
Abstract
Fe-(oxyhydr)oxide-encrusted filamentous microstructures produced by microorganisms have been widely reported in various modern and ancient extreme environments; however, the iron-dependent microorganisms preserved in hydrothermal quartz veins have not been explored in detail because of limited materials available. In this study, abundant well-preserved filamentous microstructures were observed in the hydrothermal quartz veins of the uppermost dolostones of the terminal-Ediacaran Qigebulake Formation in the Aksu area, northwestern Tarim Basin, China. These filamentous microstructures were permineralized by goethite and hematite as revealed by Raman spectroscopy and completely entombed in chalcedony and quartz cements. Microscopically, they are characterized by biogenic filamentous morphologies (commonly 20–200 μm in length and 1–5 μm in diameter) and structures (curved, tubular sheath-like, segmented, and mat-like filaments), similar to the Fe-oxidizing bacteria (FeOB) living in modern and ancient hydrothermal vent fields. A previous study revealed that quartz-barite vein swarms were subseafloor channels of low-temperature, silica-rich, diffusive hydrothermal vents in the earliest Cambrian, which contributed silica to the deposition of the overlying bedded chert of the Yurtus Formation. In this context, this study suggests that the putative filamentous FeOB preserved in the quartz veins might have thrived in the low-temperature, silica- and Fe(II)-rich hydrothermal vent channels in subseafloor mixing zones and were rapidly fossilized by subsequent higher-temperature, silica-rich hydrothermal fluids in response to waning and waxing fluctuations of diffuse hydrothermal venting. In view of the occurrence in a relatively stable passive continental margin shelf environment in Tarim Block, the silica-rich submarine hydrothermal vent system may represent a new and important geological niche favorable for FeOB colonization, which is different from their traditional habitats reported in hydrothermal vent systems at oceanic spreading centers or volcanic seamounts. Thus, these newly recognized microfossils offer a new clue to explore the biological signatures and habitat diversity of microorganisms on Earth and beyond. Key Words: Filamentous microfossils—Fe-oxidizing bacteria—Uppermost Ediacaran—Quartz vein—Submarine hydrothermal feeder system—The earliest Cambrian. Astrobiology 15, 523–537.
1. Introduction
T
Using Fe(II) as their energy source, FeOB grow in acidic to neutral and oxic to anoxic waters (Konhauser, 1998; Fortin and Langley, 2005; Kappler and Straub, 2005; Hedrich et al., 2011; Konhauser et al., 2011; Templeton, 2011; Dubinina and Sorokina, 2014). FeOB cells, excreting extracellular polymeric substances, are often rapidly entombed by Fe-oxyhydroxides due to metabolic activities (Kappler and Newman, 2004; Chan et al., 2009, 2011; Miot et al., 2009; Schädler et al., 2009). Particularly, microaerobic FeOB likely produce distinctive extracellular filamentous structures with Fe-oxyhydroxide encrustations (Edwards et al., 2004; Emerson et al., 2010; McAllister et al., 2011; Posth et al., 2014), such as the representatives of helical stalks of Gallionella and Mariprofundus, and the tubular sheath of Leptothrix and Sphaerotilus (Emerson et al., 2007; Chan et al., 2011; James et al., 2012; Krepski et al., 2013). Increasing pieces of morphological and molecular evidence show that the activity of FeOB must have played an important role in the formation of modern Fe-(oxyhydr)oxide filaments in freshwater and marine environments (Emerson and Moyer, 2002, 2010; Edwards et al., 2004; Emerson and Weiss, 2004; Little et al., 2004; Emerson et al., 2010).
Benefiting from constrained degradation by Fe ion binding, rapid silicification, and diagenetic stability, FeOB filamentous structures can be preserved exceptionally well as recognizable microfossils or casts in the geological record (Ferris et al., 1988; Toporski et al., 2002; Preston et al., 2011; Glasauer et al., 2013; Li et al., 2013; Li and Li, 2013; Sun et al., 2013; Posth et al., 2014). Putative FeOB microfossils with various Fe-(oxyhydr)oxide-encrusted filaments, stalks, and sheaths have been reported in a vast variety of geological deposits spanning from recent times to ∼3.2 Ga (Hofmann and Farmer, 2000; Boyce et al., 2003; Little et al., 2004; Kiyokawa et al., 2006; Slack et al., 2007; Hofmann et al., 2008; Planavsky et al., 2009; Fru et al., 2013). Widely distributed FeOB have the potential to play a considerable role in iron and carbon cycling in various geological niches and protracted geological history (Fortin and Langley, 2005; Kappler and Straub, 2005; Emerson et al., 2010; McAllister et al., 2011; Raiswell and Canfield, 2012; Posth et al., 2014). Thus, exploring the distribution and activity of iron-dependent life has significantly expanded our understanding of iron biogeochemistry and biosignatures on Earth and potentially beyond.
