
Abstract
The low-temperature alteration of submarine volcanic glasses has been argued to involve the activity of microorganisms, and analogous fluid-rock-microbial-mediated alteration has also been postulated on Mars. However, establishing the extent to which microbes are involved in volcanic glass alteration has proven to be difficult, and the reliability of resulting textural biosignatures is debated, particularly in the early rock record. We therefore propose a hierarchical scheme to evaluate the biogenicity of candidate textural biosignatures found in altered terrestrial and extraterrestrial basaltic glasses and serpentinized ultramafic rocks.
The hierarchical scheme is formulated to give increasing confidence of a biogenic origin and involves (i) investigation of the textural context and syngenicity of the candidate biosignature; (ii) characterization of the morphology and size range of the microtextures; (iii) mapping of the geological and physicochemical variables controlling the occurrence and preservation of the microtextures; (iv) in situ investigation of chemical signatures that are syngenetic to the microtexture; and (v) identification of growth patterns suggestive of biological behavior and redox variations in the host minerals. The scheme results in five categories of candidate biosignature as follows: Category 1 indicates preservation of very weak evidence for biogenicity, Categories 2 through 4 indicate evidence for increasing confidence of a biogenic origin, and Category 5 indicates that biogenic origin is most likely.
We apply this hierarchical approach to examine the evidence for a biogenic origin of several examples, including candidate bacterial encrustations in altered pillow lavas, granular and tubular microtextures in volcanic glass from the subseafloor and a Phanerozoic ophiolite, mineralized microtextures in Archean metavolcanic glass, and alteration textures in olivines of the martian meteorite Yamato 000593. The aim of this hierarchical approach is to provide a framework for identifying robust biosignatures of microbial life in the altered oceanic crust on Earth, and in extraterrestrial altered mafic-ultramafic rocks, particularly on Mars. Key Words: Bioalteration—Volcanic glass—Hierarchical biogenicity criteria. Astrobiology 15, 901–921.
1. Introduction—The Subseafloor as a Potential Habitat for Life
O
In the late 1930s, scientists first began to suspect that the subseafloor hosted life (ZoBell, 1938). As opposed to seafloor sediments, many decades elapsed before direct evidence of life was reported from subseafloor basaltic-ultramafic environments. Clues have come from the study of ecosystems at hydrothermal vents, also textural evidence in volcanic glasses (Fisk et al., 1998; Furnes et al., 2001a; Thorseth, 2011), but the biogenicity of the latter have been much debated (e.g., McLoughlin et al., 2010; Fisk et al., 2013). Only in recent years have advances in seafloor drilling technologies combined with new methodologies in molecular and genomic sciences provided a growing body of evidence for microbial life in the deep oceanic crust (e.g., reviews by Edwards et al., 2011, and Orcutt et al., 2011). Some of the most persuasive evidence has been obtained from laboratory success in cultivating microorganisms recovered from the subseafloor, combined with genetic and isotopic data obtained from Integrated Ocean Drilling Program (IODP) drill cores (e.g., Lever et al., 2013). However, the search for subsurface life in deep time on Earth and on other planetary bodies cannot rely on recovering nucleic acids, which are unlikely to be preserved intact in the rock record, will have undergone hydrolysis, and are subject to racemization within 106 years (Bada et al., 1999). They are also susceptible to mixing with exogenous organics. Furthermore, recent work has strongly questioned and in some cases rejected the biogenicity of some purported biosignatures in metavolcanic glass (e.g., Lepot et al., 2011; McLoughlin and Grosch, 2014). Consequently, there is a strong need for a systematic approach in the search for terrestrial and extraterrestrial biosignatures that combine high-resolution textural, mineralogical, and (bio)geochemical evidence.
2. Candidate Biosignatures in Altered Oceanic Crust—State of Knowledge
The low-temperature alteration of basaltic glass produces palagonite, a fine-grained mixture of clays with iron-oxyhydroxides and zeolites (Stroncik and Schmincke, 2001). Abiotic alteration processes are argued to produce a planar interface between the alteration products and remaining glass, also “rounding off” angular corners (Fig. 1 Staudigel et al., 2008). In contrast, when the boundary between glass and alteration products becomes ramified, with the development of micron-sized tunnels and granular etch pits, the role of microbes has been invoked (e.g., Fisk et al., 1998; Furnes et al., 2001a; Staudigel et al., 2006). Several lines of evidence have been proposed to support biotic mediation of this alteration process, and these will be summarized below.

A petrographic overview of micro-alteration textures in altered volcanic glass and olivine. (
Some of the first indications of biologically mediated volcanic glass alteration were found on subglacial volcanic breccias from Iceland where bacteria were discovered in, and near, microscopic etch pits on the glass surface, matching the dimensions of the hemispherical pits (Thorseth et al., 1991, 1992). These authors suggested that the microbes locally modified the pH and thereby accelerated glass dissolution (Thorseth et al., 1992). The first observations from the in situ oceanic crust came from Ocean Drilling Program (ODP) Hole 896A from Costa Rica rift, where granular and tubular structures were found at the interface between the fresh and altered glass, with DAPI stains binding to these alteration textures suggesting the presence of nucleic acids (Furnes et al., 1996; Torsvik et al., 1998). The microtextures were found to be concentrated along sites of early fluid infiltration and extend into the fresh glass. Subsequently, abundant granular and tubular microtextures have been reported from a large number of sites across the oceans (e.g., Fig 1d–1f) by many different workers (e.g., Fisk et al., 1998; Furnes et al., 2001a; Walton and Schiffman, 2003); and a wide diversity of microtunnel morphologies is now recognized, including straight, curved, branched, and spiral-shaped (Fisk and McLoughlin, 2013). Granular microtextures (Fig. 1e) are by far the most abundant type and are widely regarded as indicative of biological activity (e.g., Staudigel et al., 2014, and references therein); however, some workers have highlighted spherical and hemispherical alteration fronts in volcanic glass that were interpreted to involve slow, abiotic chemical exchange leading to pit formation (e.g., Fig. 4b, 4d Kruber et al., 2008). Notwithstanding this possibility, such granular and tubular microtextures have been termed bioalteration textures and will be described below.
Another line of textural evidence for subseafloor life comes from candidate bacterial encrustations in submarine glass alteration products (Fig. 1a–1c), particularly palagonite and zeolites (see review by Thorseth, 2011). For example, dredged basalts that are less than 100,000 years old from the Arctic Knipovich and Mohns Ridges contain filamentous, coccoid, ovoid, and rod-shaped microtextures in palagonite found in fractures and vesicles (Thorseth et al., 2001; Kruber et al., 2008). Transmission electron microscopy (TEM) and nanoscale secondary-ion mass spectrometry (nanoSIMS) analysis revealed localized enrichments in C and N within the microtextures and enrichment in Mn in the mineral-encrusted rims, supporting the idea that these are fossilized bacteriomorphs (McLoughlin et al., 2011). Comparable 200,000-year-old samples from the Mid-Atlantic Ridge contain hollow rod-shaped structures (Fig. 1a) that are sometimes associated with S and C enrichments and are argued to be fossilized cells (Cockell et al., 2010). These should be distinguished from mineralized spheres that are filled, not hollow, and can merge laterally to form sheets, which are therefore regarded as abiotic mineral precipitates (e.g., Fig. 1b). In zeolite-filled vesicles, mineral-encrusted spherical and filamentous structures have also been described and interpreted as cryptoendolithic microbes (e.g., Cavalazzi et al., 2011), as have large organic filaments of argued fungal origin (e.g., Schumann et al., 2004), suggesting that amygdales may also be a location for submarine life. In this study, we will also formulate biogenicity criteria tailored to this type of mineral-encrusted bacteriomorph, in addition to the glass-hosted microtextures.
