
Abstract
Nearly 40 years ago, Schoffstall and his coworkers used formamide as a solvent to permit the phosphorylation of nucleosides by inorganic phosphate to give nucleoside phosphates, which (due to their thermodynamic instability with respect to hydrolysis) cannot be easily created in water by an analogous phosphorylation (the “water problem” in prebiotic chemistry). More recently, we showed that borate could stabilize certain carbohydrates against degradation (the “asphalt problem”). Here, we combine the two concepts to show that borate can work in formamide to guide the reactivity of nucleosides under conditions where they are phosphorylated. Specifically, reaction of adenosine in formamide with inorganic phosphate and pyrophosphate in the presence of borate gives adenosine-5′-phosphate as the only detectable phosphorylated product, with formylation (as opposed to hydrolysis) being the competing reaction. Key Words: Prebiotic chemistry—Nucleoside phosphates—Phosphorylation—Borate—Formamide. Astrobiology 15, 259–267.
1. Introduction
T
Over the past several decades, several new approaches have been proposed to separately manage these two problems. First, organic materials can interact with many different mineral species (Lahav and Chang, 1976; Ferris et al., 1989; Holm et al., 1993). This interaction can, of course, catalyze reactions to transform those species. Perhaps best known is the use of clay minerals to catalyze the synthesis of RNA oligomers from nucleosides after they are modified with condensing reagents, notwithstanding the uncertainty in the availability of such reagents in aqueous prebiotic environments (Ferris et al., 1996).
More recently, minerals have been proposed to constrain undesired organic reactions, especially those that lead to “asphalts,” complex (and unproductive) mixtures of organic molecules (Benner et al., 2010, 2012). For example, the stabilization of ribose and other 5-carbon sugars by borate minerals reduces the formation of asphalts, making these species plausibly better precursors for RNA prebiotic synthesis (Ricardo et al., 2004; Kim et al., 2011; Furukawa et al., 2013). Other work related to borate and prebiotic chemistry has been done (Prieur, 2001a, 2001b; Springsteen and Joyce, 2004; Kofoed et al., 2005; Scorei and Cimpoiasu, 2006; Amaral et al., 2008; Rauchfuss, 2008; Cossetti et al., 2010; Grew et al., 2011; Scorei, 2012; Stephenson et al., 2013).
Separately, the “water problem” has been approached through the suggestion that key steps in the assembly of the building blocks of RNA occurred in non-aqueous solvents. Many solvents have been proposed, including supercritical carbon dioxide (Schreiber et al., 2012). However, already 40 years ago, Schoffstall and his coworkers suggested that formamide might have served as a prebiotic solvent for the assembly of phosphate ester linkages, one class of bonds in oligomeric RNA that is quite unstable in water thermodynamically (Schoffstall, 1976; Schoffstall et al., 1982; Schoffstall and Laing, 1985). Formamide is, of course, the product of the reaction of hydrogen cyanide with water, both indisputably present on early Earth. Formamide remains a liquid from 0°C to 210°C. More recently, Saladino and his associates noted that formamide is in many ways a better precursor for nucleobases than hydrogen cyanide (Saladino et al., 2003, 2012), a classical prebiotic precursor for adenine and parts of other nucleobases (Oró and Kimball, 1961, 1962; Sanchez et al., 1967).
These observations motivated us to ask whether mineral species might be combined with formamide to control, in one pot, organic reactivity at steps where hydrolytically unstable bonds are synthesized. In particular, we were interested in learning whether borate species might control the regiochemistry of phosphorylation of nucleosides in formamide, the “Schoffstall phosphorylation.” We report here results that show they can.
2. Materials and Methods
2.1. Materials
Reagent-grade adenosine (>99%), formamide (>99.5%), sodium metasilicate (Na2SiO3), sodium pyrophosphate dibasic (Na2H2P2O7), trisodium trimetaphosphate (Na3P3O9; >95%), and zinc phosphate (Zn3(PO4)2; >99.998%) were obtained from Sigma-Aldrich and used as received. Sodium monophosphate (NaH2PO4·H2O; ACS-grade) and sodium tetraborate decahydrate (Na2B4O7·10H2O; ACS-grade) were from Fisher Scientific. Deuterated formamide (formamide-d3, 99.2 atm % D) was from CDN Isotopes. Standards used for HPLC analysis were 2′-AMP, 5′-AMP, 2′,3′-cAMP, 3′,5′-cAMP, 2′,5′-ADP, 3′,5′-ADP, 5′-ADP, 5′-ATP. The 3′-AMP was from Santa Cruz Biotechnology. Other standards were all from Sigma-Aldrich. Tetrabutylammonium bromide (TBA-Br) and potassium phosphate (KH2PO4; ACS-grade) were used to make an ion-pair buffer eluent for HPLC analysis. They were from Tokyo Chemical Industry and Fisher Scientific, respectively. Acetonitrile (anhydrous, 99.8%) used for HPLC was from Sigma-Aldrich.
