
Abstract
During space missions, radiation represents a major hazard for human health and involves all body organs and tissues. Regarding thyroid function, it has been shown that ultraviolet radiation (UVC) has dose-dependent apoptotic effects on FRTL-5 cells, a normal strain of rat thyrocytes. We examined the effects of a sublethal dose of UVC on FRTL-5 cell growth and gene expression. Cells exposed to 10 J/m2 UVC showed no differences in viability compared to control cells after 24 h, but the BrdU incorporation was reduced, indicating a cytostatic effect. Quantitative RT-PCR carried out at 24 and 48 h after irradiation demonstrated that the mRNA levels of thyroglobulin (Tg), thyroperoxidase (Tpo), and sodium/iodide symporter (Nis) were transiently decreased at 24 h in treated cells, while the mRNAs of the thyroid transcription factors TTF1, Foxe1, and Pax8 were not affected. In cells cultured with TSH-free medium, the basal transcription of Tg, Tpo, and Nis genes was equally impaired by radiation and no longer stimulated by TSH. Overall, the results demonstrate that a sub-apoptotic dose of UVC compromises not only thyrocyte proliferation but also the expression of genes involved in thyroid hormone production. These findings might contribute to explaining the histological, biochemical, and clinical features of hypothyroidism observed in both animals and humans during spaceflight, and suggest that free thyroxine levels of astronauts during prolonged space missions should be monitored. Key Words: Thyrocyte—Thyroglobulin—Thyroperoxidase—Sodium/iodide symporter—Ultraviolet radiation—Spaceflight. Astrobiology 13, 536–542.
1. Introduction
M
Different studies have evaluated the plasma levels of key hormones involved in the regulation of fluid volume, electrolyte concentrations, and energetic metabolism in spaceflight conditions and have found multiple changes in the response of the endocrine system (Lane and Feeback, 2002). Among these, functional alterations of the thyroid suggestive of hypothyroidism have been documented in both animals and humans (Stein et al., 1999; Strollo, 1999; Grindeland et al., 2000; McMonigal et al., 2000; Macho et al., 2001; Lane and Feeback, 2002; Hellweg and Baumstark-Khan, 2007; Dayanandan, 2011). In particular, decreased triiodothyronine (T3) and elevated thyroid-stimulating hormone (TSH) plasma levels have been detected in astronauts during spaceflights compared with preflight values, and both hormones returned to the normal level in the postflight period. Similarly, decreased thyroxine (T4) and T3 plasma concentrations have been shown in rats flown aboard a biosatellite, together with morphological and histochemical changes consistent with reduced thyroid activity, that is, significant reduction of thyrocyte size, accumulation of colloid drops in the cytoplasm, decrease of iodinated thyroglobulin in the colloid, and lower T4 and T3 content per unit of thyroid tissue mass (Plakhuta-Plakutina et al., 1990). In vitro studies performed either on a normal strain of human thyrocytes or on the normal Fisher rat thyroid cell line FRTL-5, as well as on differentiated thyroid cancer–derived cell lines, have demonstrated that thyroid cells are sensitive to both gravity variations and UV radiation (Knopp et al., 1983; Ambesi-Impiombato and Villone, 1984; Meli et al., 1998, 1999a, 1999b; Chang et al., 1999; Loginov, 1999; Grimm et al., 2002a, 2002b; Kossmehl et al., 2002, 2003; Del Terra et al., 2003; Infanger et al., 2004, 2006; Kostic et al., 2010). Microgravity leads to changes in the cytoskeleton organization, altered expression of genes encoding extracellular matrix proteins, decrease of thyroglobulin, free-T3 and free-T4 secretion, reduced cAMP response to TSH, and increased apoptosis. Ultraviolet radiation (UVC, 254 nm) was shown to induce a dose-dependent activation of the apoptotic process in FTRL-5 cells cultured in the presence of TSH (Meli et al., 1999b; Del Terra et al., 2003). In the same cells flown in a stratospheric balloon, a similar effect appeared to be the result of modifications of the nuclear lipid metabolism, that is, augmentation of sphingomyelin degradation and phosphatidylcholine synthesis (Albi et al., 2009). A subsequent study performed on FRTL-5 cells cultured inside the International Space Station during the Eneide and Esperia missions demonstrated that despite TSH addition the proliferation rate was substantially lower compared to FRTL-5 cells on the ground and that cells were induced into a pro-apoptotic state (Albi et al., 2010).
To date, however, no information is available regarding the effects of UV radiation on the expression of thyroid-specific genes, that is, thyroglobulin (Tg), thyroperoxidase (Tpo), sodium/iodide symporter (Nis), and TSH receptor (Tshr), as well as of thyroid transcription factors TTF1/Nkx2-1 (thyroid transcription factor 1/NK2 homeobox 1), Foxe1/Ttf-2 (forkhead box E1/thyroid transcription factor 2), and Pax8 (paired box 8).
