
Abstract
To evaluate effects of microgravity on virulence, we studied the ability of four common clinical pathogens—Listeria monocytogenes, methicillin-resistant Staphylococcus aureus (MRSA), Enterococcus faecalis, and Candida albicans—to kill wild type Caenorhabditis elegans (C. elegans) nematodes at the larval and adult stages. Simultaneous studies were performed utilizing spaceflight, clinorotation in a 2-D clinorotation device, and static ground controls. The feeding rate of worms for killed E. coli was unaffected by spaceflight or clinorotation. Nematodes, microbes, and growth media were separated until exposed to true or modeled microgravity, then mixed and grown for 48 h. Experiments were terminated by paraformaldehyde fixation, and optical density measurements were used to assay residual microorganisms. Spaceflight was associated with reduced virulence for Listeria, Enterococcus, MRSA, and Candida for both larval and adult C. elegans. These are the first data acquired with a direct in vivo assay system in space to demonstrate virulence. Clinorotation reproduced the effects of spaceflight in some, but not all, virulence assays: Candida and Enterococcus were less virulent for larval worms but not adult worms, whereas virulence of MRSA and Listeria were unaffected by clinorotation in tests with both adult and larval worms. We conclude that four common clinical microorganisms are all less virulent in space. Key Words: Virulence—Spaceflight—Nematodes—Clinorotation—Microgravity—Microorganism. Astrobiology 13, 1081–1090.
1. Introduction
S
Clinorotation to generate suspension cultures on the ground provides some properties of microgravity conditions, specifically low shear, colocalization of biologics of differing size and density, and randomization of the gravity vector (Hammond and Hammond, 2001). Clinorotation culture conditions have substantial effects on the growth and phenotype of diverse cells, microorganisms, and phyla (Hammond and Hammond, 2001). However, to maintain cells in suspension, gravity must be continuously offset with an equal and opposite force, most typically shear force (Hammond and Hammond, 2001). It follows that during suspension culture on Earth, the cells are exposed to shear stresses. However, in the true microgravity of space, shear can be almost entirely removed, providing a unique operating range of culture conditions not available in ground-based experiments (Hammond and Hammond, 2001).
Application of space-based culture conditions to the study of microorganisms can inform us about risks to astronauts with possible changes in bacterial virulence, and in addition, the mechanistic knowledge obtained is relevant to a number of ground-based applications. Yet experiments designed to address questions of the nature and mediators of virulence changes in space continue to produce confounding results. Most, but not all, bacteria divide more rapidly in spaceflight, and growth change is linked to motility (Kacena et al., 1999; Benoit and Klaus, 2007). Microgravity also affects the phenotypes of bacterial colonies. For example, Pseudomonas sp. in spaceflight grows in both planktonic and biofilm forms (Collins, 2011). Candida grown in a rotating wall vessel transition more rapidly from single cells to a filamentous phenotype, form biofilm communities (Altenburg, 2008; Searles et al., 2011), and become more resistant to the antifungal agent amphotericin B, all of which suggest increased virulence. Salmonella grown under clinorotation are more virulent when injected into mice (Nickerson et al., 2000; Wilson et al., 2007). Salmonella grown in space and returned as live cultures at ambient temperature also appear to be more virulent when injected into mice following return, reentry, and unloading and handling delays (Wilson et al., 2007). However, none of these approaches directly evaluate host-pathogen interactions under microgravity.
The worm C. elegans is a powerful model system with which to study host-pathogen interactions (Harvill and Miller, 2000; Sifri et al., 2005; Hammond et al., 2009). We have adapted this model for use in spaceflight (Hammond et al., 2009). The natural diet of C. elegans is bacteria and fungi in the soil, but they eat and are infected by a variety of pathogens in the lab setting (Sifri et al., 2005; Hammond et al., 2009). In the presence of a virulent microorganism, the worm is killed; thus the quantity of surviving microorganisms becomes an indicator of worm mortality and hence virulence (Smith et al., 2002; Hammond et al., 2009). These methods allow the study of the subtle interplay of host organism and bacterial factors, as host defenses interact with bacterial virulence factors directly in spaceflight (Harvill and Miller, 2000; Sifri et al., 2005; Hammond et al., 2009).
The purpose of the present study was to use our spaceflight-adapted nematode assay to evaluate the effect of microgravity on the virulence of four common clinical pathogens: Listeria, methicillin-resistant Staphylococcus aureus (MRSA), Enterococcus, and Candida. Listeria is typically a food-borne pathogen responsible for gastroenteritis, meningoencephalitis, and spontaneous abortions. Enterococcus causes urinary tract infections, endocarditis, and meningitis. Candida infections range in severity from vaginitis to septic shock (Searles et al., 2011; Gow et al., 2012). MRSA infections are one of the most prevalent and costly infectious conditions within the health care system.
2. Materials and Methods
2.1. Test organisms
The following microorganisms were obtained from the American Tissue Type Collection (Manassas, VA): Listeria monocytogenes, ATCC 7644; Enterococcus faecalis OG1RF, ATCC 47077; methicillin-resistant Staphylococcus aureus FPR3757, ATCC BAA-1556; and Candida albicans SC5314. Wild type N2 Bristol nematodes were purchased from the Caenorhabditis Genetics Center at the University of Minnesota, Twin Cities.