For the measurement of water contents in formamide, water (HPLC), lithium chloride (>99%), potassium carbonate (>99%), sodium bromide (>99%), sodium chloride (>99%), and potassium sulfate (>99%) were obtained from Sigma-Aldrich and used as received.
2.2. Methods
Phosphorylation experiments were conducted in formamide (2 mL) containing adenosine (30 mM), NaH2PO4 (4 equivalents, 120 mM), and (as needed) Na2B4O7·10H2O (1 equivalent, 30 mM) or Na2SiO3 (1 equivalent). After incubation at 90°C to reproduce Schoffstall's conditions, aliquots of solution (10 μL) were sampled, mixed with water (990 μL), and analyzed with HPLC.
Proton NMR experiments were conducted with Varian Mercury 300 NMR at room temperature by using deuterated formamide (0.5 mL) as the solvent. The solutions contained adenosine (35 mM) and were added with zero, 0.2, or 0.5 equivalent moles of borate from sodium tetraborate decahydrate, or 0.5 equivalent moles of silicate from sodium metasilicate.
High-performance liquid chromatography analysis was performed with a C-18 reversed-phase narrow bore column (3 mm i.d., 150 mm length, 5 μm; SunFire; Waters) on a Waters 2695 separation module equipped with 996 photodiode array detector, which was also used to collect spectra. Standards were used to determine the mobility of adenosine, nine kinds of adenosine nucleotides, and adenine at ambient temperature. The column was eluted with a gradient of (A) aqueous 20 mM KH2PO4 with 5 mM TBA-Br (pH 3.2, adjusted by phosphoric acid) and (B) 100% acetonitrile. The elution program created a linear gradient that started from 99% A to 2.5 min, 97% at 5.5 min, 89.5% at 17.5 min, and 65.0% at 23.5 min with the total flow rate at 0.8 mL/min. Peak detection and integration were conducted with the signal at 260 nm; full spectra were also obtained.
Several products (
We measured the water content in formamide equilibrated with air maintained in sealed desiccator chambers at several humidity levels by having the air humidity maintained by saturated salt solutions. Formamide and water were mixed to give two stock solutions, one containing 50% formamide by weight, the other 20% formamide by weight. Five chambers were prepared in triplicate (15 jars total), each containing the following open vials: (a) one vial contained 10 mL of a saturated salt solution whose choice determined the humidity (chosen from each: LiCl, K2CO3, NaBr, NaCl, or K2SO4) over several grams of excess solid salt; (b) one vial contained 750 μL of pure formamide; (c) one vial contained 1.5 mL of pure formamide; (d) one vial contained 750 μL of the 50% formamide-water mixture; (e) one vial contained 1.5 mL of the 50% formamide-water mixture; and (f) one vial contained 1.5 mL of the 20% formamide-water mixture. All formamide-containing vials were previously weighed (tared). The jars were then tightly closed and placed either in the refrigerator (4°C), at room temperature (air-conditioned, 23°C), or in an incubator at 40°C, to let the various formamide-water mixtures equilibrate under the relative humidity controlled by the salt. After 22 days, the jars were reopened and all the formamide-containing vials weighed again. The equilibrium percentage (by weight) of water in formamide at a given relative humidity and temperature was calculated from the weight increase or decrease of the vials. The relative humidities of the various salts were taken from the work of Greenspan (1977).
3. Results
3.1. Adenosine in formamide forms a complex with borate but not with silicate
Lambert et al. (2010) suggested that silicate might be an important mineral to stabilize ribose. To detect complexation between adenosine and borate or silicate, proton NMR spectra were recorded of the adenosine in perdeuterated formamide (which provided the deuterium lock) with increasing amounts of sodium tetraborate decahydrate (0.2 or 0.5 mole equivalents of borate) and with sodium metasilicate (0.5 mole equivalents of silicate). The samples were premixed in an Eppendorf tube and vortexed to dissolve. The sodium borate dissolved completely; the sodium silicate did not.
With the addition of borate, several signals arising from adenosine (D′, J′, and K′ in Fig. 1b and 1c) appeared next to each proton signal of adenosine (D, J, and K in Fig. 1a–1c). These were assigned to be a borate complex of adenosine. The intensities of these new peaks increased with the amounts of borate added, whereas the intensities of original proton signals from adenosine decreased. Because the signals J and K are derived from the 2′- and 3′-protons, this result suggests that adenosine forms 2′,3′-cyclic complex with borate (Fig. 2) in formamide.