The evidence advanced to support biotic-mediated alteration of volcanic glass is now briefly reviewed, accompanied by key caveats. Culture-independent sequencing studies have shown that the microbial population that inhabits the subseafloor is distinct from that found in both overlying seawater and seafloor sediments and is up to four times more diverse (e.g., Santelli et al., 2008; Mason et al., 2009). Early biological staining efforts revealed that nucleic acids can be found at the interface of fresh and altered glass near tubular and granular microtextures and sometimes within the microtunnels (e.g., Giovannoni et al., 1996; Torsvik et al., 1998; Banerjee and Muehlenbachs, 2003). There remains, however, the possibility that these organics may be unrelated to the tunnel-forming process, and rather derived from temporally and spatially distinct microbial sources that were later adsorbed onto the microtextures. Analysis of disseminated carbonate in pillow basalt rims compared to interiors suggests that biological activity has lowered the <lc grk delta>δ13C of the rims relative to the cores (e.g., Furnes et al., 2001c). These data are, however, only consistent and not diagnostic of biological activity, and being bulk measurements are spatially, only loosely related to the microtextures. Likewise, sulfur isotopes in secondary sulfides of altered oceanic crust are highly suggestive of microbial activity in the volcanics as a whole but spatially not directly tied to the microtextural evidence (e.g., Rouxel et al., 2008; Alford et al., 2011). Lastly, controlled laboratory experiments have established that enhanced, localized dissolution occurs in volcanic glass inoculated with microorganisms, relative to abiotic controls (Staudigel et al., 1995; Thorseth et al., 1995a, 1995b); but crucially, these types of experiments have been unable to produce microtunnels. In summary, there are several lines of evidence that support the hypothesis of biologically mediated alteration of volcanic glass, but this is not universally accepted because it has not been possible to cultivate microorganisms in the laboratory that create microtunnels and granular forms. Thus, alternative abiotic mechanisms that are described and discussed below (Section 3) still need to be borne in mind and tested.
In the lower parts of the oceanic crust in mantle-derived rocks, there is now also growing evidence for microbial activity, in olivine-bearing dunnites in particular. Textural evidence includes microtunnels and galleries in the margins of altered olivines, which contain clay alteration products and in some cases nucleic acids (Fisk et al., 2006). These authors compared the microtextures to those found in volcanic glass and stated that “Biogenicity of the tunnels and galleries has not yet been proven, but on the other hand abiotic mechanisms capable of generating tunnels of similar morphology and size have not been identified.” In such environments, serpentinization reactions caused by alteration of the olivines could potentially support hydrogen-based metabolisms (e.g., McCollom and Seewald, 2013). Thus, high concentrations of organic matter associated with serpentine-hosted hydrogarnets have been argued as evidence for life in deep subseafloor environments (Ménez et al., 2012). There is now also growing support for this idea from phylogenetic studies of gabbroic samples from the lower oceanic crust (e.g., Mason et al., 2010; Orcutt et al., 2011), and strongly fractionated S and C isotopes from serpentinized ultramafics support microbial activity as well (e.g., Delacour et al., 2008; Alt et al., 2013). In this study, we will also propose biogenicity criteria that can also be applied to alteration textures in altered olivines from both terrestrial samples and martian meteorites.
The oldest in situ oceanic crust is at most 170 million years old; thus candidate evidence for the microbial alteration of subseafloor volcanic glass before this time comes from ophiolites and Precambrian greenstone belts. At several locations where pillow lava have escaped strong deformation and experienced greenschist facies or lower grades of metamorphism, filamentous titanite (CaTiSiO4) microtextures have been argued to represent mineralized bioalteration textures (for comprehensive reviews, see Furnes et al., 2008). Examples include the late Ordovician c. 443 Ma Solund-Stavfjord ophiolite of Western Norway (Fliegel et al., 2011); the Paleoproterozoic c. 2.0 Ga (billion-year-old) Pechenga Greenstone belt of northwest Russia, although these examples have been contested (Fliegel et al., 2010a; McLoughlin et al., 2012); the late Archean c. 2.7 Ga Abitibi greenstone belt of Canada (Bridge et al., 2010); and Archean pillow lavas of the Pilbara craton, Western Australia (Banerjee et al., 2007; also Fig. 1i, 1j), and Barberton Greenstone Belt, South Africa (Furnes et al., 2004; Banerjee et al., 2006; Staudigel et al., 2015; also Fig. 5e, 5f), also recently contested (Grosch and McLoughlin 2014, 2015). At these locations, clusters of filamentous and segmented titanite (Fig. 1i, 1j) have been compared to microtunnels from the modern oceanic crust largely on the basis of argued morphological similarities and interpreted as early microbial microtunnels, or trace fossils preserved by the growth of infilling metamorphic titanite (e.g., Furnes et al., 2008; Staudigel et al., 2015). Additional lines of evidence argued to support their biogenicity include carbon isotope fractionations of disseminated carbonate in the pillow lava rims and X-ray element mapping of apparent organic linings to the microtextures; however, these lines of evidence are now strongly contested, as explained below.
3. Contrasting Biotic and Abiotic Models of Volcanic Glass Alteration
A bioalteration model for volcanic glass alteration has been proposed in a series of studies (e.g., Furnes et al., 2001a, 2008; Staudigel et al., 2006, 2008, 2014). In this conceptual model, it is proposed that microbial cells colonize the glass and exude a range of substances (e.g., acids, chelating agents) that dissolve the underlying glass, creating a hemispherical cavity that mirrors the dimensions of the microbial cell, producing a granular etch pit (see Fig. 14 Staudigel et al., 2008; Figs. 2 and 3 Staudigel et al., 2014). Once this first cavity is formed, the cell is argued to divide or move and create a second cavity nearby such that the two cavities coalesce and develop a “sponge-like” network of granular alteration. Dissolution of the glass is envisioned to yield chemical components that may be used as nutrients or energy sources by the microbe while also changing the chemistry of the fluid and causing precipitation of authigenic minerals, such as clays and zeolites within the granular cavity. In this way, bioalteration is thought to lead to the development of a complex microbial biofilm–authigenic mineral layer that eventually seals the system and cuts off communication between the microbe and circulating fluid. It is at this stage in the hypothesized bioalteration process that the role of cell extensions similar to fungal hyphae has been postulated, enabling tunneling through the chemical barrier to access fresh glass by the creation of a microtunnel (e.g., Fig. 15 Staudigel et al., 2008; Fig. 3 Staudigel et al., 2014). This hypothesized process has been compared to tunneling by fungal hyphae in soil feldspars, where localized dissolution at the tip of the hyphae causes tunneling, and enables separation of dissolution at the tip of the tunnel from waste production at the trailing end (Fig. 14 Staudigel et al., 2008). However, this process has never been directly observed in volcanic glass substrates, which, unlike feldspars or carbonate, lack crystallographic planes of weakness. The involvement of fungal hyphae in microtunnel formation is at present only a hypothesis. Questions also remain about the relative timing of granule and tubule formation, as textural evidence does not always support the onset of granular before tubular alteration, as there are several examples of advanced granular alteration in which ghosts or earlier microtunnels are still preserved (e.g., Fig. 30 Fisk and McLoughlin, 2013). In the absence of direct observations of microbial tunneling in volcanic glass, this hypothesized bioalteration mechanism remains unproven, and we therefore also need to entertain abiotic scenarios.
