encapsulated organic matter was carbonized by step heating treatments of the sealed tubes during short times (15 min) with temperature increasing by 50°C at each step. The cw-EPR spectrum was recorded at room temperature after each thermal step. The thermal step at 550°C gives an EPR line with almost the same lineshape (Lorentzian) and the same linewidth as the 1.9-billion-year-old Gunflint chert (Skrzypczak-Bonduelle et al., 2008).
3. Results
3.1. cw-EPR of carbonaceous matter
The three types of CM (F, M, and C) give a cw-EPR signal at 9.4 GHz consisting of a single narrow line representing the first derivative of the microwave absorption (Fig. 1). This signal is due to polyaromatic fragments containing unpaired electrons with spin S=1/2 delocalized in 2p π orbitals of the sp2 carbon backbone (Binet et al., 2002; Robertson, 2002). Such single EPR lines are ubiquitous in carbonaceous materials, including nanographene and nanographite (Joly et al., 2010; Barbon and Brustolon, 2012). The magnetic moments may originate from defects or from localized states in the edge of the polyaromatic fragments (Enoki and Kobayashi, 2005). The CM of coals, meteorites, and non-metamorphosed (or slightly metamorphosed) cherts can be viewed as a network of polyaromatic (sp2 carbons) fragments linked together by more or less complex diamagnetic (S=0) aliphatic chains (sp3 carbons). Only a very small fraction of the polyaromatic units contain unpaired electron spins and are thus detected by EPR.
In addition to the narrow EPR line of CM, cherts and meteorite samples studied in this work contain other signals that have been reported in previous papers (Binet et al., 2002; Skrzypczak-Bonduelle et al., 2008). Meteoritic materials exhibit a broad ferromagnetic resonance signal due to magnetite and other residual magnetic minerals (Binet et al., 2002). However, this ferromagnetic resonance signal is so broad that it only contributes to a slight deviation of the baseline of the narrow CM line. The three chert samples exhibit other paramagnetic signals close to the CM line. In addition to a weak Fe3+ signal at g≈4.2, we always observed E′-center of quartz (an electron trapped at a dangling sp3 orbital of
The two EPR parameters of importance in single line spectra are the peak-to-peak linewidth ΔB pp and the g-factor, defined as g=hν/βB res, where h is Planck's constant, ν the microwave frequency, β the electron Bohr magneton, and B res the magnetic field at the center of the resonance line. These parameters are gathered in Table 1 for the samples studied in this work. The parameters for the molecules encapsulated in silica are given in Table 1 for the thermal step 550°C, a temperature for which the EPR lines are very similar to that of Gunflint chert. In addition to ΔB pp and g, the EPR lineshape of radicals is an interesting proxy that depends on the age of the CM and thus provides a test for syngenicity of the organic matter in Archean cherts (Bourbin et al., 2013). It may be argued that the slight distortion of some EPR lines in Fig. 1 could reflect a Dysonian profile (Barbon and Brustolon, 2012) resulting from electronic conductivity. A Dysonian line being homogeneous, however, it cannot give spin echoes in contrast with the experimental results presented below.
The linewidth ΔB pp reflects the broadening of the EPR line by unresolved hf interaction with hydrogen (1H) nuclei and by the dipolar interaction between radicals (Skrzypczak-Bonduelle et al., 2008; Bourbin et al., 2012). This can be seen by artificial aging, in which the evolution of ΔB pp is monitored during isochronal thermal treatments. Figure 2a shows the results for the different precursors (BSA, DNA, ATP, glucose) encapsulated in SiO2 and the values measured for the 45-million-year-old Clarno chert treated under the same thermal conditions (interrupted line). The linewidth decrease is due to the loss of hydrogen atoms that occurred during the polymerization of the CM (Skrzypczak-Bonduelle et al., 2008). It appears that the carbonization of molecules encapsulated in SiO2 follows the same evolution of ΔB pp as a young chert containing non-mature CM (Clarno in Fig. 2a). The linewidths measured for Gunflint (0.23 mT, G in Fig. 2a) and Warrawoona (0.12 mT, W in Fig. 2a) are the same as those for Clarno and organic molecules heat treated at ∼550°C and ∼700°C, respectively.
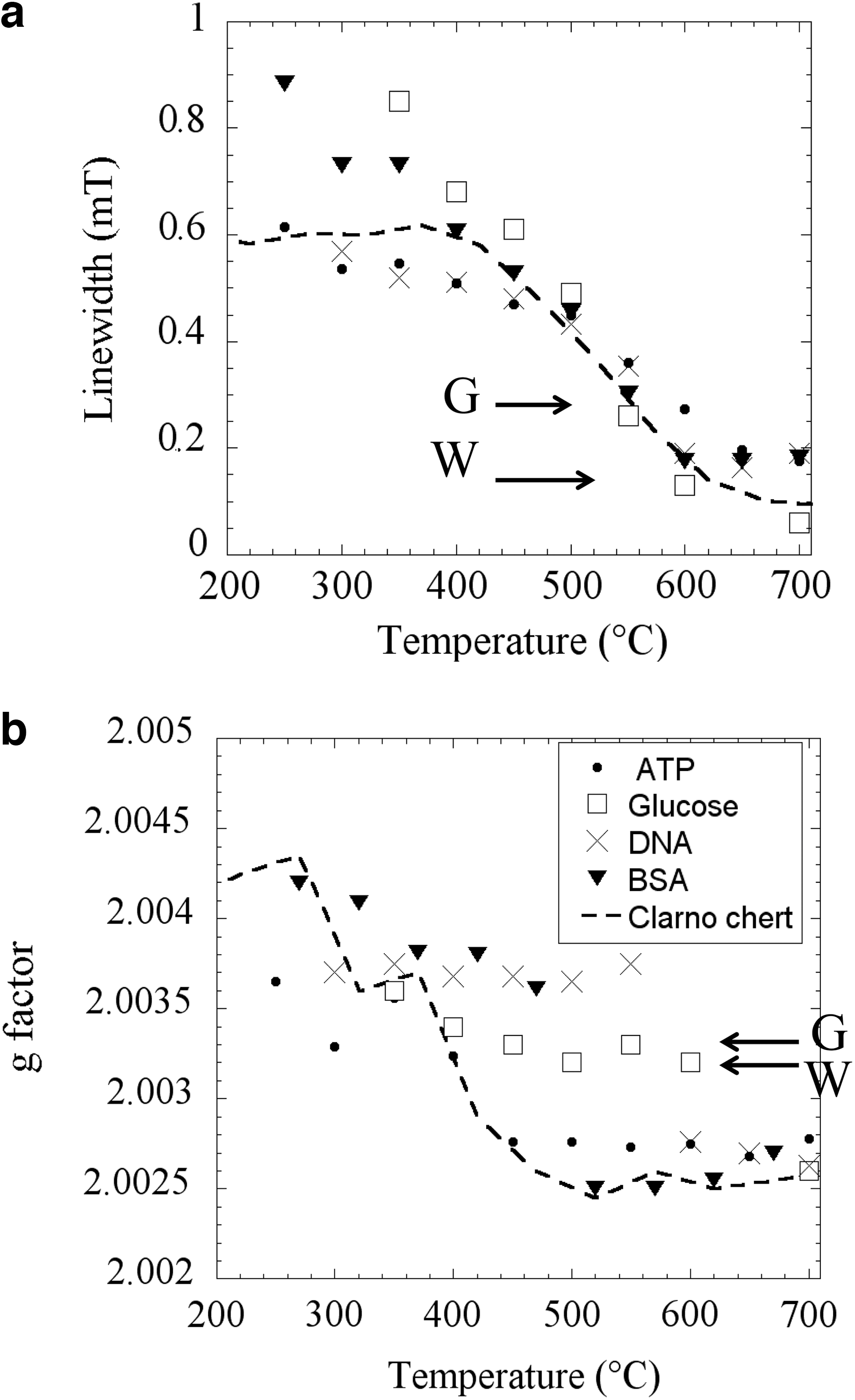
Variation of EPR parameters with thermal treatment temperature for molecules encapsulated in silica. The discontinuous line represents the evolution for the Clarno chert (45 Myr). (