Worms were expanded on nematode growth media plates seeded with E. coli, and eggs were prepared by standard techniques (Smith et al., 2002; Hammond et al., 2009).
2.2. Chemicals and reagents
All chemicals and reagents were purchased from Sigma Chemical Co. (St. Louis, MO).
2.3. Spaceflight hardware
To conduct experiments in spaceflight, we made use of the Fluid Processing Apparatus (FPA) (Hoehn et al., 2004; Hammond et al., 2009). The FPA is a glass tube that allows fluids, biologicals, and other reagents, all of which are initially separated by stoppers, to be serially mixed via a bypass channel (Hoehn et al., 2004) (Fig. 1). The FPA can be configured to isolate four separate volumes between moveable rubber septa (Hoehn et al., 2004; Hammond et al., 2009). An outer sheath houses the activation plunger. When depressed, the plunger pushes the septa forward, forcing the upstream fluid through a small bypass channel molded into the glass barrel and into the adjacent distal volume. Staged mixing of up to three fluids (total≤3.25 mL) into the distal fourth chamber (≤3.5 mL) can be achieved in this manner. Through controlled, two-stage series of fluid additions, experiments can be activated and terminated in sequential steps. Gas exchange is enabled through the use of hydrophobic PTFE Teflon membranes [pore size 0.02 μm, >700 kPa (100 psi) water entry pressure] (Hammond et al., 2009) in the septum located at the far end of the FPA most distal to the plunger (Hoehn et al., 2004; Hammond et al., 2009).

Clinorotation setup. The photograph shows the orientation of FPAs in GAPs for clinorotation experiments. The photograph shows FPAs with three chambers. The actual FPAs used for the studies in this report had four chambers, as shown in the schematic below the photograph.
The FPAs are assembled in groups of eight around the perimeter of a cylindrical container known as a Group Activation Pack (GAP) and plunged simultaneously with a crank pressure plate mechanism and crank handle (Hoehn et al., 2004; Hammond et al., 2009). In the typical configuration, C. elegans or buffer is placed in the first chamber, growth media in the second, and microorganisms in the third. The first three chambers are mixed by manual cranking once the hardware reaches the International Space Station (ISS). Forty-eight hours later, the experiment is terminated by a second turning of the crank to further advance the plunger, bringing all four proximal stoppers together and adding the 0.5 mL of paraformaldehyde to the other solutions as a fixative. The now-stabilized FPAs can be returned to ground and OD620 measured as a quantification of microorganisms.
The exact configuration and design of the clinorotation device employed has been described in detail previously (Hoehn et al., 2004; Hammond et al., 2009). To match the conditions for spaceflight and ground controls as much as possible, the FPAs were loaded into the same GAPs used in spaceflight. The GAP was mounted in a clinorotation device such that it rotated around its central longitudinal axis (Fig 1). The FPA thus moved in a circle around the GAP's axis of rotation, as opposed to each FPA rotating on its own axis. The GAP had a diameter of 10.2 cm, and FPAs rotated around the center axis at a radius of 3.5 cm. The axis of rotation was perpendicular to Earth's gravitational field, resulting in a complete randomization of the gravity vector. Given the rotation axis, and a rotation speed of 5 rpm, cells in the FPA would experience a centripetal acceleration of approximately 9.7×10−4 g (Klaus et al., 1998). Centripetal motion of the nematodes or bacterial cells would be negligible under such forces compared with nonrotating, unit gravity controls. This clinorotation model approximates one aspect of microgravity in terms of preventing net sedimentation without stirring. Constant reorientation of the culture effectively nullifies cumulative sedimentation of the bacteria but does not necessarily result in uniform distribution of the suspended organisms (Klaus et al., 1998; Klaus, 2001). Furthermore, when rotating particles of differing sizes or densities, such as bacteria with nematodes, the rotation rate cannot be set so as to maintain both components in a motionless state; there will be varying degrees of relative motion between the different components and the fluid (Horneck et al., 2010). We used conditions that have previously been shown to optimally maintain bacteria in suspension in this specific hardware (Klaus et al., 1998). The rotation rate was determined by using the equations linking the density of the particle and the medium, the viscosity of the medium, the effective Stokes radius of a bacterial cell, and the radius of the container (Klaus et al., 1998). Although the nematodes are far larger than the bacteria, our model does not rely on the forces of the clinorotation apparatus to keep the two colocalized. The nematodes are highly motile, which keeps them in suspension and allows them to swim freely toward the bacteria in response to chemotactic signals.