H1NMR spectra in perdeuterated formamide (D-C(=O)ND2) of (
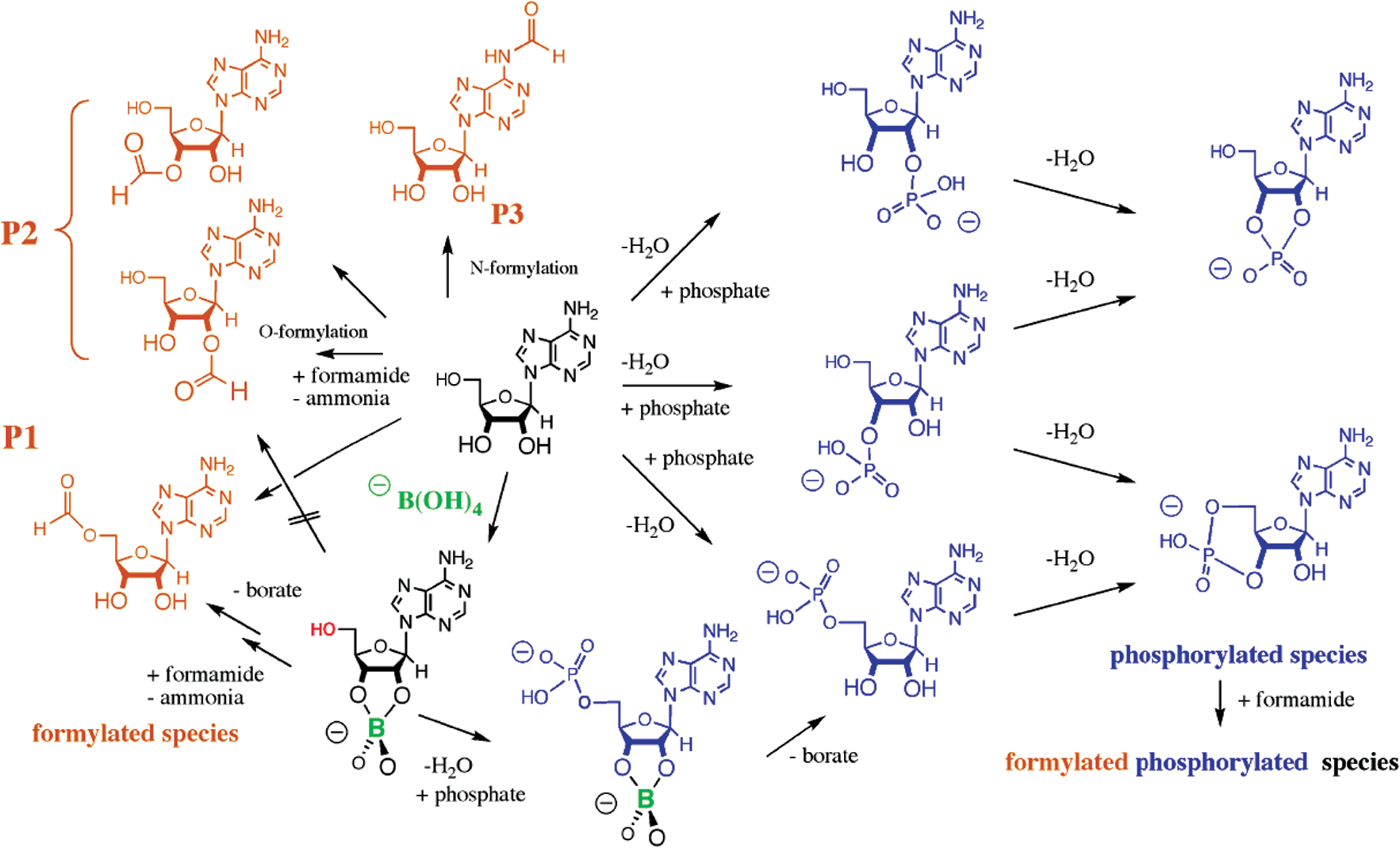
Schematic showing possible products derived from adenosine (center, black), including reactions with formamide reacting as an electrophile (left) and phosphate as an electrophile (right). Borate complexation is expected to moderate the regiochemistry of both classes of reactions by blocking nucleophilic attack from the 2′- and 3′-hydroxyl groups of adenosine. (Color graphics available at
On the other hand, the NMR signals of adenosine were not affected by the addition of silicate (Fig. 1d). Only a slight broadening was observed and attributed to the presence of particles of incompletely dissolved silicate. This indicates that adenosine does not form a complex with silicate in formamide.