However, less is known about iron-dependent microorganisms hosted by hydrothermal veins, as only a few examples have been reported to date (Hofmann et al., 2008). In this study, we report the rare occurrence of filamentous structures hosted by hydrothermal quartz-barite vein systems in the uppermost Ediacaran Qigebulake Formation, Aksu area, northwestern Tarim Basin, China, which have provided us an opportunity to explore the diversified distribution of microorganisms in a geological environment. By comparison with the well-known biogenic morphologies and mineral assemblages, the biogenicity of the filaments, habitat, and fossilization process of related microorganisms are evaluated in detail. The implications of the microorganisms colonizing hydrothermal veins are further discussed.
2. Geological Setting
Tarim Block (geographically Tarim Basin) is a main tectonic unit and one of the oldest continental blocks in China (Fig. 1A). During the Ediacaran–Cambrian (E–C) transition, the Tarim Block gradually drifted away from the Kimberly area of northwest Australia (Li et al., 1996, 2008; Li and Powell, 2001) and then evolved from a rift to a passive continental margin basin due to continuous regional extension (Lin et al., 2008; Turner, 2010; Lin et al., 2012). The Aksu area, located in the Kalpin Uplift in the northwestern margin of Tarim Basin, was linked to the infant South Tianshan Ocean to the north during the Early Cambrian (Yu et al., 2009). In this area, the E–C boundary succession crops well out in several localities (Fig. 1B) and consists of the uppermost Ediacaran Qigebulake and the Lower Cambrian Yurtus Formations (Fig. 1C). Based on the geological history of the Kalpin Uplift in the Aksu area, the E–C boundary succession there may have reached the maximum burial depth (about 4500 m) in the Early Carboniferous (Chang et al., 2012), corresponding to a peak burial temperature of 145±10°C.

(
The Qigebulake Formation consists of thin- to thick-bedded dolostone with microbialite. The upper part of this formation was subject to subaerial exposure and ensuing karstification during the terminal Ediacaran. Biostratigraphic and C-isotope stratigraphic data suggest that the Qigebulake Formation is roughly correlated with the Dengying Formation of the uppermost Ediacaran (551–541 Ma) in the Yangtze Block, South China (Gao et al., 1985; Yao et al., 2005; He et al., 2007). The Yurtus Formation consists of three lithostratigraphic units (Zhou et al., 2015): the lower chert-phosphorite (about 0.2–7 m in thickness), the middle black interbedded shale-chert with abundant diagenetic barite concretions (about 1–10 m in thickness), and the upper thin-bedded dolostone intercalated with gray to greenish argillaceous dolostone and calcareous shale (about 10–20 m in thickness). The Yurtus Formation was deposited mainly in a continental shelf environment (Yu et al., 2009). The lower part of this formation, commonly bearing small shelly fossils and acritarch assemblages, is suggested to correspond to the lower Meishucunian Stage (∼535–521 Ma) in the Yangtze Block, South China, or lower Nemakit-Daldynian Stage elsewhere (Yao et al., 2005).
The Yurtus Formation unconformably overlies the karstic dolostone of the Qigebulake Formation (Gao et al., 1985). The unconformity between the Qigebulake and the Yurtus formations is characterized by a karstification surface and dissolution-related disruption zone underneath, indicating a large-scale sea-level fall or major tectonic uplift (He et al., 2010). This unconformity was considered to separate the syn-rift strata (the Qigebulake Formation and underlying siliciclastic strata) from the post-rift deposits (the Yurtus Formation and overlying carbonate succession upward to the Upper Ordovician) (Lin et al., 2012), representing a transition from a phase of syn-rift fault-controlled subsidence to a phase of post-rift thermal-controlled subsidence (Turner, 2010). Under this circumstance, a submarine silica-rich hydrothermal system that consists of silica-rich vein swarms in the uppermost Qigebulake dolostones and overlying bedded cherts and black shales in the lowermost Cambrian Yurtus Formation was formed in the Aksu area in the earliest Cambrian (Zhou et al., 2014).
In the Aksu area, silica-dominant vein swarms extensively occur in the uppermost part of the Qigebulake dolostones and terminate immediately under the overlying bedded cherts of the Lower Cambrian Yurtus Formation. The silica-dominant veins preferentially followed the antecedent karstified permeable zone, where abundant previous pore spaces (fissures/fractures, solution pipes, channels, and cavities) could have provided the preferential conduits for hydrothermal fluid circulation. These veins are dominantly plugged by different silica components from poorly crystallized silica (chalcedony and spherulite) to crystallized quartz crystals (micro- to megaquartz) in several generations. Subordinate infills are locally present and include cube-shaped pyrite, euhedral barite crystals, and poorly crystallized Fe-(oxyhydr)oxides. The barite shows similar 87Sr/86Sr ratios (∼0.7087–0.7088, unpublished data) to that of the Early Cambrian seawater, likely indicating the inheritance from the signature of the coeval downward-circulating seawater. Fluid inclusion microthermometry of vein fills suggests that quartz and barite crystals had precipitated from low- to medium-temperature (40–200°C) hydrothermal brine fluids (11.2–25.8 wt % NaCl equivalent) (Zhou et al., 2014). This vein system has been proposed to represent a subseafloor low-temperature stockwork-like feeder zone, which had channeled silica- and barium-rich hydrothermal fluids upward into the seawater, contributing to the deposition of overlying Yurtus siliceous rocks (Zhou et al., 2014).
