
Abstract
Basaltic glass (BG) is an amorphous ferrous iron [Fe(II)]–containing material present in basaltic rocks, which are abundant on rocky planets such as Earth and Mars. Previous research has suggested that Fe(II) in BG can serve as an energy source for chemolithotrophic microbial metabolism, which has important ramifications for potential past and present microbial life on Mars. However, to date there has been no direct demonstration of microbially catalyzed oxidation of Fe(II) in BG. In this study, three different culture systems were used to investigate the potential for microbial oxidation of Fe(II) in BG, including (1) the chemolithoautotrophic Fe(II)-oxidizing, nitrate-reducing “Straub culture”; (2) the mixotrophic Fe(II)-oxidizing, nitrate-reducing organism Desulfitobacterium frappieri strain G2; and (3) indigenous microorganisms from a streambed Fe seep in Wisconsin. The BG employed consisted of clay and silt-sized particles of freshly quenched lava from the TEB flow in Kilauea, Hawaii. Soluble Fe(II) or chemically reduced NAu-2 smectite (RS) were employed as positive controls to verify Fe(II) oxidation activity in the culture systems. All three systems demonstrated oxidation of soluble Fe(II) and/or structural Fe(II) in RS, whereas no oxidation of Fe(II) in BG material was observed. The inability of the Straub culture to oxidize Fe(II) in BG was particularly surprising, as this culture can oxidize other insoluble Fe(II)-bearing minerals such as biotite, magnetite, and siderite. Although the reason for the resistance of the BG toward enzymatic oxidation remains unknown, it seems possible that the absence of distinct crystal faces or edge sites in the amorphous glass renders the material resistant to such attack. These findings have implications with regard to the idea that Fe(II)-Si-rich phases in basalt rocks could provide a basis for chemolithotrophic microbial life on Mars, specifically in neutral-pH environments where acid-promoted mineral dissolution and utilization of dissolved Fe(II) as an energy source is not likely to take place. Key Words: Basaltic glass—Chemolithotrophic—Microbial—Iron oxidation—Mars. Astrobiology 15, 331–340.
1. Introduction
I
Our interest in the potential for microbial oxidation of Fe(II) in basaltic glass (BG) stems from recent suggestions that near-surface hydrothermal venting may have occurred during past periods of active volcanic/tectonic activity on Mars (Lanz and Saric, 2009), which could have produced BG phases that may have served or may serve as energy sources for chemolithotrophic microbial activity. This speculation is supported by the recent observation that chemolithoautotrophic Fe(II)-oxidizing organisms are capable of growth via oxidation of biotite, a primary Fe(II)-silicate mineral present in soils derived from granitic rock weathering (Shelobolina et al., 2012b). However, Earth's ocean floor and other rocky planets in our solar system are dominated by basaltic rocks (Taylor and McLenna, 2009). Basaltic rocks contain Fe(II) in the form of both highly stable silicate minerals such as plagioclase and clinopyroxene, as well as olivine and BG, which are generally much less stable and subject to rapid weathering at Earth's surface (Eggleton et al., 1987) and the flanks of mid-ocean ridges (Bach and Edwards, 2003). Hence, Fe(II) in BG represents a potentially significant source of energy for chemolithotrophic Fe(II)-oxidizing organisms on Earth (Bach and Edwards, 2003; Edwards et al., 2003a).
Studies of both marine and terrestrial BG have provided evidence for circumneutral-pH microbial colonization and associated textural alteration (Thorseth et al., 1992; Torsvik et al., 1998; Staudigel et al., 2008; Cockell et al., 2009). Previous studies on marine basalts led to the isolation of a Fe(II)-oxidizing member of the Gammaproteobacteria, which is potentially capable of using BG as an energy source for growth (Edwards et al., 2003b), as well as a marine Pseudomonas isolate and a marine hydrothermal vent enrichment culture that showed colonization and cell growth on basaltic substrates (Bailey et al., 2009). Studies on terrestrial basalts revealed surface colonization by heterotrophic communities (Cockell et al., 2009). Characterization of the interior of terrestrial BG suggests that it is buffered against rapid changes in temperature and is therefore likely to be hospitable to microbial life (Bagshaw et al., 2011). The recent discoveries in Gale Crater by the Mars Curiosity rover (Grotzinger et al., 2014) emphasize the importance of circumneutral-pH systems with variable Fe redox chemistry as potential habitats for past microbial life on Mars.
Despite the well-documented association of microorganisms with altered terrestrial and marine BG, to date there has been no direct demonstration of microbially catalyzed Fe(II) oxidation in BG. This work examined the potential for oxidation of Fe(II) in BG by three different circumneutral-pH FeOB culture systems (see Materials and Methods), including an established chemolithoautotrophic Fe(II)-oxidizing, nitrate-reducing enrichment culture known for its ability to grow via the oxidation of both soluble and solid-phase (mineral-associated) Fe(II) compounds (Straub et al., 1996; Weber et al., 2001; Blöthe and Roden, 2009b; Shelobolina et al., 2012b). For each of these systems, soluble Fe(II) and/or chemically reduced NAu-2 smectite (RS) were used as positive controls to verify the presence of enzymatic Fe(II) oxidation activity. Nitrate, rather than oxygen, was used as the electron acceptor, because (unlike oxygen) it does not react spontaneously with soluble Fe(II) or smectite, thus allowing for a straightforward evaluation of Fe(II) oxidation potential.