3.2. Adenosine is phosphorylated by inorganic NaH2PO4 in formamide
In all experiments, NaH2PO4·H2O dissolved completely in formamide. Upon incubation at 90°C, multiple products were observed by HPLC to form, including 5′-AMP, 3′-AMP, 2′-AMP, 2′,3′-cAMP, 3′,5′-cAMP, 2′,5′-ADP, 3′,5′-ADP, and 5′-ADP (Figs. 2 and 3a). All of these were assigned in the HPLC trace by using authentic standards. In addition, products were seen by HPLC that did not correspond to any of the standards added, three of these in particular labeled
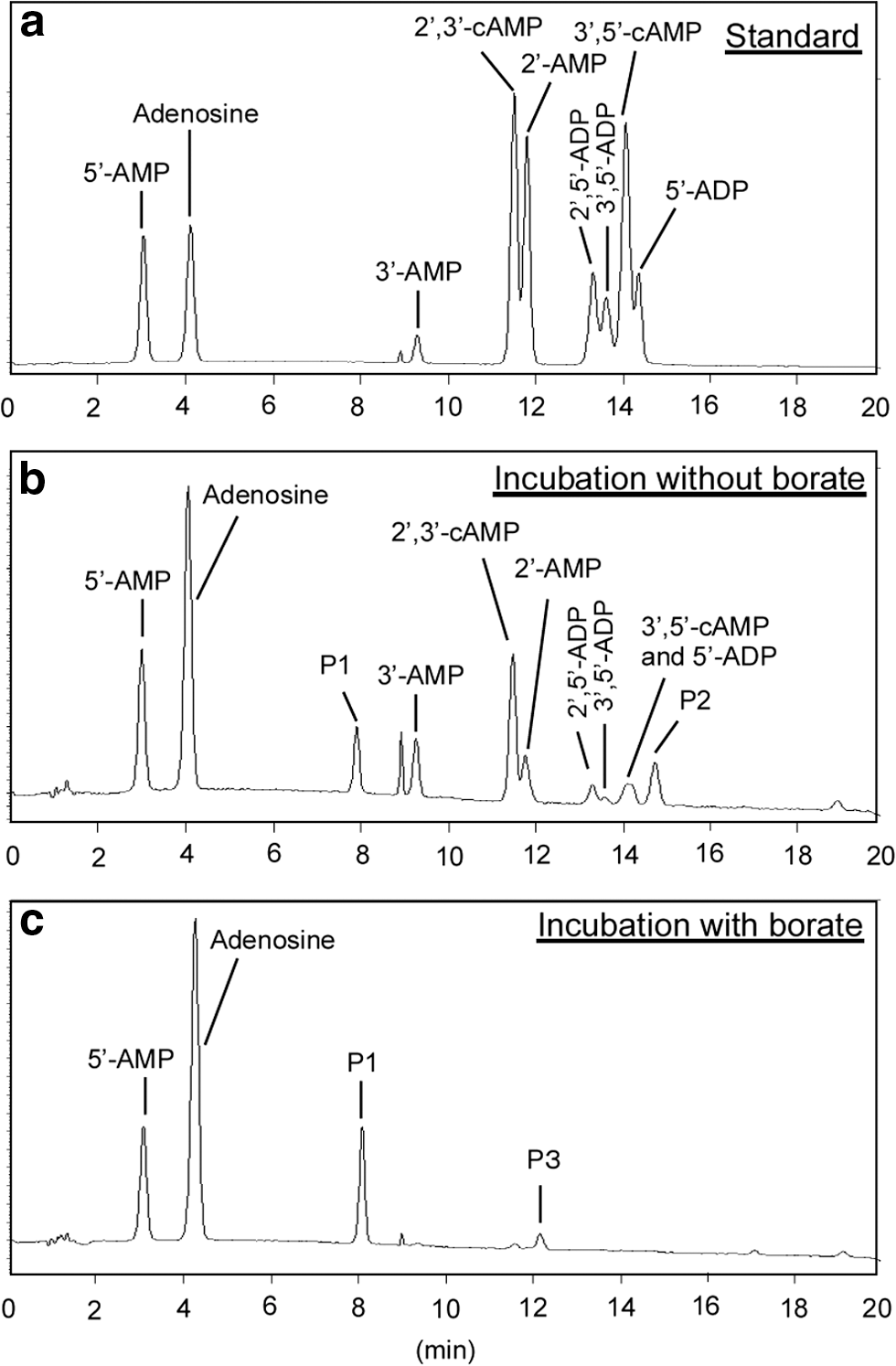
HPLC chromatograms (260 nm) resolving products arising from phosphorylation of adenosine and adenosine-borate complexes. (
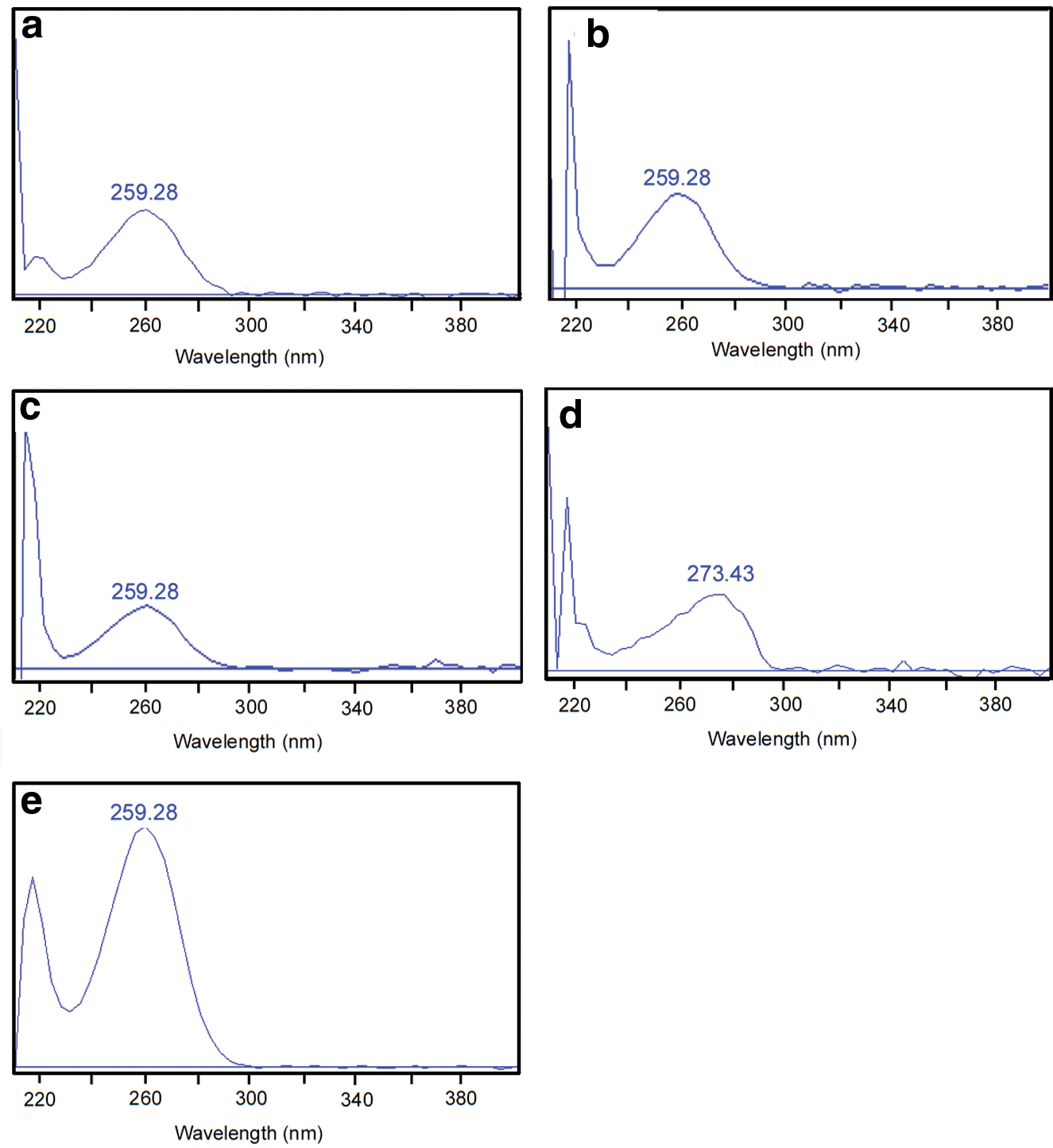
Photodiode array spectra of HPLC fractions: (
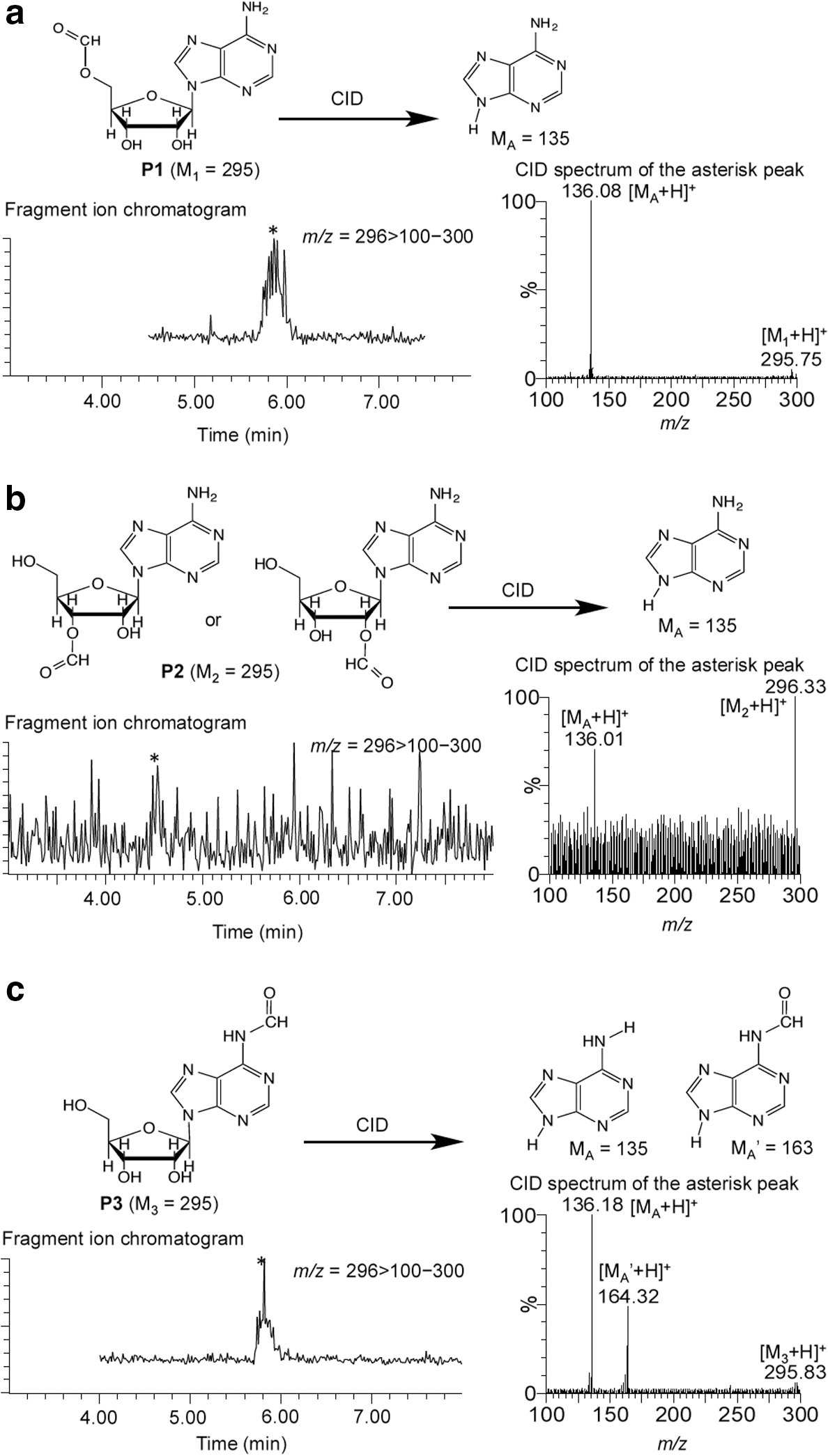
Collision-induced dissociation (CID) spectra of