Differences between thermally treated cherts and carbonized molecules encapsulated in SiO2 are more pronounced in the evolution of g-factor (Fig. 2b). The deviation of g from the free-spin g-value ge =2.0023 reflects the presence of heteroelements heavier than carbon (mainly oxygen) in the vicinity of the electron spin. The decrease of the g-factor during carbonization is due to oxygen loss in the form of H2O and CO2. It appears that the thermal evolution found for Clarno is close to that of BSA and ATP but very different from that of glucose and DNA, except at the end of the thermal treatment. This suggests that proteins may have dominated the biomolecular precursors at the origin of the CM fossilized in Clarno. Arrows in Fig. 2b indicate the g-factors for Gunflint (G) and Warrawoona (W) cherts. Interestingly, these values are close to those measured for the CM originating from glucose precursor, which may indicate that carbohydrates could be the main precursors at the origin of the CM of Precambrian cherts.
This analysis of the evolution of resonance lines by cw-EPR can be also extended to the lineshape, which evolves from a Lorentzian line toward a stretched Lorentzian line upon natural and artificial aging (Skrzypczak-Bonduelle et al., 2008; Bourbin et al., 2013). This joined evolution of the g-factor, ΔB pp, and the lineshape upon natural and accelerated aging clearly indicates that the EPR of radicals in fossil CM would be strongly influenced by any thermal event that occurs during metamorphic episodes subsequent to the formation of the sediment. However, a detailed study of the EPR lineshape (Bourbin et al., 2013) shows that the evolution of radicals in fossil CM is controlled by metamorphism for cherts younger than 2 Gyr, while it is controlled by the age for cherts older than 2 Gyr with metamorphic grade not higher than the greenschist facies.
The following part of this paper focuses on the analysis of the unresolved hf structure of the EPR line, which contributes to the EPR linewidth. It is partly due to the interaction of the radical electron spin with various nuclei (1H, 13C and 31P, 29Si,…) inside or around these radicals. The hf interactions being controlled by the chemical bonding and the structure of radicals in the CM, these interactions should be much more sensitive to the origin of the molecular precursors than the g-factor, the linewidth, and the lineshape.
3.2. Pulsed EPR of cherts and meteorites
Nuclear modulation effects of spin echo amplitude in pulsed EPR spectroscopy give a direct measurement of the hf interactions between the unpaired electron spins of the aromatic radicals and various nuclear spins inside or in the vicinity of the radicals. Typical HYSCORE spectra are shown in Fig. 3 for Clarno (45 Myr), Gunflint (1.9 Gyr), and Orgueil and Tagish Lake meteorites, plotted as the intensity versus radio frequencies ν 1 and ν 2 (in MHz). These spectra are in the range of nuclear frequencies, as usually measured in NMR spectroscopy. For the sake of comparison, all the spectra of Fig. 3 are recorded with the same interval τ=136 ns between the two first π/2 pulses. This condition is a good compromise to detect the hyperfine spectrum of both 1H and 13C in the same spectrum. In principle, the shape of the 1H-hyperfine spectrum depends on τ by the so-called “blind spot” effect, and low τ are generally preferred to avoid lineshape distortion (Schweiger and Jeschke, 2001). However, we did not observe significant blind-spot effects by varying τ, and as shown below, the most important information was obtained by using high τ values. Despite the fact that the spectrum of the Tagish Lake meteorite suffers from a low signal-to-noise ratio, it is clearly of the same type as that of the Orgueil meteorite. It appears that all the HYSCORE spectra are similar to those found for coals (Ikoma et al., 1998, 2002); however, comparison of spectra in Fig. 3 shows that the biogenic CM of cherts (type F) is different from that of meteorites (type M), despite the fact that the latter has the same chemical composition as a terrestrial coal (Hayatsu et al., 1977; Sephton et al., 1999). The characteristics of HYSCORE spectra in the different kinds of materials are summarized in Table 2 and are detailed below.

HYSCORE spectra at 9 K and τ=136 ns of two cherts from the Gunflint (1.9 Gyr) and Clarno (45 Myr) formations (F-type carbon) and for the CM of Orgueil and Tagish Lake meteorites (M-type carbon). Spectra on the right represent the contour map of HYSCORE (projection in the frequency plane). Color images available online at
The potential biosignatures are represented in bold type.
The lack of 1H signal in the Warrawoona chert is due to the total loss of hydrogen in this highly mature carbonaceous matter (Skrzypczak-Bonduelle et al., 2008).