2. Materials and Methods
2.1. Preparation of BG
Basaltic glass [nominal chemical formula (FeII,Mg)SiO2] was collected by U.S. Geological Survey volcanologist Frank Trusdell from the TEB flow in Kilauea, Hawaii (see Fig. 1A). Molten lava was quenched in water, air-dried, and shipped without further processing. Within 6 months of collection, chunks of BG were ground by a SPEX shatter box for 10 min and sieved to obtain flakes with particle sizes below 75 μM (see Fig. 1B). Silt and clay-sized fractions (≤20 μM) were obtained through sonication and settling for 30 min (calculated by using Stokes' law) followed by centrifugation and decantation. Extraction with HF-phenanthroline (Komadel and Stucki, 1988) revealed that the BG particles contained 7–8 wt % Fe, virtually all of which (>98%) was present as Fe(II). X-ray diffraction revealed a complete absence of crystalline mineral phases. BG stock suspensions (see Fig. 1C) were prepared anaerobically by adding BG particles to N2-bubbled distilled water at a concentration of 1 g mL−1. The suspensions were autoclaved for 15 min at 121°C. X-ray diffraction analysis showed that autoclaving had no impact on the amorphous nature of the BG. In addition, dissolved Fe(II) was below the detection limit (ca. 2 μM with Ferrozine reagent; see below) in autoclaved BG medium, which indicates that the sterilization procedure did not cause significant mineral dissolution.

Basaltic glass from Kilauea, Hawaii. USGS geologist sampling a lava tube (
2.2. Fe(II)-oxidizing cultures
The first neutral-pH FeOB system employed in this study was the “Straub culture” (Straub et al., 1996; Blöthe and Roden, 2009b), which is known to be capable of growth via the oxidation of insoluble Fe(II)-bearing phyllosilicate phases such as biotite (Shelobolina et al., 2012b), as well as non-silicate Fe(II)-bearing minerals such as siderite and magnetite (Weber et al., 2001). Blöthe and Roden (2009b) showed that the Straub culture is dominated by a betaproteobacterium related to the chemolithoautotrophic FeOB Sideroxydans lithotrophicus strain ES-1 (Emerson and Moyer, 1997; Emerson et al., 2010). The second system was Desulfitobacterium frappieri strain G2, a mixotrophic organism that is capable of nitrate-dependent oxidation of structural Fe(II) in smectite (Shelobolina et al., 2003). The last system was indigenous microorganisms enriched from a neutral-pH groundwater Fe seep located in the Baraboo Hills of Wisconsin. Previous studies have demonstrated the presence of abundant and diverse populations of FeOB in neutral-pH groundwater Fe seeps (e.g., Emerson and Revsbech, 1994; Haaijer et al., 2008; Blöthe and Roden, 2009a; Haaijer et al., 2012; Roden et al., 2012); hence, this system was assumed to contain a range of indigenous Fe(II)-oxidizing taxa with the potential to utilize various forms of Fe(II) as an energy source. For each of these systems, soluble Fe(II) and/or chemically reduced NAu-2 smectite (RS) were used as positive controls to verify the presence of enzymatic Fe(II) oxidation activity. NAu-2 is a natural Fe-bearing smectite phase [nominal formula
All the Fe(II)-oxidizing cultures were grown on anaerobic, bicarbonate-buffered medium (pH 6.8) with nitrate as the electron acceptor, as previously described (Shelobolina et al., 2012b) with either BG, soluble Fe(II) (FeCl2·2H2O), and/or chemically reduced (sodium dithionite) NAu-2 smectite, prepared as described by Benzine et al. (2013) (reaction with excess sodium dithionite in bicarbonate buffer at 70°C for 20 min, followed by five washings with anoxic water and autoclaving). The latter two Fe(II) forms served as positive controls for Fe(II) oxidation activity, since both have been shown to be utilized for growth by a variety of neutral-pH FeOB. The amount of BG or RS added was based on the amount of Fe(II) liberated during a 24 h extraction with 0.5 M HCl; that is, the mass-normalized 0.5 M HCl-extractable Fe(II) content [ca. 140 μmol Fe(II) g−1 for BG, ca. 2400 μmol Fe(II) g−1 for RS] was used to determine how much solid phase was added to 10 mL of medium to obtain 5–15 mmol of 0.5 M HCl-extractable Fe(II) per liter of fluid. We make no specific assumption that 0.5 M HCl-extractable Fe(II) content corresponds to the “microbially available” Fe(II) content of the solids; rather, this extraction simply provided a convenient means by which to track the loss of Fe(II) from insoluble minerals over time (Weber et al., 2001; Shelobolina et al., 2012b). Sterile controls mimicked the experimental conditions without the addition of an inoculum. Experiments to determine the potential for abiotic oxidation of BG and RS by nitrite were likewise conducted in sterile, uninoculated medium.