In the present study, the expression at the mRNA level of the above-mentioned genes has been analyzed in FRTL-5 cells with, and without, exposure to a sub-apoptotic dose of UVC radiation.
2. Materials and Methods
2.1. Materials
The nutrient mixture Ham's F-12 Coon's modified, TSH, somatostatin, transferrin, hydrocortisone, insulin, glycil-L-histidyl-L-lysine acetate, and primers were acquired from Sigma Chemical Co. (St. Louis, MO, USA). The Isol-RNA Lysis Reagent was purchased from Eppendorf (Hamburg, Germany). The Cell Proliferating Reagent WST-1, the Cell Proliferation ELISA BrdU kit, and the FastStart DNA MasterPLUS SYBR Green I kit were from Roche Applied Sciences (Mannheim, Germany).
2.2. Cell cultures
FRTL-5 cells were cultured in Ham's F-12 Coon's modified medium supplemented with 5% FBS and a mixture of 6 hormones (medium 6H), comprising insulin (1 μg/mL), hydrocortisone (10 nM), transferrin (5 μg/mL), somatostatin (10 μg/mL), glycil-L-histidyl-L-lysine acetate (10 ng/mL), and TSH (1 mU/mL) in 5% CO2 at 37°C, as previously described (Ambesi-Impiombato and Villone, 1984). To evaluate the effects of acute TSH stimulation on the expression of thyroid-specific genes in some experiments, the cells were cultured in the same medium and conditions but without TSH (medium 5H) for 5 days prior to the hormonal treatment.
2.3. Ultraviolet radiation experiments
Two days before the experiments, FRTL-5 cells were seeded to achieve 70–80% confluence at the time of UVC exposure. For the latter, the cells were washed twice in phosphate buffered saline (PBS), the liquid was removed, and the cells were exposed to either 10 or 50 J/m2 UVC (254 nm) in the Bio-Link BLX-E (Vilber Lourmat, Marne-la-Vallée, France). The two UVC doses were chosen on the basis of previously published data that show a strong induction of apoptosis in FRTL-5 cells exposed to 50 J/m2 UVC and indicate 20 J/m2 as the lower apoptosis-inducing dose (Del Terra et al., 2003). Following UVC exposure, fresh medium was added, and the cells were incubated for an additional 24 or 48 h.
2.4. Cell viability and proliferation assays
The cells were cultured in 96 well plates, exposed or not to UVC as described above. The cells were incubated for 24 or 48 h. Subsequently, the cell viability and growth were analyzed through colorimetric assays. The cell proliferating reagent WST-1 or BrdU was added to the cells in order to measure metabolic activity and DNA synthesis, respectively, according to the manufacturer's instructions, using a plate reader (DAS, Rome, Italy).
2.5. FACS analysis
The cells were cultured on 10 cm diameter Petri dishes, exposed or not to UVC as above described, and incubated for 48 h. The supernatants were then collected, and the cells were washed with PBS, harvested by incubation for 5 min at 37°C in PBS with 0.1% EDTA, and centrifuged at 1200 rpm for 5 min together with their supernatant. After a wash in PBS, the cells were resuspended in 70% ice-cold ethanol, labeled with propidium iodide, and analyzed for the DNA content as previously described (Ulisse et al., 2007) using the FACScalibur Flow cytometer and CellQUEST software (BD Biosciences, San Jose, CA, USA).
2.6. Analysis of gene expression
The cells were cultured in complete medium (6H) or starved for 5 days in 5H medium. Subsequently, they were exposed or not to UVC, and fresh 5H or 6H medium was added to all the cultures. After a 24 and 48 h incubation, total RNA was extracted with the Isol-RNA Lysis Reagent as previously described (Cianfarani et al., 2010; Ulisse et al., 2011); 2 μg of RNA were reverse-transcribed and the cDNA used for the subsequent quantitative PCR amplifications of Tg, Tpo, Tshr, Nis, Pax8, TTF1, Foxe1, and Gapdh (glyceraldehyde-3-phosphate dehydrogenase), the latter as internal control. The primers used are listed in Table 1. Controls for DNA contamination were performed by omitting the reverse transcriptase in parallel samples. The reaction mix was prepared with the FastStart DNA Master SYBR Green I kit, and the amplification was performed with the LightCycler instrument (Roche Diagnostics, Milano, Italy). The samples were processed in triplicate, and PCR-grade water was used as negative control. Standard run curves were generated for each gene by using 5-fold serial dilutions of a cDNA mixture. The PCR amplicons were visualized on 2% agarose gel, and their specificity was checked by automated DNA sequencing (Primm, Milano, Italy). Calculation of data was accomplished by means of the LightCycler relative quantification software 1.0 (Roche Diagnostics, Milano, Italy), and finally the expression values of the different genes in UVC-exposed cells were normalized against those found in control cells.
