
Abstract
Bacteria and archaea isolated from crystals of halite 104 to 108 years old suggest long-term survival of halophilic microorganisms, but the results are controversial. Independent verification of the authenticity of reputed living prokaryotes in ancient salt is required because of the high potential for environmental and laboratory contamination. Low success rates of prokaryote cultivation from ancient halite, however, hamper direct replication experiments. In such cases, culture-independent approaches that use the polymerase chain reaction (PCR) and sequencing of 16S ribosomal DNA are a robust alternative. Here, we use amplification, cloning, and sequencing of 16S ribosomal DNA to investigate the authenticity of halophilic archaea cultured from subsurface halite, Death Valley, California, 22,000 to 34,000 years old. We recovered 16S ribosomal DNA sequences that are identical, or nearly so (>99%), to two strains, Natronomonas DV462A and Halorubrum DV427, which were previously isolated from the same halite interval. These results provide the best independent support to date for the long-term survival of halophilic archaea in ancient halite. PCR-based approaches are sensitive to small amounts of DNA and could allow investigation of even older halites, 106 to 108 years old, from which microbial cultures have been reported. Such studies of microbial life in ancient salt are particularly important as we search for microbial signatures in similar deposits on Mars and elsewhere in the Solar System. Key Words: Ancient DNA—Halite—Haloarchaea—Long-term survival. Astrobiology 14, 553–560.
1. Introduction
R
Recently, five enrichment cultures from three genera of halophilic archaea (Halorubrum, Natronomonas, and Haloterrigena) were obtained from buried salts, 22–34 ka in age, from Death Valley, California (Schubert et al., 2010a). Microscopy revealed that the particular salt crystals that yielded living archaea contained dozens of prokaryote-sized particles per crystal, which were preferentially located within fluid inclusions (Schubert et al., 2009a; Lowenstein et al., 2011). Those prokaryote-sized particles are similar in appearance to starved forms, such as miniaturized coccoids, reported from the oceans and soils (Morita, 1997; Grant et al., 1998). This suggests that once inside fluid inclusions, halophilic archaea exploit starvation survival strategies, but the timing and triggering mechanisms are not yet characterized (Schubert et al., 2010a).
Green algae microscopically similar to Dunaliella, the dominant primary producer in hypersaline lakes, have also been found inside fluid inclusions in ancient halite from Death Valley (Lowenstein et al., 2011). Algal cells within fluid inclusions likely contain glycerol, the carbon source that may allow co-entombed halophilic archaea to maintain low-level metabolic activity, as they do in modern hypersaline ecosystems. It was proposed that those fluid inclusions in buried salt that contain halophilic prokaryotes and algae are an ideal environment for long-term survival of halophilic archaea (Schubert et al., 2009b, 2010b).
Replication of results is an important criterion for verifying the authenticity of cultured ancient microbes (Willerslev et al., 2004). Low success rates of culture-based approaches, for example, less than 1% in ancient Death Valley halite (Schubert et al., 2010a), make replication of results difficult. In such cases, a culture-independent approach that uses polymerase chain reaction (PCR) to amplify fragments of DNA, followed by cloning and sequencing, can serve as an alternate means of verification.
Here, we present molecular evidence for preservation of DNA in ancient subsurface halite crystals, 22 and 34 ka in age, from Death Valley. Crystals were specifically chosen from three depths of a subsurface salt core, 13.0 m (∼22 ka), 17.8 m (∼34 ka), and 17.9 m (∼34 ka), from which halophilic archaea were previously cultured (Schubert et al., 2010a) (Fig. 1). Fragments of the archaeal 16S rRNA gene, ∼550 bp in size, comprising hypervariable regions V3–5, were amplified by PCR. The resulting amplicons were cloned, and a subset of clones was sequenced. Similarity searches and phylogenetic analyses were used to identify sequences similar to two of the three previously cultured halophilic archaeal genera from Death Valley (Schubert et al., 2010a).