3.2.1. 1H hyperfine interactions
The shape of the 1H-HYSCORE signal reflects the nature of the C-H bonds in radicals of the CM. These radicals are composed of a polyaromatic core terminated by hydrogen atoms or more or less branched aliphatic chains. Radicals in the different types of CM differ by the shape and the size of the aromatic part and by its branching level. Three types of hydrogen atoms can be considered as schematized in Fig. 4. The unpaired electron is delocalized on carbon 2p π orbitals of the aromatic fragment and interacts with nuclear spins by two different hf interactions: the isotropic interaction A iso, which is proportional to the electron spin density at the nucleus, and the dipolar interaction T, which depends on distance R between electron and nuclear spins as T α R −3. The first type of hydrogen atoms to be considered belongs to the aliphatic chains. As these 1H nuclei are the most distant from the electron spin (they are noted Hd in Fig. 4), they interact with the electron spin only by a weak dipolar interaction that gives the central peak along the diagonal ν 1=ν 2 at the nuclear frequency ν n=14.5 MHz of 1H (matrix line). The second type of hydrogen atoms is localized in benzylic positions (noted Hb in Fig. 4) and characterized by a hf interaction dominated by the isotropic coupling term A iso, with only a weak dipolar contribution T. They appear in the HYSCORE spectrum as pairs of spots along an axis ν 1=−ν 2 crossing the diagonal ν 1=ν 2, with a spitting equal to A iso. In the case of a distribution of hf couplings for Hb atoms, these spots overlap and give a spectrum in the form of a linear ridge along the antidiagonal ν 1=−ν 2 (Gourier et al., 2008). The third type of hydrogen atoms is localized on the edges of the aromatic rings (noted Ha). These aromatic hydrogen atoms are characterized by a significant dipolar interaction T in addition to the isotropic interaction A iso. In a disordered CM, this gives peaks and ridges that are positively shifted from the antidiagonal and exhibit a “horn shape.” The splitting between the extrema of the ridge is equal to A iso+2T, and the shift from the antidiagonal is 9T 2/32ν n (Pöppl and Kevan, 1996; Schweiger and Jeschke, 2001) (Fig. 4).
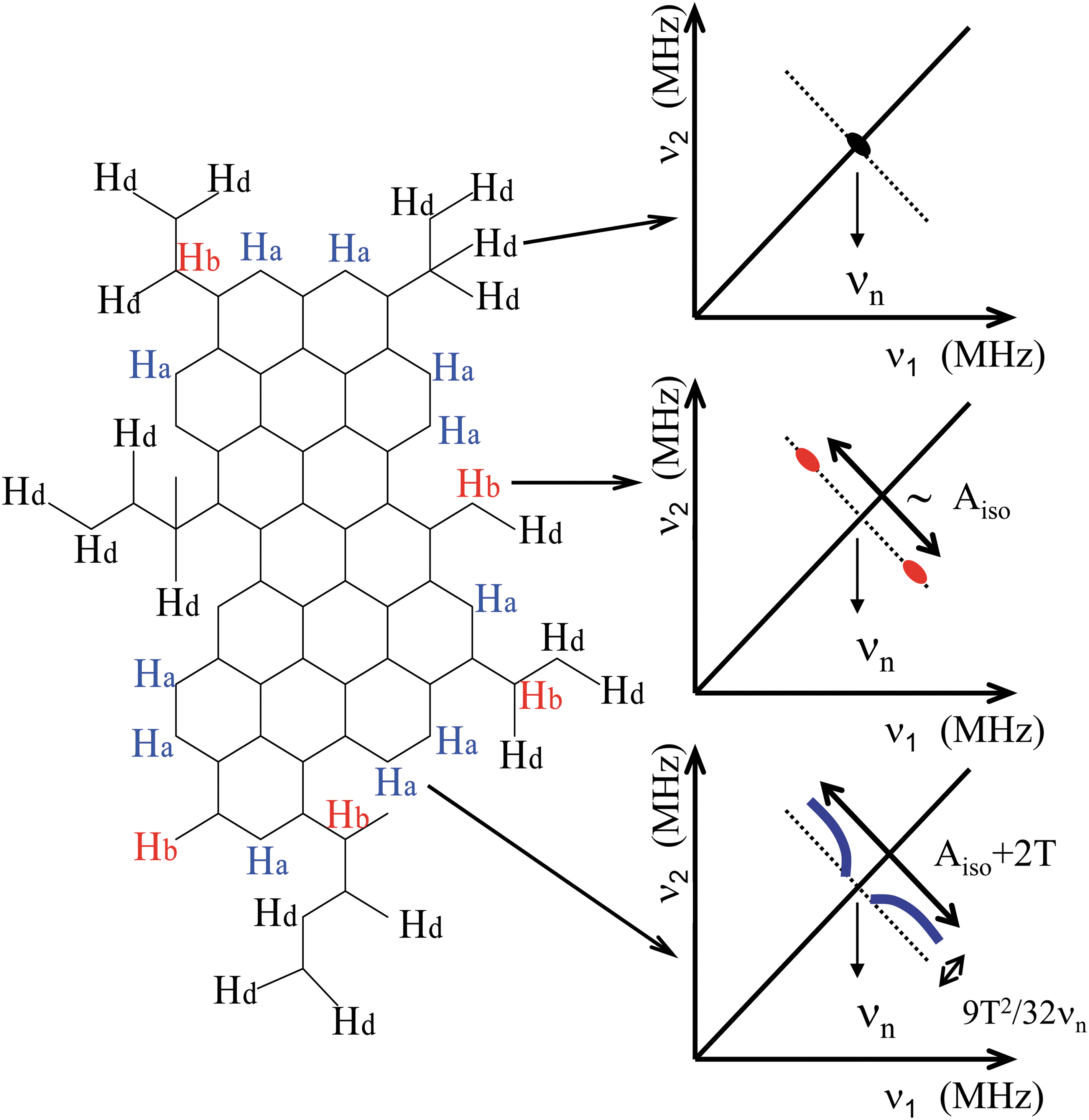
Schematic representation of a hypothetical branched polyaromatic fragment in primitive CM. The electron is delocalized on the 2p
π
orbitals of aromatic rings. Distant hydrogen (Hd), benzylic hydrogen (Hb), and aromatic hydrogen (Ha) represent the three types of hydrogen atoms that can be identified in HYSCORE spectra. Schematic HYSCORE spectra for Hd, Hb, and Ha hydrogen atoms are represented with the indication of the hyperfine parameters. A
iso and T are the isotropic and dipolar components of the hyperfine interaction, respectively. Color images available online at
It thus appears that the shape of the 1H-HYSCORE spectrum should mainly depend on the branching level of aromatic radicals. A poorly branched radical is dominated by aromatic (Ha) hydrogen atoms, which gives the “horn shape” spectrum observed for the biogenic CM (F-type) fossilized in cherts (Fig. 3). Alternatively, Ha atoms are much less abundant in highly branched radicals corresponding to meteoritic CM (Gardinier et al., 2000; Cody and Alexander, 2005), which are dominated by benzylic (Hb) hydrogen atoms as shown by the linear 1H ridge observed in meteoritic (type M) CM (Gourier et al., 2008).