2.7. Statistical analysis
All the experiments were performed in triplicate, and the statistical evaluation of the data was accomplished by the Student t test. The results were considered significantly different if the pertaining p values were lower than 0.05.
3. Results
Initially, the dose-dependent apoptotic effect of UVC on FRTL-5 cells was analyzed by colorimetric assay. The results showed no significant variation of the viable cell population following treatment with the low UVC dose (10 J/m2), but at higher dose (50 J/m2) it was significantly reduced to 0.363±0.165–fold (p<0.01) after 24 h and to 0.256±0.058–fold (p<0.01) after 48 h (Fig. 1A). Likewise, the FACS analysis demonstrated that the low UVC dose did not induce significant apoptosis in FRTL-5, while the high UVC dose triggered a massive apoptosis, detected as accumulation of sub-G0/G1 nuclei in irradiated cells (Fig. 1B). To further analyze the effects of UVC on FRTL-5 proliferation, BrdU incorporation following exposure was determined. As reported in Fig. 2, both UVC doses induced a significant reduction of BrdU incorporation at 24 h with respect to control cultures. However, unlike the cells subjected to 50 J/m2, those which received 10 J/m2 displayed a significant recovery at 48 h. Based on such evidence, the dose of 10 J/m2 was used to examine by real-time RT-PCR the UVC effects on the expression of thyroid-specific genes, that is, Tg, Tpo, Nis, and Tshr, and of the transcription factors TTF1, Pax8, and Foxe1. Data reported in Fig. 3 show that the exposure to UVC radiation transiently decreased the Tg (p<0.05), Tpo (p<0.01), and Nis (p<0.01) transcripts by 50% or less. After 2 days, the Tpo mRNA level had increased significantly, and those of Tg and Nis returned to control values (Fig. 3). Conversely, no alteration of Tshr mRNA was observed (data not shown). In light of the above findings, we decided to investigate the expression of the three main transcription factors that regulate thyroid-specific genes. As shown in Fig. 4, the TTF1, Pax8, and Foxe1 mRNA levels were not significantly affected by the irradiation of FRTL-5 cells with 10 J/m2 UVC.
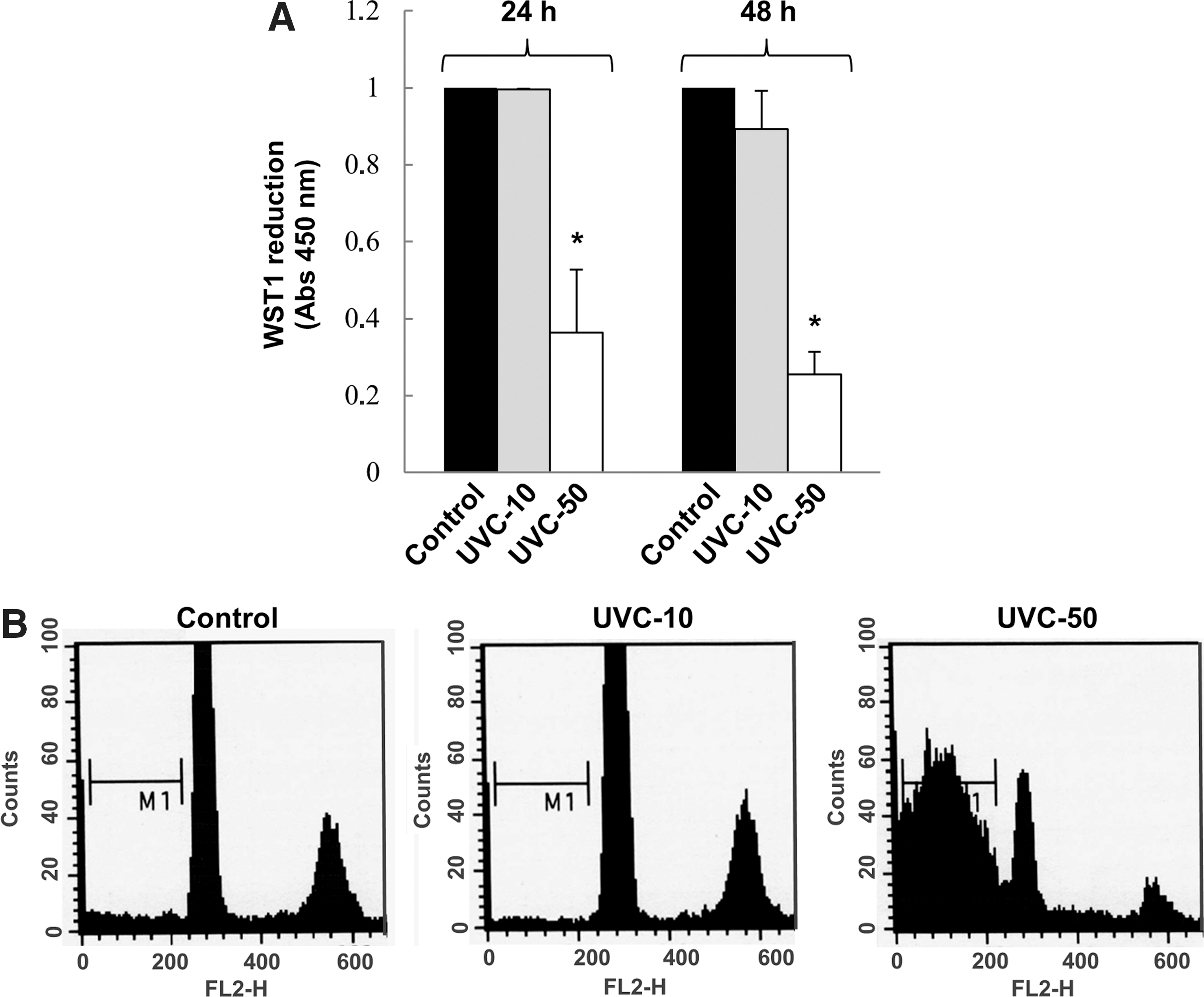
FRTL-5 cells have been exposed to 10 or 50 J/m2 UVC (254 nm) and then cultured for 24 and 48 h in standard conditions. (
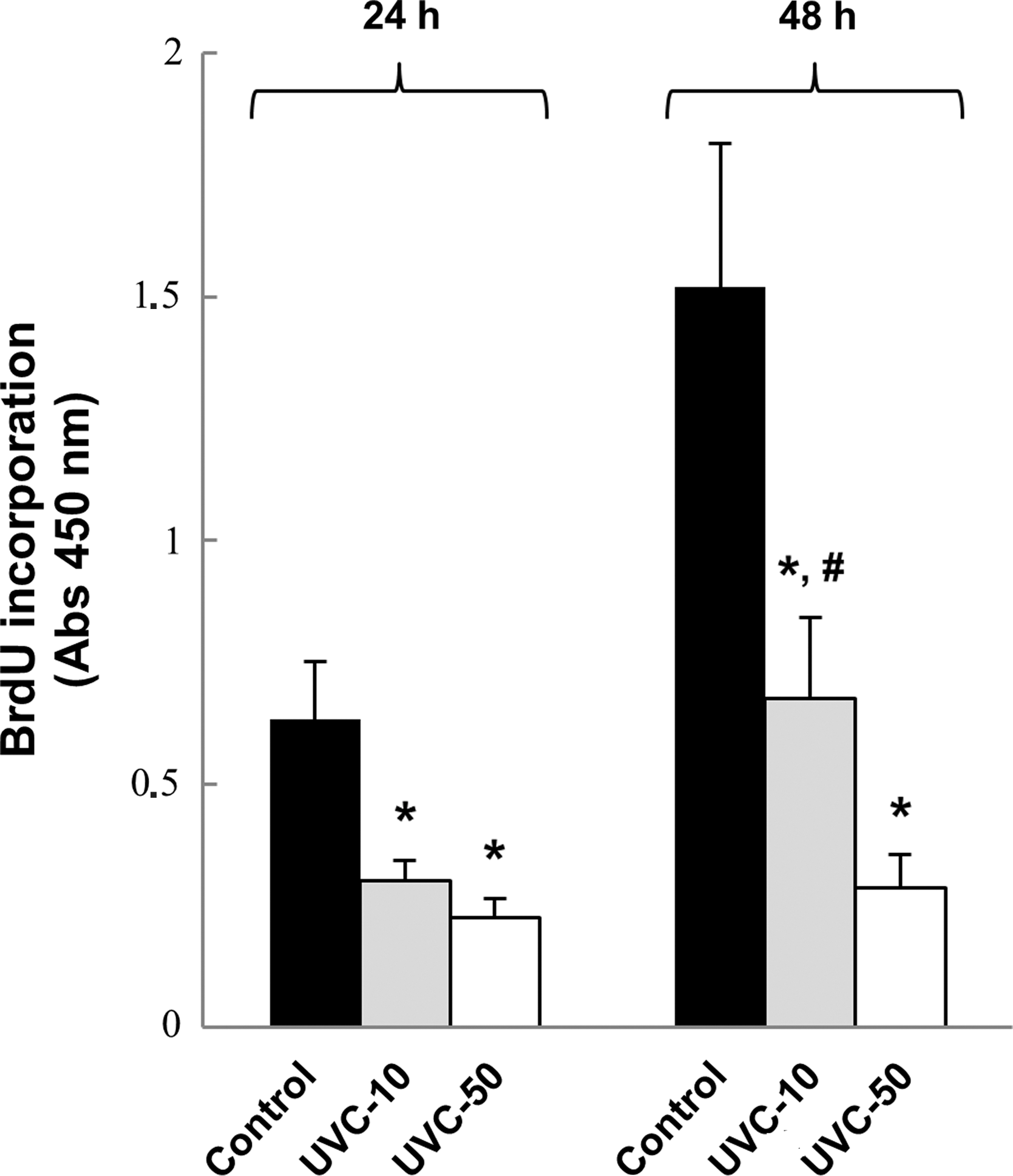
BrdU incorporation assay of FRTL-5 cells irradiated with 10 or 50 J/m2 UVC (254 nm) and then cultured for 24 and 48 h in standard conditions. *p<0.01 (UVC vs. control), # p<0.05 (48 h UVC-10 vs. 24 h UVC-10).