(
2. Materials and Methods
2.1. Death Valley core DV-93
Core DV-93 from Death Valley is composed of interbedded halite and mud and contains a 90 m long, 100,000-year record of Pleistocene closed-basin paleoenvironments that range from hot, dry mudflats and saline pans to perennial lakes (Fig. 1A) (Lowenstein et al., 1999). Core DV-93 was radiometrically dated by using the uranium-series isochron method (Ku et al., 1998). Large numbers of prokaryotic microorganisms, up to 107 cells/mL, were found exclusively in fluid inclusions from one halite interval (13.0–18.0 m; 22–34 ka) when perennial saline lakes existed in Death Valley, which suggests that the trapping and preservation of microbes was paleoenvironmentally controlled (Lowenstein et al., 1999; Schubert et al., 2010a). Halite from that interval is composed of vertically oriented crystals that in cross section widen upward in a V pattern (Fig. 1B). Such halite structures indicate crystal growth as a crust at the bottom of a brine lake. Their preservation in core DV-93 indicates that the halite has not recrystallized since formation. Brine inclusions in bottom-growth halite crystals are rectangular prisms, cubic, or irregular in shape (Fig. 1C). Their orientation in planar arrays parallel to halite crystal growth faces is accepted as evidence for “primary” trapping of brine inclusions during a single salt crystallization event (Goldstein and Reynolds, 1994). Primary fluid inclusions can remain completely sealed and isolated from the external environment for periods of 500 million years (Lowenstein et al., 2001). Taken together, well-preserved halite growth structures and primary fluid inclusions indicate that the salt studied here has not recrystallized and that microbes have been sealed inside fluid inclusions since crystallization from surface brines. Prokaryotes in fluid inclusions are coccoid-shaped and “miniaturized,” with cell diameters <1 μm (Fig. 1D), which is much smaller than the straight or curved rods and coccoid-shaped halophilic prokaryotes found in modern hypersaline lakes (Schubert et al., 2009a). Laser Raman spectroscopy targeting red and orange masses associated with algal cells confirmed that organic materials, such as carotenoids, are also preserved in fluid inclusions (Fig. 1E) (Winters et al., 2013).
Ancient halite samples were obtained from depths of 13.0 m (∼22 ka), 17.8 m (∼34 ka), and 17.9 m (∼34 ka) of the Death Valley core DV-93. Halite crystals with primary fluid inclusions were screened for microbial cells or cellular structures with a Zeiss AXIO Imager A1 (Plan Apochromatic 100×/1.4 NA oil objective, Zeiss AxioCam MRm for grayscale images and Nikon Coolpix 8800 for color images). Similar fluid inclusions in halite from the same core depths were previously used to document long-term survival of halophilic archaea via cultivation experiments (Schubert et al., 2010a) and were used in the present study to document DNA preservation.
2.2. Surface sterilization and DNA extraction
Halite was surface sterilized by using a rigorous protocol that was developed to destroy DNA contaminants on crystal surfaces (Gramain et al., 2011; Sankaranarayanan et al., 2011). Halite crystals were “spiked” with a known quantity of purified human DNA. The “spiked” halite samples were soaked in 4 mL of 10 N HCl for 20 min. The halite crystals were then twice soaked in 4 mL of halite-saturated Na2CO3 solution for 15 min to ensure complete neutralization of acid. The halite crystals were then soaked in 4 mL of halite-saturated bleach for 20 min. A final set of four washes in 4 mL of halite-saturated brine (20 min) was performed to completely remove the bleach.
Surface-sterilized crystals were dissolved in 2.5 mL of DNA-free water (Roche, PCR grade water) and concentrated to a volume of 100 μL in Amicon centrifugal filters (Ultra-0.5, 50 kDa, YM-50, Millipore) (Sankaranarayanan et al., 2011). An additional 100 μL of PCR-grade water was added to bring the total volume of extract to 200 μL. Qiagen DNeasy plant mini kits were used to purify DNA from this extract.
Surface sterilization and DNA extraction were performed under a Class IIA laminar flow hood in a dedicated ancient DNA facility. PCR with human mitochondrial HV1 primers was used to verify successful removal of the intentionally introduced surface contaminant. Lack of amplification products on an agarose gel indicated successful surface sterilization.
