
Abstract
Water is the fundamental molecule for life on Earth. Thus, the search for hibernating life-forms in waterless environments is an important research topic for astrobiology. To date, however, the organizational patterns containing microbial life in extremely dry places, such as the deserts of Earth, the Dry Valleys of Antarctica, or Mars analog regolith, have been poorly characterized. Here, we report on the formation of bacterial biosaline self-organized drying patterns formed over plastic surfaces. These emerge during the evaporation of sessile droplets of aqueous NaCl salt 0.15 M solutions containing Escherichia coli cells. In the present study, scanning electron microscopy (SEM) and energy dispersive X-ray spectrometry (EDS) analyses indicated that the bacterial cells and the NaCl in these biosaline formations are organized in a two-layered characteristic 3-D architectural morphology. A thin filmlike top layer formed by NaCl conjugated to, and intermingled with, “mineralized” bacterial cells covers a bottom layer constructed by the bulk of the nonmineralized bacterial cells; both layers have the same morphological pattern. In addition, optical microscopic time-lapsed movies show that the formation of these patterns is a kinetically fast process that requires the coupled interaction between the salt and the bacterial cells. Apparently, this mutual interaction drives the generative process of self-assembly that underlies the drying pattern formation. Most notably, the bacterial cells inside these drying self-assembled patterns enter into a quiescent suspended anhydrobiotic state resistant to complete desiccation and capable of vital reanimation upon rehydration. We propose that these E. coli biosaline drying patterns represent an excellent experimental model for understanding different aspects of anhydrobiosis phenomena in bacteria as well as for revealing the mechanisms of bacterially induced biomineralization, both highly relevant topics for the search of life in extraterrestrial locations. Key Words: Escherichia coli—Self-organized bacterial drying pattern formation—Hibernating life—Biosaline biomineralogical life formations—Anhydrobiosis—Biologically induced biomineralization (BIM). Astrobiology 14, 589–602.
1. Introduction
W
Desiccation refers to the extreme dehydration of cells when the hydration shell of molecules is gradually lost (Billi and Potts, 2000). It is well known that the removal of water through air-induced drying damages membranes, proteins, and nucleic acids and is lethal to the majority of organisms (Potts, 1994). Nevertheless, anhydrobiotic organisms such as certain vegetative cells of bacteria, cyanobacteria, and higher organisms, for example, tardigrades, can survive extreme desiccation (Potts, 1994; Billi and Potts, 2002; Jönsson, 2007). Anhydrobiosis refers to the state in which an organism stops its vital functions temporarily due to partial or total desiccation (Potts, 1994). This quiescent state is characterized by an extreme reduction of measurable metabolism (Billi and Potts, 2002). Desiccated cells do not grow, and the time during which they remain dried may represent the greater part of their life span. When rehydrated, anhydrobiotes revive and resume active metabolism. Interestingly, the maximal longevity of microorganisms in a metabolically inactive, desiccated state is currently unknown (Kennedy et al., 1994). Numerous cellular mechanisms responsible for enhanced desiccation resistance in bacteria have been identified, including de novo trehalose synthesis, production of extracellular polymeric substances (EPS), and uptake of exogenous glycine betaine (Billi and Potts, 2002).
Related to the previous issue, desiccation has other notable consequences as well for both abiotic and biotic systems. For instance, it is well documented that the evaporation of water drives the formation of the different self-organized patterns that emerge in drying conditions, such as in small droplets containing different kinds of solutes (Deegan, 2000). Drying dissipative patterns have been studied for many kinds of suspensions and solutions of colloidal particles, linear-type synthetic and bio-polyelectrolytes, water-soluble neutral polymers, ionic and non-ionic detergents, gels, colloidal polymer-complex, and dyes (Kaya et al., 2010; Okubo, 2011). In addition, drying patterns have been observed with biological macromolecules, for example, salt solutions of DNA (Jing et al., 1998; Okubo et al., 2010) and proteins (Shiba et al., 2003; Chen and Mohamed, 2010; Ma et al., 2013) and human fluids (e.g., blood, Brutin et al., 2011). However, despite the fact that bacteria are the most abundant cellular organisms that thrive on Earth (Staley et al., 2007) and numerous fundamental biological issues have been decoded and unraveled when using these organisms as biological models, the study of drying patterns and bacteria has not been previously reported.
During the motility study of Escherichia coli cells inside a simple microcosmos, a droplet of NaCl physiological saline solution (9 mg/mL, 0.15 M), we found unexpectedly that bacterial cells underwent self-assembly in conjunction with the NaCl (driven by water evaporation) that produced complex biosaline drying patterns on plastic surfaces. In this way, the evaporating water generates regular patterns with a good reproducibility for the several common wild-type E. coli K-12 laboratory and mutant strains assayed in this study (see Materials and Methods).