2.4. Assay details
Each experiment was set up in triplicate. One set of GAPs was flown in space on the ISS. Ground controls included a second set of GAPs rotated on a clinorotation device to maintain cells in suspension, and a third set of GAPs was maintained under static conditions on the ground. Spaceflight and ground controls were performed in identical hardware and with identical timing, except that the ground control procedures were offset 30 min to allow for any delays in astronaut communications from the ISS. All experiments were matched to spaceflight ambient temperature. Specific details of nematode management, buffers, and procedures, including feeding, bacterial growth controls, and optimization of the assay, are detailed in our methods paper (Hammond et al., 2009). Figure 2 summarizes the experimental design. In brief, the first chamber, next to the gas exchange membrane, contained 2 mL of S-basal medium with cholesterol plus 5000 N2 C. elegans eggs. In ground controls >80% of the worms hatched. In experiments designed to measure virulence toward larval worms, no additional food was provided in the first chamber, which causes the hatched eggs to arrest at the L2 life-cycle larval stage. In experiments designed to measure virulence toward adult worms, the first chamber also included 1×109 heat-killed E. coli. This food source allows the eggs to progress to the L3/L4 stage of their life cycle during the 2–3 day period before they were mixed with the test microorganism. The quantity of E. coli was adjusted so that this food source was depleted approximately one-half day before the test microorganisms were introduced to the worms. By culturing the worms at ambient temperature, as opposed to the usual 30°C used in most C. elegans studies, we were able to slow their growth, thereby minimizing egg laying and completion of the life cycle within the interval of the virulence assay. To measure the growth of test microorganisms in the absence of worms, control FPAs were also set up with just 2 mL of S-basal medium plus cholesterol adjacent to the gas exchange membrane. The second chamber contained 1 mL of growth medium for the microorganisms. Brain heart infusion (BHI) was used for MRSA and Listeria, BHI with kanamycin was used for Enterococcus, and yeast extract peptone dextrose was used for Candida. The third chamber contained 0.75 mL of phosphate-buffered saline with 1×107 live microorganisms in static phase. In feeding control studies, heat-killed E. coli replaced live microbes. The last chamber contained 0.5 mL of 12% paraformaldehyde fixative.
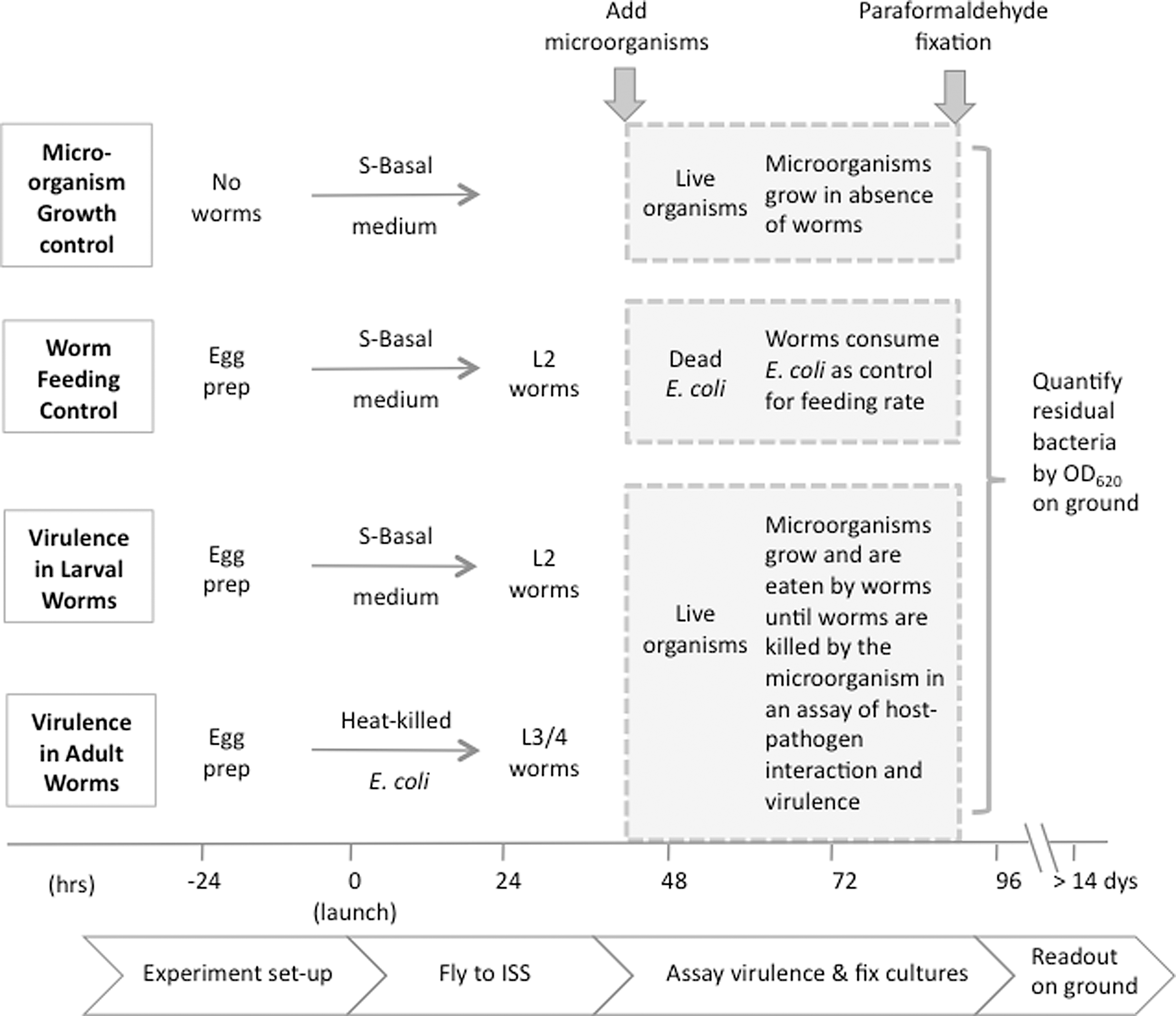
Design and timeline of the four experiments. Growth controls were set up with live microorganisms grown in the absence of worms. Worm feeding controls were conducted with L2 larvae and heat-killed E. coli. Virulence in adult and larval worms was evaluated by preparing C. elegans eggs 1 day before launch. All GAPs were loaded 1 day before launch and passed off. Eggs hatching in basal media are growth arrested at the L2 stage, whereas larvae hatching in the presence of killed E. coli as a food source can mature into L3/4 larvae. Virulence assays were activated approximately 40 h after launch and terminated by fixation after an additional 48 h. The timeline reflects hours prior to and after launch, which is defined as zero.
