
Abstract
The delivery of extraterrestrial organics to early Earth provided a potentially important source of carbon and energy for microbial life. Optically active organic compounds of extraterrestrial origin exist in racemic form, yet life on Earth has almost exclusively selected for L- over D-enantiomers of amino acids. Although D-enantiomers of proteinogenic amino acids are known to inhibit aerobic microorganisms, the role of concentrated nonproteinogenic meteoritic D-amino acids on anaerobic metabolisms relevant to early Earth and other anoxic planets such as Mars is unknown. Here, we test the inhibitory effect of D-enantiomers of two nonproteinogenic amino acids common to carbonaceous chondrites, norvaline and α-aminobutyric acid, on microbial iron reduction. Three pure strains (Geobacter bemidjiensis, Geobacter metallireducens, Geopsychrobacter electrodiphilus) and an iron-reducing enrichment culture were grown in the presence of 10 mM D-enantiomers of both amino acids. Further tests were conducted to assess the inhibitory effect of these D-amino acids at 1 and 0.1 mM. The presence of 10 mM D-norvaline and D-α-aminobutyric acid inhibited microbial iron reduction by all pure strains and the enrichment. G. bemidjiensis was not inhibited by either amino acid at 0.1 mM, but D-α-aminobutyric acid still inhibited at 1 mM. Calculations using published meteorite accumulation rates to the martian surface indicate D-α-aminobutyric acid may have reached inhibitory concentrations in little over 1000 years during peak infall. These data show that, on a young anoxic planet, the use of one enantiomer over another may render the nonbiological enantiomer an environmental toxin. Processes that generate racemic amino acids in the environment, such as meteoritic infall or impact synthesis, would have been toxic processes and could have been a selection pressure for the evolution of early racemases. Key Words: Microbial iron reduction—Amino acids—Toxicity. Astrobiology 15, 238–246.
1. Introduction
L
In addition to organics common on Earth, there are several compounds in meteorites that are rare or nonexistent in the terrestrial biosphere. Of particular interest is the amino acid class of organic compounds, most of which are nonproteinogenic. For example, the nonproteinogenic amino acids, norvaline (Nor) and α-aminobutyric acid (AABA), have been detected in several carbonaceous chondrites, with Nor occurring in the Murchison (Kvenvolden et al., 1971; Ponnamperuma, 1972; Lawless, 1973; Cronin and Pizzarello, 1983) and Almahata Sitta (Glavin et al., 2010) meteorites and AABA detected in the Murchison (Kvenvolden et al., 1971; Ponnamperuma, 1972; Ehrenfreund et al., 2001; Botta and Bada, 2002), Murray (Ehrenfreund et al., 2001; Botta and Bada, 2002), Orgueil, and Ivuna meteorites (Ehrenfreund et al., 2001).
Common terrestrial D-amino acids have been shown to partially or fully inhibit growth of a number of aerobic bacteria, including Lactobacillus arabinosus (Fox et al., 1944), Bacillus subtilis (Trippen et al., 1976), strains of Erwinia (Grula, 1960) and Thiobacillus (Lu et al., 1971), Corynebacterium collunge (Trippen et al., 1976), and Escherichia coli (Davis and Maas, 1949; Rowley, 1953a, 1953b). Conversely, Zhang and Sun (2014) recently demonstrated that common aerobic soil bacteria are capable of consuming D-enantiomers of alanine, aspartic acid, glutamic acid, and leucine at 4 mM concentration, proposing an enzymatic reverse mechanism that not only detoxifies the D-amino acids but renders them a bioavailable source of carbon and nitrogen. They propose that, in doing so, bacteria carry out a global ecosystem service to prevent the buildup of D-amino acids to toxic concentrations. However, the majority of amino acids tested in this and the previously mentioned studies are proteinogenic, and most of the strains that have been subjected to these tests operate an aerobic and heterotrophic mode of metabolism. Therefore, it remains to be seen whether D-forms of nonproteinogenic amino acids of extraterrestrial origin inhibit redox-driven anaerobic microbial metabolisms, such as microbial iron reduction, which would be more relevant for an anoxic early Earth, when impact fluxes were higher than today, and anoxic worlds, such as Mars.
Here, we demonstrate the inhibitory effect of two chiral nonproteinogenic amino acids that are common to meteorites, Nor and AABA, on the ability of a number of iron-reducing microorganisms to carry out microbial iron reduction (MIR).
2. Materials and Methods
2.1. Microorganisms
To investigate the effects of D-amino acids, we used iron-reducing microorganisms. These organisms were chosen because they are anaerobic and iron reduction is a widespread and possibly ancient metabolic capability (Vargas et al., 1998; Lovley, 2013). Actively growing cultures of Geobacter bemidjiensis (Nevin et al., 2005; DSM-16622), Geopsychrobacter electrodiphilus (Holmes et al., 2004; DSM-16401), and Geobacter metallireducens (Lovley et al., 1993; DSM No-7210) were obtained from the Deutsche Sammlung von Mikroorganismen und Zelkulturen GmbH (DSMZ; Braunschweig, Germany). Although isolates allow for controlled data, natural communities that involve interactions between a variety of microorganisms have not been investigated before. As such, a MIR enrichment culture was also tested, which was established with waterlogged sediments from the bed of an iron-rich stream near Borgarnes, Iceland (64°29′N, 21°31′W).