Samples containing iron-rich filamentous structures were collected from quartz veins in the uppermost part of the Qigebulake dolostones at Linkuanggou (40°53′44″N, 79°48′14″E), Xiaoerbulake (40°54′41″N, 79°50′44″E), and Aksu (41°01′00″N, 80°02′37″E) sections in the Aksu area (Fig. 1D, 1E).
3. Material and Methods
Filamentous structures were studied by a combination of optical microscopy, field emission scanning electron microscopy (FESEM) with energy-dispersive spectrometry (EDS), and Raman spectrometry. Thin sections (∼30 and ∼300 μm thick, respectively) were studied to evaluate the biological nature of the filamentous-like objects. During preparation of the thin sections, precautions were taken to limit the risk of contamination.
Ultramicroscopic fabrics and microstructures were mainly examined under a Supra 55 field emission scanning electron microscope at China University of Geosciences (Beijing). Thin sections and freshly broken fragments coated with platinum were used in FESEM observation. FESEM study of most samples was operated at 10–15 kV with a working distance of 3.0–9.4 mm. Semiquantitative element concentrations of micron-sized spots were analyzed by an Oxford energy dispersive X-ray spectrometer connected to a field emission scanning electron microscope, operated at 15 kV with a working distance of 8.5 mm.
To determine the properties of filamentous materials, some filamentous structures were observed in the sections by a LabRAM HR800 Raman spectrometer at the Institute of Geology and Geophysics, Chinese Academy of Sciences (IGGCAS). The system was equipped with automated x–y micro-sampling stage, 1800 grooves/mm diffraction grating, and charge-coupled device detector. The Raman microscope operating confocally at laser excitation wavelength of 532 and 633 nm with a spectral footprint of 1 μm gave a spectral resolution better than 1.5 cm−1. An Olympus 100× objective was used. Each spectrum required 1 min total scan time.
4. Morphology and Composition of Filaments
4.1. Optical microscopy and scanning electron microscopy observation
Microscopically, filamentous structures that are dark, ocherous, or rusty in color are conspicuously distributed in the quartz veins. They are not ubiquitous in the siliceous cement but are more abundant in boundary zones between the microquartz or macroquartz cement and the dolostone host rock (Fig. 2A). Generally, they occur either as discrete filaments (Fig. 2B) or are intertwined together as porous, mycelium- or mat-like networks that radically grow outward from the substrate and extend into the pore space available (Fig. 2C). The filamentous structures vary in a wide range of size and morphology, including straight to highly curved, sheath-like, segmented forms (Fig. 2D–2I). The straight to highly curved filaments, which appear to be flexible, are abundant and frequently intertwined with each other. Sheaths are characterized by hollow tube-like structures with smooth or rough outlines (Fig. 2D). Segmented filaments extend discontinuously as long chain-like structures (Fig. 2E, 2F). The filaments are commonly 20–200 μm (some up to several hundreds of micrometers) in length and 1–5 μm in diameter with walls of about 1 μm in thickness. Most of the filaments have a constant width relative to their length. All the filaments were permineralized by poorly crystallized Fe-(oxyhydr)oxides along filaments (Fig. 2G, 2H). They were entombed in spherulite, chalcedony, and/or crystallized quartz (micro- to macroquartz). In some cases, filaments extend across the crystal boundaries of quartz (Fig. 2D) or growth rings of spherulite (Fig. 2I).