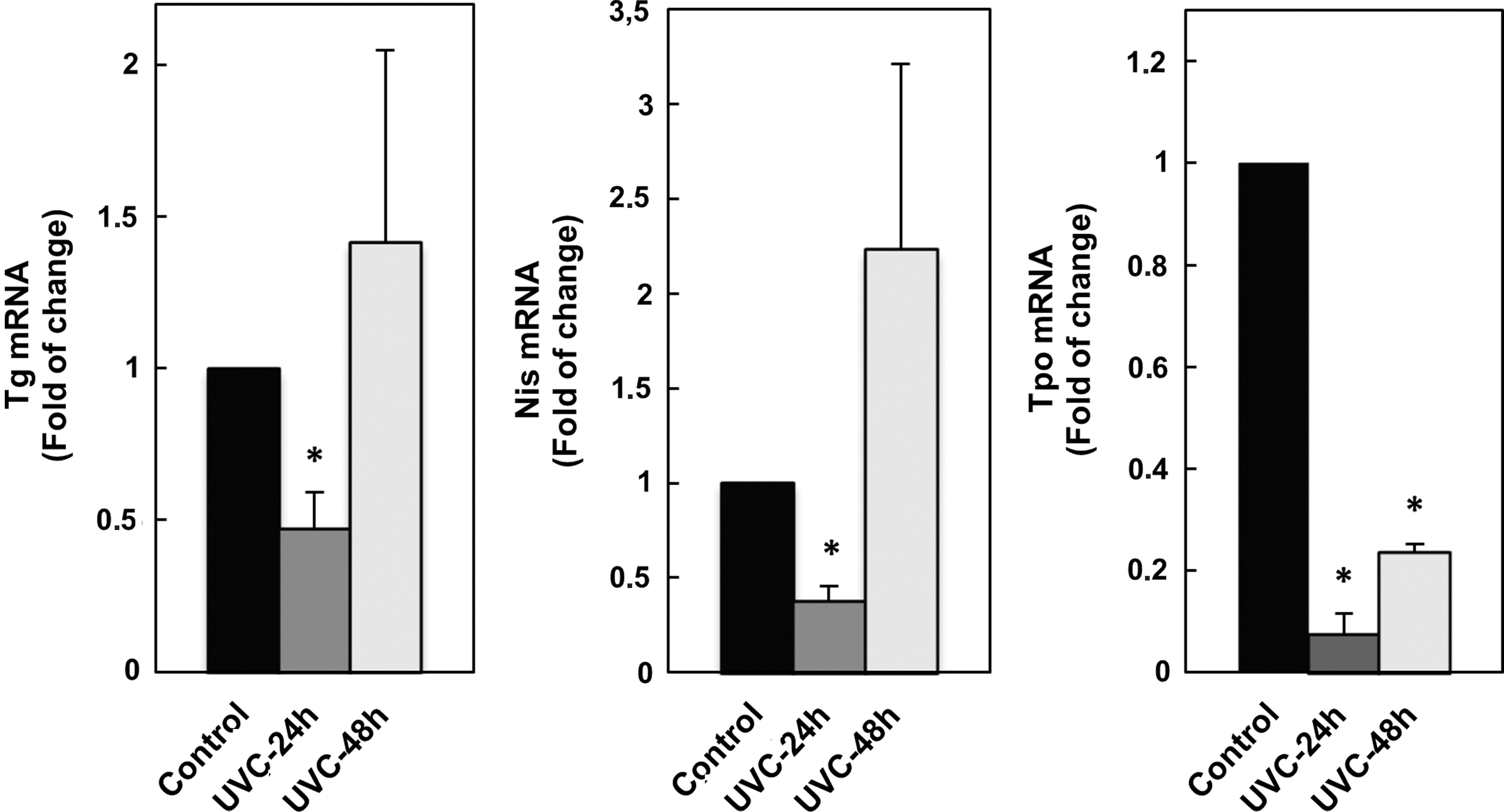
Evaluation of the Tg, Nis, and Tpo mRNA levels in FRTL-5 cells by quantitative RT-PCR. Cells have been irradiated with sub-apoptotic UVC dose (10 J/m2) and then cultured for 24 and 48 h in standard conditions. *p<0.01.