2.2. Cultivation and growth conditions
Strict anaerobic culturing techniques were used throughout (Miller and Wolin, 1974). G. bemidjiensis and G. metallireducens were grown on freshwater bicarbonate-buffered medium, containing (per L) 2.5 g NaHCO3, 0.25 g NH4Cl, 0.06 g NaH2PO4, 0.1 g KCl, 10 mL vitamin solution, and 10 mL trace elements solution. The vitamin solution contained (per L) 2.0 mg biotin, 2.0 mg folic acid, 10.0 mg pyridoxine-HCl, 5.0 mg riboflavin, 5.0 mg thiamine, 5.0 mg p-aminobenzoic acid, and 5.0 mg thioctic acid. The trace elements solution contained (per L) 1.5 g NTA, 3.0 g MgSO4, 0.5 g MnSO4·H2O, 1.0 g NaCl, 0.1 g FeSO4·7H2O, 0.1 g CaCl2·2H2O, 0.1 g CoCl2·6H2O, 0.13 g ZnCl, 0.01 g CuSO4·5H2O, 0.01 g AlK(SO4)2·12H2O, 0.01 g H3BO3, 25.0 mg NaMoO4, 25.0 mg Na2WO4·2H2O, and 24.0 mg NiCl2·6H2O. In all experiments, Fe(III)-citrate (50 mM) was added from sterile anoxic stock as the electron acceptor, and in inhibition experiments acetate (10 mM) was supplied as the electron donor. The final pH of the growth medium was adjusted to 6.8–7.0, and both strains were incubated in the dark at 30°C.
Geopsychrobacter electrodiphilus was grown on a basal medium containing (per L) 0.6 g KH2PO4, 0.3 g NH4Cl, 0.5 g MgSO4·7H2O, 0.1 g CaCl2·2H2O, and 20 g NaCl. After autoclaving, this medium was supplemented with (per 10 mL) 0.35 mL 10% (w/v) NaHCO3, 0.01 mL SL-10 trace element solution, 0.01 mL selenite-tungstate solution, and 0.01 mL vitamin solution (as above). All supplements were made from sterile anoxic stocks with sterile needles and syringes that were flushed with N2. The SL-10 trace element solution contained (per L) 10 mL 7.7 M HCl, 1.5 g FeCl2·4H2O, 0.07 g ZnCl2, 0.1 g MnCl2·4H2O, 6.0 mg H3BO3, 0.19 g CoCl2·6H2O, 2.0 mg CuCl2·2H2O, 24.0 mg NiCl2·6H2O, 36.0 mg Na2MoO4·2H2O, and 0.5 g Na2-EDTA. The solution was adjusted to pH 7.0. The selenite-tungsten solution contained (per L) 0.5 g NaOH, 3.0 mg Na2SeO3·5H2O, and 4.0 mg Na2WO4·2H2O. This strain was used in inhibition tests only, in which the electron donor and acceptor supplied were acetate (10 mM) and Fe(III)-citrate (50 mM), respectively. The strain was incubated in the dark at room temperature.
The MIR enrichment culture was initiated in an anaerobic chamber by adding sediment to a sterile serum vial containing sterile anoxic freshwater basal medium at pH 6.8–7.0. The interior of the anaerobic chamber was cleaned with ethanol prior to use, as were the gloves used to transfer sediment to the serum vial. The basal medium contained (in g L−1 deionized water) NaHCO3 (2.5), NH4Cl (0.25), NaH2PO4·H2O (0.06), KCl (0.2), and vitamins (10 mL) and trace elements (10 mL). The vitamin mixture contained (in mg L−1 deionized water) biotin (2.0), folic acid (2.0), pyridoxine-hydrochloride (10.0), riboflavin (5.0), thiamine (5.0), nicotinic acid (5.0), pantothenic acid (5.0), vitamin B-12 (0.1), p-aminobenzoic acid (5.0), and thioctic acid (5.0). The trace elements mixture contained (in g L−1 deionized water) nitrilotriacetic acid (1.5), MgSO4 (3.0), MnSO4·H2O (0.5), NaCl (1.0), FeSO4·7H2O (0.1), CaCl2·2H2O (0.1), CoCl2·6H2O (0.1), ZnCl2 (0.13), CuSO4·5H2O (0.01), AlK(SO4)2·12H2O (0.01), H3BO3 (0.01), NaMoO4 (0.025), NiCl2·6H2O (0.024), and NaWO4·2H2O (0.025). Vitamin and trace elements mixes were stored at 4°C in the dark. The headspace was flushed with N2/CO2 (80:20) through a 0.2 μm syringe filter. The enrichment was amended with acetate (10 mM) and lactate (10 mM) as the electron donors and poorly crystalline Fe(III) oxide (approximately 100 mM) as the terminal electron acceptor. Iron oxide was synthesized as previously described (Lovley and Phillips, 1986b) and confirmed with X-ray diffraction. FeCl2 (1.3 mM) was added to the medium as a mild reducing agent. The enrichment culture was incubated in the dark at room temperature and periodically measured for production of ferrous iron as a proxy for MIR with the ferrozine assay (Stookey, 1970; Lovley and Phillips, 1986a, 1986b). A second-generation culture was initiated by transferring a 10% v/v inoculum to fresh medium upon detection of complete reduction of ferric iron.