More details on the hyperfine coupling can be deduced from the analysis of 1H ridges, although the lack of well-resolved hyperfine peaks in the spectra indicates a distribution of hf parameters due to the strongly disordered character of the CM. Figure 5 shows the 1H-HYSCORE spectra of Gunflint chert recorded at τ=128 ns and τ=348 ns. One type of hydrogen dominates the spectrum at τ=128 ns, while a more complex structure appears at τ=348 ns, revealing the presence of several types of hydrogen atoms. At both τ values, the dominant hydrogen atoms are characterized by A iso≈−3 MHz and T≈+7 MHz. The significant contribution of the dipolar interaction and the negative value of the isotropic coupling indicate that these dominant hydrogen atoms are bonded to aromatic carbon atoms (Ha hydrogen in Fig. 4). The same features are observed for Clarno (45 Myr). The spectrum for Warrawoona (3.5 Gyr) exhibits no 1H signal, which is in agreement with the high maturity of its CM. Indeed, the CM has lost its hydrogen as shown also by its very narrow cw-EPR linewidth (Fig. 1 and Table 1). On the contrary, the lack of vertical shift in the proton ridge of meteoritic CM (Fig. 3) indicates a distribution of A iso values with very small dipolar contributions, pointing to dominant hydrogen atoms in benzylic position (Hb type). This shows that aromatic radicals in meteoritic CM are more branched (less aromatic hydrogen) than in cherts, as previously inferred from 13C-NMR spectroscopy (Gardinier et al., 2000; Cody and Alexander, 2005).
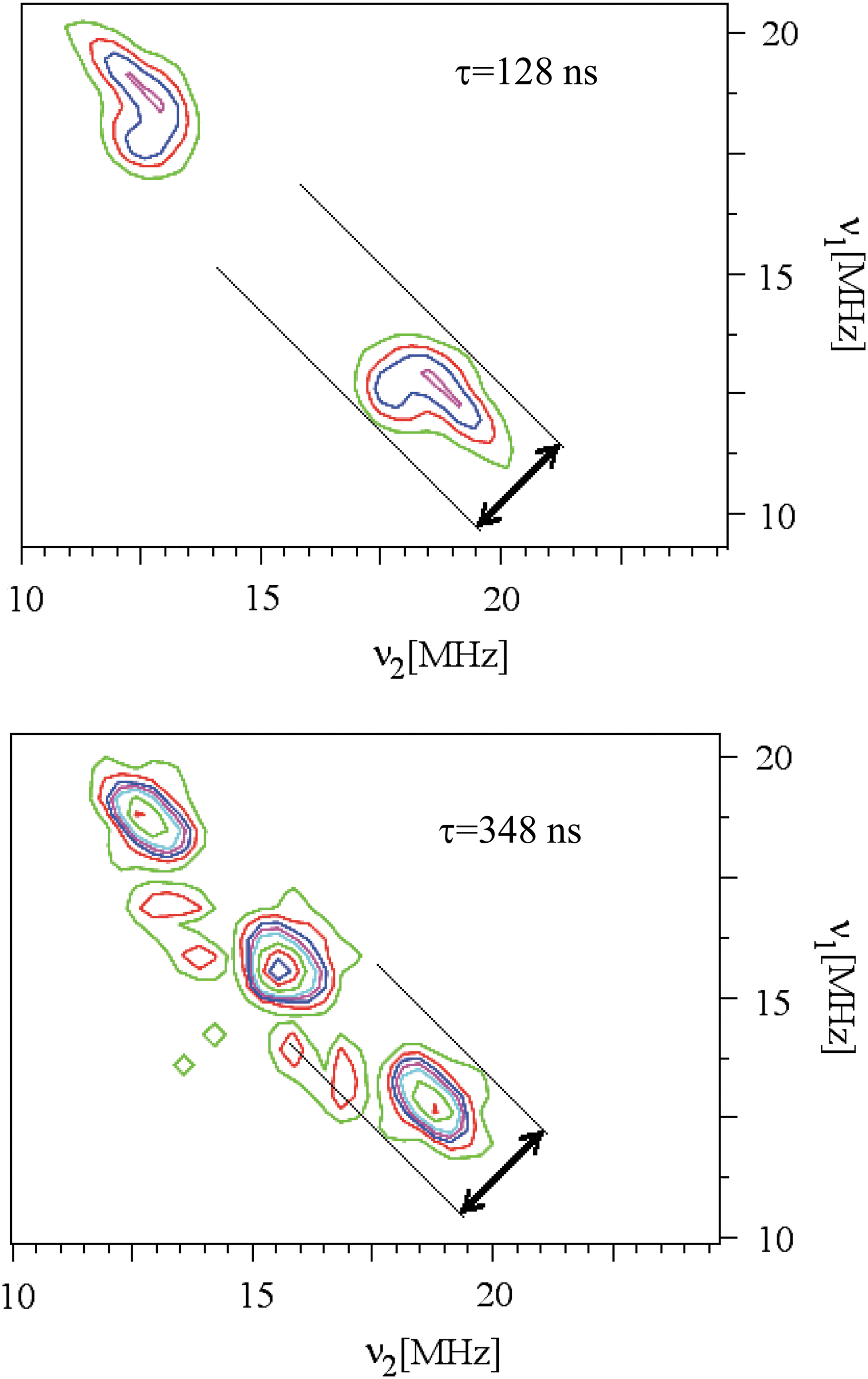
HYSCORE spectra at 4 K of Gunflint chert in the 1H frequency range, recorded at two τ values. The vertical frequency shift is represented by an arrow. Color images available online at
3.2.2. 13C hyperfine interaction
13C-HYSCORE spectra of biogenic CM and meteoritic CM are also clearly different (Fig. 3 and Table 2). The carbon signal centered at the nuclear frequency ν n=3.7 MHz of 13C is weak and narrow in cherts, while it is broad and intense in meteorites. Because of the low 13C abundance (1.1%), the intensity and shape of carbon signal are very sensitive to the geometry and environment of the radical. Ikoma et al. (1998, 2002) showed that the signal intensity ratio 13C/1H increases with the number of equivalent carbon nuclei interacting with the unpaired electron of the radical. This ratio is clearly larger in meteorites than in cherts, which may be related to the high aliphatic branching of aromatic units in meteoritic CM as already deduced from the shape of the 1H-HYSCORE spectra and from 13C NMR (Gardinier et al., 2000; Cody and Alexander, 2005). The large number of carbon atoms located at short distances from the polyaromatic unit increases the probability for the unpaired electron to interact with a 13C nucleus. This is not the case with the biogenic CM in cherts, which exhibits a weak 13C signal due to distant carbons and a 1H signal typical of aromatic hydrogens (Ha). These differences in the 13C/1H ratio may also reflect size differences of aromatic fragments (Ikoma et al., 1998, 2002). This would indicate that aromatic radicals of fossil CM in cherts are smaller than those of meteoritic CM. Nevertheless, HYSCORE spectroscopy shows that 1H and 13C signatures of biogenic CM can be easily distinguished from those of meteorites, despite the fact that they exhibit the same cw-EPR signal (Fig. 1).