Approximately 40 h into spaceflight, the GAPS were plunged to mix the worms, growth media, and test microorganisms. The microorganisms and worms were allowed to interact for 48 h at ambient temperature. The cultures were then fixed by further advancing the plunger to introduce paraformaldehyde through the same bypass channel. Fixed cultures were stored at ambient temperature on the ISS until returned to ground for analysis by spectrophotometry and light microscopy. Assays of virulence for Candida, Listeria, and Enterococcus were done concurrently in spaceflight, static ground, and clinorotation. Assays for MRSA virulence were done in two parts: spaceflight with matched static ground control and, at a separate time, clinorotation with matched ground control.
The primary evidence that this assay has a broad dynamic range to observe virulence changes is derived from our earlier demonstration (Hammond et al., 2009) that nonpathogenic E. coli killed few worms in this assay, while Salmonella typhimurium killed nearly all the nematodes. In the present studies, post-spaceflight examination of worms grown with microorganisms revealed a mixture of C. elegans with curved shapes and needle shapes (data not shown). The needle shape is characteristic of dead worms, whereas the curved shape is characteristic of live worms. This result verifies that some worms survived the 48 h co-culture with microorganisms and were alive at the time that paraformaldehyde was added. Thus, our virulence assay was conducted within the dynamic range with both dead and live worms present at the end of the co-culture with microorganisms.
2.5. Statistics
Statistics were performed by using the Student two-tailed t test. Data are presented as the mean±1 SEM. Assays were performed in quadruplicate unless otherwise noted.
3. Results
Figure 3 compares the ability of C. elegans to consume nonpathogenic microbes in static/ground, spaceflight, and clinorotation cultures. All three conditions were inoculated with killed E. coli at an optical density (OD) of 0.24. The OD620 after 48 h was 0.128±0.006 for ground control, 0.114±0.005 for spaceflight, and 0.124±0.003 for clinorotation. The three results are not statistically different, verifying that the intrinsic ability of C. elegans to ingest microbes was not affected by spaceflight or clinorotation. This control illustrates that the freely motile nematodes were able to scavenge bacteria equally well whether the microorganisms were sedimented on the floor of the culture vessel in a static ground-based assay or dispersed through the medium in the microgravity of spaceflight or maintained in suspension through rotation.

Feeding controls in spaceflight and clinorotation. The ability of C. elegans L2 larvae to ingest killed E. coli is shown in static/ground, spaceflight, and clinorotated cultures. All three conditions were inoculated with killed E. coli at an OD620 of 0.24. After 48 h, the cultures were fixed with paraformaldehyde, and the quantity of remaining E. coli was estimated by the OD620. Error bars indicate±1 SEM of quadruplicates.
Figure 4 illustrates the virulence of the four microorganisms toward nematodes in static/ground condition, clinorotation on ground, and microgravity of spaceflight. When cultured alone as growth controls, the four microorganisms grew in spaceflight or clinorotation as well as or better than they did in matched ground-based cultures (Table 1). Clinorotation induced a striking increase in the growth of Candida (2.4-fold increase in OD620 relative to ground controls), and spaceflight induced a striking increase in the growth of MRSA (2.3-fold increase in OD620 relative to ground controls). A comparison of worm/microorganism co-cultures with the growth controls revealed that the raw OD620 of microorganisms incubated with worms was higher than the raw OD620 of microorganisms cultured alone (Table 1). This does not reflect light absorption by the worms, as they do not absorb light at this wavelength (data not shown). Instead, it appears the increased OD620 in the microorganism/worm mixtures reflects debris from worms that have been killed by the microorganism. We therefore calculated, for each condition, the difference in OD620 for microorganisms mixed with worms minus the OD620 for microorganisms cultured alone. Table 1 provides the raw data used to calculate the delta (Δ) OD620, and Fig. 4 is based on those ΔOD620. We used the delta (Δ) OD620 as an index of virulence to compare spaceflight or clinorotation conditions with static/ground controls (Fig. 4). For example, if microgravity increased virulence, we would expect to see a greater positive ΔOD620 in the spaceflight samples due to both more rapid killing of C. elegans to generate worm debris and to the greater numbers of unconsumed microorganisms that continue to multiply during the incubation period.