2.3. Phylogenetic analysis
To confirm the existence of iron-reducing microorganisms in the MIR enrichment culture, DNA was extracted and sequenced for bacterial 16S ribosomal RNA (rRNA) by using the Roche 454 Pyrosequencing platform (Research and Testing Laboratories, Austin, TX, USA). DNA was extracted from a 500 μL subsample with the FastDNA SPIN Kit for Soil (MP Biomedicals, Illkirch, France). Initial attempts were unsuccessful, and DNA was only retrieved after incorporating the protocol of Direito and others (2012) to reduce adsorption of DNA onto clay matrices. Specifically, the FastDNA SPIN kit phosphate buffer was substituted for an equal volume of 1 M sodium phosphate buffer in 15% molecular-grade ethanol and corrected to pH 8.0. Samples were incubated in a heating block at 80°C for 40 min following the lysing step in the FastDNA SPIN kit protocol. All subsequent processing was the same as in the manufacturer's protocol.
Following 16S rRNA PCR amplification and pyrosequencing, the resulting sequence reads were subject to analysis and processing with the Research and Testing Laboratories pipeline. This analysis is split into two stages. Firstly quality trimming, denoizing, and chimera checking, and secondly phylogenetic analysis. The methods employed in the first stage are described elsewhere (Edgar, 2010, 2011; Edgar et al., 2011). The second stage involves clustering sequences into operational taxonomic units and use of the distributed .NET algorithm that utilizes BLASTN+(KrakenBLAST) to query the seed sequence of each cluster against a database of high-quality sequences compiled from the National Center for Biotechnology Information (NCBI). Sequences are then resolved to taxonomic levels based on the BLASTN+derived sequence identity percentage. Species-level identification is assigned to sequences with greater than 97% identity, genus level is assigned to between 95% and 97% identity, family level to between 90% and 95% identity, order level to between 85% and 90% identity, class level to between 80% and 85% identity, and phylum level to between 77% and 80% identity. All sequences characterized by a match of less than 77% are discarded. Data reported in this study were taken from the Counts file, and sequences with equal similarity to more than one taxa omitted.
2.4. Experimental setup
To assess inhibition, D- and L-Nor and AABA were added to media already containing an electron donor and acceptor, at a final concentration of 10 mM. Additional tests assessing concentration sensitivity were conducted at 1 and 0.1 mM with G. bemidjiensis. The electron donor and acceptor supplied to pure strains were Fe(III)-citrate (50 mM) and acetate (10 mM), respectively. The enrichment culture was supplied with an equal mix of acetate and lactate (10 mM each) serving as the electron donors and synthetic ferrihydrite as the terminal electron acceptor (c. 20 mM). All amino acid supplements were carried out from sterile anoxic stocks. For pure strains, replicates were inoculated with 250–900 μL of the appropriate culture in late log-phase. Each replicate of the enrichment experiment was inoculated with 1 mL second-generation active culture that had fully reduced the ferric iron in the medium. All inhibition tests were inoculated with 250–900 μL of the appropriate culture in the late log-phase of growth.
All experiments were conducted in triplicate and included positive controls [known electron donor (or donors) and acceptor supplied] and blanks (electron donor and acceptor, no culture).
2.5. Analytical methods
Microbial iron reduction was determined by measuring the concentration of HCl-extractable ferrous iron with time as previously described (Lovley and Phillips, 1986a, 1986b). Briefly, 100 μL samples were anaerobically removed and digested in 0.5 N HCl for 1 h to dissolve Fe(II) minerals, and Fe(II) was quantified with the ferrozine assay (Stookey, 1970). Cell numbers were monitored in the 10 mM inhibition tests with G. bemidjiensis by epifluorescence microscopy. Samples (100 μL) were anaerobically removed at the start and end of the experiments with a syringe and needle and fixed with glutaraldehyde (2.5% final concentration). Mean changes in ferrous iron reduction and cell numbers between inhibition tests compared with corresponding positive controls were assessed for statistical significance by using a Student t test (2-tailed, type 2, critical vale of 0.05), and corresponding p values discussed below.
3. Results
3.1. Phylogenetic diversity of the MIR enrichment culture
Family-level taxonomic identification of sequences was retrieved from 16S rRNA sequencing of the enrichment culture. More than half the sequences identified at this level belong to the iron-reducing Geobacteraceae family (62.1%), with the majority of remaining sequences belonging to Holophagaceae (34.7%). All sequences assigned to Holophagaceae were identified at genus level as Geothrix species. Species-level identification was resolved for iron-reducing species Geobacter pelophilus (41.3%), Geobacter psychrophilus (2.4%), and Geothrix fermentans (1.9%).
3.2. Inhibition of MIR by D-amino acids
The effect of adding D- and L-forms of both chiral amino acids on MIR by Geobacter bemidjiensis is shown in Fig. 1. Specifically, the addition of 10 mM D-Nor to acetate- and ferric iron–containing media led to the production of 1.77±1.52 mM Fe2+ after 15 days, compared with 50.30±3.85 mM production in the positive control. This difference is statistically significant (p value<0.05). Addition of 10 mM D-AABA led to the production of 1.99±1.13 mM Fe2+, a similarly significant difference to the positive control (p value<0.05). The presence of 10 mM L-Nor resulted in a change of 14.68±4.54 mM Fe2+, significantly different to the mean change in Fe2+ concentration in the positive control (p value <0.05). Addition of 10 mM L-AABA did not result in a significantly different mean change in Fe2+ concentration (p value >0.05). The effect of adding the racemic form of Nor or AABA was not assessed for this strain.