3.2.3. Evidence of 31P and 14N in biogenic carbonaceous material
For particular τ values, a narrow HYSCORE peak appears at 12.2 MHz in cherts, which is lacking in meteoritic CM, as shown in Fig. 6 for Clarno, Gunflint, and Warrawoona cherts recorded with τ=136, 156, and 136 ns, respectively. As this peak does not correspond to 13C or 1H nuclear frequencies, it indicates the presence of a heteroelement localized inside or sufficiently close to carbonaceous radicals to interact with the electron spin via a dipolar interaction. There are no nuclei with such a high nuclear frequency; however, this value matches with a double frequency transition of 31P (I=1/2, ν n=6.1 MHz, abundance 100%) that should appear at 2ν n=12.2 MHz. Compared with the 1H signal, the intensity of the 31P signal increases with the age of the CM and is the dominant signal for the oldest sample (Warrawoona, 3.5 Gyr). However, this attribution of the peak at 12.2 MHz to a double frequency transition of 31P may appear speculative in absence of a direct observation of a 31P spectrum around 6.1 MHz.
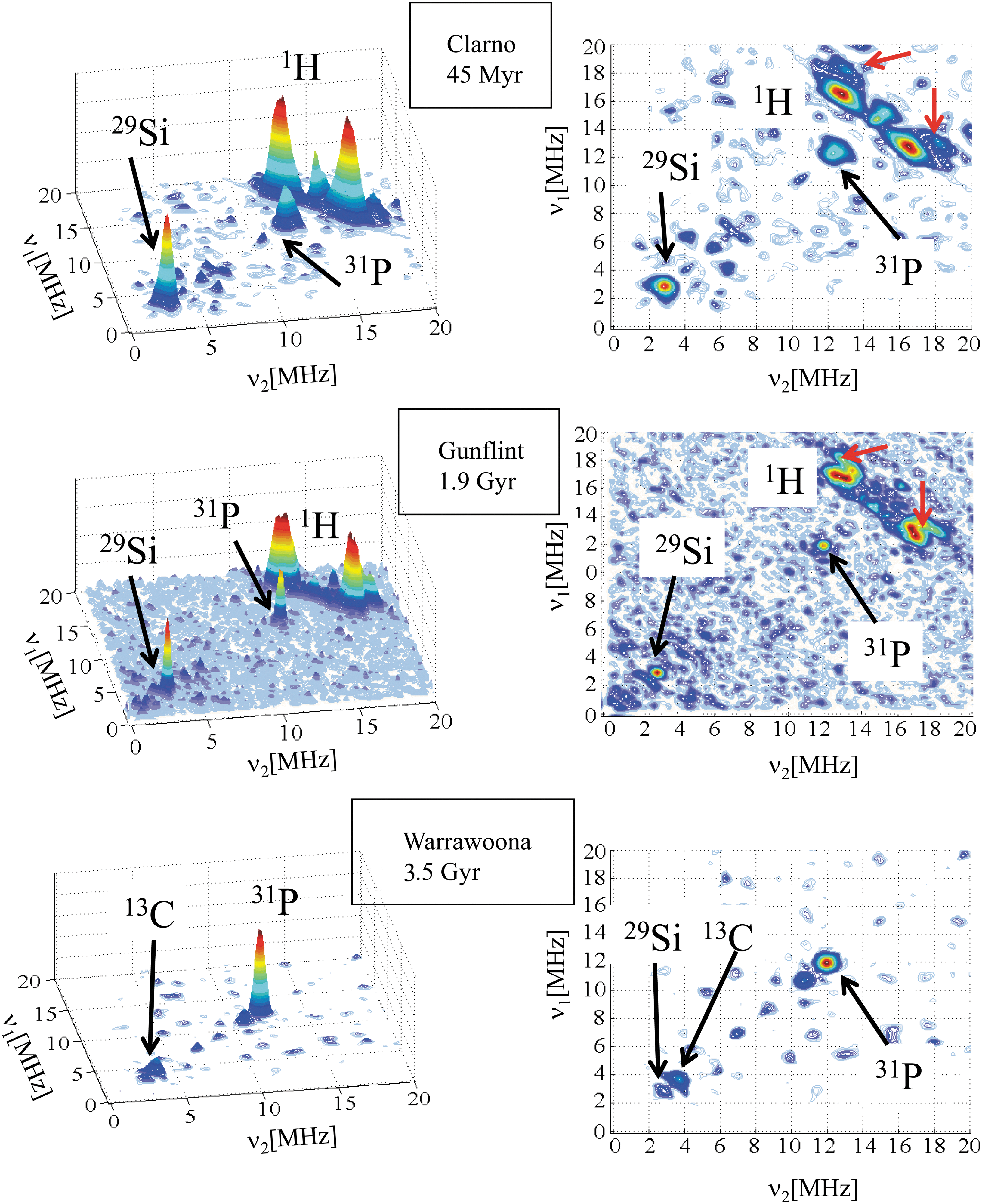
HYSCORE spectra at 9 K showing the double frequency 31P transition in three cherts: Clarno (τ=136 ns), Gunflint (τ=256 ns), and Warrawoona (τ=136 ns). Color images available online at
Convincing proof of the presence of 31P was obtained by studying the CM extracted from the Warrawoona chert by the HF/HCl extraction procedure (see Section 2.1). As the sample is an almost pure carbonaceous material after this demineralizing reaction, the HYSCORE intensity is much higher, and the spectrum reveals features that cannot be observed in a pristine chert, which contains only a small amount of CM. The HYSCORE spectrum at 4 K (Fig. 7) of the extracted CM shows two clear indications for the presence of 31P nuclei in addition to other nuclei and confirms the lack of hydrogen already observed in the chert (Fig. 6) and expected for such primitive CM (3.5 Gyr). In the positive quadrant (+,+) of Fig. 7, corresponding to hyperfine coupling regime A<2ν n for nuclear spin I=1/2, three peaks can be distinguished that are centered at 5.8 MHz. They correspond to double frequency transitions of 29Si (ν n=2.9 MHz). At a slightly higher frequency, three peaks centered at the frequency of 31P (ν n=6.1 MHz) correspond to hf interaction parameters A iso=8 MHz and T=1 MHz with phosphorous. Double quantum (dq) transitions of 14N are also clearly observed at low frequency. The negative quadrant (+,−) corresponds to hyperfine coupling regime A>2ν n, with peaks centered at frequency |A|/2, and split by 2ν n. In addition to double quantum peaks of 14N, two pairs of peaks can be distinguished that correspond to 13C (|A|/2=7.8 MHz) and 31P (|A|/2=6.9 MHz), identified by their splitting 2ν n=3.7 and 12.2 MHz, respectively. This result clearly shows that at least two types of 31P nuclei are bonded to the radical and characterized by their hyperfine interaction A≈8 MHz and A≈13.8 MHz, respectively. The strong 13C interaction A≈15.6 MHz is compatible with carbon atoms of the polyaromatic rings, which are directly in contact with the unpaired electron spin. The presence of several types of 14N nuclei in such mature CM indicates that these nitrogen atoms are incorporated in the aromatic rings, which protected them from elimination during diagenesis and metamorphism. In contrast, the 14N signal is very weak or lacking in the meteoritic CM.

HYSCORE spectrum at 4 K and τ=130 ns of the CM extracted from the Warrawoona chert, showing the hyperfine interactions with 31P, 13C, 14N, and 29Si nuclei (dq indicating a double quantum transition). The (+,+) quadrant corresponds to ν
1>0 and ν
2>0, and the (+,−) quadrant corresponds to ν1>0 and ν2<0. Color images available online at