2.3. Polymerase chain reaction
Polymerase chain reactions were carried out in 12.5 μL volumes containing 1× PCR buffer (Invitrogen), 200 μM dNTPs each, 1.5 mM MgCl2, and 0.5 μM primers. Three microliters of DNA template were added to each reaction. Forward primer A344F (Casamayor et al., 2002) was used with reverse primers 518R (Muyzer et al., 1993) and A915R (Casamayor et al., 2002) to amplify ∼200 and ∼550 bp fragments of the archaeal 16S rRNA gene, respectively. Primer pairs L15996 and H16401 were used to amplify ∼400 bp of the human mitochondrial HV1 region (Vigilant et al., 1991). PCRs with primer pairs A344F/518R and L15996/H16401 were run with the following conditions: initial denaturation at 94°C, followed by 50 cycles of 30 s at 94°C (denaturation), 30 s at 55°C (annealing), and 30 s at 72°C (extension), and a final extension of 10 min at 72°C. PCR conditions for primer pair A344F/A915R were identical with the exception of 45 s at 72°C during cycling and a final extension of 30 min at 72°C. This final extension is recommended to ensure addition of the 3′A to PCR products, which improves ligation efficiency. PCR products were visualized on a 1.2% agarose gel, stained with ethidium bromide. PCRs were set up in a separate room in the ancient DNA facility.
2.4. Cloning and sequencing
Purified amplicons from the ∼550 bp PCR were cloned (original TA cloning kit, Invitrogen), and inserts from positive clones (blue-white screening) were amplified with M13 primers and sequenced with ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Life Technologies).
2.5. Data analysis
Base calling was performed with ABI Sequencing Analysis Software v5.1 (Applied Biosystems, Life Technologies). Sequences were aligned in CLC sequence viewer and trimmed by removing vector and primer regions. Chimeric sequences were removed from the data set with the UCHIME program (Edgar et al., 2011). Sequences were assigned genus-level taxonomies by using the Ribosomal Database Project classifier (Wang et al., 2007; Cole et al., 2009). Sequences were also compared to 16S rDNA from cultured Death Valley archaea (FJ492047–FJ492052) by using nucleotide BLAST (Altschul et al., 1990). Sequences >99% similar to cultured Death Valley archaea were retrieved and aligned with MEGA (Tamura et al., 2011), imported into DnaSP (Librado and Rozas, 2009), and converted into .rdf files for use with Network (Bandelt et al., 1999). Median-joining networks were then constructed in Network by using the default parameters. Sequences used in our analysis are deposited in NCBI (GenBank ID: KF058934–KF059022, KJ494931–KJ495688).
3. Results
DNA extracts from ancient halite did not show any amplification (absence of bands on an agarose gel) with human mitochondrial HV1 primers, indicating successful surface sterilization. Extracts from surface-sterilized ancient halite showed ∼44–50% PCR success rates (n=16) with primers amplifying a ∼550 bp fragment of the 16S rRNA gene. Cloning and sequencing of these amplicons, followed by chimera filtering, resulted in the recovery of 846 16S rRNA sequences from the three depths (DV427, 13.0 m, n=296; DV582, 17.8 m, n=291; and DV588, 17.9 m, n=259) (Fig. 2). Taxonomic assignments made with the Ribosomal Database Project classifier showed 16S rRNA sequences from haloarchaeal genera, including Natronomonas, Halorubrum, Haloarcula, and Halobiforma, and several as-yet unclassified haloarchaea (between ∼30% and 60% of clone libraries) (Fig. 2).