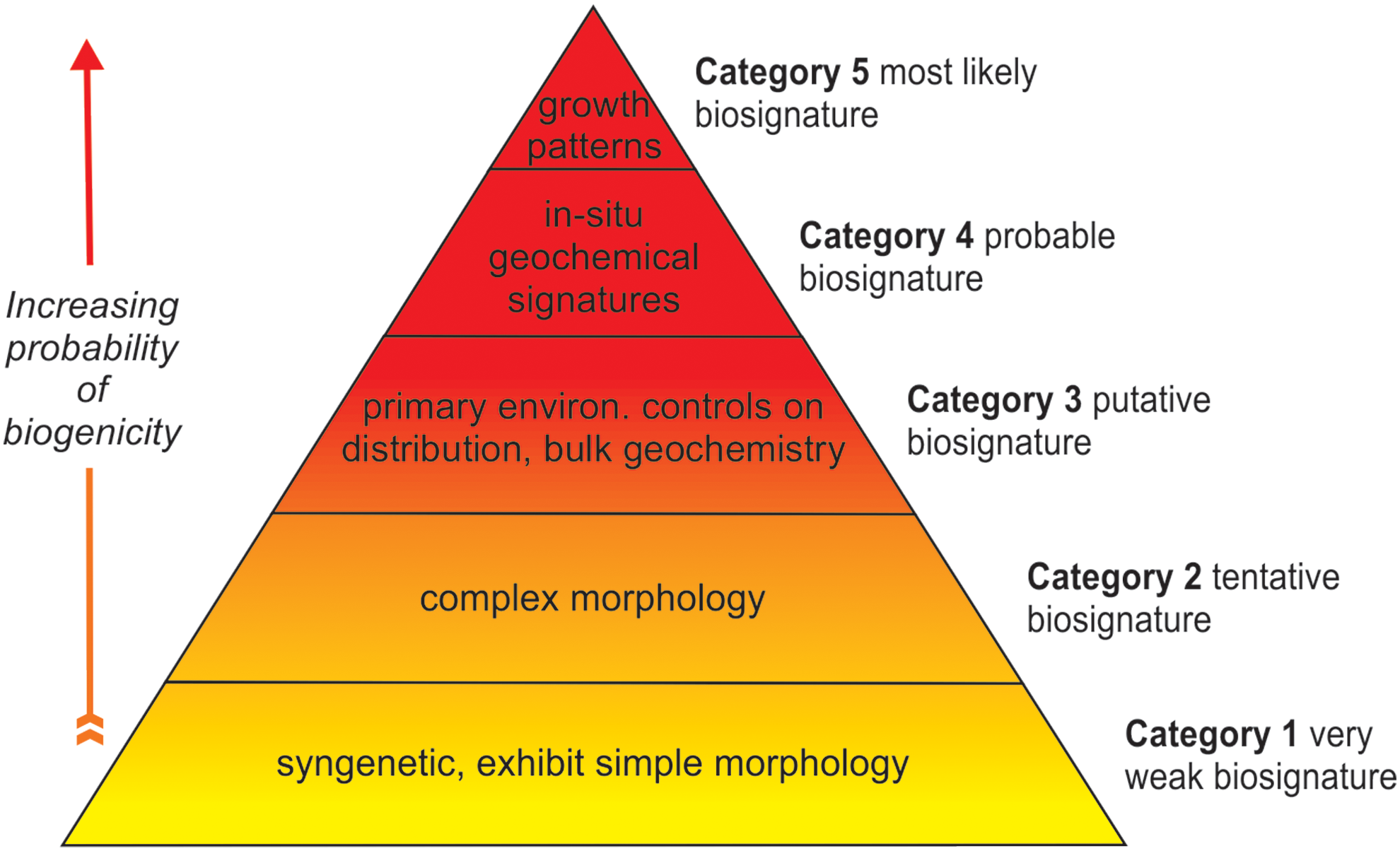
A pyramid summarizing the hierarchical biogenicity scheme and the lines of evidence that need to be combined to investigate a biogenic origin for a candidate biosignature. These criteria are additive, so to ascend the pyramid and achieve a higher category of biosignature requires fulfillment of all the criteria. Thus, for example, to achieve Category 3, termed a putative biosignature, requires that all the criteria for Categories 1, 2, and 3 be met. A detailed explanation of each criterion can be found in Table 1. (Color graphics available at

A flow diagram summarizing the sequence of data that need to be collected to test the hierarchical biogenicity criteria outlined here, with possible biotic or abiotic outcomes suggested. This sequence starts at the outcrop scale and progresses to the micron scale or nanoscale and is formulated in a way that could be applied to material collected by Mars sample return missions. (Color graphics available at
There are a number of abiotic mechanisms that have been suggested for granular and microtubular formation in volcanic glass. The processes envisioned include purely physicochemical dissolution of preexisting heterogeneities in glass, ambient inclusion trail–type processes, fluid-inclusion migration trails, and radiation-damage trails (e.g., McLoughlin et al., 2010; Lepot et al., 2011), which will be summarized now. Recent hydrofluoric acid etching experiments have brought the idea of chemical dissolution to the fore, as microtunnels of argued similarity in size and shape to candidate bioalteration textures were formed in laboratory experiments with volcanic and man-made glasses exposed to 1% HF (Fisk et al., 2013). These authors cautioned that formation of such microtextures by abiotic corrosion of basaltic glass is a viable hypothesis and that resolution of these alternative hypotheses, that is, biotic versus abiotic, is required before the microtunnels can be reliably interpreted as biosignatures. This is an important reminder that we need to continue to work to test such abiotic physicochemical processes, even if it is difficult to envisage how concentrations of strong acid such as those used in the laboratory experiments could be naturally sustained within the highly permeable oceanic crust. In addition, it is useful to consider what processes might give rise to preexisting heterogeneities in volcanic glass that could be exploited by purely chemical dissolution, for example, compositional variations resulting from heterogeneous cooling or phase separation in the glass and/or compositional and ultrastructural variation caused by flow banding in the cooling glass. These mechanisms remain hypothetical but could conceivably give rise to fine-scale heterogeneities in volcanic glass.
An alternative abiotic mechanism of tunneling that has been widely explored is that of ambient inclusion trails (AITs), which classically involve the migration of a mineral inclusion in a chert substrate (Tyler and Barghoorn, 1963; Knoll and Barghoorn, 1974) and more recently have been argued to involve migrating organic blebs leaving a hollow tubular trail in their wake (e.g., Lepot et al., 2011). This process is well known in cherts and phosphorites (e.g., Wacey et al., 2008b), with fewer putative cases described from metavolcanic glass (Lepot et al., 2009, 2011). The defining characteristics of the AIT process are longitudinal striae, a constant diameter, and polygonal cross section, sometimes with terminal inclusions (Wacey et al., 2008a). Although when considering the recent hypothesis involving the migration of organic blebs, these characteristics are not required; rather, a terminal organic bleb to a microtunnel that radiates from a site of initial fluid pressure is the only requirement. AITs appear, however, to be relatively uncommon in surveys of volcanic glass from the in situ oceanic crust, perhaps due to the scarcity of mineral and organic inclusions in glass and/or the absence of directional fluid pressure needed to propel the inclusion (for further arguments, see McLoughlin et al., 2010). Alternative abiotic mechanisms proposed for microtunnel formation in (meta)volcanic glass, namely, fluid inclusion trails and/or radiation damage trails, need to be taken into consideration but can be recognized due to their tendency to intersect.