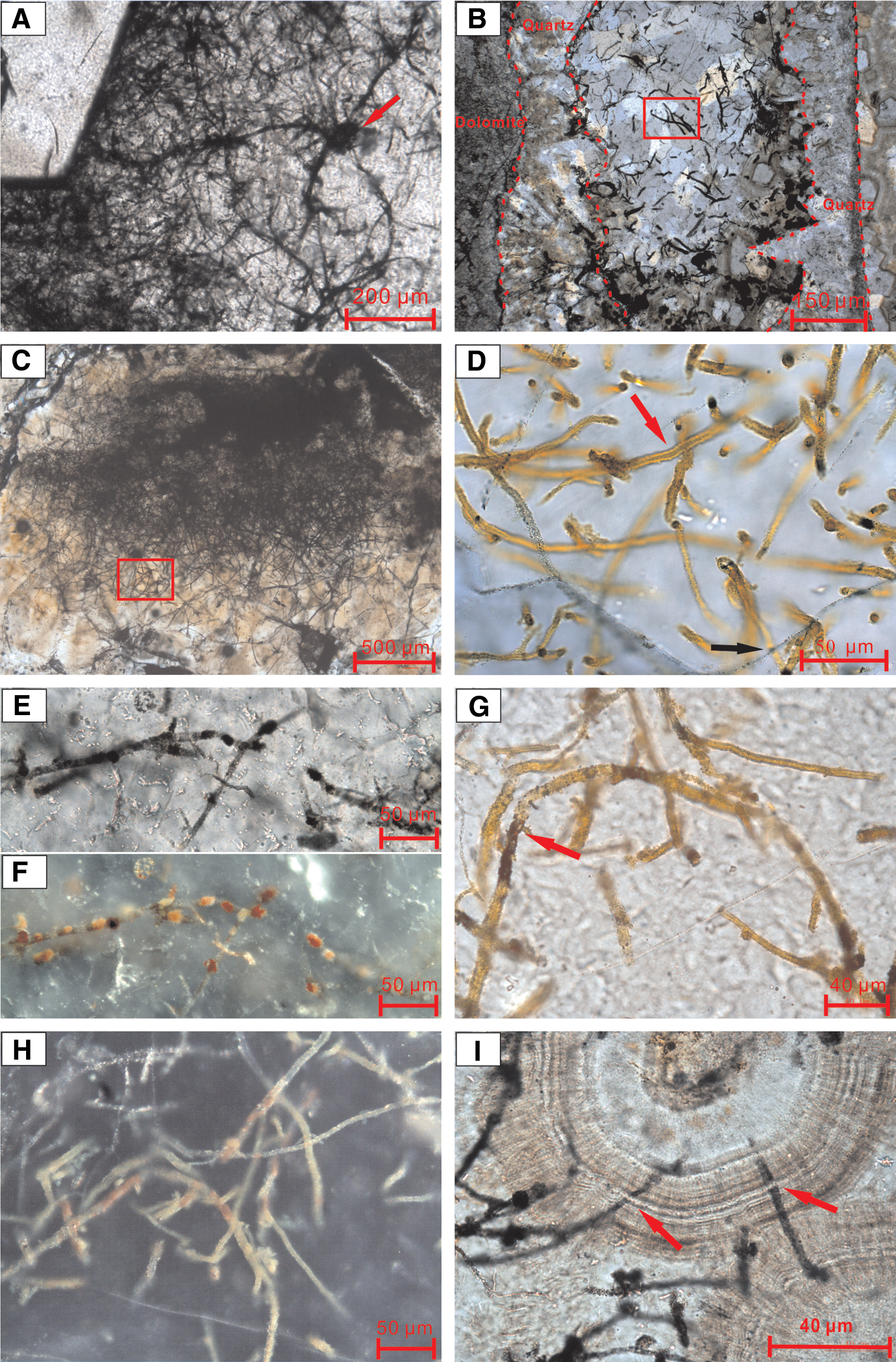
Photomicrograph of filamentous microstructures in thin sections from quartz veins in the uppermost Qigebulake dolostone in Aksu area. (
Scanning electron microscopy observations did not show any distinct filament structures. However, fibril-like fabrics that resemble organomineralization textures (likely mineralized extracellular polymeric substances) have been discovered near the quartz crystals (Fig. 3A, 3B). EDS analysis results suggest that the surface of these structures is dominated by Si, Fe, and O (Fig. 3C, 3D).

(
4.2. Raman spectroscopy
Raman spectroscopy is a useful method to detect bond types, functional groups, and compound classes (Ellery et al., 2004; Villar and Edwards, 2006; Marshall et al., 2010; Jehlička et al., 2014) and thus has been widely used in evaluation of purported fossils (Schopf et al., 2002; Edwards et al., 2007; Schopf and Kudryavtsev, 2009; Marshall et al., 2011; Heim et al., 2012; Tang et al., 2013). Raman spectroscopy was used to identify the material of filament structures in the thin sections (Fig. 4). All analyzed spots show diagnostic vibrational bands of quartz at 127, 206, and 465 cm−1 (Villar and Edwards, 2006), which is consistent with microscopic observations. Importantly, Raman spectroscopy analyses also revealed that the Fe-encrusted filaments mostly consist of goethite with Raman bands at 300, 387, 483, 558, and 686 cm−1 and hematite with Raman bands at 613 and ∼1318 cm−1 (Villar and Edwards, 2006; Marshall et al., 2011), though there is a lack of carbonaceous material (Fig. 4B and 4D). Raman bands at 1464 and 1527 cm−1, visible at laser excitation wavelength of 532 nm, remain to be identified.