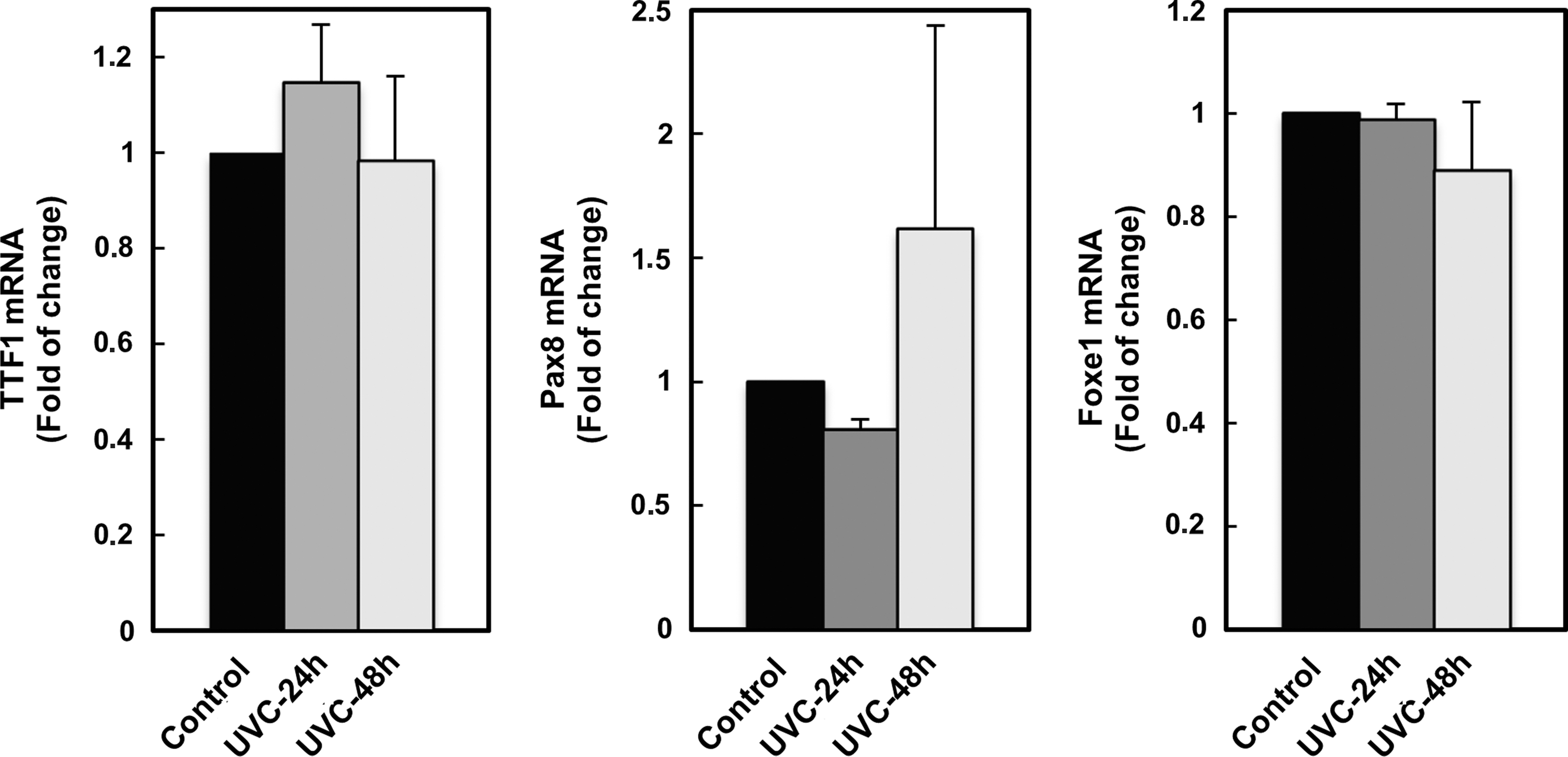
Evaluation of the TTF1, Pax8, and Foxe1 mRNA levels in FRTL-5 cells by quantitative RT-PCR. Cells have been irradiated with sub-apoptotic UVC dose (10 J/m2) and then cultured for 24 and 48 h in standard conditions.
In response to the TSH, FRTL-5 cells synthesize, secrete, and iodinate thyroglobulin, displaying an increased gene expression of Tg, Tpo, and Nis (Aza-Blanc et al., 1993; Kogai et al., 1997; Takano and Amino, 1997). Thus, the next step was that of verifying whether the reduced expression of Tg, Tpo, and Nis following UVC exposure was due to impaired basal and/or TSH-induced transcription. To this end, cells were starved for 5 days in 5H medium, exposed to UVC, and then replaced in culture for an additional 24 h in either 5H or 6H medium. The results, shown in Fig. 5, demonstrate a significant reduction of both basal and TSH-induced Tg and Tpo mRNA levels (p<0.01) and of TSH-induced Nis expression (p<0.05).