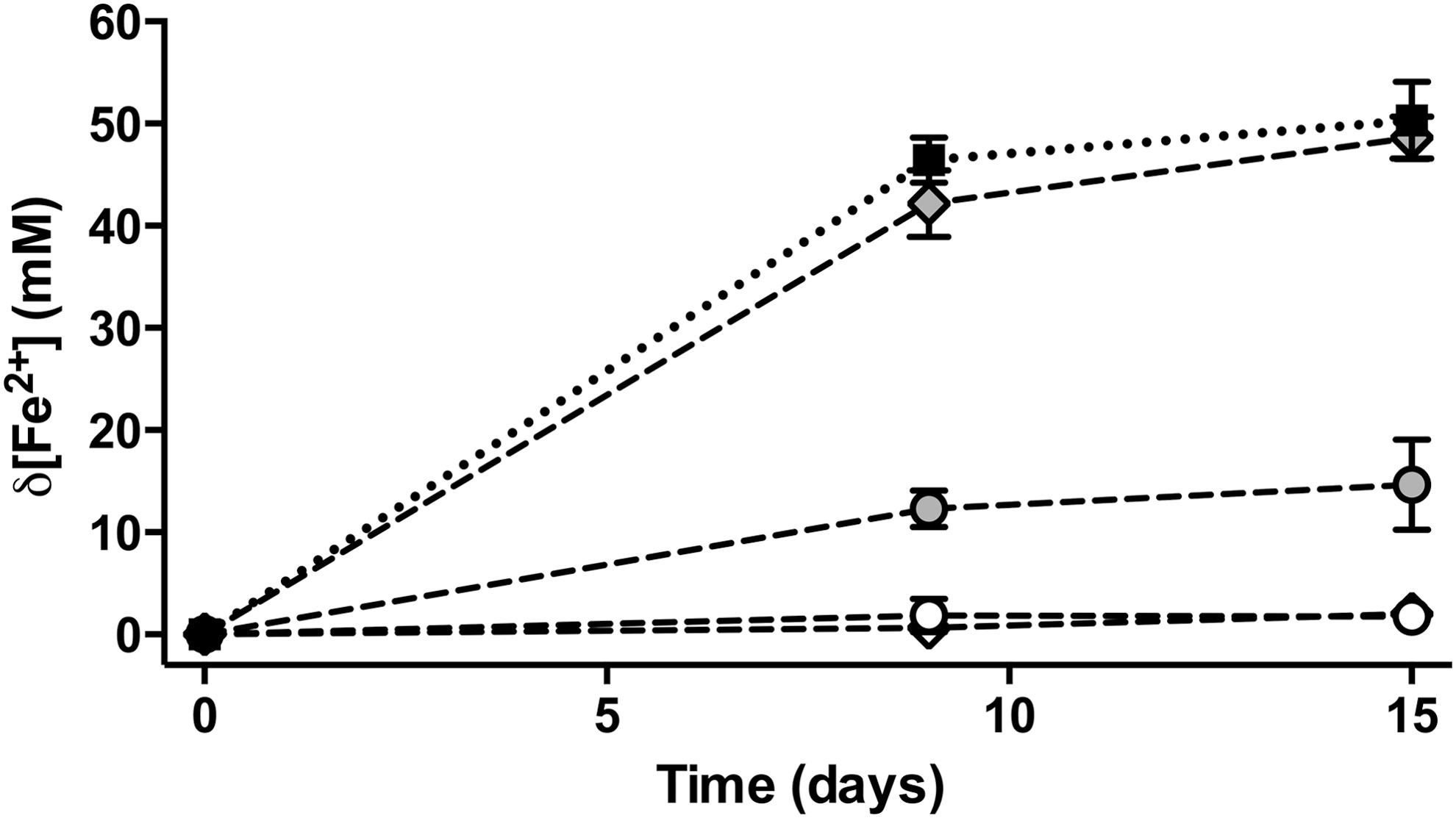
Inhibition of MIR by D-enantiomers. Data represent mean change in Fe2+ concentration (mM) with time (days) when G. bemidjiensis is grown on acetate (10 mM) and Fe(III)-citrate (50 mM) spiked with chiral amino acids (dashed lines). D-Nor (empty circles), L-Nor (shaded circles), D-AABA (empty diamonds), and L-AABA (shaded diamonds) were added at a final concentration of 10 mM. Dotted black line and black squares represent positive control (no amino acid added). Error bars represent standard deviation of triplicate measurements.
The effect of adding 10 mM D- and L-forms of the chiral amino acids on MIR by Geobacter metallireducens is shown in Fig. 2a. Addition of D-Nor and D-AABA gave rise to changes of 4.60±1.49 mM Fe2+ and 1.12±0.53 mM Fe2+, respectively, compared to an increase of 36.14±5.50 mM Fe2+ in the positive control over the course of 5 days. Both D-inhibited means are significant compared with that of the positive control (p values <0.05). No inhibition was evident in the presence of 10 mM of the L-form of either, compared to the positive control (L-Nor: 38.18±4.03 mM Fe2+, p value >0.05; L-AABA: 41.01±4.34 mM Fe2+, p value >0.05).