Concerning microtextures in greenschist facies metavolcanic glass, the mechanisms of growth of the metamorphic mineral, typically titanite that forms the microtextures, need to be investigated. In a study that will be discussed in more detail below, Grosch and McLoughlin (2014) demonstrated, on the basis of radiometric dating, metamorphic mapping, and morphological analysis, that the growth of the titanite is related to much younger contact metamorphism that forms filamentous porphyroblasts. In a separate study of titanite microtextures found in late Archean volcanic glass fragments, it was argued on the basis of the textural context of the microtextures and their ultrastructure shown by scanning transmission electron microscopy (STEM) of focused ion beam (FIB) wafers that they are secondary, metamorphic mineral dendrites and not subseafloor microtextures (Lepot et al., 2011). In both of these cases, an understanding of the geological context as revealed by scientific drilling was important (Grosch et al., 2009a, 2009b; Philippot et al., 2009), as was petrographic analysis to reconstruct the local metamorphic history of the host rock. Both studies rejected a model for the microtextures involving metamorphic titanite growth infilling microbial microtunnels that formed earlier in the subseafloor environment.
4. Introducing the Hierarchical Biogenicity Scheme
The systematic characterization scheme proposed here involves a hierarchical sequence of information that is required to evaluate a microbial origin for a candidate biosignature. We propose a series of meso- to nanoscale criteria that are additive and yield increasing confidence of a biogenic origin from Categories 1 to 5. The criteria are tailored to microtunnels (MT) in volcanic glass or olivines and for candidate bacterial encrustations (BE) in alteration products such as palagonite. The scheme is illustrated in a pyramid shown in Fig. 2 and aims to evaluate the degree of support for a biogenic origin. The most useful biosignatures are those that have the strongest evidence of a microbial origin and are unlikely to be the result of abiotic processes (Category 5 in our scheme). So-called possible or candidate biosignatures (Categories 1–4 in our scheme) constitute only a working hypothesis, not a confirmation of detection of life, and need to be the target of further study using the types of approaches and criteria outlined herein. All these criteria are based on experience and arguments developed in studies of volcanic glass by a large number of authors, and similar principles and processes have more recently been proposed for microtextures in altered olivines. There is one notable difference, however, which is the much smaller size of microtextures in altered olivines, and this will be discussed further in Section 5.5 below.
The biosignature hierarchical classification system begins with observations at the outcrop or thin section scale and then proceeds to higher magnifications of investigation, for example, in situ micron to nanoscale geochemistry. The system is represented in tabular form with the biogenicity criteria (e.g., “Textural context and syngenicity”; see detailed explanation in text below) arranged in columns and the resulting categories of biosignatures in rows (Table 1). All criteria need to be satisfied at a given category before the candidate biosignatures can be evaluated at the next level. This new biogenicity scheme builds upon generalized biogenicity criteria for endolithic microborings proposed by McLoughlin et al. (2007). Here, we extensively update and extend previous criteria to include new discoveries and techniques that have been reported in recent years, and formulate these into a hierarchical approach for testing biogenicity. Our aim is not to provide statistics or quantify the probability of a biogenic origin, as this will require a community-wide effort to agree and quantify the limits on biologically controlled characteristics. Rather, we aim to provide a framework and a series of qualitative tests that can be used to evaluate the likelihood of a biological origin.

4.1. Textural context and syngenicity
The first criterion involves demonstrating that the microtexture is integral to the host rock and is syngenetic, that is, contemporaneous with early (subseafloor) alteration. Thus, for in-hand specimens or thin sections, it needs to be established that the microtextures are hosted by early phases and, if possible, cross-cut by late-stage phases or mineral veins. In seafloor basalts, for example, it is necessary to show that the microtextures are rooted in fractures along which early fluids circulated and are not associated with late-stage veins hosting neptunian sediments or perhaps seafloor fauna. In the case of candidate bacterial encrustations, it is helpful to show that these are hosted by early alteration phases in the paragenetic sequence, preferably on the outer margins of the fracture, and not in the late-stage alteration minerals at the center of fractures. In ophiolite-hosted samples, late-stage features resulting from post-obduction groundwater circulation should be avoided, and may be distinguished by oxidative alteration and/or the presence of immature organics.
In Precambrian greenstone belts, the metamorphic history is often more complex, and it is thus necessary to show that any candidate biosignatures pre-date secondary metamorphic mineral growth due to later regional or contact metamorphic events. In some instances, it may be possible to obtain absolute radiometric ages on some of these events by, for example, U-Pb dating of metamorphic titanite growth (e.g., Grosch and McLoughlin, 2014, and below). Where direct radiometric dating is not possible, relative age relationships from cross-cutting textural observations have to be utilized.
In meteorite samples, it is necessary to show that the microtextures and alteration phases of interest are pre-terrestrial in origin. Thus, fusion crusts on the outside of the meteorite should cross-cut or truncate the alteration features. Sometimes, it is also possible to use radiometric dating to show that the alteration minerals are pre-terrestrial, related to earlier fluids, and therefore much older than the age of terrestrial impact (e.g., Swindle and Olson, 2004).
In summary, demonstration of an early syngenetic origin is an essential first step to evaluating a candidate biosignature. In addition, evidence of at least simple morphological variation (see below) is necessary to fulfill the requirements for a Category 1 biosignature, considered to be of only very weak biogenic origin. If a microtexture cannot be shown to be syngenetic to early seafloor or early subsurface hydrothermal alteration, it is not worth pursuing as a candidate biosignature.
4.2. Analysis of microtexture size and shape variation
The second criteria concern variation in the shape and size of the microtextures, also known as morphospace analysis, and there are two guiding principles to test a biological origin. Firstly, microtextures of biogenic origin are expected to show a restricted size range typical of biogenic populations, as opposed to abiotic features that can encompass a wider range of dimensions (see explanation by Brasier et al., 2006). Secondly, a biological origin is supported by morphological sophistication, in particular, evidence of ornament or internal structure to the microtexture, which if possible can be shown to have a biological function. In parallel, abiotic origins for the morphological complexity must also be tested and rejected, as increased morphological complexity does not simply imply an increased probability of biogenic origins.
Concerning the size range of candidate bioalteration textures, it has been found that microtunnels in volcanic glass of the in situ oceanic crust typically range in diameter between 0.5 and 1μm, with an average width of c. 2 or 3 μm, and granular textures are typically less than 1.4 μm in diameter (Figs. 6 and 7 Furnes et al., 2007). This relatively narrow size distribution is consistent with a biological origin, and such a population therefore deserves further investigation, especially the geochemical signatures (see below) to ascertain biogenicity. In comparison, the average width of titanite microtextures in Archean metavolcanic glass has been found to be greater (Furnes et al., 2007) and more variable (Fig. 1 Grosch and McLoughlin 2014), and this has been significant in recent studies contesting the biogenicity of the microtextures.
Morphological variation in candidate biosignatures has been well studied and is a valuable tool in evaluating putative origins. Two morphological groups are identified in volcanic glass, namely, granular and tubular microtextures, with the tubular type being further subdivided by using seven morphological characteristics: (1) length and width, (2) density, (3) curvature, (4) roughness, (5) variation in width, (6) branching, and (7) tunnel contents (Fisk and McLoughlin, 2013). An illustrated guide to these morphological features based upon the study of 119 samples from across the world's oceanic basins is available in a recent petrographic atlas (Fisk and McLoughlin, 2013). Complex morphological features to look out for include internal spirals, tunnel divisions or septae, annulations, and putative cell remains within the glass-hosted microtunnels (e.g., Fisk and McLoughlin, 2013). If found, such features warrant further investigation to confirm if they are primary and if they have any biological significance, and to exclude possible abiotic origins, for instance, AITs. In the case of candidate bacterial encrustations in palagonite or zeolites, any variations in the cell wall ultrastructure and internal bodies should be investigated (Thorseth et al., 2003). Several high-magnification imaging techniques have been developed over recent years to better document such submicron-scale features, for example, FIB-SEM (e.g., Wacey et al., 2014) and laser scanning microscopy (e.g., Dultz et al., 2014). At the same time, there are several morphological features that are diagnostic of abiotic processes and should be avoided. Most notably, microtunnels with terminal inclusions or crystals, perhaps also longitudinal striae indicative of AIT processes (see above).