In this work, we characterize the macroscopic and microscopic structural organization of these bacterial drying biosaline formations and report the reversible vitality exhibited by the bacterial cells that form these patterns. Finally, we discuss the significance of these findings from an astrobiological perspective, which could be important for the search for hibernating extraterrestrial resistant organisms. Additionally, we suggest that these patterns might serve as biomarkers detectable by imaging systems during planetary exploration.
2. Materials and Methods
2.1. Escherichia coli strains, media, and growth conditions
The Escherichia coli MG1655 K-12 (Blattner et al., 1997; Dragosits et al., 2013), MG1655 strR (Gauger et al., 2007), W3110 (Khankal et al., 2009), and NMHK8 (BW25113 sdiA::cat) (Høyland-Kroghsbo et al., 2013), NM525 and GSO553 (NM525 ΔflhDC::kan) (Thomason et al., 2012) strains were used in this work. The media used was Luria-Bertani (LB) medium (Gómez-Gómez et al., 1996): 1.0% (10 g/L), Difco Bacto-tryptone (Lot n°3113075), 0.5% (5 g/L), Difco yeast extract (Lot n°6257032) and 0.5% (5gr/L NaCl Merck, Lot n°1064041000). The LB medium was jellified with Agar Bacteriológico Europeo (ABE, Lot n°15100184) at the concentration indicated. The physiological saline solution of NaCl (9 mg/mL, 0.15 M) employed was commercially made and distributed by the Fresenius Kabi España S.A. company under the name Cloruro de Sodio Meinsol. Similar results were obtained with a solution of NaCl (obtained from VWR Chemicals) prepared with Milli-Q water.
2.2. Experimental setup to study biosaline bacterial drying patterns
The experimental protocol for droplet evaporation was carried out with the following procedure: the different E. coli K-12 strains used in this work were grown in Petri dishes prepared with LB ABE agar 1.5% (15 g/L). The cells obtained from a colony grown from these plates were inoculated with a toothpick at the center of an 8.5 cm Petri dish containing 30 mL of liquid LB medium, harnessed with 0.6% (6 g/L) ABE agar. The plates were sealed with Parafilm to prevent loss of water and cultivated at 37°C. The plastic Petri dishes made of polystyrene are manufactured by the Sterilin company (
The in vitro setup was established as follows: approximately 5×109 bacterial cells from an E. coli K-12 strain macrocolony biofilm grown at 37°C during 14 days, similar to the one shown in Fig. 1A (the colonial pattern-morphotype exhibited by these colonies was described by Gómez-Gómez and Amils, 2014), were carefully removed from the agar plate and suspended into a droplet (200 μL) of physiological salt NaCl solution (9 mg/mL, 0.15 M) that was placed on the plastic surface of a Petri dish. This simple experimental microcosmos simulates environmental situations where the continuous supply of nutrient inflow is insufficient to support active bacterial growth, and the bacterial behavior observed necessarily might depend exclusively on bacterial motility. The evolution in appearance and behavior of this bacterial community inside the droplet at room temperature (<25°C) was observed directly under the objective of an Ultralyt ULNM-90-10000 microscope, which was coupled to a digital camera, over that period of time sufficient for complete droplet desiccation. The plates were photographed with reflected light with a Kodak Easy Share Z710 digital camera.
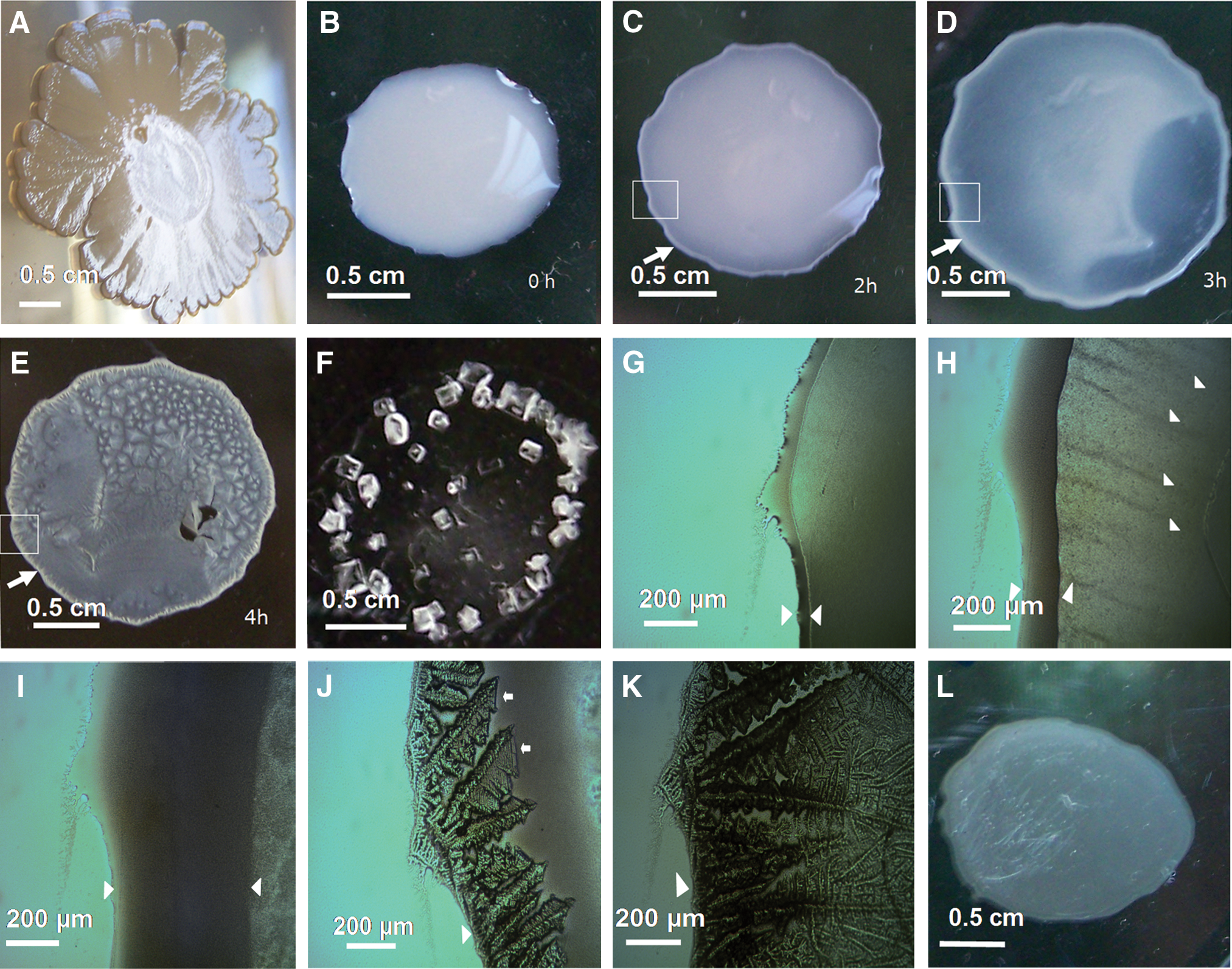