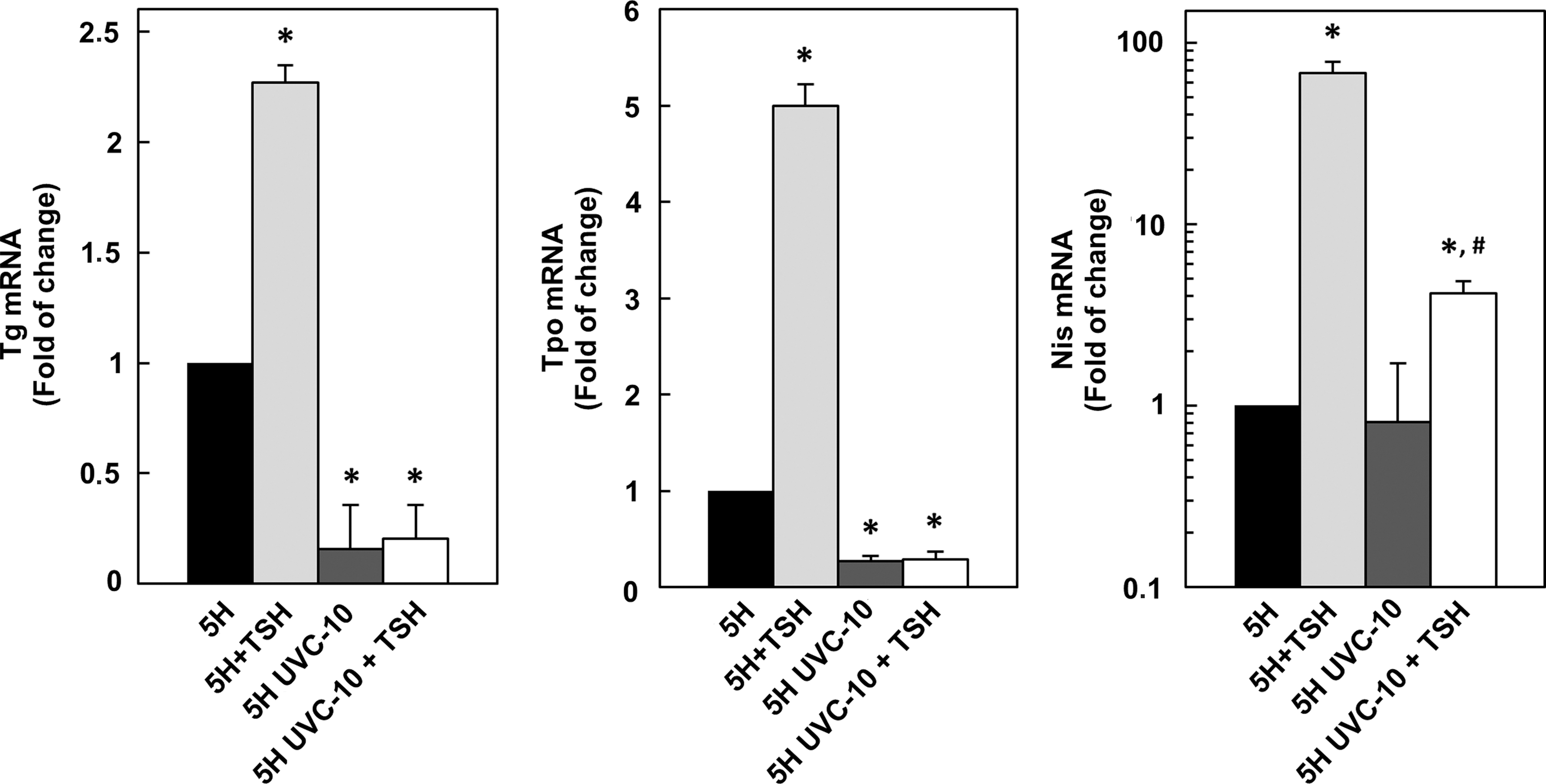
Quantitative RT-PCR analysis of Tg, Tpo, and Nis mRNAs in FRTL-5 cells starved in medium 5H for 5 days then irradiated with sub-apoptotic UVC dose (10 J/m2) and further cultured in fresh 5H or 6H medium for 24 and 48 h. *p<0.01, # p<0.05 (5H UVC-10 vs. 5H UVC-10+TSH).
4. Discussion
During spaceflight, astronauts are subjected to hypogravity and radiation, both of which appear to compromise the normal functions of many body organs and systems, including the thyroid gland. In this regard, alterations of the human thyroid function indicative of hypothyroidism have been documented in both animals and humans. However, experimental studies on humans, animals, or cell cultures inside spacecraft have not been able to make the distinction as to whether the effects are due exclusively to hypogravity or radiation.