The Straub culture was maintained in medium containing ca. 5 mM soluble Fe(II) (FeCl2) and 5 mM
Desulfitobacterium frappieri strain G2 was similarly maintained in medium containing 5 mmol Fe(II) L−1 in the form of RS, with 1 mM acetate, 5 mM nitrate, and 0.05% yeast extract. To obtain mineral-free cell inocula for use in BG oxidation experiments, the culture was transferred into nitrate-reducing medium with 1 mM acetate, 5 mM nitrate, 0.05% yeast extract, and 5 mL of H2, which replaced RS as the primary energy source. Cells grown for the third generation under these conditions were transferred into BG medium, with RS used in parallel as the positive control.
The groundwater Fe seep enrichments were initiated with streambed samples collected from Hemlock Draw Preserve, Sauk County, Wisconsin, in April 2013. Water and Fe(III) oxide–containing sediment was collected in sterile 50 mL Falcon tubes and immediately stored at 4°C. One-milliliter samples of the suspension were used to inoculate medium with either BG or RS as the sole electron donor and 5 mM nitrate as the electron acceptor.
Cell growth was not directly monitored in the Fe(II)-oxidizing cultures; hence, our experiments correspond to Fe(II) oxidation assays as opposed to formal growth experiments. However, it should be noted that prior work has demonstrated the ability of both the Straub culture (Straub et al. 1996; Blöthe and Roden, 2009b) and D. frappieri strain G2 (Shelobolina et al., 2003) to couple Fe(II) oxidation to cell growth with nitrate as the electron acceptor. Preliminary studies showed that, by analogy to D. frappieri strain G2 (Shelobolina et al., 2003), the Straub culture could be repeatedly transferred in medium with RS as the energy source (Shelobolina, unpublished data). Hence, it seems likely that cell growth was coupled to Fe(II) oxidation in these cultures.
2.3. Sampling and analysis
One-milliliter culture samples were collected with a N2-flushed needle and syringe every 3–7 days and placed in an anaerobic chamber. The samples were transferred into 2.0 mL microcentrifuge tubes and centrifuged at ca. 13,000 rpm for 5 min. The supernatant was removed, and the pellet was washed twice with sterile culture medium lacking nitrate. One milliliter of 0.5 M HCl was added to the pellet to extract Fe for 24 h. To measure aqueous Fe(II), 0.5 mL of the supernatant was transferred into a 15 mL conical tube with 6 mL of 0.5 M HCl. Both aqueous and HCl-extractable Fe(II) were determined colorimetrically by using Ferrozine (Stookey, 1970). As discussed above, loss of HCl-extractable Fe(II) was used as an indicator of mineral-associated Fe(II) oxidation. Nitrate was determined colorimetrically according to Cataldo et al. (1975). Nitrite was determined by using a colorimetric assay adapted from Wetzel and Likens (1991). The concentration of total dissolved Fe in D. frappieri strain G2 cultures was determined by inductively coupled plasma atomic emission spectroscopy (ICP-OES); ICP-OES was required to determine the low levels of dissolved Fe present in the cultures.
2.4. DNA extraction and 16S rRNA gene sequencing
DNA was extracted from the seep inoculum with the Mo-Bio PowerSoil DNA Isolation Kit according to the manufacturer's protocol. 16S rRNA genes were amplified by using the universal prokaryotic primers 515f/806r (Caporaso et al., 2011). Single samples were multiplexed into the plate, and libraries were prepared by 454 Titanium technology at the University of Wisconsin Biotechnology Center. The sequence data were processed (including denoising) by using the Quantitative Insights Into Microbial Ecology (QIIME) pipeline (Caporaso et al., 2010) and employing the Ribosomal Database Project (RDP) classifier (Wang et al., 2007) for taxonomic assignments. Only the top 10 most abundant taxa that could be classified at the phylum level or better are reported here.
A conventional clone library was used to assess the microbial community in an active smectite-oxidizing enrichment derived from the Fe seep. DNA was extracted as above and PCR amplified with primers 338F and 907R as previously described (Mohanty et al., 2008). The clone library was constructed by using Promega pGEM-T vector and JM109 competent cells. Eighty-four sequences were analyzed with the BioEdit program, and comparisons were achieved with BLAST (Altschul et al., 1997). No further classification was used aside from BLAST.
3. Results
3.1. Activity of Fe(II)-oxidizing cultures
The Straub culture oxidized both soluble Fe(II) (added as FeCl2·2H2O) and structural Fe(II) in RS (Fig. 2A, 2B), as determined by loss of aqueous and/or 0.5 M HCl-extractable Fe(II) content. Although Fe(III) oxides produced during soluble Fe(II) oxidation dissolved in the 0.5 M HCl extracts, this was not the case for Fe(III) in the oxidized RS, because the solubility of oxidized smectite is much lower than that of reduced smectite (Kostka et al., 1996). For simplicity, Fe(II) oxidation was therefore assessed by monitoring the loss of 0.5 M HCl-extractable Fe(II) over time. In contrast to results obtained with soluble Fe(II) and RS, there was no evidence for Fe(II) oxidation in the BG cultures (Fig. 2C). All (within measurement error) of the Fe recovered in the 0.5 M HCl extracts was initially present as Fe(II), and this remained true over time in this and the other culture experiments described below (data not shown). Although nitrate concentrations were not determined in these experiments, previous studies have shown that the Straub culture oxidizes Fe(II) with nitrate according to the 5:1 ratio expected for reduction of nitrate to N2 (Straub et al., 1996; Blöthe and Roden, 2009b; Shelobolina et al., 2012b).