In summary, evidence of a restricted range in size and of complex morphology is necessary to confirm a Category 2 biosignature, which is regarded as a tentative biosignature in this hierarchical scheme. In the case of candidate bacterial encrustations in authigenic minerals, this means internal structure, and for microtunnels in volcanic glass, ornament (e.g., spirals) on the microtunnel walls.
4.3. Geological and physicochemical setting
The third criterion involves mapping of the geological and physiochemical variables that control the occurrence and preservation of the candidate biosignatures. Collection of samples in outcrop or drill core across lithological and environmental gradients allows for investigation of the impact of biologically relevant variables (e.g., thermal environment, water depth, pore fluid geochemistry, nutrient concentration) on the distribution of candidate microtextures, and can help identify those samples that are best preserved. Moreover, any correlation between variations in the size, shape, abundance, or location of the candidate biosignatures with primary environmental variables can strengthen the case for biogenicity or potentially weaken it. Quantitative studies of this type are few in number, but one exception is a study of the abundance of microtunnels in basaltic hyaloclastite across a subglacial-marine transect in Neogene lavas from Antarctica (Cousins et al., 2009). Here, microtunnel densities were measured across salinity and elevation gradients, and it was found that the abundance of microtunnels was greatest in the most marine-influenced samples, suggesting a connection between salinity and microtunnel formation. The challenge is then to establish whether this trend reflects microbial salinity tolerance or purely physicochemical dissolution.
Further support for a biogenic origin could be obtained from establishing a correlation between primary geochemical parameters, such as subsurface fluid composition, with the abundance of the microtextures. A compilation of data from the subseafloor has shown that the abundance of granular and tubular microtextures reaches a peak at c. 150 m depth, and this has been compared to parameters such as porosity, permeability, and clay mineralogy (Furnes and Staudigel, 1999; Furnes et al., 2001a). The challenge is to interpret this pattern, however, because the abundance of the microtextures is a time-integrated product across different fluid environments (temperature, oxidation state, flux). Thus, to demonstrate a unique biological control is difficult. The task is perhaps easier when comparing phylogenetic data from the active microbial community in the subseafloor to borehole fluid properties, or C and S isotope fractionations, for instance (e.g., Lever et al., 2013), which allows for the most likely microbial metabolisms to be inferred at different depths and conditions in the subseafloor. If these types of data sets could be linked to the textural evidence in volcanic glass, an important step forward could be made in this field.
In the case of metavolcanic glass, investigating the distribution of the microtunnels across metamorphic field gradients can help distinguish preservational gradients from primary, possibly biologically controlled, variations in the abundance of the microtextures. Thus, in the seafloor, an increase in metamorphic grade due to burial metamorphism or zones of hydrothermal alteration may affect the distribution of the microtextures. In ophiolites, metamorphic facies changes related to obduction processes may affect the distribution of microtextures, but to date this scenario has not been documented by systematic metamorphic mapping studies. In metavolcanic glass of Archean greenstone belts, the metamorphic history may be yet more complex with regional metamorphic overprinting and/or contact metamorphic events needing to be quantified and their impact on the microtextures investigated (see case study below). In Archean early life studies, scientific drilling can be very valuable in yielding continuous drill core samples across various geological and geochemical environments (e.g., Grosch and McLoughlin, 2013) to test potential biotic and abiotic controls on the distribution of candidate microtextures.
In summary, to establish a biosignatures as Category 3, termed a putative biosignature, it is necessary to demonstrate all the features of Category 2 and that the distribution and abundance of the microtextures is controlled by primary environmental factors. In addition, a strong case should be made to illustrate that the geochemical evidence such as organics and bulk isotopic signatures are syngenetic and likely indicative of microbial origin (see below).
4.4. Geochemical evidence syngenetic to the microtexture
Geochemical evidence of microbial activity is required to further strengthen the case for biogenicity, and this is most powerful if it is in situ data that are texturally tied to the candidate biosignature. Bulk geochemical data can be instructive, but spatially resolved elemental or isotopic data are more powerful. In all instances, it is essential to demonstrate that the geochemical evidence is syngenetic to the microtextures and related to early processes. There are a whole range of analytical techniques that can be employed, which include electron microprobe (EMP) mapping of host minerals; Raman mapping of carbonaceous remains and associated minerals; FIB preparation of ultrathin wafers across microtextures that can then be analyzed by TEM for elemental data, by nanoSIMS for elemental or isotopic measurements, and by synchrotron-based X-ray techniques for redox data. Several of these techniques will be discussed in the case studies below.
In a notable study, Storrie-Lombardi and Fisk (2004) proposed a quantitative method for discriminating between biotic and abiotic alteration of subseafloor glass based on the composition of the alteration phases. This study involved altered volcanic glass from three sites that was attributed to biotic or abiotic origins on the basis of the morphology of the boundary between the glass and alteration products. EMP analysis was then conducted to determine the composition on the glass and alteration products, and 11 key elements were used for principal component analysis (PCA). Three factors were identified as accounting for 80.5% of the variance in the data and served as inputs to a hierarchical cluster analysis algorithm to identify clusters in the data. Finally, the PCA factors and the objectively detected cluster assignments (cluster identified without a human observer and evaluation) were used to train a nonlinear stochastic artificial neural network to predict the Bayesian likelihood of correct classification, that is, as biotic, abiotic, or unaltered glass. At these locations, the autonomous classification algorithm agreed with the man-made petrographic evaluation in 14 of the 15 biotic clays studied. Thus, in this small data set it appears possible that quantitative discrimination between biotic and abiotic alteration can be made on the basis of chemical composition. The challenge is to extend this parameterization approach to other localities, compositions of glass, and other types of evidence, for example, microtunnel morphology, to test whether biotic and abiotic morphologies can be discriminated quantitatively.
One of the most discussed geochemical lines of evidence for biologically mediated alteration is carbonaceous material associated with microtextures in altered volcanic glass and olivines. In some settings, for example meteorites, it may be possible to extract bulk carbonaceous material and use techniques such as flash pyrolysis–gas chromatography–mass spectrometry and time-of-flight secondary-ion mass spectrometry to investigate the macromolecular structure and isotope composition of these organics to evaluate possible sources (e.g., Sephton et al., 2002, 2003; McKay et al., 2006). A stronger case for biogenicity can be made if the organics can be shown to be early and related to the microtextures; this can be investigated at the micron to submicron level with, for example, Raman spectroscopy (Ménez et al., 2012), FIB-SEM (Wacey et al., 2014), or both. The presence or absence of heteroatoms (N, P, S) within the carbonaceous material can be mapped with TEM-EELS and nanoSIMS (e.g., McLoughlin et al., 2012; Wacey et al., 2014), and if sufficiently abundant the carbon isotope signature of the carbonaceous material can be measured in situ by nanoSIMS to test possible sources (e.g., Wacey et al., 2008b). We note that EMP mapping of light elements, especially carbon (e.g., Furnes et al., 2001a; Banerjee et al., 2007), has proven to be unreliable in this context, as discussed further below (Section 5.4). In meteorite samples, all the same techniques can be applied to investigate pre-terrestrial organics (e.g., McKay et al., 2011).