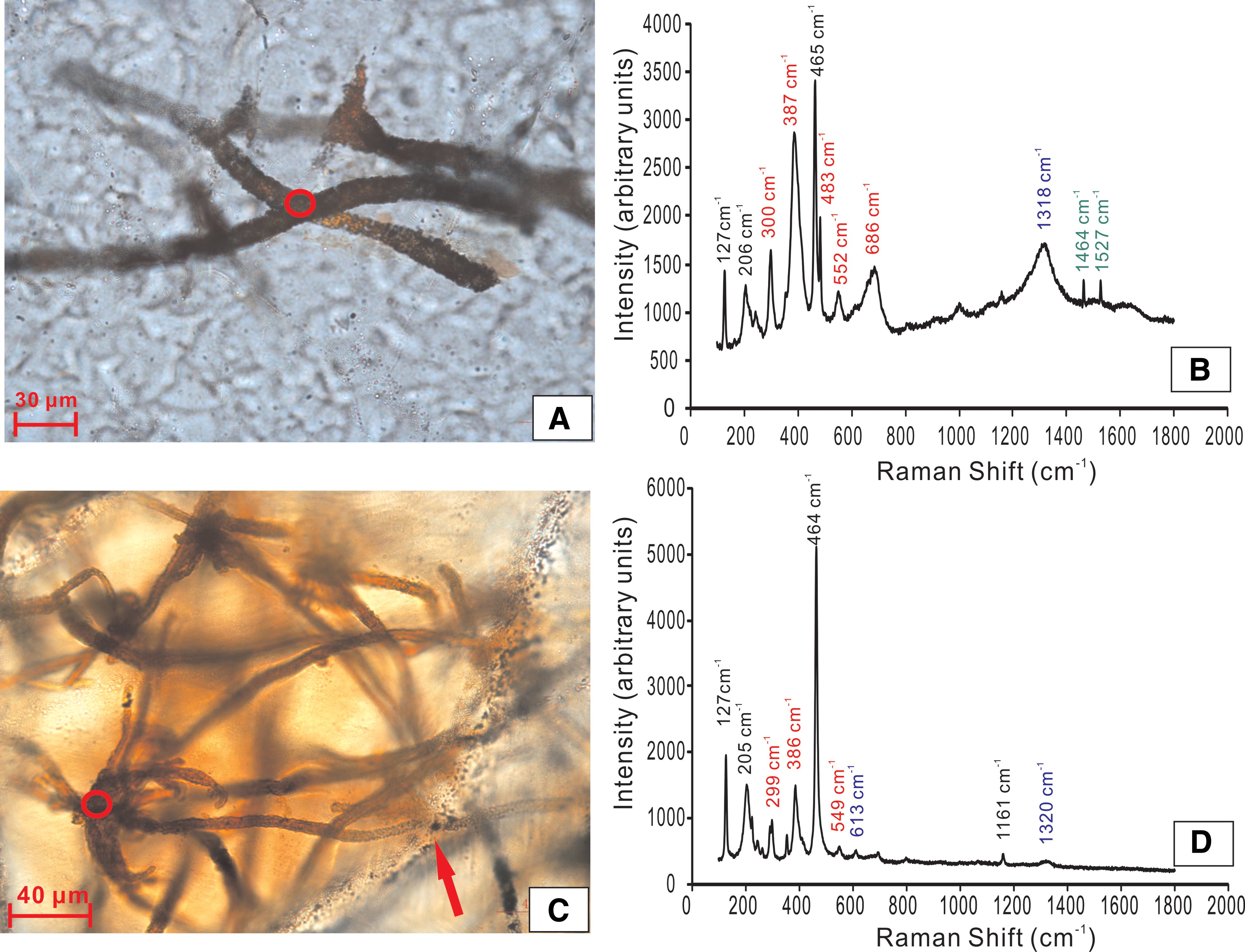
(
5. Discussion
5.1. Biogenic origin of filamentous microstructures
Two competing hypotheses, biological (microbial filaments) or abiotic (mineral fibers) origin, have been proposed to explain the formation of the filamentous microstructures in various geological environments (Hofmann et al., 2008). Although silica/barium carbonate biomorphs have been synthesized under abiotic conditions (García-Ruiz, 1998; García-Ruiz et al., 2002, 2003, 2009), filaments permineralized by Fe-(oxyhydr)oxide have been proposed to be formed by abiotic processes only in rare cases, such as in the submarine hydrothermal self-organized agate (Hopkinson et al., 1998). The criteria and reliability for identifying filamentous microfossils from ancient rocks have been summarized in recent review papers (Cady et al., 2003; Little et al., 2004; Hofmann et al., 2008). Generally, microbial filaments bear distinctive morphological structures (segmented filaments, tube-like sheaths, twisted stalk, branching patterns, internal septate structures, terminal knobs, dendritic and mat-like networks) and geometric parameters (including size, widths, bending, directionality, and tortuosity), which are fundamentally different from abiotic fabrics and valuable for comparison and evaluation of the biogenicity of filaments. Due to diagenetic alternation of organic and mineralogical biomarkers and ambiguous interpretation of geochemical signatures (i.e., Fe isotope) (Planavsky et al., 2009; Moeller et al., 2014), Fe-(oxyhydr)oxide filaments with complex structures are therefore the most valuable biosignatures.