Macroscopic and optical microscopic study of E. coli saline drying patterns in aqueous solution of NaCl 0.15 M on plastic Petri dish surfaces. (
Figure images were prepared with the Microsoft Photo Editor program and Microsoft PowerPoint software.
2.3. Cell viability assay
Cells extracted from the drying biosaline droplets were resuspended in 200 μL of fresh LB medium, and 100 μL of this was used to prepare serial dilutions in LB medium. One hundred microliters obtained from the appropriate dilutions was spread in Petri dishes prepared with 30 mL of LB medium harnessed with 1.5% ABE agar (15 g/L). The plates were then incubated overnight at 37°C, after which the colonies were counted. During each day, as shown in Fig. 6P, the viability of three independent drying biosaline patterns was measured.
2.4. Scanning electron microscopy and energy dispersive X-ray spectrometry
The scanning electron microscopy (SEM) and energy dispersive X-ray spectrometry (EDS) analyses were performed with a FEI-Quanta 200FEG electron microscope that used an acceleration voltage of 5.00 keV. The scanning electron microscope images were obtained with scanning modes by the detection of secondary electrons and backscattered electrons.
3. Results
3.1. Motility behavior of E. coli cells inside the saline droplet (0.15 M NaCl) prior to formation of the drying pattern
To inhabit and maintain a population in soil, E. coli must have the capacity to resist soil environmental stresses, one of which is the desiccation stress associated with natural soil wetting-drying cycles (Zhang and Yan, 2012). To learn how E. coli cells respond to the exposure of water evaporation under in vitro controlled conditions, the E. coli cells obtained from the colonies grown in semisolid LB agar (Fig. 1A) were resuspended in small droplets of a NaCl 0.15 M solution. Monitoring the evaporating droplet under the microscope revealed that several minutes (∼5–7 min) after their inoculation, the bacterial cells rapidly began an active accumulation at the “edge” of the droplet (Fig. 1B) that formed the exterior boundary along the perimeter of the droplet (Fig. 1C, 1D). This edge increased its thickness with time through a massive bacterial accretion driven apparently by a process of the cell-to-cell aggregation (Fig. 1G–1I). Furthermore, we observed the generation of the fine bacterial “streams” forming stripes aligned in the direction of the edge of the biofilm inside the droplet (Fig. 1H and see Movie S1 in the Supplementary Material, available online at
In this respect, it is well known that swarming is a form of collective motility. Thus, during swarming motility, the E. coli “swarmer” cells carry out a collective movement (i.e., this kind of bacterial migration is a social rather than individual trait, Copeland and Weibel, 2009) over solid surfaces (generally over semisolid agar surfaces with nutrients supporting the bacterial growth and swarming movement) and participate in the formation of coordinated patterns of group activity that emerge as a consequence of the collective motion, which is something akin to the behavior exhibited by a swarm of bees (Harshey, 1994). The E. coli swarming behavior is driven by flagella and requires metabolically produced energy, which is necessary to power the flagellar rotatory motors that control the rotation of the flagella that propel the bacteria in this active movement (Gómez-Gómez et al., 2007; Kearns, 2010).