The amounts of 5′-AMP, 3′-AMP, and 2′-AMP increased in the initial hours of incubation, reaching a maximum at ca. 24–48 h (Figs. 2 and 6a); upon further incubation, their amounts gradually decreased, evidently as they were further transformed. As one candidate product for further transformation, 2′,3′-cAMP continuously increased in amount throughout the incubation (which was stopped after 168 h). Thus, in the first stage (until ∼100 h), 5′-AMP was the principal product. After ∼144 h, 2′,3′-cAMP became the major product (Fig. 6a).

Time course of the phosphorylation of adenosine (30 mM, Schoffstall conditions) in formamide resolved using ion-pair HPLC (tetrabutylammonium bromide, potassium phosphate) with (
3.3. In the presence of borate, adenosine is regioselectively phosphorylated by various phosphorylating agents
To determine whether borate complexation could constrain the complexity of the product mixture, these experiments were repeated in the presence of borate. As shown in Figs. 3 and 5, the amounts of 3′-AMP, 2′-AMP, 2′,3′-cAMP, 2′,5′-ADP, and 3′,5′-ADP, and 5′-ADP were all considerably less in the presence of borate than in its absence. Instead, 5′-AMP and the unassigned
Sodium dihydrogen pyrophosphate (60 mM, if all dissolved), sodium trimetaphosphate (30 mM, if all dissolved), and zinc phosphate (60 mM, if all dissolved) were also examined as potential sources of phosphate in the presence of borate. Unfortunately, these phosphates did not dissolve completely in formamide. Nevertheless, with pyrophosphate, comparable amounts of 5′-AMP and
3.4. Regiochemistry of phosphorylation of adenosine is not influenced by silicate
Sodium metasilicate proved to be difficult to dissolve in formamide. Perhaps for this reason, the complexity of the phosphorylation product mixture was almost the same as seen in the experiments without borate (Figs. 3 and 6e). Whatever silicate was available conferred no evident regioselectivity in the Schoffstall phosphorylation reaction.