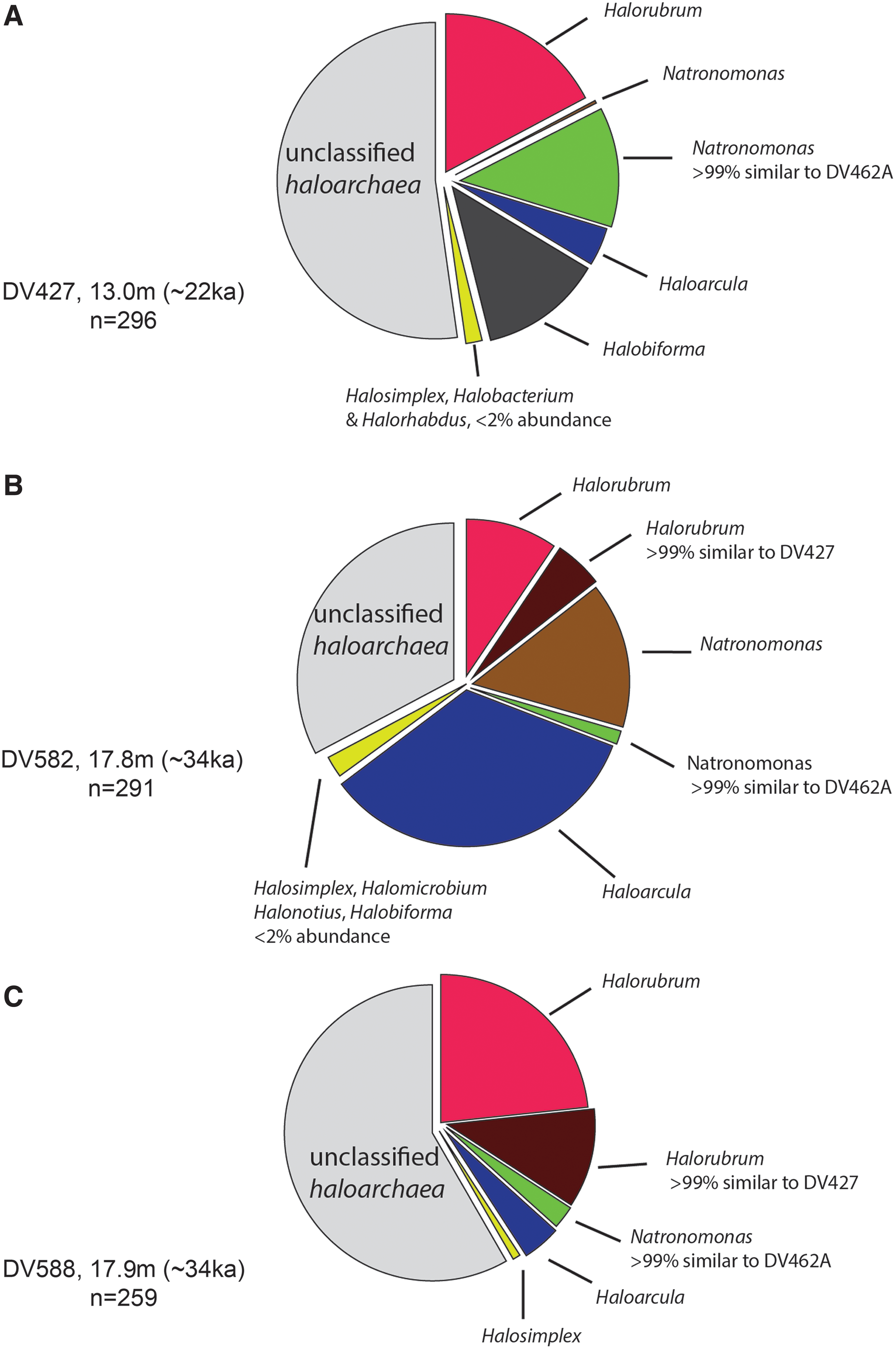
Taxa summary plots (genus level) for clone libraries from ancient Death Valley halite. (
Sequences from cloned PCR amplicons were then compared to 16S rDNA from previously cultured Death Valley archaea (FJ492047–FJ492052, including Natronomonas DV462A and Halorubrum DV427) by using BLAST (Altschul et al., 1990). Sequences identical to Natronomonas DV462A were identified from halite at all three depths tested (Fig. 3A). Additionally, sequences >99% similar to Natronomonas DV462A were recovered from the same depths. Within clone libraries from the three sampled depths, sequences >99% similar to Natronomonas DV462A had relative abundances ranging from ∼10% (13.0 m) to ∼1–2% (17.8 and 17.9 m) (Fig. 2). Sequences >99% similar to Halorubrum DV427 were also identified from halite at depths of 17.8 and 17.9 m (Fig. 3B), with relative abundances of ∼4% (17.8 m) and ∼9% (17.9 m).