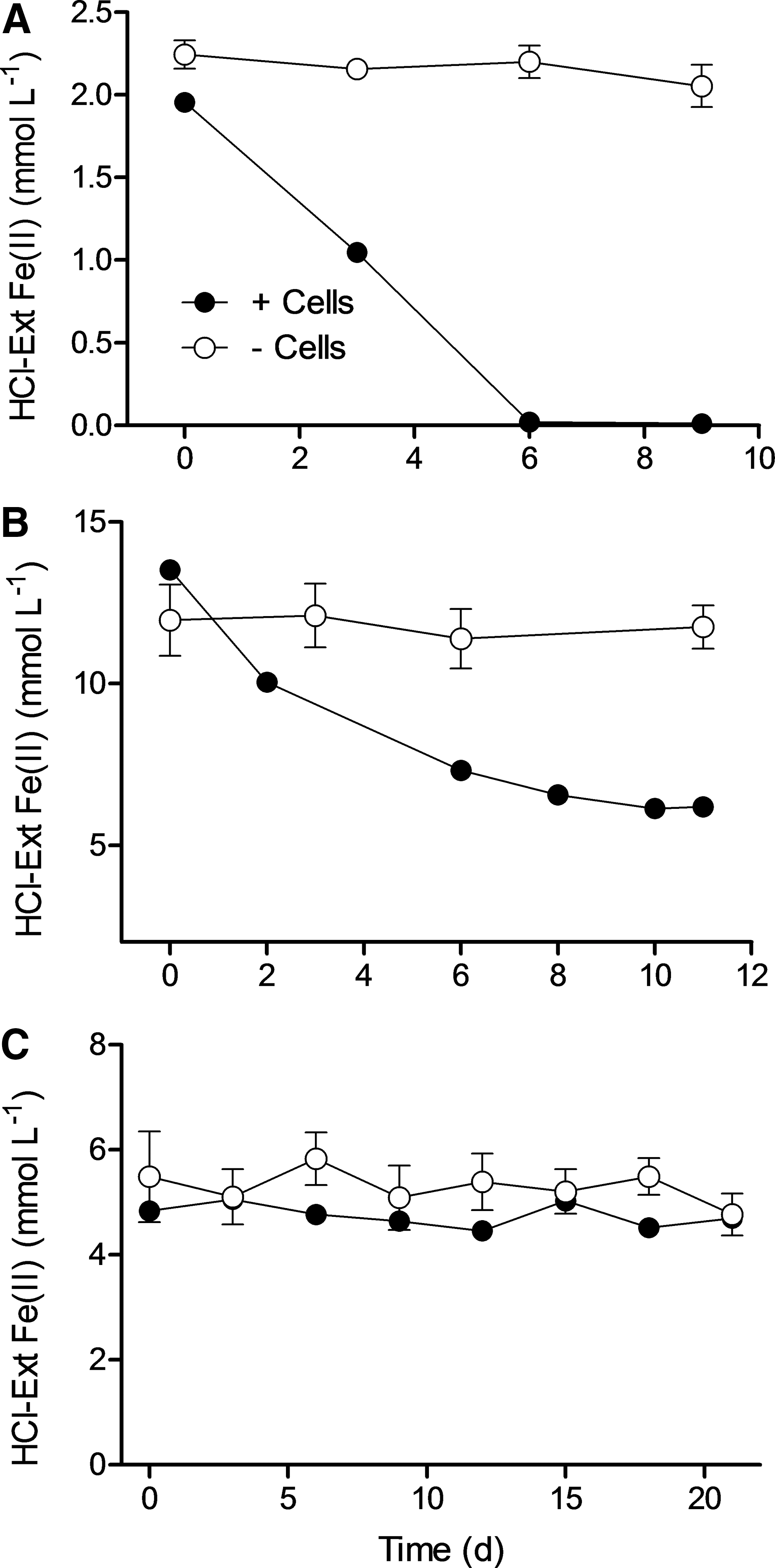
Fe(II) concentrations in Straub cultures containing soluble Fe(II) (
Desulfitobacterium frappieri strain G2 showed substantial oxidation of RS with concomitant consumption of nitrate (Fig. 3A, 3B). The amount of nitrate consumed was much greater in terms of electron equivalents than the amount of Fe(II) oxidized, which is attributed to organotrophic metabolism of yeast extract included in the mixotrophic medium. Nitrite was initially present at a concentration of 0.5 mM and increased with the onset of nitrate reduction (Fig. 3B). Because nitrite oxidizes Fe(II) in RS under the conditions of our experiments (Fig. 4A, 4B), it is possible that some of the observed Fe(II) oxidation took place abiotically through reaction with nitrite. Previous studies with D. frappieri strain G2 have shown, however, that not all RS oxidation activity is linked to abiotic reaction with nitrite, and that cell growth is linked to Fe(II) oxidation (Shelobolina et al., 2003).