Although UVC is characterized by lower energy levels with respect to cosmic rays, UVC with wavelengths of 254 nm has sufficient energy to cleave the bonds in the polymer chains, producing free radicals, which can initiate further reactions with molecular oxygen and cause the splitting of macromolecules (El Ghazaly and Alzahrani, 2013). Such events are substantially analogous to those triggered more intensely by ionizing radiation, and this makes UVC a suitable and easily manageable model for in vitro studies.
In the present work, we sought to examine the effects of a low UVC dose on the thyrocyte function in terms of viability, proliferation, the expression of thyroid-specific genes and relative transcription factors. Our results demonstrate that the exposure of FRTL-5 cells to a sub-apoptotic UVC dose causes provisional growth arrest and reduction of Tg, Tpo, and Nis mRNA levels. The cell replication delay is presumably due to UVC-induced lesions of DNA such as pyrimidine dimers, pyrimidine–pyrimidone (6–4) photoproducts, and DNA adducts, which in turn trigger activation of cell cycle checkpoints and intervention of nucleotide-excision repair processes (Kiefer, 2007). Otherwise, the cells would become less responsive to the TSH stimulus, as also indicated in a previous report monitoring the proliferation of FRTL-5 cultures inside the International Space Station (Albi et al., 2010). In addition, oxidative/reductive events are known to modulate various cellular functions through changes in the gene expression profile, ensuing from alterations in transcription factor activity. In thyrocytes, it has been demonstrated that the DNA binding of Pax8 and TTF1, thyroid transcription factors that regulate Tg, Tpo, Nis, and Tshr genes, decreased upon exposure to oxidating agents and was fully restored by reducing agents (Arnone et al., 1995; Kambe et al., 1996; Tell et al., 1998). In particular, in vitro experiments have shown that only the reduced form of the paired domain of Pax8 is capable of assuming the right conformation for sequence-specific DNA binding (Tell et al., 1998). Similarly, it has been indicated that TTF1 oxidation leads to the formation of disulfide bridges that stabilize preexisting TTF1 dimers and generate TTF1 oligomers inactive in DNA binding (Arnone et al., 1995). The transcriptional activities of Pax8 and TTF1 were found to have increased following treatment of FRTL-5 cells with TSH, and it has been purported that such stimulation might be masked by adding dithiothreitol to cell extracts, indicating that TSH upregulates the DNA binding of both factors, at least in part, by reducing preexisting, oxidized forms (Kambe et al., 1996). The fact that the rat Tg and Tpo promoters contain specific sequences for Pax8, TTF1, and Foxe1 should be taken into consideration. The rat Nis enhancer bears multiple TTF1 and Pax8 recognition sites, and a further TTF1 sequence is present at the rat Nis promoter. The TTF1 binds also to rat Tshr promoter. Such information provides a likely explanation for our findings. Indeed, the observed decrease of Tg, Tpo, and Nis transcription rate after UVC exposure might be ascribed to a transitional oxidation of TTF1 and Pax8, enduring until the cellular responses against oxidative damage reset the pre-irradiation redox homeostatic level. The insensitivity of the Tshr mRNA to UVC might be due to differences in the Tshr promoter structure, such as the lack of Foxe1 and Pax8 recognition sites, and the presence of a single TTF1 binding site. The basal transcription of Tg, Tpo, and Nis genes in cells cultured without TSH is equally affected by UVC. Moreover, the addition of TSH after UVC exposure elicits a lower increase in Nis mRNA and is no more effective in stimulating the Tg and Tpo genes compared to control cells. These results are in agreement with the above-mentioned study and demonstrate that TSH boosts the binding activities of TTF1 and Pax8 factors by promoting their oxidative-reductive transition (Kambe et al., 1996).
In conclusion, our data indicate that the exposition of normal rat thyrocytes to a low sub-apoptotic dose of UVC causes not only provisional growth slowdown but also impairment of cell-specific transcriptional activation of the genes involved in iodine uptake and thyroid hormone synthesis. This might account, at least in part, for the hypothyroidism registered in both animals and humans during spaceflights, and along with the results from different independent studies suggests that the free thyroxine levels of astronaut crews during prolonged space missions should be monitored.
Footnotes
Acknowledgment
This work was supported by the Italian Space Agency (ASI).
Author Disclosure Statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Abbreviations
Foxe1/Ttf-2, forkhead box E1/thyroid transcription factor 2; Gapdh, glyceraldehyde-3-phosphate dehydrogenase; Nis, sodium/iodide symporter; Pax8, paired box 8; PBS, phosphate buffered saline; T3, triiodothyronine; T4, thyroxine; Tg, thyroglobulin; Tpo, thyroperoxidase; TSH, thyroid-stimulating hormone; Tshr, TSH receptor; TTF1/Nkx2-1, thyroid transcription factor 1/NK2 homeobox 1.