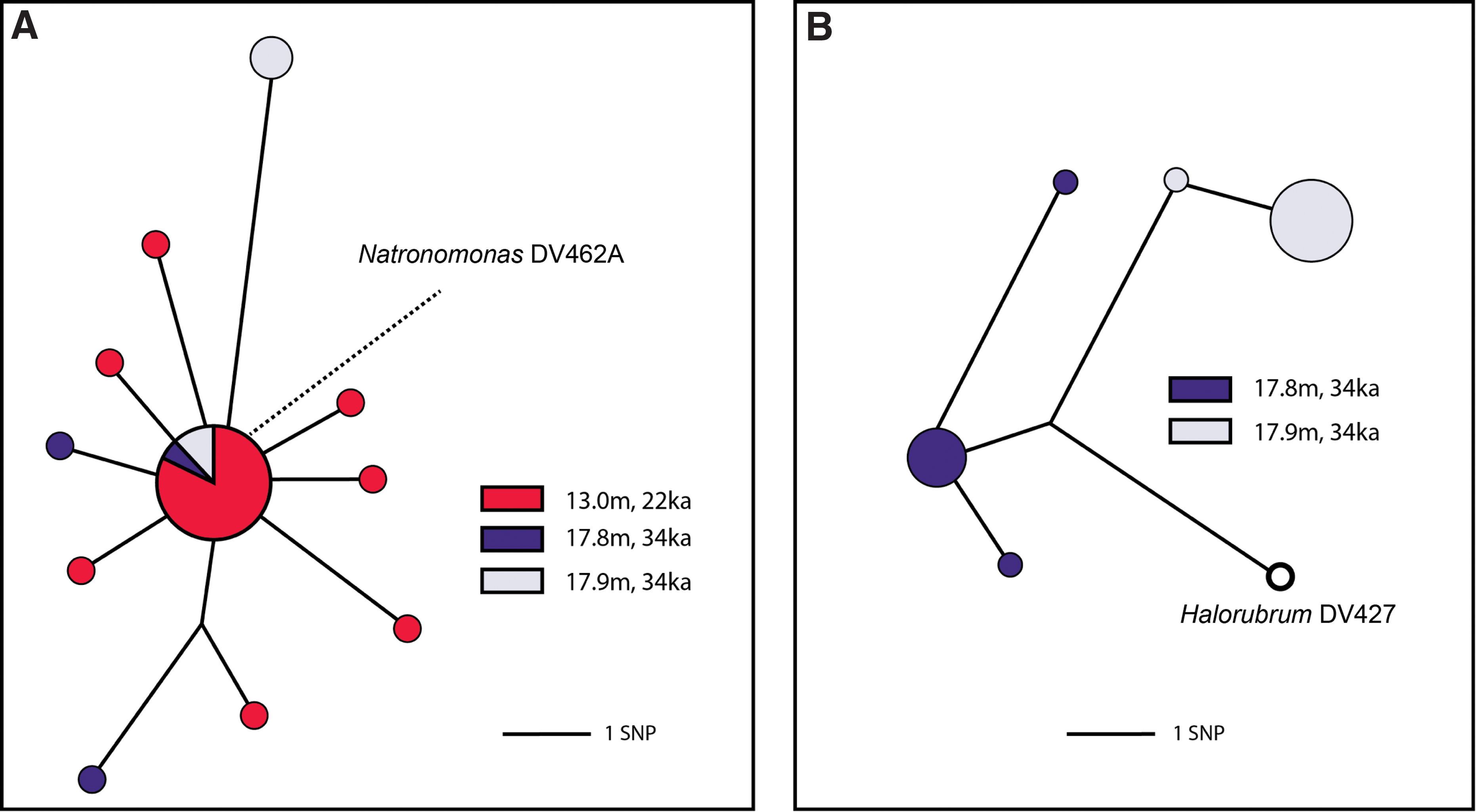
Median-joining network (Bandelt et al., 1999) of sequences >99% similar to (
The recovery of ancient DNA sequences identical, or nearly so, to Natronomonas DV462A and Halorubrum DV427 provides independent support for the authenticity of these cultured organisms. Further, the recovery of 16S rDNA sequences with <1% divergence from the cultured microbes suggests the existence of a larger community of closely related organisms in the ancient Death Valley halite.