To determine whether this collective bacterial movement observed in drying, evaporating droplets is an active process that requires flagella, we examined the behavior of 2.5×109 E. coli NM525 ΔflhDC::kan cells loaded in a NaCl 0.15 M droplet (this E. coli strain lacks the flagellar master regulator FlhDC and is thereby unable to produce flagella, Thomason et al., 2012) in comparison to its wild-type NM525 motile strain under the microscope. The swarming-like stream patterns generated by the collective motion of the E. coli wild-type motile strain were not observed in this mutant strain that lacks the flagella (data not shown), which indicates that flagella are required for the emergence of stream patterns of bacterial collective movement within the NaCl evaporating droplet. These results appear to rule out the hypothesis that forces of nonbiological origin generate “patterns of flow” in the fluid (e.g., convective streams) of the droplet that drag the bacteria and mimic the illusion of an active bacterial collective motion. Here, we suggest the possibility that the emergence of this collective streaming bacterial movement, under these crowded bacterial conditions in drying NaCl droplets, could be interpreted as a kind of swarming-like collective behavior. It is clear, however, that additional future experiments will need to be carried out to clarify this hypothetical supposition.
Intriguingly, for the E. coli NM525 ΔflhDC::kan mutant that is defective in the production of flagella, a residual bacterial movement was observed that generated an edge in the droplet Interestingly, E. coli movement on a surface mediated by type 1 pili has been recently described (Kurihara et al., 2011). These results along with our findings suggest the possibility that several active motility modes (flagellar and nonflagellar dependent requiring energy) could be acting in evaporating droplets. However, the possibility that passive movement (i.e., random Brownian motion) could occur in conjunction with active bacteria-produced movement to generate, as a final net outcome, the global bacterial collective movement observed in evaporating droplets cannot be completely ruled out. Future experiments will clarify the nature of the phenomenon of collective bacterial movement in drying saline droplets.
3.2. Complex bacterial biosaline drying pattern formation on plastic surfaces
The evolution of bacterial movement inside the NaCl-saline droplet was monitored during approximately 4 h at room temperature. After this elapsed time, the water evaporation triggered a rapid process of salt crystallization. This process was initiated suddenly from the edge of the biofilm (Fig. 1J) and advanced along the periphery of the droplet to completely cover the bacterial edge (Fig. 1K). The ongoing water evaporation generates a centripetal drying pattern formation process toward the center of the biofilm (Fig. 1J and see Movie S2 in the Supplementary Material) in a manner such that, after a few minutes, a complete biosaline pattern of the border is achieved for the whole of the droplet (Fig. 1E). It was also observed that this process apparently dragged the self-assembly of the E. coli bacterial cells into complex drying patterns (Fig. 1J). In addition, different foci of pattern formation are observed in the interior of this class of biofilms and show, in some cases, confluence in their borders (Fig. 1E and Fig. 2E, 2F). That is, the bacterial cells underwent a process of crowding and aggregation that was salt-induced and driven by water evaporation, leading to the formation of a “dried salt-entrapped bacterial tapestry” composed of distinct “E. coli biosaline drying patterns.” To our knowledge, this kind of bacterial biosaline self-organized pattern has not been reported before. Here, in the context of this work, self-organization means that the acquisition of order and structure during the pattern formation process occurs through interactions internal to the system, without intervention by external directing influences. In other words, the self-organized pattern is an emergent property of the system, rather than a property imposed on the system by an external ordering influence (Ball, 1999; Camazine et al., 2001).

Typical microscopic biosaline drying patterns observed in dried E. coli biofilms and typical abiotic inorganic NaCl crystal. (
On the other hand, in the Petri dish control plates containing just the NaCl salt but no bacteria, only the faceted, blocky shapes of typical NaCl abiotic crystals were observed (lattice constant: 5.65 Å) (Fig. 1F and Fig. 2L). In addition, in the absence of NaCl, no patterns of this kind are generated by the bacterial cells alone (Fig. 1L). Therefore, both bacterial cells and NaCl are necessary for these biosaline drying patterns to emerge.