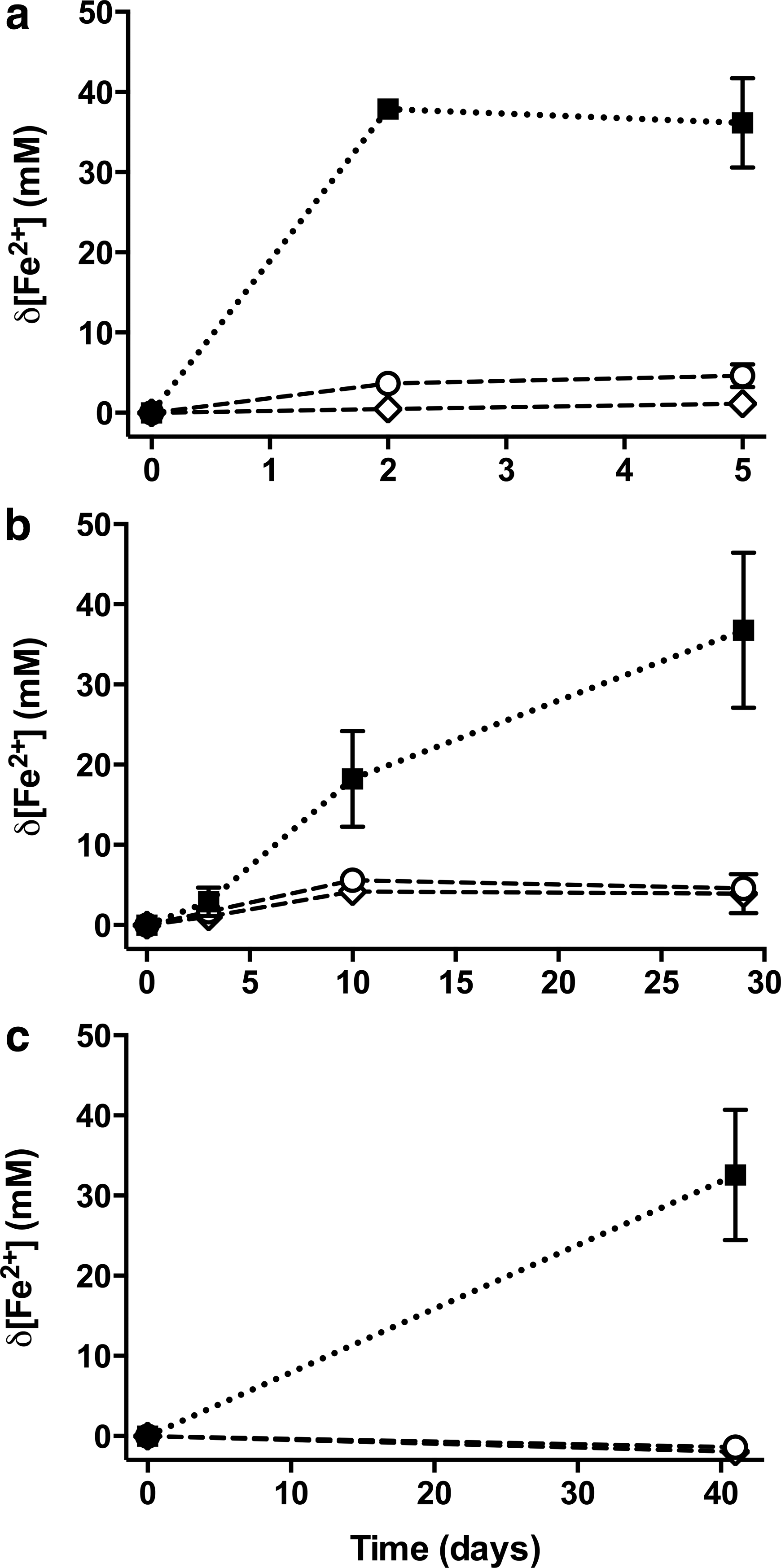
Inhibition of MIR by other iron-reducing microorganisms. Data represent mean change in Fe2+ concentration (mM) with time (days) when cultures of G. metallireducens (
In the absence of any amino acids, Geopsychrobacter electrodiphilus reduced 36.76±9.82 mM Fe2+ over the course of 29 days. As shown in Fig. 2b, MIR was significantly lower in the presence of 10 mM of either D-Nor (4.57±1.40 mM Fe2+, p value <0.05) or D-AABA (3.92±2.50 mM Fe2+, p value <0.05). No inhibition was evident in the presence of 10 mM L-Nor (30.24±9.07 mM Fe2+, p value >0.05) or 10 mM L-AABA (39.24±9.99 mM Fe2+, p value >0.05).
With the exception of L-Nor, the addition of 10 mM of all forms of amino acids inhibited MIR by the enrichment culture over the course of 41 days (Fig. 2c). Compared with an increase of 32.56±7.90 mM Fe2+ in the positive control, addition of 10 mM of D-Nor and D-AABA led to significantly lower Fe2+ production, with overall changes in concentration of −1.37±0.87 mM (p value <0.05) and −1.97±1.21 mM (p value <0.05), respectively. The presence of 10 mM L-AABA also led to significantly lower Fe2+ production, albeit to a lesser extent (5.27±4.11 mM Fe2+, p value <0.05), whereas the presence of 10 mM L-Nor did not significantly change Fe2+ concentration compared to the positive control (13.90±25.03 mM Fe2+, p value >0.05). However, an increase in Fe2+ concentration was detected in only one of the three replicates ([1] 43.58 mM compared with [2] −0.67 mM and [3] −1.22 mM).