The clear difference between the HYSCORE spectrum of the pristine chert, which contains only a double frequency peak of 31P with no measurable hf interaction, and the HYSCORE spectrum of the extracted CM, with measurable 31P hf interactions of two distinct phosphorous atoms and no double frequency peaks, may be a direct consequence of the demineralization procedure. The presence of multifrequency 31P transitions in pristine cherts (four-frequency transitions at 24 MHz are also observed, not shown) indicates that P atoms belong to a phosphorous-rich component in contact with the CM, so that there are no direct C-P bonds. This separation of the CM component and P-rich component explains the presence of magnetically equivalent 31P nuclei responsible for the multifrequency transitions, which weakly interact by dipolar interaction with the electron spins of the CM, giving a single peak. This strongly suggests that these P-rich components are embedded in the carbonaceous microstructures, as already observed for other inorganic compounds in Precambrian rocks (van Zuilen et al., 2012) and in meteorites (El Amri et al., 2005). The extraction of the CM by the demineralization procedure could have destroyed the P-rich microprecipitates, with formation of direct C-P bonds in the carbonaceous network that are responsible for measurable hf couplings with both isotropic and dipolar contributions. Whatever the nature of these P-rich components (clusters, nanoparticles, etc.), their systematic presence in contact with biogenic carbonaceous particles in cherts is a strong indication of the biological origin of phosphorous. They may originate from the decomposition of nucleic acids and other P-rich biomolecules during diagenesis. The fact that only 31P, 13C, and 14N interactions are detected in the Warrawoona chert is due to the complete hydrogen loss in this very mature CM. In the younger Clarno chert, the less mature CM gives a strong 1H signal and a weaker 31P signal, indicating less abundant and (or) less organized P-rich phases. The Proterozoic chert (Gunflint) is an intermediate situation. Also, the presence of various types of 14N in the biogenic CM extracted from cherts, which are lacking in the meteoritic CM, is a strong indication of the biologic origin of nitrogen, which may originate from the decomposition of proteins and nucleic acids.
3.2.4. Interaction with 29Si nuclei
As the CM is embedded in a Si-rich mineral matrix in cherts and in meteorites, it is not surprising to observe the systematic presence of a 29Si signal (4% natural abundance), with characteristics depending on the origin of the CM. This signal is weak and broad in the meteoritic CM, while it appears narrow and intense in cherts except for the oldest one (Warrawoona). However, a 29Si signal is clearly observed when the CM is extracted from this chert (Fig. 7). The differences in the 29Si signals may reflect different types of chemical bonding between the CM and the mineral matrix. We expect that direct C-Si bonds should give stronger hyperfine interactions (broader 29Si signals) than C-O-Si bonds. If this interpretation is correct, it means that carbonaceous particles would be dominantly linked to the mineral by C-O-Si bonds in cherts and by C-Si bonds in meteorites.
3.3. Effect of thermal treatments on the hyperfine interactions
It was shown in Section 3.1 that thermal treatments of a young chert and of biomolecules encapsulated in a SiO2 matrix give the same evolution of the cw-EPR line of CM. To evaluate the effect of thermal metamorphism on HYSCORE biosignatures, we compared in more detail the radicals produced by carbonization of biomolecules encapsulated in SiO2 (C-type CM) and the radicals produced by natural aging in cherts (F-type CM).
For each type of encapsulated molecules (BSA, DNA, ATP, glucose), the HYSCORE spectra were recorded at three τ values (136, 156, and 200 ns) after each thermal step (400°C, 450°C, 500°C, 550°C, 600°C, and 650°C). Figure 8 displays examples of HYSCORE spectra recorded at 9 K and τ=136 ns after the thermal step at 550°C. This temperature was chosen because it gives a cw-EPR line with the same linewidth ΔB pp (related to the H/C ratio of the CM) as Gunflint chert.