4. Discussion
In general, work with mineral-organic interactions seeks evidence that these interactions facilitate reactions. For example, Saladino and his coworkers noted the ability of many mineral species to create nucleobases from formamide and other precursors; this was important work that helped to develop our understanding of how these RNA components might have arisen on early Earth (Saladino et al., 2003, 2012).
This work moves into largely unexplored territory by seeking minerals that might limit the complexity of products arising under prebiotic conditions. We and others have previously shown that borate constrains the reactivity of carbohydrates, preferentially stabilizing 5-carbon products generated from formaldehyde (Kim et al., 2011) and, among 5-carbon pentoses, ribose (Furukawa et al., 2013). Here, it is interesting to examine the details of the Schoffstall phosphorylation reaction, which can also generate a reasonably complex manifold of products (Fig. 6).
In the absence of borate, essentially all the products expected from nonspecific phosphorylation (Figs. 2 and 6) were seen. Initially, 5′-phosphorylation was the major product, but with substantial amounts the 2′- and 3′-phosphorylation products were observed as well. The 2′,3′-cAMP was the major product at the end of the reaction without borate.
One interpretation of these results is consistent with the model that the initial phosphorylation occurs at the sterically least hindered 5′-alcohol. This phosphorylation requires, of course, the removal of a single molecule of water, a removal that is thermodynamically acceptable in formamide having little water. However, should the activity of water be extremely low in the environment, a second molecule of water might be removed to create a cyclic phosphate. Products evolved over time, as seen in Fig. 5, with 2′,3′-cyclic AMP becoming an increasingly dominant product as the incubation proceeded.
The ability of borate to control this diversity was striking. The fact that borate in formamide forms a complex that “ties up” the 2′- and 3′-alcohols of adenosine suggests that borate might direct Schoffstall phosphorylation away from the 2′- and 3′-OH groups and toward the 5′-OH group. This expectation was met. In the presence of borate, 5′-phosphorylation generated the overwhelmingly dominant product.
The cleanliness of the result led us to ask what the unassigned products,
Silicate did not form any detectable (by NMR) complex with adenosine in formamide. This result led us to expect no influence by silicate on the regiochemistry of the Schoffstall phosphorylation. Essentially none was observed. It is still unclear whether the absence of an effect is better explained by the low apparent solubility of silicate in formamide or the inability of silicate to form a complex with adenosine.
From simple stoichiometry, it can be seen that pyrophosphate can generate phosphate esters directly from nucleosides without the need to remove a water molecule. Therefore, the only theoretical advantage to using formamide with pyrophosphate as a phosphorylation reagent is to ensure that the product is not subsequently hydrolyzed. Phosphorylation was observed in formamide with sodium pyrophosphate as the reagent in the presence of borate. Interestingly, the initial rate of phosphorylation with sodium pyrophosphate was quite similar to the slope of the line with phosphate. This might be interpreted as evidence for a “common intermediate” followed by a rate-limiting step. Here, it is hypothesized that the ultimate phosphorylating reagent is a formamide molecule carrying a phosphoryl group on its oxygen. Further work is needed to explore this hypothesis.
Even without borate, these results have prebiotic relevance. Abiotic phosphorylation of nucleosides is a key chemical step that may have provided nucleotides to the prebiotic world. This phosphorylation yields several types of nucleoside monophosphate (NMP): 2′-monophosphate (2′-NMP), 3′-monophosphate (3′-NMP), 5′-monophosphate (5′-NMP), 2′,3′-cyclic monophosphate (2′,3′-cNMP), and 3′,5′-cyclic monophosphate (3′,5′-cNMP). The 2′,3′-cNMP that formed after prolonged incubation under dehydrating conditions is particularly interesting. Its strain provides it energy that might be used to form oligomers (Westheimer, 1968; Gerlt et al., 1975).
However, the 5′-NMP nucleosides that are selectively formed in the presence of borate are also of interest. Given condensing agents, these are the substrates for the Orgel-Ferris condensation reactions to give oligomers in the presence of clays (Schwartz and Orgel, 1985; Ferris et al., 1996).