The appearance of microscopic drying patterns changes drastically depending on the location in the dried film (Fig. 1E); however, a number of different patterns can be observed. We briefly describe the most common one, a pattern that exhibits fractal-like growth and exhibits a characteristic cross-symmetrical organization (Fig 2A–2K). In the center of this pattern there is a typical morphology of a NaCl crystal (Fig. 2A, 2B, 2F–2I, and 2K), indicating that the cross-symmetry exhibited by this pattern is inherited from the cubic habit of an initial nucleating founder (Adam) NaCl crystal. Furthermore, the patterns have a rich internal morphological structure that changes with the distance from the nucleating center of the pattern. Thus, in the presence of bacterial cells, an Adam NaCl crystal develops dendritic structures that arise initially from the corner of the cubic formation to create a characteristic fractal-like pattern (Fig. 2H). A transition zone is observed where the pattern changes from fractal to bushy fern-leaf morphology. Therefore, the different zones can be identified in this complex pattern and are denoted as follows: the nuclear core (nc), fractal-like zone (fl), and a fern-leaf bushy zone (fb), which terminates in the boundary of the pattern (bp) that forms in the confluence zone with other similar patterns (Fig. 2F and 2I, 2J). In addition, in this kind of pattern, there are zones where a complete confluence among the dendritic structures can be observed (Fig. 2E), as well as zones where this contact does not occur (Fig. 2F and 2I, 2J).
Interestingly, the drying biosaline patterns generated by the E. coli NM525 ΔflhDC::kan strain are similar to those observed in wild-type strains (for instance, cross-symmetrical patterns similar to those shown in Figs. 2 and 3 are observed in drying droplets of this E. coli strain; data not shown), indicating that during the process of biosaline drying pattern formation per se, flagella and flagella-driven motility are not factors involved directly in its formation.

Scanning electron microscope images of a typical nucleating core exhibiting a cross-symmetry fractal-like E. coli biosaline drying pattern. (
The dynamics of the NaCl+bacteria pattern formation process was studied with time-lapsed video microscopy. The movies obtained during the droplet drying process (see Movies S2 and S3 in the Supplementary Material) demonstrate that the biosaline drying pattern formation is a kinetically fast process; typical snapshots extracted from these videos are shown in Fig. 1J and 1K and Fig. 2B–2E. The time-lapse Supplementary Movie S3 shows the formation of a typical biosaline pattern that exhibits cross-symmetrical geometry. Thus, given depictions as seen in these movies, we suggest that the pattern formation is a coupled process that involves the interaction of the bacterial cells and the NaCl salt, which leads to a simultaneous self-assembly of both biotic and abiotic components in an integrated biosaline self-organized drying structure.
Taken together, these results suggest that, although the bacteria apparently do not disrupt the initial NaCl nucleation process to form a primary NaCl crystal, they have the capacity to interfere with the “normal” abiotic inorganic formation of the NaCl crystal by modulating the subsequent growth and development of the NaCl crystal pattern that develops into a complex “organic crystal habit” (Ball, 1999).
3.3. Scanning electron microscopy studies and energy dispersive X-ray spectrometry analyses of the biosaline drying patterns
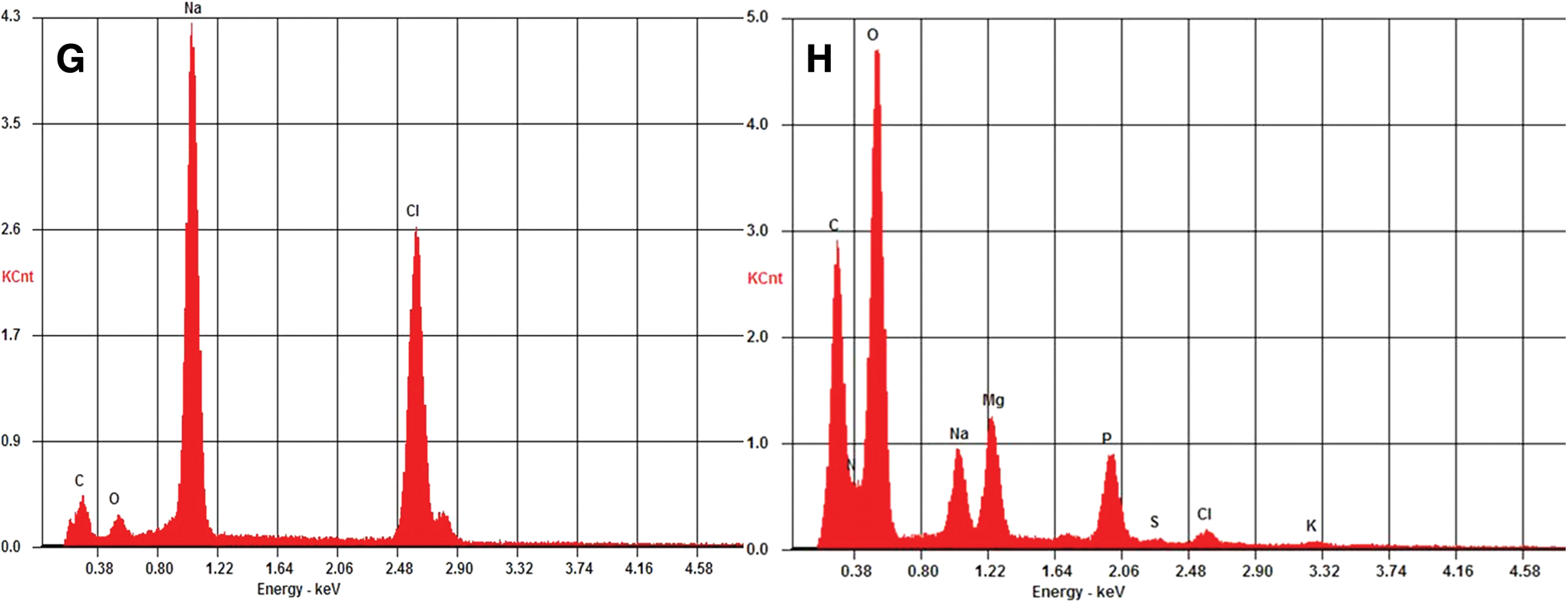