HYSCORE spectra at 9 K and τ=136 ns of molecular precursors encapsulated in sol-gel silica and treated at 550°C. Color images available online at
Several features of the C-type CM are similar to those observed in meteoritic (M-type) CM but differ from F-type CM (Figs. 8 and 3). First, the shape of the 1H ridges is linear and very similar in C-type and M-type CM. This linear ridge is the signature of the distribution of highly branched aromatic radicals. This similitude of C- and M-type CM is confirmed by the 13C signal, which is broad and intense in both types of carbonaceous materials, while it is weak and narrow in F-type CM. It is also confirmed by the 29Si signal, which is broad in both M- and C-type CM, while it is narrow and generally intense in F-type CM. The presence of similar 1H, 13C, and 29Si hf structures in C- and M-type CM that are different from those of F-type CM suggests that the organic precursors at the origin of the meteoritic CM were probably submitted to a thermal event before incorporation in the parent body. However, this thermal episode was not strong enough to eliminate hydrogen atoms. The cosmochemical implications of this result are beyond the scope of the present paper, and more work is needed to explore the thermal history of the meteoritic CM by EPR spectroscopy.
The second characteristic of C-type CM is the absence of 31P signal even after carbonization of P-rich precursors (DNA and ATP), while this signal is always present in F-type CM. This indicates that P-rich molecules produced during thermal treatment of DNA and ATP are not embedded in the CM in the form of P-rich phases as in F-type CM. Instead, they are localized at large distances from the electron spins. As dipolar interactions are proportional to R −3, where R is the distance between the 31P nuclei and the unpaired electron spin of radicals, the amplitude of the nuclear modulations of the spin echo vanishes at large distances, which explains the lack of 31P signal for carbonized DNA and ATP.
This possible effect of temperature on the separation of phosphorous-rich precipitates from the CM also occurs for thermal treatment on a young chert. The 2-D projections of the HYSCORE spectra at τ=200 ns of Clarno treated at 300°C and 400°C compared with the pristine sample are shown in Fig. 9. No HYSCORE spectra could be detected for thermal treatments at higher temperature. The pristine sample shows a very weak 13C signal, the double frequency peak of 31P and a 1H ridge with two peaks (arrows) corresponding to the anisotropic hyperfine interaction of Ha hydrogen atoms. These features are the main characteristic of F-type CM in cherts. Thermal treatments modify the 1H spectrum, with the loss of Ha hydrogen atoms, as revealed by the linearization of the proton ridge and the lack of shifted peaks, indicating that the radicals are more branched than in pristine chert. The vanishing of the double frequency 31P peak can also be observed. This confirms that a thermal episode can destroy the P-rich phases and modifies the branching level of aromatic moieties. Consequently, 1H, 13C, and 31P HYSCORE spectra of CM provide nuclear magnetic biosignatures only in cherts that have not been submitted to a metamorphic episode. However, the fact that these biosignatures are observed in Warrawoona and Gunflint cherts shows that they are not strongly affected by a metamorphic grade reaching the greenschist facies. This confirms the results obtained for the evolution of the cw-EPR lineshape (Bourbin et al., 2013).
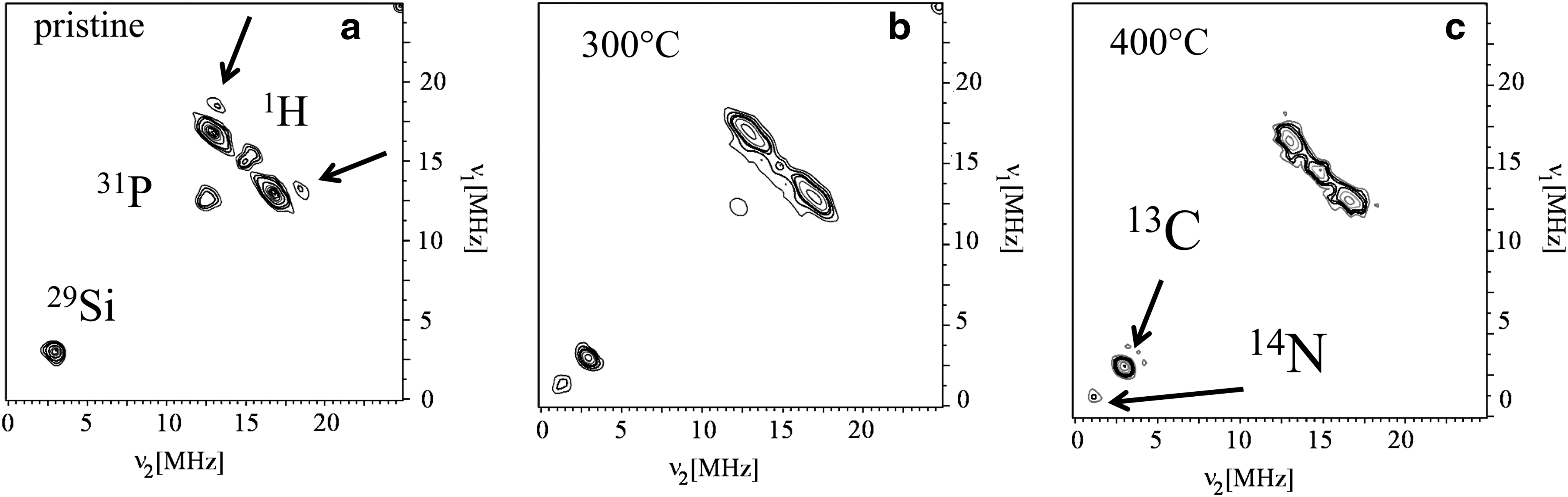
HYSCORE spectra at 30 K of Clarno chert recorded with τ=200 ns. (
4. Discussion
Organic radicals are always present in primitive CM (Skrzypczak-Bonduelle et al., 2008; Binet et al., 2002) and can thus be used as internal probes to study their chemical environment at a subnanometer scale by the indirect detection of nuclear resonances of nuclei with nonzero nuclear spin by using pulsed EPR spectroscopy. The main results are summarized in Table 2, highlighting the features that can be considered as possible biosignatures of primitive life.