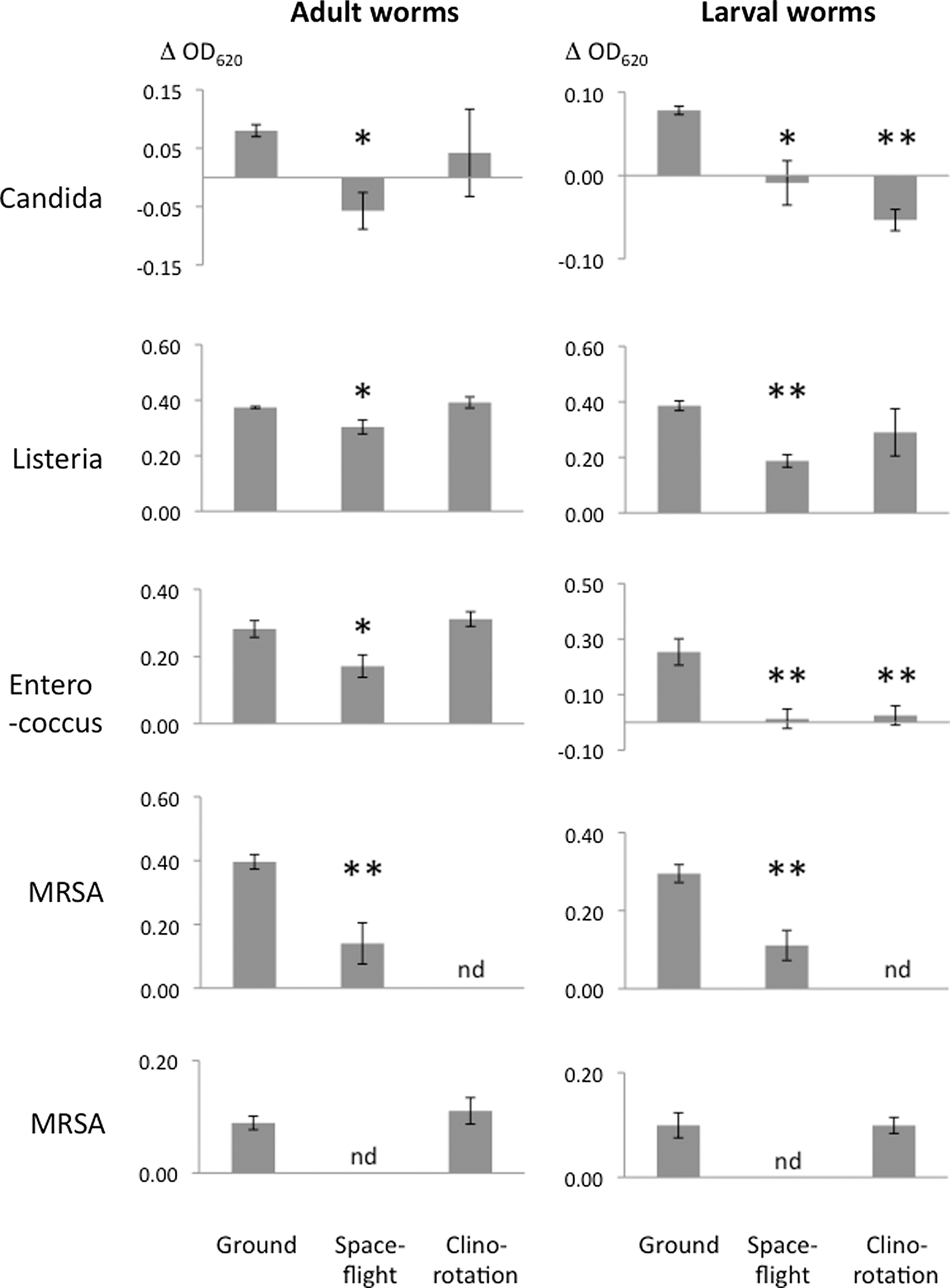
Virulence of four microorganisms toward adult and larval worms in ground/static, spaceflight, and clinorotation. Microorganisms were cultured with nematodes in identical hardware under static conditions, clinorotation, and spaceflight and fixed with formaldehyde after 48 h of co-incubation. Delta (Δ) OD620 values are the OD620 for microorganism in the presence of larval or adult worms minus the OD620 for microorganism cultured alone. Error bars indicate±1 SEM. The t test was used to estimate the significance between the ΔOD620 for spaceflight versus static assays or clinorotated samples versus static assays. * indicates p<0.05, ** indicates p<0.01. Assays with larvae were conducted in quintuplicate; assays with adult worms or medium alone were in quadruplicate with the exception of Listeria in clinorotation where one sample was lost, leaving triplicates, and MRSA where all assays were in quintuplicate. The positive ΔOD620 with the static control suggests that there is some debris generated when microorganisms are incubated with larvae or adult worms (or that the microorganisms grew better in the presence of C. elegans). The smaller ΔOD620 seen in spaceflight indicates more consumption of microbes and/or less generation of debris under spaceflight. Either explanation is consistent with decreased virulence. Assays of virulence for Candida, Listeria, and Enterococcus were done concurrently in spaceflight, static ground, and clinorotation. Assays for MRSA virulence were done in two parts: spaceflight with matched static ground control and, at a separate time, clinorotation with matched ground control. nd=not done.