HCl-extractable Fe(II) (
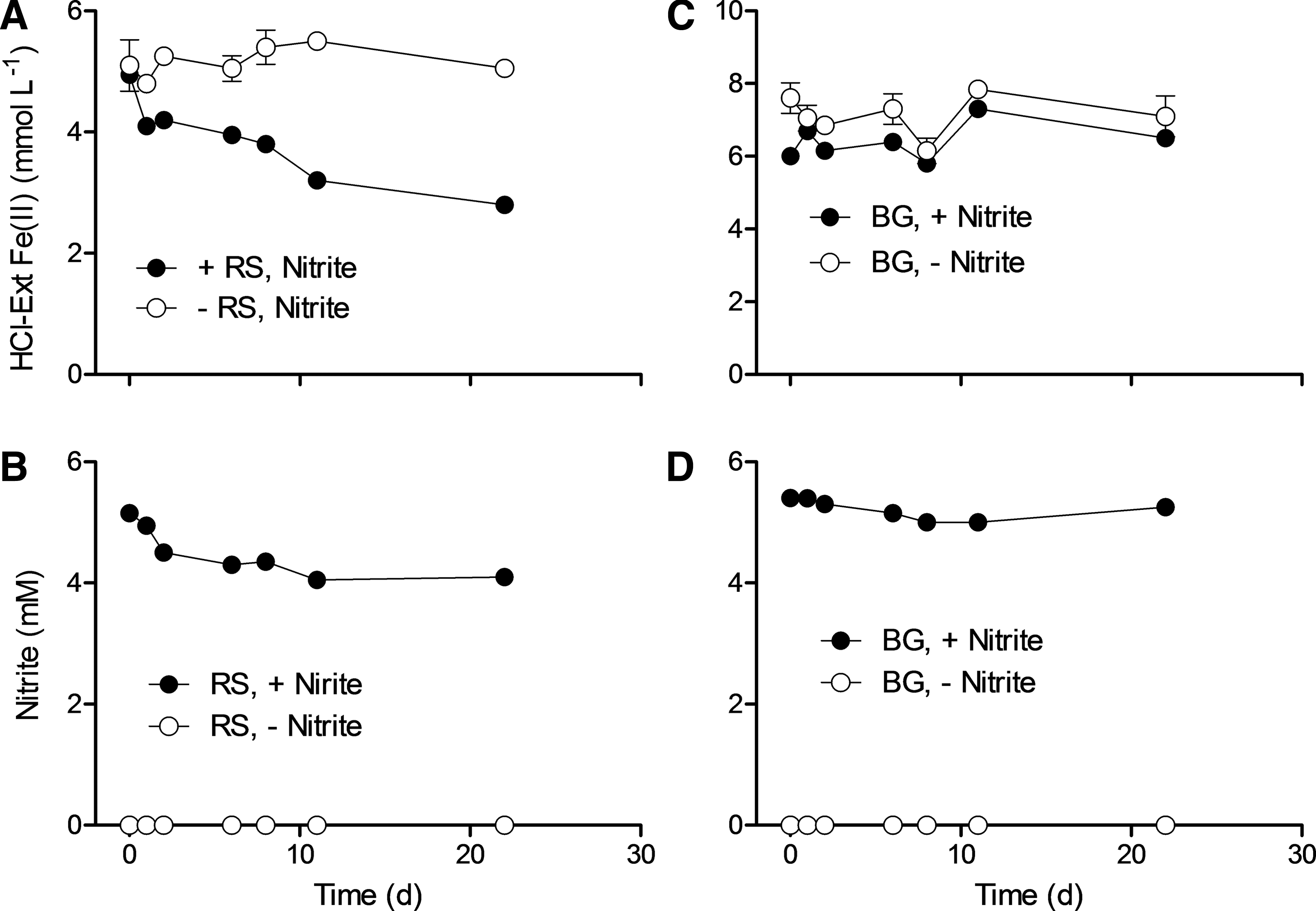
HCl-extractable Fe(II) (
In BG containing D. frappieri strain G2 cultures, the concentration of Fe(II) fluctuated between 8.1 and 7.7 mM compared to between 7.8 and 6.7 mM in the sterile controls (Fig. 3C); thus, like the Straub culture, D. frappieri strain G2 did not oxidize Fe(II) in BG. Although nitrite accumulated in the medium through organotrophic nitrate reduction (Fig. 3D), parallel studies showed that nitrite was unable to oxidize BG in our culture system (Fig. 4C, 4D).
The Fe seep enrichments showed active oxidation of soluble Fe(II) (Fig. 5A) and RS (Fig. 6A) but not BG (Fig. 6C). The large amount of nitrate consumption and substantial accumulation of nitrite in these experiments (Fig. 6B, 6D) was likely driven by oxidation of organic matter in the streambed inoculum. As was the case for the D. frappieri strain G2 cultures, some of the observed Fe(II) oxidation may have been linked to abiotic reaction with nitrite. However, the experiment with soluble Fe(II) (Fig. 5A) showed Fe(II) oxidation with only small amounts (≤ca. 0.04 mM) of nitrite formed (Fig. 5B), which suggests that organisms capable of enzymatic Fe(II) oxidation were present in the seep materials. This explanation is consistent with a 16S rRNA gene pyrosequencing library for the enrichment culture inoculum, which was dominated by operational taxonomic units (OTUs) related to the known FeOB taxa Gallionellaceae (Emerson et al., 2010) and Comamonadaceae (Blöthe and Roden, 2009a) (Fig. 7). The library also contained abundant OTUs associated with the genus Crenothrix, a methane-oxidizing organism (Stoecker et al., 2006) that is often associated with iron-oxidizing microbial mats (Quaiser et al., 2014) and other analogous environments (Wolfe, 1960; Tuhela et al., 1997). In addition, a conventional Sanger-based clone library of 16S rRNA genes from the RS enrichment showed that it was dominated by organisms of the Acidovorax genus (approximately 85% of the clones shared 97% similarity to Acidovorax delafieldii strain PCWCS4), multiple strains of which have been demonstrated to oxidize Fe(II) mixotrophically (Muehe et al., 2009; Byrne-Bailey et al., 2010).