The normalized effect of adding D- and L-forms of Nor and AABA to the different iron-reducing cultures is shown in Fig. 3.

Normalized effect of adding D- and L-Norv and AABA on MIR by all strains. Media contained acetate (and lactate for the enrichment culture) and Fe3+ as electron donor and acceptor, respectively. Results are expressed as the percentage of Fe2+ produced over the course of experiments compared to corresponding positive control (no amino acid added) for each strain, where 100% represents no inhibition. Error bars represent standard deviation of triplicate measurements.
3.3. Inhibition of MIR at different concentrations
Inhibition of MIR on Geobacter bemidjiensis was tested at the lower concentrations of 1 and 0.1 mM, the normalized results of which are summarized in Fig. 4. Compared with the significant reduction in Fe2+ production in the presence of 10 mM discussed above, addition of D-Nor did not significantly affect the change in Fe2+ concentration at a concentration of 1 mM (p value >0.05) or 0.1 mM (p value >0.05). In contrast, the presence of 1 mM D-AABA did result in a significantly lower production of Fe2+, resulting in a change of 2.08±0.94 mM compared with 35.17±2.98 mM in the positive control (p value <0.05) over the course of 17 days. The presence of 0.1 mM D-AABA did not lead to significantly different changes in Fe2+ compared with the positive control (p value >0.05).

Normalized effect of adding D- and L-Norv and AABA at different concentrations on MIR by G. bemidjiensis. Final concentrations of amino acids were 10, 1, and 0.1 mM. Results are expressed as the percentage of Fe2+ produced over the course of the experiment compared to the corresponding positive control (no amino acid added), where 100% indicates no inhibition. Error bars represent standard deviation of triplicate measurements.
3.4. Inhibition of microbial growth
To assess whether inhibition affected microbial growth as well as MIR, cells were enumerated in the 10 mM inhibition test with Geobacter bemidjiensis. The average number of cells±standard deviation across all replicates at the start of the experiment was 1.14×107±5.09×106 cells mL−1 sample. In the positive control, the average number of cells increased from 1.48×107±3.47×106 to 3.45×107±6.91×106 cells mL−1, the difference between the two numbers being statistically significant (p value <0.05). The average number of cells decreased in the presence of D-Nor from 1.09×107±1.89×106 to 3.68×106±1.47×106 cells mL−1, representing a significant overall change of −7.24×106 ±2.39×106 cells mL−1 over time (p value <0.05). The number of cells in tests with added L-Nor also significantly decreased by 9.44×106±3.41×106 cells mL−1 over the course of the experiment, from 1.37×107±2.50×106 to 4.25×106±2.32×106 cells mL−1 (p value <0.05). A similarly significant decrease in cell numbers was measured in the presence of D-AABA, with a reduction of 5.33×107±1.34×106 cells mL−1 between the start and the end (p value <0.05). In each case, the change in cell numbers over the course of the experiment was statistically significant when compared with that of the positive control (p values <0.05). In contrast, there was a significant increase in the number of cells from 8.80×106±4.41×105 to 6.33×107 cells mL−1 with the addition of L-AABA (p value <0.05), an overall change of 5.46×107±2.93×107 cells mL−1 that is not significantly different to the change measured in the positive control (p value >0.05).
These results indicate that the presence of D-Nor, L-Nor, and D-AABA inhibited microbial growth compared with the positive control, whereas addition of L-2-aminobutyric acid did not.
4. Discussion
During the early history of Earth, meteoritic infall and endogenous synthesis of organic compounds could have provided a potentially important source of carbon and energy for microorganisms. However, these sources are generally racemic or contain substantial proportions of both enantiomers of optically active compounds. In this study, we sought to investigate the effect of nonproteinogenic D-amino acids on anaerobic microorganisms. We were motivated to investigate whether, once life takes an evolutionary branch point of predominantly using one enantiomer or the other, the unused enantiomer is a passive compound in the environment, or whether it plays a role in biological interactions. The above results demonstrate that two D-amino acids inhibit dissimilatory iron reduction and growth of three pure cultures of iron-reducing deltaproteobacteria and one enrichment culture containing iron-reducing deltaproteobacteria.
Inhibitory concentrations measured in previous studies are much lower than those found to inhibit MIR in this study. Only two published studies have tested microbial inhibition by Nor and AABA. The first (Rowley, 1953a) involved testing the effects of adding racemic mixtures of both amino acids to 100 strains of the Gram-negative bacterium Escherichia coli, and demonstrated that 75 were inhibited by DL-Nor and 7 by DL-AABA, though the final concentrations of each were not reported. A follow-up study (Rowley, 1953b) extended these tests to 365 strains of E. coli and reported inhibition by DL-Nor (8.5 μM) in 261 strains and by DL-AABA (9.9 μM) in 4.