In the present study, we also observed many morphological variants of filamentous microstructures in silica-filled veins in the Qigebulake dolostones (Fig. 2). These structures were completely entombed in the vein-filling silica components (Fig. 2) and locally extended across the crystal boundary or the growth ring of chalcedonic spherulites (Fig. 2D, 2I), indicating that they lived in the open space of veins prior to silica precipitation. However, entombment of filaments by silica precipitation must have followed immediately, which is essential for better preservation of filaments of high abundance (Fig. 2). In the absence of known abiotic processes that created similar Si/Fe-based morphologies (García-Ruiz et al., 2003; Hofmann et al., 2008; Chan et al., 2011), these micron-scale fibril-like fabrics nearby quartz crystals (Fig. 3) display typical biogenic features, which may be associated with organomineralization by filamentous bacteria. Their occurrence may imply that silica-filled vein space harbored microorganisms prior to filling or cementation (Fig. 3A). This is further supported by the clumped distribution pattern of curved elongate filaments (Fig. 2B), which is commonly recognized as a signature of filamentous bacteria, rather than abiotic precipitation (Trewin and Knoll, 1999). The porous networks of mycelium-like filament textures probably represent microbial growth forms (Fig. 2C). Collectively, these iron-rich filaments are quite different from morphologies associated with abiogenic mineral fibers and self-organized patterns (Hopkinson et al., 1998; Hofmann et al., 2008) and mineralization from synthesized silica/barium carbonate biomorphs (García-Ruiz, 1998; García-Ruiz et al., 2002, 2003, 2009). Thus, they are likely of biogenic origin.
5.2. Microfossils of Fe-oxidizing bacteria
Filamentous microstructures in quartz veins in the Qigebulake Formation have shown to be derived from microorganism activities. However, the microorganisms responsible for their formation are yet unknown. Traditionally, it has been understood that the filamentous microstructures in extreme environments were formed by microorganisms such as FeOB (Little et al., 2004), archaea (Mason et al., 2007, 2008), fungi (Kretzschmar, 1982; Schumann et al., 2004; Ivarsson et al., 2012), or uncertain life-forms (Hofmann and Farmer, 2000).
In the present study, discrete encasement and preservation of filaments in quartz crystals without visible dissolution are inconsistent with the scenario produced by boring endoliths, which were definitely introduced into the minerals later and dwelled in fractures and vein walls (McLoughlin et al., 2007, 2010; Ivarsson and Holm, 2008). There is no clear evidence of branching, anastomosis, and septa in filamentous structures, indicating that they are not likely to be fossilized fungi (Schumann et al., 2004; Ivarsson et al., 2012). As for the unique occurrence in subsurface quartz veins where sunlight was absent, filaments could not have derived from the Fe-mineralized cyanobacteria or other filamentous photosynthetic bacteria (Krepski et al., 2013). In contrast, they can be best interpreted as microfossils of FeOB as discussed below.
Morphologically, the various filamentous microstructures in Qigebulake quartz veins (Fig. 2) bear striking resemblance to the remains of FeOB preserved in the geological record (Little et al., 2004; Hofmann et al., 2008) and modern seafloor Fe-Si deposits associated with oceanic hydrothermal vents (Table 1). In particular, Fe-(oxyhydr)oxide tubular sheaths with widths of ∼1–5 μm and lengths of approximately tens of micrometers in the quartz veins (Fig. 2) are comparable with those of the modern freshwater FeOB Sphaerotilus/Leptothrix group (van Veen et al., 1978; Emerson and Revsbech, 1994; Spring, 2006; Fleming et al., 2011) and a novel group of marine iron-oxidizing Zetaproteobacteria that generally occur in hydrothermal vent fields such as Loihi Seamount (Emerson and Moyer, 2002; Fleming et al., 2013), Franklin Seamount (Boyd and Scott, 2001), Juan de Fuca Ridge (Kennedy et al., 2003a, 2003b, 2003c), submarine volcanoes along the Kermadec Arc (Hodges and Olson, 2009), and iron–silica-rich precipitates of the Lau Basin (Sun et al., 2013). Typical stalk-like filaments have not been recognized yet.
D: diffuse vent. LHV: low-temperature hydrothermal vent.
Mineralogically, filaments encrusted by goethite and hematite (Fig. 4), which may have transformed from metastable ferrihydrite during later diagenesis, are consistent with the metabolic products of FeOB (Boyd and Scott, 2001; Kennedy et al., 2004; Posth et al., 2014). Notably, because of the relatively low maximum burial temperature (145°C±10°C) of E–C strata in the Aksu area (Chang et al., 2012) and complete entombment by silica cements (Posth et al., 2014), the bacteriogenic goethites in the quartz veins were well protected and thus mostly survived the complete transformation into the hematite during later burial and thermal evolution over a protracted time period. Nevertheless, the coexistence of hematites (Fig. 4B) does reflect to some extent phase transformation from goethite by thermal dehydration during the late burial. On the other hand, it is reasonable that organic material has not been detected within the studied filament structures by laser Raman spectroscopy because chemolithotrophic FeOB generally contain only a few weight percent of organic carbon (Boyd and Scott, 2001; Emerson and Moyer, 2002; Suzuki et al., 2011; James et al., 2012; Bennett et al., 2014), which would have been readily lost by rapid decomposition in the oxidized environment or by mineralization (Toporski et al., 2002; Posth et al., 2014). In summary, abundant Fe-(oxyhydr)oxide-encrusting filaments in quartz veins share the common morphological and mineralogical characteristics of iron-rich filaments found in modern and ancient hydrothermal vent fields as described above and are distinctive enough to be the FeOB microfossils.