Bulk geochemical data can be useful for investigating whether there was microbial activity at a given study site and can be used as a reconnaissance tool for identifying areas to target for further textural and in situ geochemical evidence. Thus, sulfur isotope analyses of secondary sulfides in altered glass and mafic and ultramafic rocks from the in situ oceanic crust have found δ34S fractionations that are comparable to those resulting from bacterial sulfate reduction, especially at low metabolic rates (Delacour et al., 2008; Rouxel et al., 2008; Alt and Shanks, 2011; Ono et al., 2012; Lever et al., 2013). Thus, δ34S values in pillow lava–hosted sulfides that are strongly shifted away from mantle values have been reported from ophiolites (Parnell et al., 2014) and Archean greenstone belts (McLoughlin et al., 2012) and are argued to support microbial sulfate reduction. Carbon isotopes on disseminated carbonate in pillow rims have been found to be shifted away from both seawater and mantle values; thus the carbonate has been argued to be a by-product of subseafloor microbial activity (e.g., Furnes et al., 2001b; Cockell et al., 2010). But unlike secondary sulfides that can texturally be shown to be early alteration phases in vesicles and fractures, disseminated carbonates are of less certain syngenetic origin; thus alternative secondary processes need to be excluded. Likewise, bulk δ13C measurements from organic carbon found in palagonite have been interpreted to be consistent with microbial activity (e.g., Kruber et al., 2008), but the limitation of such data sets is that they are not diagnostic of specific microbial metabolisms and cannot be related to particular alteration textures. We therefore stress again that the most powerful geochemical information comes from in situ data that are texturally tied to the candidate biosignatures.
Variations in the composition of the matrix surrounding the microtextures are also of interest and can be used to test formation mechanisms. For example, investigations of so-called leached zones in recent volcanic glasses, which are typically <20 nm wide and contain elemental depletions that surround the microtunnels, suggest incongruent dissolution of the host glass during microtunnel formation (e.g., Alt and Mata, 2000; Fliegel et al., 2012; Knowles et al., 2012). Alteration minerals are also of interest; for instance, the concentration of transition metals, particularly Fe and Mn in the rims of candidate bacterial encrustations, can be investigated to explore potential microbial metabolisms (e.g., McLoughlin et al., 2011) and bacterially induced biomineralization processes (e.g., Pedersen et al., 2015). In greenschist facies metavolcanic glasses, thermodynamic modeling of mineral compositions surrounding candidate microtextures can be used to map metamorphic temperature variations on a microscale and then to test the origin of the microtextures and investigate their preservation (e.g., Grosch et al., 2014, and case study below). Redox variations in the minerals surrounding candidate biosignatures in altered volcanic glass have been explored in only a small number of studies using synchrotron-based X-ray techniques (e.g., Templeton et al., 2009; Fliegel et al., 2012; Knowles et al., 2013). For this reason, mapping redox variations has been included as a requirement of a Category 5 biosignature. Redox data measured on iron- and manganese-bearing phases need to be interpreted in terms of either changes in the redox conditions of the alteration fluids or locally controlled microbial processes—this can be challenging and is thus at present a desirable requirement of a Category 5 biosignature.
In summary, to establish a biosignature as Category 4, termed a probable biosignature, it is necessary to demonstrate all the features of Category 3 plus in situ elemental and isotopic signatures preserved both in the microtexture itself and the surrounding matrix. To satisfy all these requirements is extremely demanding and in many instances will not be possible, because the quality of preservation is insufficient to find, for example, organic remains.
4.5. Characterization of putative growth patterns
This criterion concerns the identification of growth patterns that are indicative of biological behavior. A key requirement for a biogenic origin is that the microtexture must not intersect, whether they are microtunnels in altered glass or olivines, and the microtextures must avoid one another. Conversely, abiotic structures such as radiation damage trails, AITs, and fluid inclusion trails can be expected to intersect, as they do not need to share the substrate. The absence of intersecting microtextures is one of the most important growth and distribution criterion for a biogenic origin, but this alone is not definitive proof of biogenicity (e.g., Fliegel et al., 2010b). In addition, there are several other examples of the selective distribution or preferred orientations of microtextures that have been interpreted to support a biogenic origin, although we stress that, at most, these are only consistent with biological activity and rely upon experimentally untested observations. For this reason, growth patterns suggestive of biogenic behavior form part of a Category 5 biosignature and are one of the most challenging criteria to establish.
The distribution of microtunnels around phenocrysts in volcanic glass, vesicles, and along flow banding has been interpreted to record microbial behavior. For example, the apparent preference of microtunnels for olivine phenocrysts has been suggested to reflect a preference for iron-bearing minerals (Walton, 2008) but alternatively may reflect stresses in the glass caused by differential cooling around a phenocryst, which would need to be distinguished. Likewise, microtunnels that radiate from vesicles in volcanic glass have been interpreted to reflect microbes tunneling from initial sites of fluid infiltration (Fig. 5 Furnes et al., 2007, and references therein), but again this distribution pattern could record the tunneling process exploiting stresses and weaknesses in the glass. Putative examples of microtunnels exploiting “flow banding” or microstructural weakness in once-viscous, silicate glass have also been reported (e.g., Fig. 4a, 4b in Furnes et al., 2001a). Quantitative image analysis of these types of textural observation will help test different growth hypotheses, and efforts have been made to modify the ichnofabric index developed for trace fossils in sedimentary rock so it can be applied to microborings in volcanic glass (e.g., Montague et al., 2010). The most robust demonstration of putative biological growth patterns will come, however, from in situ incubation of microbes on glass substrates under controlled experimental conditions.
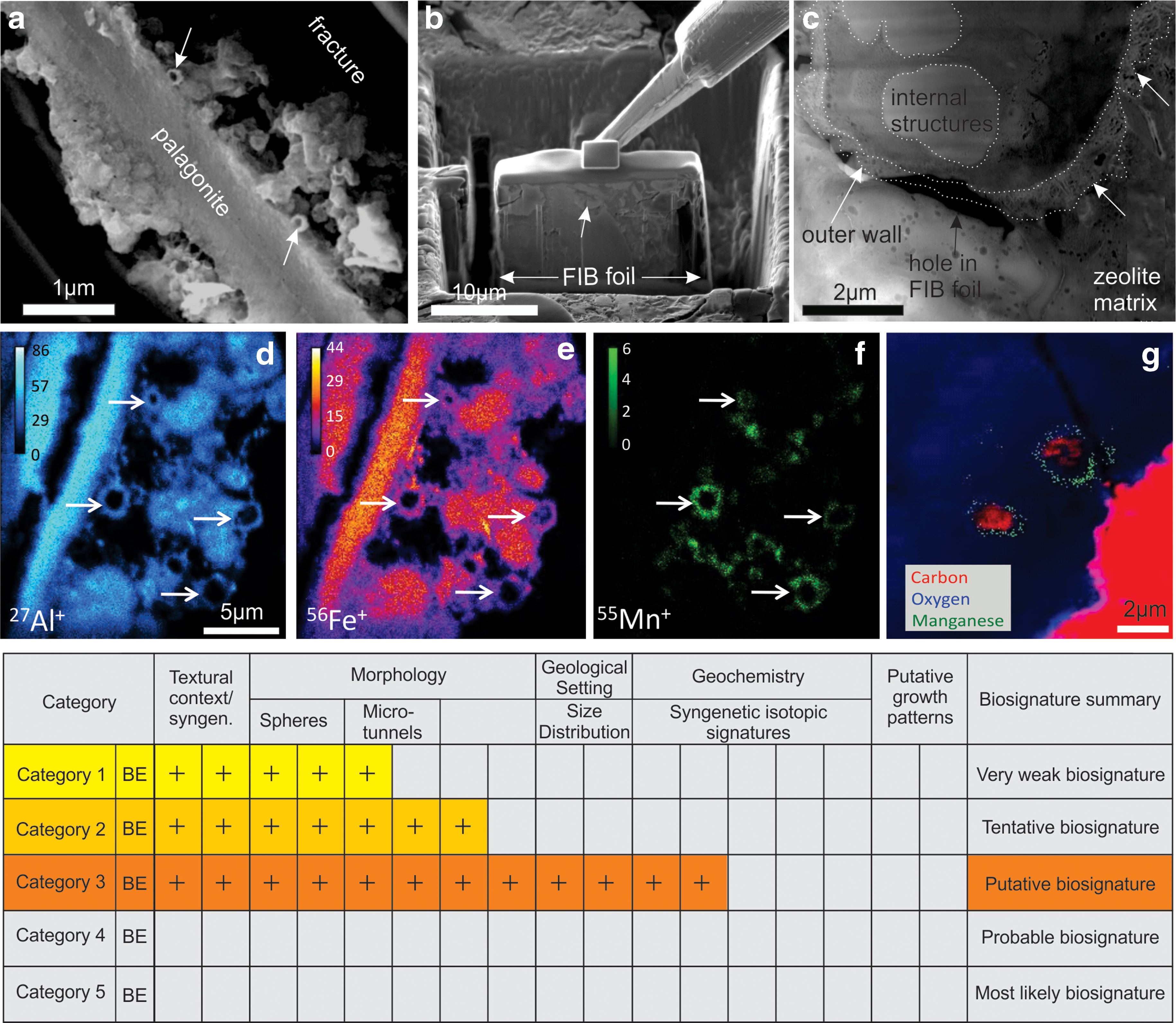
Candidate bacterial encrustations from subseafloor environments. (

Microtextures in altered volcanic glass from the in situ oceanic crust near Costa Rica, a Category 1 biosignature (a, b); from the Cretaceous Troodos ophiolite of Cyprus, a Category 3 biosignature (c, d); and Archean metavolcanic glass of the Barberton greenstone belt, South Africa, which fails the biogenicity criteria (e, f). (
In short, demonstration of a Category 5 biosignature, the origin of which is most likely to be biogenic, requires characterization of biologically controlled growth patterns in the form of preferred orientations or distributions of the microtextures. Category 5 also requires the mapping of local redox variations in the matrix surrounding the microtextures, as described in the geochemistry section above. To confirm the presence of microbial activity in terrestrial volcanic glass and especially martian samples, a Category 5 biosignature is most desirable.