4. Discussion
The use of stringent surface sterilization and extraction protocols (see Materials and Methods for details), which were performed in a dedicated ancient DNA facility, in a building separate from the laboratory where the previous culturing study was conducted, addresses the major criteria (surface decontamination, inter-laboratory reproducibility, sterile work environment) (Cooper and Poinar, 2000; Willerslev et al., 2004; Hebsgaard et al., 2005; Willerslev and Cooper, 2005) considered essential for validation of ancient DNA/culturing experiments.
Studies comparing microbial diversity obtained by using culturing and PCR-based approaches from several sources, including soil, freshwater, and marine habitats (Suzuki et al., 1997; Felske et al., 1999; Floyd et al., 2005), report that organisms well represented in cultures are often underrepresented in clone libraries, which is reflective of differences in ease of cultivability. In the ancient Death Valley halite, culturing approaches are further limited to targeting a subset of organisms that survived entombment for tens of thousands of years, in addition to ease of cultivability. In contrast, PCR-based approaches obtain estimates of microbial diversity driven by relative abundance of 16S rDNA template molecules. Thus, the recovery of 16S rDNA sequences >99% similar to the two strains Natronomonas DV462A and Halorubrum DV427 from halite samples similar to those from which they were originally cultured provides conclusive support that demonstrates their presence in ancient Death Valley halite.
Sequences >99% similar to the two cultured strains comprise <10% of the clone libraries from ancient Death Valley halite, which suggests that these organisms were a subset of a larger microbial community trapped in the halite. DNA sequences similar to strains of the third cultured genus, Haloterrigena, were not observed in the clone libraries reported here. Further sampling would be necessary to conclusively verify the authenticity of the three Haloterrigena strains recovered from the previous culturing study (Schubert et al., 2010a). The culture-independent approach used here suggests that the relative abundance of Haloterrigena 16S rDNA is possibly lower than the detection limits of our current clone libraries, which suggests the relative ease of cultivability of these strains. On the other hand, improvements in culturing strategies could lead to the recovery of additional haloarchaeal strains, such as Haloarcula, that are relatively abundant in the 16S rDNA clone libraries from the ancient Death Valley halite (Fig. 2, ∼4–30%).
5. Conclusion
The findings reported here collectively support long-term survival of halophilic archaea and preservation of DNA within fluid inclusions for periods of tens of thousands of years. Additionally, these results demonstrate the applicability of PCR-based techniques for verification of the authenticity of ancient microbial cultures. In contrast to the low success rates of culturing techniques (Schubert et al., 2010a), the sensitivity of PCR allows for consistent retrieval of DNA from geologically ancient samples with small amounts of preserved DNA. The relative ease of reproducibility for PCR-based approaches makes them ideally suited to the investigation of older halite, millions of years in age, from which haloarchaeal cultures have been reported (Stan-Lotter et al., 1999, 2002; Vreeland et al., 2000, 2007). Further study of microbial life in ancient salt is particularly important as we search for microbial signatures in similar deposits on Mars (Fendrihan and Stan-Lotter, 2004; Squyres et al., 2004).
Footnotes
Acknowledgments
This work was funded by an NSF Geobiology & Low Temperature Geochemistry grant (EAR 1024692). Sequences described in this work are deposited in the NCBI GenBank database (GenBank ID: KF058934–KF059022, KJ494931–KJ495688).
Author Disclosure Statement
No competing financial interests exist.
Abbreviation
PCR, polymerase chain reaction.