5.3. Habitat and fossilization
The presence of filamentous FeOB microfossils in Qigebulake quartz veins may provide valuable information for a particular paleoenvironment. Generally, morphologically recognizable extracellular structures, such as tubular sheath and twisted elongate stalk, are exclusively produced by obligate neutrophilic, microaerobic Fe-oxidizers (Kappler and Straub, 2005; Emerson et al., 2010; Konhauser et al., 2011; Posth et al., 2014). They grow in aqueous fluid habitats at neutral pH and redox states that are characterized by low concentrated but high and steady fluxes of both Fe(II) and O2 (Sobolev and Roden, 2001; James and Ferris, 2004; Emerson et al., 2010; Krepski et al., 2013). Correspondingly, they generally live in a wide spectrum of environments from freshwater (e.g., wetlands, lakes, springs, water wells and pipelines, groundwater discharge zones, and aquatic sediments) (Emerson and Revsbech, 1994; Emerson and Moyer, 1997; Neubauer et al., 2002; Emerson and Weiss, 2004; James and Ferris, 2004; Sobolev and Roden, 2004; Baskar et al., 2008; Blöthe and Roden 2009; Preston et al., 2011; Kato et al., 2012) to marine environments (e.g., shallow bay iron seeps, seafloor basalts, oceanic crust, deep-ocean hydrothermal vents) (Emerson and Moyer, 2002, 2010; Hanert, 2002; Emerson et al., 2007, 2010; McAllister et al., 2011; Templeton, 2011).
However, filamentous microfossils of FeOB that dwelled in the Qigebulake quartz veins are quite enigmatic in view of their unique habitat. Tectono-thermal evolution and burial history reveal that the uppermost Ediacaran unit in the Aksu area might have been uplifted to a subaerial environment only during the terminal Ediacaran prior to Yurtus Formation deposition and recent time (since the Cenozoic) (Chang et al., 2012). Since this unit was deeply buried in the subsurface (up to a several kilometers' depth due to the vast thickness of Cambrian–Permian succession), this kind of extreme environment has not been reported to be hospitable for microaerobic, filamentous FeOB yet (Hofmann et al., 2008; Emerson et al., 2010). On the other hand, geological evidence does not support the presence of subaerial siliceous hydrothermal systems from the terminal Ediacaran onward, because related siliceous sinter deposits on the subaerial exposure surface (Jones and Renaut, 2003) have not been discovered. Furthermore, the high salinity levels (approximately 3–7 times seawater salinity) of primary fluid inclusions in the quartz veins (Zhou et al., 2014) also do not support the occurrence of paleo-subaerial hydrothermal systems, which would generally have been dominated by recirculating meteoric water. Similarly, there is no indicator to support that subaerial hydrothermal springs or hydrothermal events occurred during the Cenozoic uplift stage. Therefore, silica-rich veins in the Qigebulake Formation that contain abundant filamentous microfossils cannot be causally related to a subaerial siliceous hydrothermal system.
In contrast, the quartz-barite vein swarms in the upper part of Qigebulake dolostones have been reasonably interpreted to be subseafloor feeder remnants of a silica-rich hydrothermal vent system that was developed in the very Early Cambrian (Zhou et al., 2014). This suggestion is evidenced by the specific vertical stacking occurrence that the quartz veins terminate immediately under the overlying bedded chert successions without upward penetration. Moreover, the characteristic textural and compositional features of these veins, which are analogous to the hydrothermal silica-barium deposits on the modern oceanic seafloor (Herzig et al., 1988; Urabe and Kusakabe, 1990; Stüben et al., 1994; Bogdanov et al., 1997; Bohrmann et al., 1999; Halbach et al., 2002; Hein et al., 2008) and ancient examples (Nijman et al., 1999; Yamamoto et al., 2005; Kiyokawa et al., 2006; Van Kranendonk, 2006; Chen et al., 2009; Wang et al., 2012), also suggest a subseafloor hydrothermal origin. In this case, the low-temperature, silica-rich hydrothermal fluids were vented diffusively into the earliest Cambrian bottom seawater, promoting widespread deposition of the overlying bedded cherts in the basal Cambrian Yurtus Formation. In the subseafloor mixing zone, downward-circulating oxygenated seawaters could have mixed with the ascending hydrothermal fluids with steady fluxes of Fe(II) in the antecedent porous karstified dolostones immediately under the seafloor. Consequently, aqueous microaerophilic conditions were created there, which sustained colonization of the FeOB community and bacterial iron oxidation (Boyd and Scott, 2001; Kennedy et al., 2003c; Emerson and Moyer, 2010).