4.6. Summary of the hierarchical biogenicity categories
To summarize the hierarchical biogenicity scheme proposed here, demonstration of an early syngenetic origin and of simple morphological variation is required to fulfill the requirements for a Category 1 biosignature, which exhibits weak or questionable evidence with regard to a biogenic origin. Documentation of a restricted size range and of complex morphology (meaning internal structure for candidate bacterial encrustations, and ornament on microtunnels in volcanic glass) is necessary to confirm a biosignature in Category 2, regarded as a tentative biosignature. Additional evidence that the distribution and abundance of the microtextures is controlled by primary environmental factors, along with bulk isotope fractionations indicative of microbial activity, and characterization of organics syngenetic to the microtextures are needed to confirm a candidate biosignature as Category 3, which is termed a putative biosignature. To achieve a Category 4 biosignature, termed a probable biosignature, requires preservation of in situ elemental and isotopic signatures within the microtexture itself and the surrounding matrix. To confirm a Category 5 biosignature, fulfillment of all the above is required along with evidence of biologically controlled growth patterns and local redox gradients around the microtextures. A Category 5 biosignature has the strongest support for a biogenic origin and therefore provides the most convincing evidence of microbial activity in subseafloor volcanic glass or ultramafics—no terrestrial example yet described achieves this category. The hierarchical scheme is summarized in a flow diagram in Fig. 3 to illustrate the sequence of information that can be collected from terrestrial and extraterrestrial samples, with possible biotic or abiotic outcomes. This iterative approach encourages continuous improvement of the biogenicity criteria by combining petrological and geochemical evidence to test the biological principles upon which the criteria are built. Below, we will illustrate how these criteria can be applied to a number of examples.
5. Application of the Hierarchical System for Evaluating Biogenicity
In this section, we illustrate how these biogenicity criteria can be applied to several terrestrial examples and to the martian meteorite Yamato 000593. We emphasize that continued development of new analytical methods with which to investigate volcanic glass-hosted microtextures will aid the collection of new data to satisfy the criteria proposed here and help to further refine these criteria. Many of the candidate bioalteration examples described below rank low on our hierarchical biogenicity scheme because key evidence is lacking (see Supplementary Tables S1–S5, available online at
5.1. Candidate bacterial encrustations from the Arctic Mohns Ridge and Australian Antarctic Discordance
To illustrate the application of the new hierarchical biogenicity criteria to candidate bacterial encrustations, we will focus on examples from the Arctic Mohns Ridge (Kruber et al., 2008; McLoughlin et al., 2011) and Australian Antarctic Discordance (AAD) IODP Leg 187 (Thorseth et al., 2003; Pedersen et al., 2015), illustrated in Fig. 4. The Mohns Ridge samples contain rounded to elongated pores 0.5–2 μm across embedded in palagonite that have sizes and shapes comparable to microbial cells in <100,000-year-old seafloor basaltic lavas. Light microscopy observations confirmed that these microtextures are part of the early alteration assemblage, as they are concentrated along the glass palagonite interface and are embedded in the texturally oldest generations of palagonite (Fig. 2 McLoughlin et al., 2012). In situ elemental mapping by TEM-EDS revealed that the micropore rims are comparable in composition to the bulk palagonite and that some are enriched in manganese. Thus, nanoSIMS mapping (Fig. 4d–4g) was used to investigate these further and revealed elevated concentrations of carbon and nitrogen in some of the micropores (e.g., Fig. 5 McLoughlin et al., 2011). Hence, these structures were interpreted as fossilized bacteriomorphs of endolithic microorganisms. The preferential accumulation of Mn in some of the cell encrustations suggests the mineralization of Mn-oxidizing bacteria, as suggested from other sites on the basis of genetic evidence and synchrotron X-ray mapping (e.g., Templeton et al., 2005). The AAD samples include 0–2.5 Ma dredged seafloor basalts containing spherical, rod-, and star-shaped structures in palagonite (e.g., Fig. 1 Thorseth et al., 2003). In older 18–28 Ma drill core samples from the AAD, coccoid structures embedded in zeolites were reported that show Mn enrichments, a complex radial ultrastructure, and discontinuous carbonaceous linings as revealed by FIB-TEM (Fig. 4c). The microtextures also contain internal subspherical structures (Pedersen et al., 2015).
Applying the hierarchical scheme proposed here, the Mohns Ridge candidate bacterial encrustations are (i) hosted by early phases, and their syngenicity is confirmed by relative age constraints; (ii) exhibit relatively simple morphologies with little variation in size; (iii) preserve geochemical evidence with enrichments in Mn on the rims of the coccoids, concentrations of C, N within the microtextures, and bulk carbon isotope data with a δ13Corg of −22‰ from 0.9 wt % carbon interpreted to be derive from both living and fossilized biomass (Kruber et al., 2008). However, two of the biogenicity criteria are missing or incomplete; namely, quantitative constraints on the shape and size range of the microtextures are not reported. Secondly, clear demonstration that geological variables control the distribution of the microtextures is not reported, owing probably to the small sample set available. Thus, the Mohns Ridge bacterial encrustations achieve only a Category 3 biosignature, that is, unless these further lines of evidence are presented (Fig. 4 and Supplementary Table S1). In comparison, the AAD samples currently achieve a Category 2 biosignature given the presence of internal structures and the complex outer wall ultrastructure (Fig. 4c).