Our results are broadly consistent with other studies that have reported the inhibition of microbial growth by D-amino acids, though these are limited to proteinogenic amino acids. Gladstone (1939) reported the failure of a strain of the Gram-positive bacterium Bacillus to grow in the presence of the chiral amino acids valine (8.5 μM) and leucine (9.9 μM). Shortly after this, a study aimed at elucidating the effectiveness of the antibiotic gramidicin recognized the inhibitory effects of D-leucine on the Gram-positive bacterium Lactobacillus arabinosus (Fox et al., 1944). Davis and Maas (1949) also reported the inhibitory effect of DL-serine on E. coli, where no inhibition was evident upon addition of the L-isomer only. The aforementioned studies conducted by Rowley (1953a, 1953b) found several amino acids to inhibit different strains of the same species to varying degrees. In both studies, norleucine and norvaline inhibited the majority of strains. Other studies reported inhibition by D-isomers on E. coli (Kuhn and Somerville, 1971; Caparrós et al., 1992), as well as strains of Erwinia (Grula, 1960), Brucella (Yaw and Kakavas, 1952), Rhodospirillium (Coleman, 1959), Alcaligenes (Lark and Lark, 1959), Pseudomonas (Eisenstadt et al., 1959), and Coryneacterium (Trippen et al., 1976). A more recent study reported the prevention of biofilm formation by Staphylococcus aureus in the presence of D-amino acids (Hochbaum et al., 2011).
A number of studies found inhibition to be either short-lived (Davis and Maas, 1949; Rowley, 1953b; Coleman, 1959) or completely preventable through the addition of other, seemingly antagonistic, amino acids (Davis and Maas, 1949; Rowley, 1953a, 1953b; Lu et al., 1971). The transient nature of inhibition in the former studies was not observed in these experiments, where the presence of the D-isomer resulted in total inhibition of MIR (Fig. 3) or no inhibition at lower concentrations (Fig. 4). However, the antagonistic effects of adding different amino acids were not assessed (beyond adding the racemic form); hence it is possible that these MIR cultures would be affected differently in the presence of amino acids other than those shown to inhibit in isolation in this study.
The results show that the evolutionary choice to use one optically active form of a molecule over another had the potential to render the other an environmental toxin to early anaerobic organisms—in the case of amino acids, even nonproteinogenic D-amino acids. Furthermore, processes that generate racemic mixtures of amino acids, such as impact events (Martins et al., 2013), would not only produce molecules of benefit to life but would also be the producers of environmentally toxic biological building block molecules. Such toxins may even have contributed to the evolution of racemases.
4.1. Mechanisms of inhibition
A decrease in cell numbers of G. bemidjiensis over the course of the experiment in the presence of inhibitory amino acids was observed, suggesting cell death.
A number of mechanisms have been proposed to explain the inhibitory effects of D-amino acids. Early speculation attributed inhibition to interference with “proteosynthetic enzymes” (Kobayashi et al., 1948), with hydrolysis of peptide bonds (Yaw and Kakavas, 1952), or with growth factors in the medium (Lark and Lark, 1959). However, the prevailing theory is that D-amino acids interfere with the production of peptidoglycan, the major component of bacterial cell walls that is known to contain D-alanine (Izaki et al., 1986). Evidence to support this hypothesis was presented by Trippen et al. (1976), who demonstrated that D-serine and other D-amino acids replaced D-alanine in nucleotide-activated peptidoglycan precursors, which led to less efficient incorporation into peptidoglycan. Decreased cross-linking within peptidoglycan was also apparent (Trippen et al., 1976). Caparrós and others (1992) also reported the incorporation of D-amino acids, including D-AABA tested in this study, into peptidoglycan by E. coli, and found concentrations above 45 mM to be lethal, though incorporation of D-AABA was observed in the presence of 20 mM. Their results echoed those of Trippen et al. (1976) in that cross-linking was found to be reduced, leading them to propose that such effects may lead to less production of peptidoglycan per cell. Interestingly, β-aminobutyric acid was also assessed but was not found to be incorporated into peptidoglycan. In fact, amino acids lacking an alpha-amino group, or achiral and diamino acids, were not found to be incorporated into macromolecular peptidoglycan (Caparrós et al., 1992), suggesting that inhibition is only caused by amino acids with alpha-amino groups, like the alanine that is integral to the formation of peptidoglycan in the first instance, in addition to the chiral amino acids tested in this study. It therefore seems probable that interference with peptidoglycan formation is the cause of MIR inhibition observed in this study.
This hypothesis is further supported by the known presence of AABA in the lantibiotic “marsacidin” (Sass et al., 2008). Lantibiotics are naturally produced by, and act against, other Gram-positive bacteria, and are of two types. Type A lantibiotics are long flexible molecules that rapidly kill through the formations of pores, whereas Type B lantibiotics are globular in structure and inhibit peptidoglycan synthesis (Smith and Hillman, 2008). Marsacidin, a Type B lantibiotic, is ribosomally produced by Bacillus spp. and acts to complex the sugar phosphate head group of peptidoglycan precursor lipids, resulting in the inhibition of peptidoglycan biosynthesis (Sass et al., 2008). This strongly suggests that the inhibitory action of AABA reported here and elsewhere is related to its role in this peptidoglycan-targeted lantibiotic.