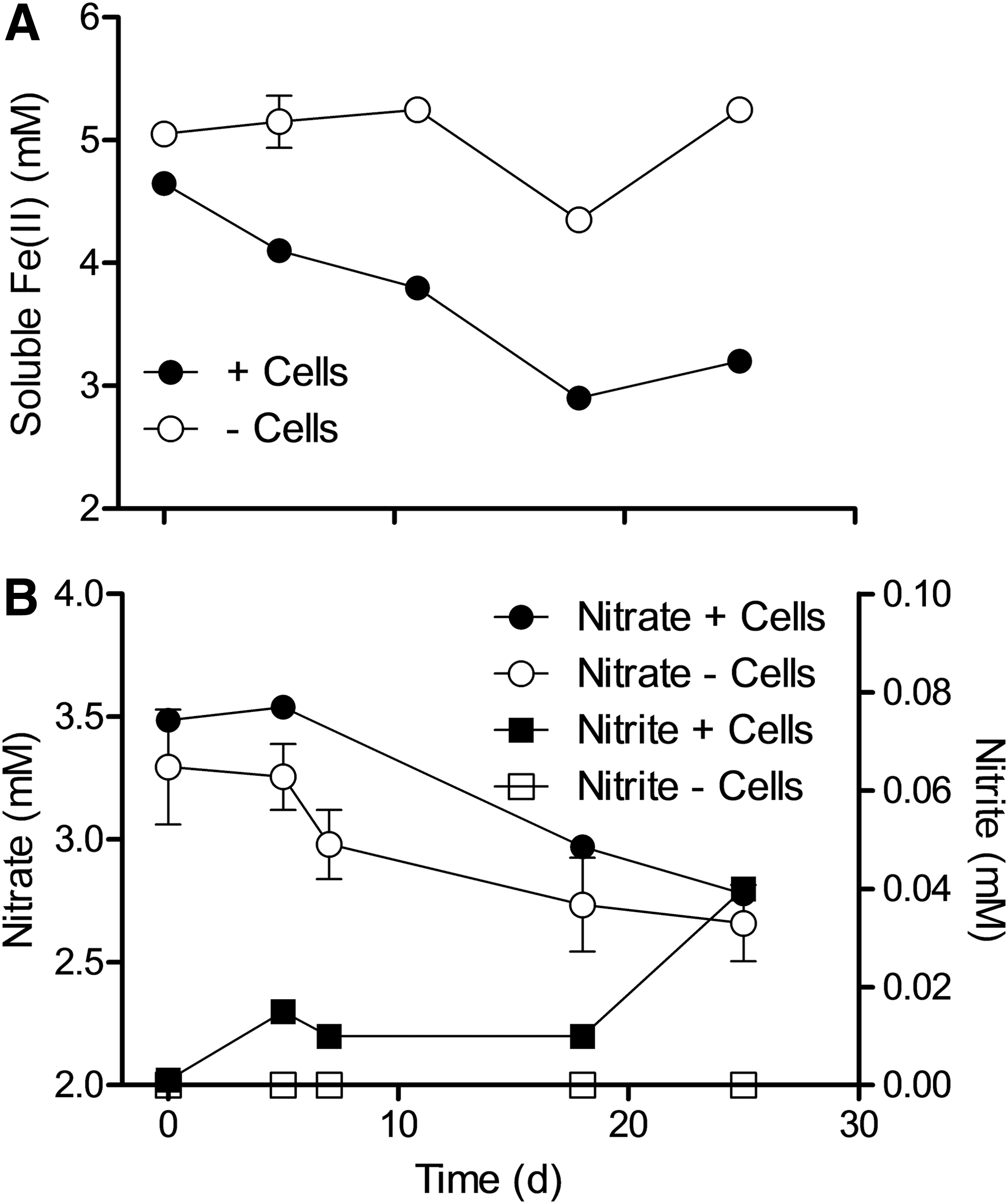
Soluble Fe(II) (

HCl-extractable Fe(II) (

16S rRNA gene pyrosequencing results for the Fe seep enrichment culture inoculum. Only the top 10 most abundant taxa that could be classified at phylum level or better are shown (2485–4346 reads). The abbreviations in parentheses indicate the level of taxonomic resolution (C=class, O=order, F=family, G=genus). (Color graphics available at
4. Discussion
The central aim of this study was to assess the potential for chemolithotrophic FeOB to oxidize Fe(II) in BG under circumneutral pH conditions. The underlying motivation for the work was the concept that Fe(II) in BG [and possibly other Fe(II)-bearing silicate minerals] could potentially support microbial life on Mars (Jakosky and Shock, 1998). Fe(II) is viewed as one of the most likely sources of energy for lithotrophic microbial life on Mars that could utilize either traces of oxygen (present at ca. 0.2% in the martian atmosphere) or nitrate as an electron acceptor (Jepsen et al., 2007). The free energy release during Fe(II) oxidation is similar with oxygen or nitrate as the electron acceptor (Jepsen et al., 2007); hence, from a thermodynamic perspective, these two oxidants are interchangeable in terms of their potential to support chemolithotrophic metabolism. In fact, many neutral-pH FeOB are capable of utilizing either oxygen (under microaerophilic conditions) or nitrate (under anoxic conditions) as an electron acceptor (Benz et al., 1998).