To better understand the microscopic architectural organization of the E. coli biosaline drying patterns, we studied them with SEM and EDS analysis. The scanning electron microscope images of the biosaline drying pattern observed in several E. coli strains—MG1655 (Fig. 3), NMHK8 and W3110 (Figs. 4 and 5)—indicate that the biosaline patterns have a complex 3-D architecture such that the NaCl and the bacterial cells share the same morphological pattern and possess a characteristic organizational mode. Indeed, the NaCl as determined by EDS analysis (Fig. 5G) is conjugate to, and intermingled with, the “mineralized” bacterial cells (Fig. 5F); thus a thin filmlike layer that constitutes the biomineralogical material is formed (Fig. 5G, 5H). Remarkably, as shown in Fig. 3H, the NaCl+bacteria interaction is extremely complex in the fractal line zone of the drying patterns, forming structures without a clear distinction between both components. Beneath this thin filmlike biomineralogical layer is the bulk of the drying pattern, which is constructed completely by bacterial cells (Fig. 4C).

Scanning electron microscope images of a typical drying E. coli bacterial-salt fern-leaf pattern. (

Scanning electron microscope images showing the appearance of a film layer with a fern-leaf morphology containing mineralized cells, and EDS analysis of relevant regions in this pattern. (
Taking all these results into account, it is tempting to suggest that the NaCl can apparently be conjugated with bacterial cells growing its “organic habit” along the “scaffold” provided by the bacterial cells that self-assemble into a specific integrated bacteria+NaCl salt pattern. That is, the bacteria participate passively during the formation of the biosaline drying patterns and apparently provide a scaffold where the NaCl deposition and conjugation are carried out. In addition, these observations suggest that the bacteria as a whole have the ability to modulate crystal growth but also that the NaCl modulates reciprocally the formation of the bacterial pattern.
Thus, although the drying pattern formation requires both the bacterial and salt components, we cannot confirm with the available data that the bacteria carry out an active sensu stricto role, that is, whether metabolic energy is used per se by the bacterial cells specifically during the process of construction of the drying biosaline patterns, for instance, to generate movement of the Na+ and Cl– ions. Future experiments will be designed and carried out to clarify this important point.
3.4. Escherichia coli cells that self-organized into biosaline drying patterns display vital reversibility
The next key hypothesis to be tested was whether the E. coli cells that enter into this self-assembled biosaline pattern in a “quiescent” state remain viable, that is, if they demonstrate the capacity to return to an active animated state (reanimation) after water is supplied to these salt-entrapped biofilms. In other words, do the E. coli bacterial cells enter into an anhydrobiotic state or do they not when they form these drying patterns? To test this hypothesis, a 7-day-old biosaline-dried bacterial biosaline biofilm (shown in Fig. 1E) was rinsed with a drop of fresh saline solution (Fig. 6A). Immediately after the drop was added (Fig. 6B), the bacterial biosaline patterns began to disassemble in a sequential and orderly fashion, releasing in this way the bacterial cells to the aqueous phase (Fig. 6F–6J). However, a remainder of tightly associated cells forming residual morphological patterns was still observed (data not shown). Then, surprisingly, both the cells that were liberated from the biosaline pattern as well as those that were liberated by mechanical disruption from these residual patterns were able to recover rapidly their “collective” activity of movement (see Movie S4 in the Supplementary Material), generating the characteristic edge of the droplet (Fig. 6K–6M and see Movies S5 and S6 in the Supplementary Material). Furthermore, as would be expected with water evaporation, the bacteria inside the biofilm again underwent (Fig. 6N, 6O) another cycle of the drying pattern formation, generating architectural patterns similar to those observed in the original foundational biosaline-drying biofilm; thus the process started all over again near the biofilm edge. The data indicate the likelihood that bacteria resist dissection when forming these drying patterns.