Inhibition of MIR by Geobacter bemidjiensis was not observed in the presence of 1 mM D-Nor or 0.1 mM D-AABA, indicating that a threshold exists at which interference of peptidoglycan biosynthesis ceases. Recently, Zhang and Sun (2014) reported that soil microorganisms were capable of consuming D- and L-isomers of the proteinogenic amino acids alanine, aspartic acid, and glutamic acid at equal or near-equal rates. They attribute this observation to the reverse racemization of D-isomers into L, a phenomenon they propose all bacteria are capable of. They observed that utilization of D-glutamic acid, D-aspartic acid, and D-leucine was prevented upon addition of rifampicin, which acts to inhibit RNA synthesis, while consumption of corresponding L-isomers and D-alanine continued unaffected. This led them to conclude that RNA synthesis and gene expression were essential to the bacteria utilizing D-enantiomers, and supports their theory that enzymatic conversion of D- to L-isomers was responsible for the observed uptake (Zhang and Sun, 2014). The final concentration of amino acids they added to their experiments was 4 mM, which lies within the range of concentrations tested on G. bemidjiensis in this study. If all bacteria are indeed capable of the enzymatic conversion of D- to L-isomers, the results of this study indicate that such ability is concentration-dependent. For instance, at concentrations below the threshold at which they cause irreversible damage through interference with peptidoglycan formation (between 10 and 1 mM for D-Nor and between 1 and 0.1 mM for D-AABA), D-isomers are enzymatically converted to the non-inhibitory L-isomers, which in the case of nonproteinogenic amino acids then serve as a source of cellular nitrogen and carbon.
The observation reported by Caparrós and others (1992) that only alpha-amino acids were found to inhibit peptidoglycan synthesis is particularly pertinent with respect to the exogenous delivery of organics to planetary surfaces. Martins and colleagues (2013) demonstrated the shock synthesis of several alpha-amino acids solely through high-velocity impact of an ice mixture containing amino acid precursors, common to cometary material. Among the amino acids formed were racemic mixtures of the two chiral amino acids tested here, norvaline and aminobutyric acid. This suggests that the impact of meteoritic and cometary material to the surface of early Earth and other planetary bodies contributed to the concentration of inhibitory amino acids.
4.2. Mars—a theoretical example
The planet Mars is anoxic and could have hosted habitable conditions. Although organic compounds have not yet unambiguously been detected on Mars, they are nonetheless expected to reside in the subsurface (Kanavarioti and Mancinelli, 1990). By using estimated rates of the accretion of meteoritic material on Mars, and concentrations of D-AABA as measured in a number of carbonaceous chondrites, a first-order estimation of the concentrations of this compound and its potential to inhibit MIR can be reached.
Flynn (1996) calculated the present-day accretion rate of meteoritic material to the surface of Mars with organic matter intact as 2.4×106 kg per year. If one assumes this material is evenly distributed across the surface of Mars, this equates to 1.7×10−5 g m−2. Ehrenfreund and others (2001) detected 13±11 ppb and 914±189 ppb AABA in Orgueil and Murchison, respectively. The concentration of AABA reported for Ivuna and Murray was 12±7 ppb and 463±68 ppb, respectively. By using the lower-bound concentration reported in Orgueil (2 ppb) and the higher bound of that measured in Murchison (1103 ppb), and assuming that all organic matter in the meteoritic material accounted for in the accretion rate resembles that of these carbonaceous chondrites, the concentration of the D-isomer of AABA that accumulates on the surface of Mars each year is in the range of 1.66×10−8 to 9.14×10−6 μg g−1 bulk carbonaceous chondrite. Assuming a rock-water ratio of 1:1 (1 g meteorite in contact with 1 mL water), and complete disintegration of meteoritic material on impact (allowing exposure to the organic component within), it would take at least 11.3 million years and up to several billion years for D-AABA at any given point on Mars to accumulate to the inhibitory concentration of 1 mM found in this study. The lack of exposure of organic matter and the presence of large volumes of water would substantially dilute these concentrations.
However, the aforementioned accumulation rate of 2.4×106 kg per year is thought to be a lower limit, as the flux of meteoritic material to the surface of Mars during the first billion years of Solar System history is estimated to have been 10,000 times that of the present day (Flynn, 1996). During this period of heightened meteoritic infall, it could have taken little more than 1000 years for the accumulation of D-AABA to reach inhibitory concentrations, assuming the higher concentrations detected in Murchison (10,000 years gives rise to 0.91 mg D-AABA per square meter; 8.86 mM in a 1:1 rock-to-water-ratio scenario).
The lack of conclusive detection of organics on the surface of Mars, coupled to the known presence of strong oxidizing agents such as perchlorate salts (Hecht et al., 2009), renders these estimates uncertain. Some studies have shown inhibitory effects to be completely prevented in the presence of antagonistic compounds; these findings place additional uncertainty on the significance of this inhibition. However, these results serve to highlight that amino acids, the building blocks of proteins in all known life on Earth, may in high enough concentrations restrict habitability.
5. Conclusions
We have shown here that D-amino acids can inhibit iron-reducing deltaproteobacteria at concentrations that could plausibly exist from concentrated organic material delivered by meteoritic infall. Other processes that produce racemic mixtures of amino acids, such as endogenous synthesis, might also lead to accumulations of these compounds in the environment. These data show that, when life chose one enantiomer over another in building biological macromolecules, the unused optically active compound was not merely a passive organic compound, but it became a toxin at concentrations relevant to the early history of Earth and Mars. The prevailing view that a greater stock of amino acids, the building blocks of proteins essential to life as we know it, increases the chances that life will prosper on planetary bodies beyond Earth is therefore too simplistic.
Footnotes
Acknowledgments
S.L.N. was supported by a PhD studentship from the STFC/UK Space Agency under the Aurora Programme.
Author Disclosure Statement
No competing financial interests exist.