Santelli et al. (2001) showed that the acidophilic FeOB Acidithiobacillus ferroxidans could grow via oxidation of Fe(II) released during dissolution of the Fe(II)-silicate mineral fayalite. Fe(II)-silicate phases (including amorphous BG) are, however, quite insoluble and slow to dissolve at neutral pH, with dissolution rates at pH 7 being 103- to 104-fold lower than at pH 2 (Bandstra and Brantley, 2008). Although a few prior studies have suggested that FeOB could grow via oxidation of BG at neutral pH (Edwards et al., 2003b; Bailey et al., 2009), none of these studies demonstrated a direct linkage between cell growth and Fe(II) oxidation. It is also important to note that recent studies of microbial communities associated with basaltic rocks in the vicinity of hydrothermal vents at the Loihi seamount [from which a variety of FeOB have been identified and isolated in pure culture (Emerson and Moyer, 2002; Emerson et al., 2007)] suggest that FeOB proliferate via oxidation of soluble Fe(II) emitted from the vents as opposed to direct attack of mineral surfaces (Templeton et al., 2009).
Our results show that Fe(II) in BG was not utilized as an electron donor by three different microbial Fe(II) oxidation systems, all of which were capable of oxidizing either soluble Fe(II) or structural Fe(II) contained in RS, a secondary phyllosilicate mineral. It is important to note that the studies of Edwards et al. (2003b) and Bailey et al. (2009) involved samples from the seafloor near active hydrothermal fields, while we sampled from a Fe seep and our pure/enrichment cultures were not from the seafloor. Thus, our culture systems are not directly comparable to those of Edwards et al. (2003b) and Bailey et al. (2009). However, the inability of the Straub culture, which is able to oxidize the highly insoluble Fe(II)-silicate mineral biotite (Shelobolina et al., 2012b), to attack Fe(II) in BG particles of comparable or smaller size compared to the biotite flakes provides convincing evidence of the inaccessibility of Fe(II) in BG toward enzymatic oxidation. Although the mechanism for biotite oxidation by the Straub culture remains unknown, there was no evidence for solubilization of Fe via production of siderophores or other Fe-chelating compounds. Hence, Shelobolina et al. (2012b) argued for direct enzymatic attack of edge sites on the biotite particles. It seems possible that the absence of edge sites in the amorphous glass renders the material resistant to such attack. The idea that specific edge sites are involved in microbially driven mineral oxidation is consistent with the observation that acidophilic FeOB preferentially attached to pyrite parallel to the {100} and {110} crystal faces (Edwards et al., 1998). Detailed spectroscopic analyses analogous to those performed on silicate melts (e.g., Jackson et al., 2005) may help provide insight into how Fe bonding environments on the amorphous BG surface could influence their susceptibility to enzymatic attack.
Desulfitobacterium frappieri strain G2 is known to solubilize Fe during mixotrophic growth on RS with nitrate as the electron acceptor [with accumulation of micromolar amounts of aqueous, presumably organically complexed Fe(III)], a process which may play a role in facilitating mineral-bound Fe(II) oxidation (Shelobolina et al., 2003). This capacity led us to evaluate whether a similar process could facilitate oxidation of Fe(II) in BG by this organism. ICP-OES analysis of the D. frappieri strain G2 BG cultures revealed very low levels (<0.1 μM) of soluble Fe, with concentrations in inoculated cultures slightly lower than those in uninoculated controls (Fig. 8). These results indicate that D. frappieri strain G2 was not able to solubilize Fe from BG, which presumably played a role in limiting the organism's ability to oxidize Fe(II) in glass. Solubilization of Fe has been suggested to play a role in BG oxidation by FeOB, in particular the strain of Marinobacter isolated by Edwards et al. (2003b), which is consistent with generation of siderophores by the closely related Marinobacter aquaeolei strain VT8 organism (Homann et al., 2009) and the presence of siderophore-related genes in the organism's genome (Singer et al., 2011). However, the role of siderophores in mineral-bound Fe(II) oxidation by M. aquaeolei, and whether or not this organism can in fact grow via BG oxidation, remains to be determined. The same is true for the wide range of other aerobic and nitrate-reducing neutral-pH FeOB that have not yet been tested on BG, including recently identified alphaproteobacteria related to the genus Bradyrhizobium that are capable of enzymatic oxidation of Fe(II) in RS or biotite (Shelobolina et al., 2012a; Benzine et al., 2013).