The silica-rich hydrothermal fluids with relatively high temperatures (40–200°C) and salinity levels (3–7 times of coeval seawater), as revealed by the fluid inclusion data of quartz and barite crystals as documented above (Zhou et al., 2014), seemingly were not habitable for microorganisms. However, these attributes likely represent the temporary pulses of hotter upwelling fluids in which FeOB could have not colonized the veins. In contrast, habitable niches for FeOB may have been created in the vein space during intervals of lower temperature and salinity level free of silica precipitation. Thus, filamentous FeOB might have colonized the subseafloor stockwork-like feeder veins during waning episodes of relatively low-temperature silica-rich hydrothermal fluids, followed by demise and rapid fossilization during waxing of higher-temperature hydrothermal venting (Fig. 5). Toward the end of the hydrothermal activity, the hydrothermal venting would have tended to be weaker such that the FeOB would have thrived to some extent. This scenario is supported by the local enrichment of filamentous microstructures within the microquartz and chalcedonic spherulite in the center of some vein fills (Fig. 2B). These fossilized FeOB thriving in subseafloor low-temperature mixing zones of hydrothermal systems may be analogous to the modern subseafloor filamentous microbial structures in hydrothermal vent fields at Axial Volcano off the coast of Oregon (Kennedy et al., 2003c).

(
5.4. Implications
In recent decades, increasing evidence has been reported on habitat distribution of FeOB in modern oceans (Table 1) and ancient records (Little et al., 2004; Hofmann et al., 2008, and references therein). Diverse chemoautotrophic FeOB with typical filamentous structures are widely found in the marine Si-Fe deposits or iron-rich mats (i.e., low-temperature alteration of shallow subsurface ocean crust), but they mostly dwell in silica- and iron-rich fluids and are permineralized at low-temperature (mainly diffuse) hydrothermal vent fields in mid-ocean ridge and back-arc seafloor spreading centers, volcanic arcs, and seamounts (Table 1). However, the FeOB filaments discovered in the Qigebulake quartz veins of the silica-rich hydrothermal system in the E–C successions in the Aksu area were not associated with traditional hydrothermal vents at oceanic spreading centers or volcanic seamounts; rather, they occurred in relatively stable, passive continental margin shelf environments (Yu et al., 2009). Thus, this type of FeOB found in subseafloor quartz veins of low-temperature hydrothermal systems provides a new analogue and clue with which to explore the geological niches adaptive for FeOB and improve our understanding of the diversity of hydrothermal vent-associated microorganisms (McAllister et al., 2011). Meanwhile, the well-preserved filamentous fossils of FeOB will be helpful in the search for morphological and geochemical biosignatures in ancient Fe-dependent microbial records and our understanding of habitat diversity and early biosphere evolution on Earth and other planets.
6. Conclusion
Abundant filamentous microstructures are present in silica-dominant vein networks in the uppermost dolostones of terminal-Ediacaran Qigebulake Formation, which is overlain by the bedded cherts and black shales of the Lower Cambrian Yurtus Formation, in the Aksu area, northwestern Tarim Basin, China. The silica-dominant vein swarms were considered to be subseafloor remnants of silica-rich hydrothermal vent systems during the earliest Cambrian age. These filaments were permineralized by Fe-(oxyhydr)oxide and completely entombed in the silica (chalcedony, quartz) cements. The identical morphological structures and parameters to those of FeOB living in modern and ancient hydrothermal venting fields point to a biogenic origin of these filaments. Therefore, these Fe-rich filaments are likely the microfossils of FeOB, which thrived in the subseafloor mixing zone of a silica-rich hydrothermal system and were rapidly fossilized in response to waning and waxing of hydrothermal venting, respectively. These newly recognized FeOB microfossils were discovered in the subseafloor hydrothermal systems in a continental shelf environment, far from oceanic spreading centers and volcanic seamounts, which provides a new perspective in exploration of habitat diversity of extremophiles and relevant biological signatures on Earth and potentially beyond.
Footnotes
Acknowledgments
This research is supported by National Science and Technology Special Project of China (grant no. 2011ZX0500803), National Basic Research Program Project (973 Project grant no. 2012CB214802), and National Natural Science Foundation of China (grant nos. 40839907, 41203030, and 41472089). We are grateful to Geoffrey Li, Zhou Zhang, and Dr. Wenfang Wu for useful suggestions and to the anonymous reviewer for constructive comments that have improved the paper greatly. We would like to thank Wenwen Wei for his assistance in the field. We appreciate the technical support in lab work provided by Bojie Wen (Raman spectroscopy) from the Institute of Geology and Geophysics, CAS.
Author Disclosure Statement
No competing financial interests exist.