OD620
OD620 of microorganisms grown in medium alone (growth) or after addition into cultures of larval or adult worms. Cultures in identical hardware were maintained under static/ground, spaceflight, or clinorotation conditions. After 48 h of growth, samples were fixed with paraformaldehyde. Values are the mean±1 SEM. Assays with larvae were conducted in quintuplicate; assays with adult worms or medium alone were in quadruplicate with the exception of Listeria in clinorotation where one sample was lost, leaving triplicates, and MRSA where all assays were in quintuplicate. Assays of virulence for Candida, Listeria, and Enterococcus were done concurrently in spaceflight, static ground, and clinorotation. Assays for MRSA virulence were done in two parts: spaceflight with matched static ground control and, at a separate time, clinorotation with matched ground control.
For each of the microorganisms, when cultured with adult or larval worms, the ΔOD620 for spaceflight microgravity cultures were significantly lower than for the corresponding static/ground controls. This indicates more consumption of the microorganism and/or less generation of debris under spaceflight conditions. Either explanation is consistent with decreased virulence under microgravity.
When assayed under clinorotation conditions, Candida and Enterococcus were less virulent for larval worms, matching what was seen in spaceflight. However, the virulence of MRSA and Listeria for larval worms was unchanged by clinorotation, which differs from what was seen in spaceflight. Clinorotation results for adult worms are even less consistent with spaceflight, as virulence of Enterococcus, Listeria, and MRSA was unchanged by clinorotation. This is in direct contradistinction to the decreased virulence seen in concurrent samples tested in spaceflight, as summarized in Table 2. Clinorotated cultures of Candida with adult worms showed such wide variability between replicates that it was not possible to determine whether they were significantly different from ground/static controls.
The effect of spaceflight and clinorotation on virulence of four microorganisms for adult and larval worms is summarized along with the morphology of the microorganisms. Statistical significance was estimated by two-tailed Student t test. ⇓indicates statistically reduced virulence compared to static/ground controls.⇔indicates virulence that is not statistically different from static/ground controls. CNE=could not evaluate differences in virulence due to wide variability in replicates.
Previous studies have shown that Candida grown in a rotating wall vessel to maintain cells in suspension under quiescent conditions increase the rate at which they transition from a single cell to a filamentous phenotype and form biofilm communities (Altenburg, 2008; Searles et al., 2011). These dimorphic shifts are associated with significantly greater resistance to the antifungal agent amphotericin B and changes in the expression of two genes associated with the yeast-hyphal transition. To evaluate whether spaceflight or clinorotation induced dimorphic shift in our Candida cultures, we examined aliquots of fixed samples by light microscopy (Fig. 5). Yeast from static/ground samples grew as single cells, whereas yeast from clinorotation and spaceflight conditions showed a dimorphic shift with branching and formation of filaments.
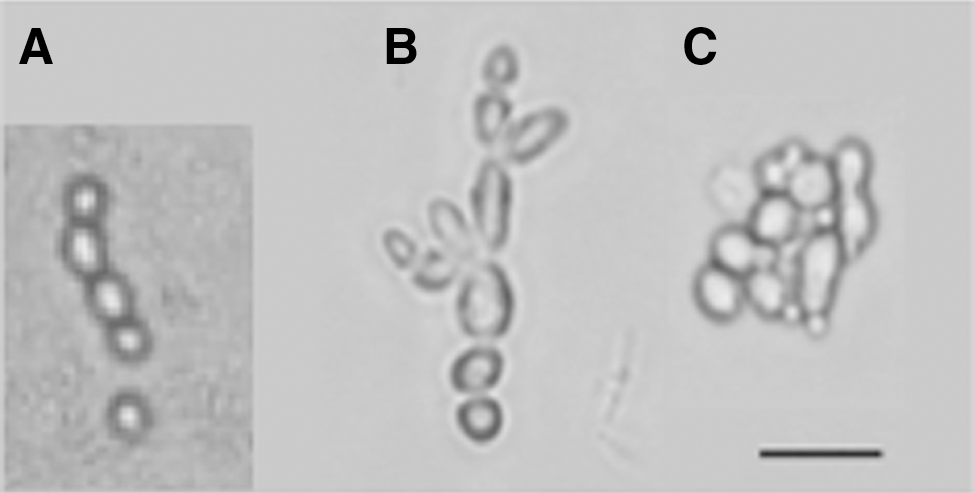
Dimorphic shift of Candida under spaceflight and clinorotation. (
4. Discussion
Results of the current experiments directly demonstrate that spaceflight is associated with reduced virulence for diverse microorganisms, including Listeria, Enterococcus, Candida, and MRSA. These are the first data acquired with a direct in vivo assay system in space to demonstrate virulence (Hammond et al., 2009). By employing the C. elegans host-pathogen model, we can assay virulence in spaceflight and terminate the assay with fixative for subsequent ground-based analysis. This eliminates any confounding variables associated with reentry and delays. In this report, we have verified that neither spaceflight nor clinorotation changed the feeding rate of nematodes so that changes in bacterial consumption accurately reflect virulence.