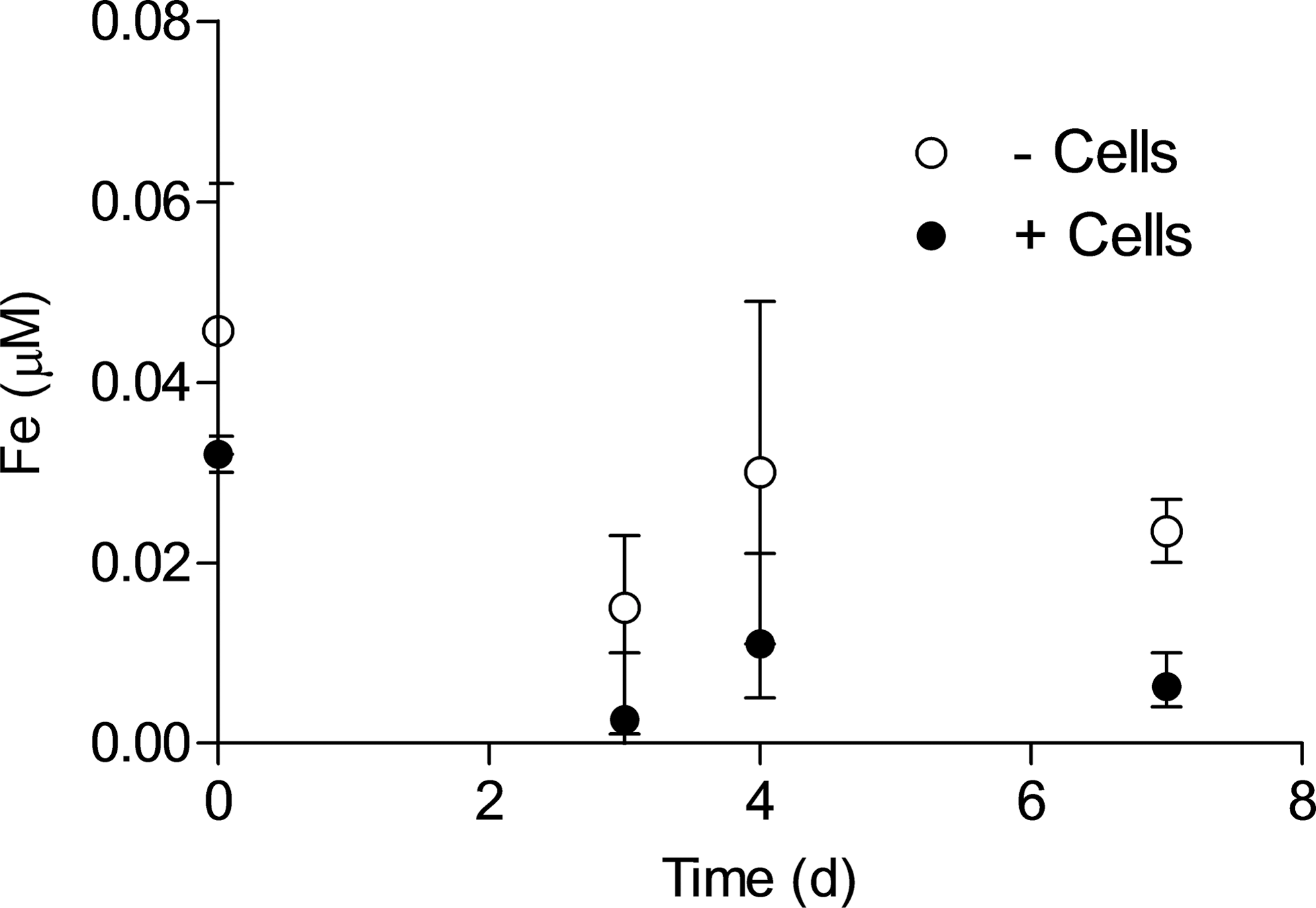
Soluble Fe concentrations in D. frappieri strain G2 BG cultures. Data represent mean±SD of triplicate cultures.
5. Conclusions
The results of this study show that Fe(II) in BG was not available for enzymatic oxidation by three different types of neutral-pH FeOB systems. These findings have important implications toward the long-standing speculation that Fe(II)-Si-rich phases in basalt rocks could provide a basis for chemolithotrophic microbial life on Mars (Jakosky and Shock, 1998), specifically in neutral-pH environments where acid-promoted mineral dissolution and utilization of dissolved Fe(II) as an energy source is not likely to take place. A thorough analysis of the capacity for various other FeOB (including both pure cultures and native FeOB communities analogous to those in the Fe seep) to oxidize BG [and other reactive Fe(II)-silicate phases such as olivine] is required to verify the generality of our findings. In particular, our results suggest the need for a broad-scale culturing campaign with the use of native BG phases as the Fe(II) source for enrichment in terrestrial and marine environments where geochemical and mineralogical data suggest that oxidation of Fe(II) in BG is taking place under neutral-pH conditions.
Footnotes
Acknowledgments
This work was supported by the NASA Astrobiology Institute. We gratefully acknowledge B. Converse for assistance with the 16S rRNA gene pyrosequencing analysis and Tao Wu for assistance with mineralogical analysis.