Concerning the cw-EPR line itself, it was previously shown that the progressive loss of hydrogen (1H) during the Precambrian was responsible for the EPR line narrowing observed upon increasing age of the chert and of a specific modification of the EPR lineshape, which moved from a pure Lorentzian shape to a stretched Lorentzian shape (Skrzypczak-Bonduelle et al., 2008). The amount of stretching increases with the age of the CM, the most stretched line corresponding to the oldest CM (∼3.5 Gyr), which allowed us to propose a method for dating the CM with respect to its mineral matrix (Bourbin et al., 2013). On the opposite, the hydrogenated CM of carbonaceous meteorites (∼4.6 Gyr) always gives a purely Lorentzian EPR line (Binet et al., 2002, 2004b). Archean CM and meteoritic CM differ also by the temperature dependence of their EPR lines (proportional to the paramagnetic susceptibility), which obeys a Curie's law specific for S=1/2 radicals in Archean cherts and a more complex law indicative of S=1 diradicals in meteorites (Binet et al., 2004a, 2004b; Delpoux et al., 2011). However, such cw-EPR analyses, which give information on the age and the origin (terrestrial versus extraterrestrial) of the CM, give no indication of their biogenic/abiotic origin. In this work, the indirect detection of nuclear magnetic frequencies by pulsed EPR showed that the intensity and shape of the 1H spectrum is sensitive to the biogenicity of the organic precursors. As anticipated, the intensity of the 1H spectrum decreased in cherts of increasing age and vanished in the oldest Archean chert. This feature can be used to detect any biogenic contamination (endolithic bacteria, for example) in Archean cherts, as the CM originating from bacteria that contaminated Archean cherts long after the sediment formation (i.e., from the Proterozoic to present) would exhibit intense 1H spectra. An important result is that the shape of the 1H HYSCORE spectrum is also sensitive to the origin of the precursors and their maturation degree. At the molecular level, this feature is interpreted as the direct consequence of the degree of aliphatic branching of aromatic radicals, which controls the ratio C=Ha/(Ha+Hb+Hd), with C=0 for totally branched radicals and C=1 for radicals with no aliphatic branching. It appeared from pulsed EPR that C≈0 is characteristic of meteoritic CM and of carbonized molecular precursors, biogenic or not. On the contrary, CM from fossilized organisms is characterized by 0<< C<1. The sensitivity to thermal treatments of the nuclear magnetic spectra of CM indicates that the biosignatures are no longer valid for CM that experienced a metamorphism higher than the greenschist facies.
However, these 1H biosignatures cannot be used for Archean cherts, as CM has lost most of its hydrogen in this case. After checking that there is no 1H spectrum in an Archean sample, indicating a very low level of biological contamination, the next step is the study of other isotopes, the most interesting ones being 13C and 31P (and possibly 14N). The 13C spectrum is complementary to that of 1H, as it is also sensitive to the branching level of aromatic radicals. The most branched radicals (meteoritic CM and CM from carbonized precursors) give strong and broad 13C ridges, while poorly branched CM (fossils) gives very weak and narrow 13C peaks, independently of its age. Phosphorous is also a promising signature of a biological origin, as we systematically found a 31P signal in cherts containing fossilized bacteria. This signal is lacking in meteorites and, more surprisingly, in CM formed by carbonization of phosphorous-rich biomolecular precursors (DNA, ATP). This indicates that only the natural fossilization process is able to maintain phosphorous atoms at close distance (subnanometer scale) from the radicals. The most likely origin of this phosphorous is the decomposition of nucleic acids (DNA, RNA) of fossilized organisms. We are presently studying Archean chert samples from South Africa (Josefsdal Formation), which fully confirm the presence of 31P close to aromatic radicals. The chemical form of this phosphorous is not yet determined and will be the object of future work.
It must be stressed that all these nuclear magnetic biosignatures are only those corresponding to CM responsible for the narrow EPR line. It might be possible that other forms of CM are also present but are not detected by cw-EPR and thus cannot be probed by HYSCORE spectroscopy. For example, more or less graphitized carbons give broad lines with various shapes, width, and g-factor anisotropy (Barbon and Brustolon, 2012). If such forms of carbon are present in our samples, their cw-EPR signals are too weak to be detected, and we checked by echo-detected EPR that all the HYSCORE spectra correspond to the narrow cw-EPR line of CM. Moreover, the presence of graphitized carbon appears very unlikely in the studied cherts, because they suffer from a metamorphism that is too low (greenschist facies or lower) to transform carbon into graphite. This was precisely the reason why we selected these well-documented samples.
It appears that cw-EPR and pulsed EPR are complementary to Raman spectroscopy for the analysis of primitive CM. We believe that combination of Raman parameters of the well-known G and D lines (width, intensity ratio, etc.), cw-EPR parameters (g-factor, width, lineshape, magnetism), and pulsed-EPR parameters (intensity and shape of 1H, 13C, 31P resonances) would constitute fingerprints of the origin, age, and evolution of primitive CM. In addition to CM, it is worth noting that EPR can also be used to detect inorganic biosignatures, as for example biogenic vanadyl VO2+ ions (Gourier et al., 2010) and biogenic MnO2 mineralizations (Kim et al., 2011).
5. Conclusion
It may be concluded that EPR spectroscopy in its different modalities constitutes a powerful tool to investigate primitive organic matter in relation with the origin of life on Earth. In this work, we showed that the intensity and the shape of EPR-detected nuclear resonances of 1H, 13C, and 31P are markers of the precursors (biological, meteoritic), the age, and the thermal history of CM. The main advantage of EPR is a unique combination of sensitivity, resolution, and selectivity. In addition, this technique is non-invasive, as pristine samples can be studied without preparation, and CM can be detected at the part-per-million level in the bulk of small (centimeter- to millimeter-sized) pieces of rocks. In association with other methods (Bandfield et al., 2001; Marzo et al., 2007; Derenne et al., 2008; Orofino et al., 2010; Westall and Cavalazzi, 2011; Bower et al., 2013), EPR offers a great potential for the search for fossil traces of life on Mars.
Footnotes
Acknowledgments
The authors acknowledge the Centre National d'Etudes Spatiales (CNES) and the Institut National des Sciences de l'Univers (INSU) of CNRS for financial support. We also thank François Robert from Museum National d'Histoire Naturelle (Paris) for providing the chert samples. The carbonaceous matter of the Tagish Lake meteorite was provided by Luann Becker, from University of California, Santa Barbara (California, USA).
Abbreviations
ATP, adenosine triphosphate; BSA, bovine serum albumin; CM, carbonaceous matter; cw, continuous wave; EPR, electron paramagnetic resonance; hf, hyperfine; HYSCORE, hyperfine sublevel correlation; NMR, nuclear magnetic resonance.