In orbit, net forces on an object (gravity and centrifugal acceleration) are effectively nulled out, resulting in a perpetual free-fall condition. In low-Earth orbit, only a small residual force, generally referred to as microgravity, remains. In contrast, clinorotation randomizes the influence of gravity so there can be no net directional acceleration or force acting on an object. Residual accelerations for the clinorotation device used in this study are less than 10−3 g, which, according to Stoke's Law, means that nonmotile bacteria will move through the media at a rate 1000-fold less than bacteria in a static system. This is important, as the magnitude of the applied g-force determines biological outcome in some systems (Brown et al., 1976). Both culture modalities, space-based and clinorotation, minimize motion in a suspension culture and prevent the microorganisms from sedimenting. Caenorhabditis elegans, by contrast, are highly motile organisms and can remain in suspension in any of these conditions. We cannot exclude effects of the transient effects of launch: 8 min and 30 s at a maximum acceleration of 3.4g hypergravity, as well as mechanical vibration. But any such effects would likely be reversed by the time the FPAs were activated in orbit, two and a half days after launch.
Caenorhabditis elegans display an innate, or immediate, immune response and share many cellular and molecular structures and control pathways with higher organisms (Harvill and Miller, 2000; Sifri et al., 2005). For example, they are able to produce antimicrobial peptides and enzymes in response to microbes (Ewbank, 2006). A brief exposure to bacteria “immunizes” the worms and allows them to survive a subsequent exposure that would otherwise prove lethal, a phenomenon referred to as “conditioning” (Anyanful, 2009). Caenorhabditis elegans do not have a true adaptive or secondary immune response that higher-order organisms generate upon repeated exposure to the same microbe. Nonetheless, several studies have shown good concordance between the virulence of Salmonella assayed in C. elegans and virulence in the mouse systemic infection assay (Paulander et al., 2007; Jelsbak et al., 2012).
We used both larval and adult worms as targets in our assays for two reasons. There are data that suggest that the life-cycle stage of the worm influences the immune responses it can generate. Arresting cell cycle at L2 by starvation allows for the nematodes to be well synchronized. Feeding the nematodes allows for the study of more mature forms but risks progression to egg production and loss of life-cycle synchrony. The adult experiments were important to allow microorganism exposure to a mature pharynx, which can be an important portal of bacterial entry (Sifri et al., 2005; Tenor and Aballay, 2008).
Spaceflight has been shown to have other effects on host-pathogen interactions (Bhattacharya et al., 2011). For instance, even short-term spaceflight affects cellular and humoral immunity, phagocytic functions, and the maturation of immune cells in Drosophila (Bhattacharya et al., 2011). This is important as, although heavily touted as a microbial virulence model (Harvill and Miller, 2000; Sifri et al., 2005), C. elegans lacks adaptive immunity. In higher-order species such as mammals, lymphocytes, which are the cellular mediators of adaptive immunity, have blunted activation during spaceflight (Cogoli et al., 1993; Hughes-Fulford, 2011), and T cell response is lost (Cogoli et al., 1993), implicating gravity as a necessary factor in normal immune function.
Unlike spaceflight, clinorotation did not affect virulence of Listeria or MRSA for either C. elegans larvae or adults and did not affect the virulence of Enterococcus for adult worms. Clinorotation did reproduce spaceflight-induced reduction in the virulence of Candida and Enterococcus for larval worms. High variability in the replicates for Candida assayed with adult worms did not allow us to conclude whether clinorotation reduced virulence or not. Thus, for these particular microorganisms, using shear to offset gravity did not consistently produce the same effect on virulence as did true spaceflight microgravity. This discordance may reflect variability in how different microorganisms are affected by the shear forces that are required to offset gravity in the clinorotation model.
Clearly, the decreased virulence of Candida in spaceflight is not what would have been predicted based on the observed dimorphic shift of the yeast during spaceflight, given that dimorphic shift is commonly associated with increased virulence (Altenburg, 2008; Searles et al., 2011; Gow et al., 2012). This suggests that there were other mechanism(s) to decrease virulence, such as intracellular genetic changes, which were more potent than the tendency to increase virulence as a result of dimorphic shift.
Benoit and Klaus (2007) reviewed the literature looking for an explanation as to why microgravity is associated with increased bacterial growth for many, but not all, bacteria. Their theory was that spaceflight indirectly affects growth by reducing the tendency of bacteria to settle out of liquid media and reducing the potential for buoyant convection in the vicinity of actively metabolizing bacterium. They postulated that the impact on microgravity would be less evident on motile bacteria that can remain dispersed throughout the liquid culture and actively stir the media in their microenvironment, whether on ground or in spaceflight. They reported that nine of nine studies with nonmotile strains (including Salmonella typhimurium, E. coli, and B. subtilis) showed increased growth in microgravity, whereas three of three studies with motile E. coli showed no difference in growth under microgravity. Motility also explained the variability in concurrence between results in spaceflight and results from clinorotation used to maintain microorganisms in suspension. Six of six nonmotile bacteria showed increased growth in clinorotation, whereas a motile strain showed no difference (Benoit and Klaus, 2007).
We postulate that motility and shear forces account, at least in part, for the discordance between spaceflight and clinorotation in our studies (Table 2). Listeria is a rod-shaped bacterium with flagella and a tumbling motility that may explain why clinorotation did not reduce virulence for adult or larval eggs. The other three microorganisms are nonmotile but are likely to adopt different profiles in microgravity. The Candida strain that we used is listed as anamorphic in the ATCC catalogue but clearly does form filaments under the culture conditions applied (Fig. 5). Hence, the filamentous Candida are several-fold larger than the bacteria we used and will incur larger shear and Coriolis forces in the clinorotation tubes. Enterococcus and Staphylococcus are both small cocci that propagate as sheets or clusters, respectively (Table 2). In summary, the effects of clinorotation on bacterial growth and virulence in the C. elegans model appear to be influenced by multiple variables, but these likely include motility and size-dependent shear and Coriolis forces incurred.