How likely is it that formamide was present on early Earth or Mars such that it might have supported prebiotic chemistry? As a closing experiment, we collected data that show that mixtures of formamide and water, if left to equilibrate with an atmosphere with defined humidity, became enriched in formamide (Fig. 7). For example, when the relative humidity of an environment is maintained at 11% by a saturated solution of LiCl, the amount of water in formamide is approximately 5%, 10%, and 15% at 40°C, 23°C, and 4°C, respectively. Thus, formamide becomes enriched in lacustrine environments, for example, in Death Valley in the summer. However, in such lacustrine solvent that is rich in formamide, strong UV radiation as a result of the lack of an ozone layer on prebiotic Earth might have destroyed adenosine and its phosphates. Therefore, the borate-moderated phosphorylation reactions reported here are conceivable in sediment pore solvent in such lacustrine environments on a prebiotic Earth where UV radiation was shielded by minerals or salts (Cockell, 1998).
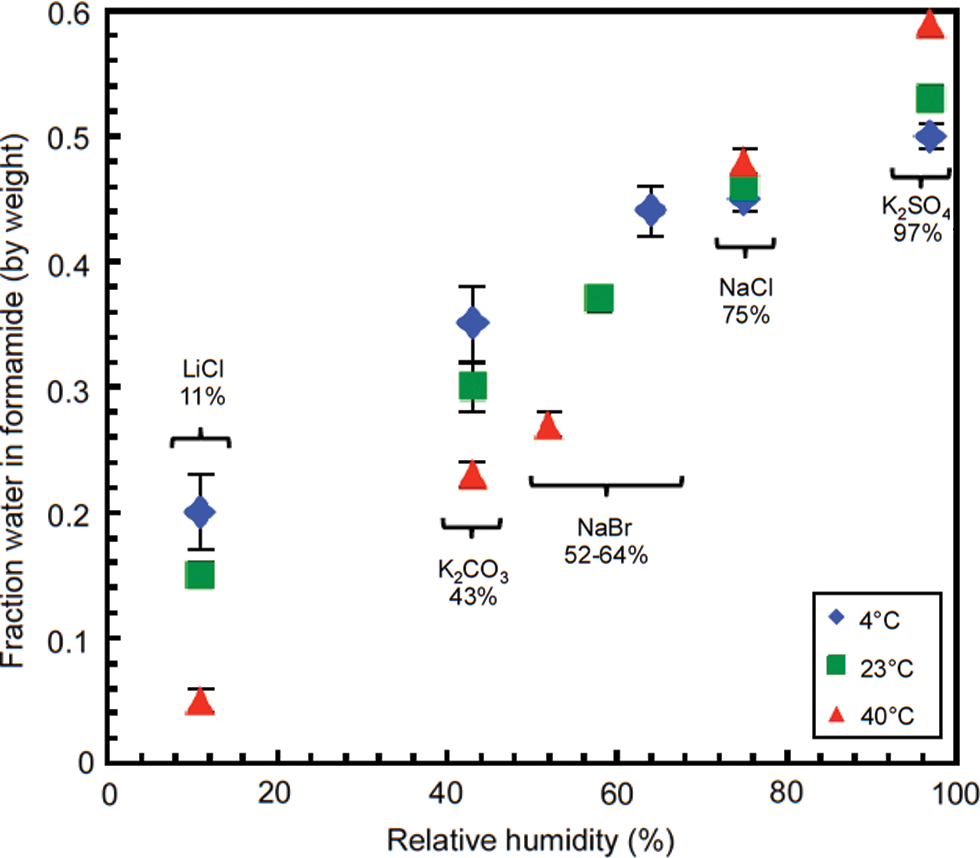
Weight fraction of water in formamide equilibrated in a closed chamber at three temperatures (4°C, 23°C, 40°C) with humidity maintained by a saturated solution of the indicated salt. (Color graphics available at
But were similar deserts present on early Earth? Briefly, we do not know. However, the inventory of water on early Earth and the absence of mature plate tectonics might have resulted in a planet that was flooded and offered very little dry land for formamide to accumulate as a solvent [pace some geological evidence suggests the occurrence of continental crusts before 4.3 Ga, possibly giving dry land (Wilde et al., 2001; Harrison, 2009)]. In this context, it is perhaps worth noting that Mars might have always had dry land (Phillips et al., 2001; Perron et al., 2007).
Footnotes
Acknowledgments
This work was supported in part by the NASA Exobiology program (NNX08AO23G) and the NASA Astrobiology Institute program on Titan (via a subcontract from the Jet Propulsion Laboratory, NNH08ZDA002C). Y.F. is indebted to Tohoku University for travel funds.
Author Disclosure Statement
No competing financial interests exist.