Escherichia coli cells are re-animated upon rehydration of a dried biosaline film. (
To confirm this suggestion, the viability of E. coli MG1655 strain cells in biosaline droplets kept at room temperature for 7 days was measured as explained in the Materials and Methods. As shown in Fig. 6P, a loss of viability (a reduction in viable cell counts) was observed between Day 1 and Day 2, but after this the viability remained stable with no additional decrease. Together, these results indicate that the bulk of bacterial cells that form a biosaline drying pattern withstand desiccation at least for 7 days after the formation of the drying pattern, and they are able to return to life (vital functions restored) after this elapsed time upon rehydration. Additionally, this experiment suggests that the collective bacterial movement observed under the microscope, when the water is supplied to the drying droplet, is likely a phenotypical manifestation of this cellular vitality.
4. Discussion
The findings presented in this work have several important astrobiological implications, which we address below.
4.1. Relevance for understanding bacterial anhydrobiosis
Escherichia coli bacteria employ similar mechanisms as those other bacteria employ to resist osmotic shock caused by increased water salinity in aquatic environments. For example, high water salinity prompts E. coli to synthesize trehalose and sucrose, uptake environmental glycine betaine, and produce EPS (Billi et al., 2000). Nevertheless, the mechanisms that E. coli employs to achieve resistance to extreme desiccation are poorly understood. From an astrobiological point of view, a relevant result of our study is that E. coli bacterial cells inside the biosaline patterns displayed vital reanimation upon rehydration (Fig. 6A–6M and 6P). Apparently, the patterns are completely desiccated structures with a water content below the level considered as the truly anhydrobiotic desiccated state; that is, the bacterial cells inside these structures can, as in toto entities, enter into a quiescent anhydrobiotic state while participating in the formation of these complex biosaline patterns.
In marked contrast, for instance, to air-dried bacterial cells, the cells in these patterns have a NaCl covering, can intermingle with this NaCl, and establish a strong interaction with this salt. Thus, we consider that this represents a new mode of drying, most likely with this saline coating shell providing protection against other environmental stressors such as cold, acids, antibiotics, UV radiation, and so on. Interestingly, it was previously reported that vegetative chemolithoautotrophic bacterial cells covered with very thin layers of a Mars analog regolith showed a higher survivability to UV radiation than bacterial cells that lack the protection provided by these covering layers (Gómez et al., 2010). Future experiments must clarify this suggestion.
4.2. Biomineralogical implications: how do the biosaline bacterial drying patterns form?
Biomineralization is the process by which living organisms produce minerals. Biologically induced mineralization (BIM) and biologically controlled mineralization (BCM) are two bacterially mediated modes that form minerals depending on the level of control the organism has over this mineral-formation process (Lowenstam, 1981; Lowenstam and Weiner, 1989; Mann, 2001). Because minerals produced under biological control have distinctive crystallographies, morphologies, and isotopic ratios that distinguish them from abiotically produced minerals with the same chemical composition (and are stable through geological time), the use of minerals that are the result of BCM processes as biomarkers (indications of past life) has been proposed for extraterrestrial life and could play an important role in the search for past or present life on Mars (Banfield et al., 2001; Blanco et al., 2006).
Several mechanisms have been proposed for modulating biomineralization by bacteria (BIM). These include metabolic processes that alter pH and supersaturation, as well as cell surface properties of EPS, cell membrane, and S-layers that may alter mineral nucleation. In BCM, organic compounds—proteins, lipids, proteoglycans, and polysaccharides—are known to control the structure, composition, shape, and organization of inorganic crystals (Mann, 2001).
Despite the study of drying patterns in evaporating droplets in abiotic systems, many aspects of how this pattern is actually formed are not yet fully understood. Are the drying biosaline bacterial patterns products of a genuine BIM formative process? A previous study conducted by Shiba et al. (2003) reported the formation of dendritic architectures in evaporating NaCl droplets containing the protein 288 (a synthetic periodic polypeptide). Although these authors were able to identify that the dendritic structures were constructed with NaCl, they were not able to clarify whether these structures contained the 288 proteins as well. Our studies with SEM, EDS analysis and video movies of the E. coli biosaline drying pattern formation show that these patterns have a layered organization, with the bacterial cells entering as whole entities (maintaining their cellular integrity) to form the drying pattern. Thus, not only bio-macromolecules, but also bacterial cells, as whole entities, can form drying biosaline patterns.