Virulence is an interplay between the microbe and the host, and both must be considered to understand changes in pathogenicity (Harvill and Miller, 2000; Sifri et al., 2005). Salmonella flown in space appear to be more virulent when returned to ground and inoculated into mice (Wilson et al., 2007), but this experimental design does not actually evaluate virulence in microgravity. It remains uncertain whether the increased virulence in those studies was due to microgravity exposure or to the effects of return to Earth, including reentry, space shuttle landing, and delay in removing samples from the orbiter for handling. These processes take more than the generation time of the bacteria in question and allow abundant opportunity for gene expression and translational and post-translational changes.
Changes in virulence during spaceflight are likely to reflect modulation of the C. elegans host as well as modulation of the microorganisms. Spaceflight is known to downregulate C. elegans genes, including those related to neuronal and endocrine signaling (Leandro et al., 2007; Selch et al., 2008; Szewczyk et al., 2008; Honda et al., 2012; Qiao et al., 2013). Inactivation of those genes on ground leads to increased longevity mediated through longevity-regulating transcription factors or through dietary-restriction signaling (Honda et al., 2012). It was suggested that the proteins function in processes related to sensory perception of food, signaling in response to food or food intake (Honda et al., 2012). We did not detect any reduction in the ingestion of killed E. coli during spaceflight as opposed to ground controls but cannot verify whether the ingestion of other microorganisms was impaired as a result of microgravity-induced modulation of C. elegans. It is also possible that microgravity affected the C. elegans' immunity. Genes downregulated by spaceflight encode proteins that act through DAF-16 and SKN-1 transcription factors (Honda et al., 2012). These are the same transcription factors involved in generation of C. elegans' antimicrobial peptides and lysozymes, as well as its MAPK pathway-mediated pathogen resistance (Millet and Ewbank, 2004). In the present report, all four microorganisms tested showed reduced virulence in spaceflight; however, we do not propose that this is purely the result of some spaceflight-mediated global enhancement of C. elegans' immunity. We have seen increased virulence in spaceflight when testing live E. coli tested against Tol1 deleted adult C. elegans (Hammond et al., 2013). We have also tested Salmonella and shown that some gene deletions significantly decrease the virulence of the Salmonella toward C. elegans, whereas other deletion strains are no more virulent in spaceflight than on the ground (Hammond et al., 2013).
Suspension culture devices continue to evolve in an effort to understand the physical parameters critical to specific biological applications for scientific and commercial applications (Hammond and Hammond, 2001; Brungs et al., 2011; Herranz et al., 2013). Our choice of a simple clinorotation device was both scientific and practical. At a scientific level, the device reproduces the quiescent, unstirred fluid conditions achievable on orbit in a system in which the forces have been cleanly quantified and defined (Klaus et al., 1998). At a practical level, we were flying large complex payloads, and the flight hardware virulence assay tubes (FPAs) in their encased cylindrical sets of eight tubes (GAPs) were facile to mount and rotate in the clinorotation device (Hoehn et al., 2004). Measuring virulence in spaceflight requires the use of biologicals that are rapidly perishable. Launches of the space shuttle were often delayed, and frequently with very late notice. When this occurred, there was often no more than 4 h of time in which to prepare a fresh set of assays and deliver them to the launch pad. To accommodate this, while awaiting launch, we prepared, every 2 days, sufficient reagents for all three conditions: static/ground, spaceflight, and clinorotation, as well as fresh worm stock. In anticipation of a launch, one of the three sets of FPAs was loaded with microorganisms, buffers, and fixative. When scheduled launch was imminent, one set of FPAs was loaded with worms and delivered to the launch pad. Once launch was confirmed, the static and clinorotation tubes were prepared with reagents from the same lot as the spaceflight. Future studies are planned to explore the full range of suspension culture options.
5. Conclusions
We conclude that spaceflight is associated with diminished virulence of several species of microorganisms. These data provide the first direct evidence for reduced virulence of motile and nonmotile microorganisms in microgravity. Differences in growth and virulence between space-flown and clinorotation samples are likely due, at least in part, to differences in applied shear forces, and motility of the microbes.
Footnotes
Acknowledgments
These studies were supported by SpaceHab/Astrotech and Astrogenetix Inc. We thank NASA for spaceflight access under the auspices of the International Space Station National Lab Pathfinder program. We thank Anthony Johnson for technical assistance.
This material is the result of work supported with resources and the use of facilities at the Durhan Veterans Affairs Medical Center and the Office of Research and Development, Department of Veterans Affairs, Veterans Health Administration. Contents do not represent the views of the Department of Veterans Affairs or the United States of America.
Author Disclosure Statement
None of the authors have any commercial associations that might create a conflict of interest.
Abbreviations
BHI, brain heart infusion; FPA, Fluid Processing Apparatus; GAP, Group Activation Pack; ISS, International Space Station; MRSA, methicillin-resistant Staphylococcus aureus; OD620, optical density at 620 nm.