Given that proteins such as 288 can modulate the macroscopic habit exhibited by NaCl, it is tempting to speculate that modulation observed in drying biosaline patterns of E. coli cells could be caused by a protein component (or components) present in the bacterial surface or even a protein secreted by the bacteria to the surrounding medium (during the phase of adaptation on the evaporating droplet) that acts as a template-scaffold and promotes and organizes the NaCl deposition. However, the possibility that other components distinct from the proteins produced by E. coli (e.g., EPS exopolysaccharides) could be involved in this process cannot be discounted. Therefore, we assume that the assembly of NaCl and bacterial cells into dendrites of various shapes is controlled by the use of bacterial assembled structures as templates that ultimately incorporate the entire cell into the self-assembly. In view of this, we suggest that the drying bacterial pattern formations could be considered as a kind of BIM process (Frankel and Bazylinski, 2003). Therefore, the formation of bacterial saline drying patterns represents an excellent mode for studying the mechanisms that underlie bacterially induced biomineralization and could have a relevant role in understanding the process of “biologically induced” or “organic matrix-mediated” BIM.
4.3. Biological implications and future research
Finally, what are the biological implications of these results? Could E. coli use these biosaline drying formations as a survival strategy in response to different stressful situations? It is tempting to speculate that indeed their formation could be used by bacteria as a “collective last resort” against extreme environmental situations for achieving biological survival. The phenomenon is “collective” because the bacterial cells inside these biosaline formations could be effectively sequestered and protected in drying situations to endure, in this manner, these extreme environmental conditions; thus this may represent an optimal strategy for survival. The cells can achieve a complete dormancy in this anhydrobiotic state, as they are protected with a saline coat. They would furthermore be refractory to any kind of environmental assaults that normally require wet conditions and of living metabolic activity that could result in damage to key cellular components (Gómez-Gómez, 2010). Therefore, the study of these kinds of biosaline dying patterns will help reveal the biological mechanisms employed by bacteria for survival in extreme dry environmental conditions.
Several kinds of pattern formations have been described in colonies of E. coli under different environmental and nutritional conditions (Budrene and Berg, 1991; Darton, 2010; Zorzano et al., 2005). However, despite intensive research in recent years, many aspects of E. coli pattern formation, as well as how the morphology of the patterns change when they are exposed to different stresses (e.g., osmotic, antibiotic, oxidative, etc.), are poorly understood. In this regard, due to the availability of their complete genomic sequence (Blattner et al, 1997), the comprehensive set of single-gene knockout mutants (Baba et al., 2006), as well as their rapid growth and easy genetic manipulation, E. coli constitutes the reference model for studying the molecular, physiological, and genetic basis that underlies the biosaline drying pattern formation in biological systems. Furthermore, the bacterial biosaline drying structures could be an appropriate vehicle for bacterial dispersal. Future experiments must clarify these fundamental issues. Interestingly, in this regard, it is has been suggested that the search for extraterrestrial life should target chloride-bearing salts, for example, halite found on Mars, particularly in layered crystalline salts with preserved growth textures and fine-scale layering (Osterloo et al., 2008). Our discovery that bacteria can participate in the formation of bio-NaCl saline layered patterns makes the study of these bioformations of unmistakable astrobiological interest.
5. Conclusions
Using sessile saline droplets containing E. coli cells in this work, we report the discovery that water evaporation drives a salt-induced coupled self-assembly of bacterial cells with NaCl salt into biosaline quiescent drying complex patterns. Thus, most interestingly, the bacterial cells as whole, and under certain environmental conditions, can undergo a controlled self-organization into spatial formations in which the bacterial cells inside these patterns are capable of vital reanimation. Moreover, we judge that these findings will have important implications for understanding the capacity of bacteria to resist and survive extreme conditions. Such stressful environments can be encountered, for example, in the harsh environmental conditions in outer space or in numerous astrobiological sites where the search for life will take place in the coming decades, as well as in the many dry ecological niches on Earth.
Footnotes
Acknowledgments
The authors express their gratitude to Illias Tagkopoulos, Paul S. Cohen, Patrick C. Cirino, Sine L. Svenningsen, and to Gisela Storz for kindly providing them with the E. coli K-12 MG1655, MG1655 strR, W3110, NMHK8B, NM525 and GSO553 derivate strains, respectively, used in this work. We also thank an anonymous reviewer for constructive comments that have improved the quality of the manuscript.
Disclosure Statement for All the Authors
The authors declare that they have no competing interests.
Abbreviations
ABE, Agar Bacteriológico Europeo; BCM, biologically controlled mineralization; BIM, biologically induced mineralization; EDS, energy dispersive X-ray spectrometry; EPS, extracellular polymeric substances; LB, Luria-Bertani; SEM, scanning electron microscopy.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.