5.2. Microtunnels in volcanic glass from the Costa Rica Rift ODP Hole 896A
To illustrate the application of the new hierarchical biogenicity criteria to candidate bioalteration textures in oceanic pillow lavas, we will focus on ODP Holes 896A and 504B from the Costa Rica Rift (Fig. 5a, 5b), where candidate microbial microtextures were described in 1996 (Furnes et al., 1996; Giovannoni et al., 1996). Textural and scanning electron microscopy (SEM) investigations found spherical alteration textures and so-called vermicular bodies <70 μm long at the interface between fresh and altered glass, up to 432 m into the volcanic basement that is ∼6 million years old (Furnes et al., 1996). The size range of the granular and tubular microtextures has been quantified together with size range data determined at a site of similar age, Hole 504B, which is also from the Costa Rica Rift and typical of microtextures known from the in situ oceanic crust that span 0–1.4 μm in diameter for the granules and 0–11 μm in diameter for the microtubes (Figs. 6 and 7 Furnes et al., 2007). The initial reports interpreted that “these textures by their size and shape strongly indicate that they were produced by the etching of microbes” (Furnes et al., 1996). In addition, EMP analysis was used to document elemental enrichments [in K2O, P2O5, ± TiO2, ± Al2O3, FeO(t)] and depletions (in CaO, Na2O, SiO2, and MgO) in the vicinity of the microtextures that were argued to reflect microbially mediated dissolution of the glass (Furnes et al., 1996). Microbiological data included fluorescent dyes, which indicate the presence of nucleic acids at the interface of fresh and altered glass, especially at the distal tips of microchannels, along with the presence of prokaryotic DNA (Giovannoni et al., 1996). Further 16sRNA work found bacterial and archaeal DNA (Torsvik et al., 1998). These workers also reported negatively fractionated δ13C values of less than −7‰ in disseminated carbonate from pillow rims, which are argued to suggest biological activity (see also Furnes et al., 2001a).

Alteration textures in olivine from martian meteorite Yamato 000593 imaged using SEM and currently evaluated as a Category 1 biosignature. (
Applying the hierarchical biogenicity criteria proposed here, we reevaluate the microtextural evidence from Hole 896A. Firstly, the relative age constraints and mineral paragenesis support an early origin for the microtextures. Secondly, they show simple tubular morphology and thus satisfy the requirements for a Category 1 biosignature that exhibits weak or questionable evidence with regard to a biogenic origin. Considering now the requirements for Categories 2 and 3, the DNA staining work was hampered by concern over the syngenicity of the fluorescent material and the possibility that these were later adsorbed onto the clay minerals infilling the microtextures. The size range of the granular and tubular microtextures is relatively restricted and typical of microtextures known from the in situ oceanic crust. Bulk δ13Ccarb isotope data are available (Furnes et al., 2001a) but provide only weak support given, again, concerns about syngenicity. The elemental data from EMP mapping lack the appropriate spatial resolution to the microtextures to critically evaluate formation mechanisms, and rather give a bulk time-averaged perspective on the seafloor alteration regime (Furnes et al., 1996; Torsvik et al., 1998). FIB-TEM studies have not found a discrete organic phase at the interface of the fresh and altered glass, which is absent from FIB wafers cut across the Costa Rica microtextures (Alt and Mata, 2000; Knowles et al., 2012) and several other examples investigated (e.g. Benzerara et al., 2007; Fliegel et al., 2012). These studies have found a leached zone in the glass rimming the microtextures, but this was not used to make strong conclusions regarding biotic or abiotic formation mechanisms (e.g., Knowles et al., 2012). Thus, the microtextures are characterized as Category 2 and are termed tentative biosignatures in the hierarchical scheme outlined here due to the absence of internal complexity to the microtextures and the lack of compelling in situ isotopic or elemental data in support of microbial processes (Supplementary Table S2). In addition, a much stronger case needs to be made for biologically relevant variables that control the distribution and abundance of the microtextures.
5.3. Microtunnels in volcanic glass of the Troodos Ophiolite, Cyprus
The hierarchical biogenicity criteria will now be applied to microtunnels in volcanic glass of the ∼92 Ma Troodos ophiolite of Cyprus, which include some of the largest and most morphologically complex microtunnels described from volcanic glass (Furnes et al., 2001b; McLoughlin et al., 2009). The variation in microtunnel morphology (Fig. 1g, 1h and Fig. 5c, 5d) at this locality includes annulated forms, complex spiral shapes, and microtunnels with internal structures (McLoughlin et al., 2009). To test the geochemical evidence associated with these microtextures, focused ion beam serial sectioning was combined with scanning electron microscopy (3D-FIB-SEM) to image the contents of the microtunnels combined with TEM mapping by Wacey et al. (2014). These analyses revealed a distinct 50–200 nm thick carbonaceous lining within the annulated microtube investigated (see Fig. 9 Wacey et al., 2014). The lining was found to be almost continuous and closely replicates the outer annulated morphology of the terminal end of the microtube. A spiral substructure previously seen when using optical microscopy was also shown to be carbonaceous, comprising sheets that, in places, span the entire width of the microtube. Elemental mapping and electron energy loss spectroscopy (EELS) confirmed that the carbon is organic and co-occurs with nitrogen. EELS and selected area electron diffraction revealed that a clay mineral infills the remainder of the microtube, with a chemistry and structure consistent with Fe-rich smectite (nontronite). Anticorrelation between carbon and the elements Al, Ca, Fe, Mg, O, Si, in the infilling clay, and the morphology of the carbon, show that the carbon lining is a discrete structure that predates clay mineral growth, which thereby excludes the later absorption of organics onto the clay. These findings also rejected a hydrothermal or metamorphic fluid-derived origin for the carbonaceous linings to the microtube, and an origin from recent post-obduction groundwater-hosted microorganisms. The source favored by these authors was thus from microbial biomass in the Cretaceous subseafloor, but they stressed that it is not possible to ascertain whether the organic material is connected to the tunneling process (Wacey et al., 2014). It is somewhat paradoxical to note that an ophiolite-hosted microtunnel preserves organic remains, whereas in situ oceanic crustal samples have not been found to contain discreet organic linings (see previous section).
To summarize the data collected on the Troodos microtextures and how these are evaluated using the hierarchical scheme proposed here, firstly, the microtextures formed early in the alteration history of the lavas. Secondly, the microtextures show two apparent size and morphological groups: the more abundant and narrower microtunnels (e.g., Fig. 2E Furnes et al., 2001b) and the wider and more morphologically elaborate microtunnels (ornamented variety) that are less abundant (e.g., Fig. 2F Furnes et al., 2001b; Fig. 5 McLoughlin et al., 2009). Thirdly, the microtunnels preserve in situ geochemical evidence of early organic linings, although their direct connection to the tunnel-making process is unclear (Wacey et al., 2014). Thus, the Troodos microtunnels are tentatively characterized as a Category 3 biosignature, although there are several outstanding concerns. Firstly, the larger and more morphologically complex microtunnels are a less abundant component of the Troodos microtunnel population and should therefore not be over-emphasized in assessments of a biological origin for all the microtunnels. In addition, these morphological and size variations in the microtunnel population need to be better quantified. Secondly, the organic-bleb AIT mechanisms of Lepot et al. (2011) need further exploration and testing for the Troodos microtunnels, given the occurrence of large vesicles, possibly organic-bearing in the vicinity of some of the larger microtunnels (Fig. 2F Furnes et al., 2001b). Thirdly, a strong case has yet to be made for whether primary environmental factors or preservational processes control the distribution of the Troodos microtextures (Supplementary Table S3).
5.4. Archean microtextures in pillow lavas of the Barberton greenstone belt, South Africa
Microtextures found in c. 3.45 Ga pillow lavas of the Barberton greenstone belt, South Africa (Fig. 5e, 5f), have been argued to represent mineralized bioalteration textures formed by microbial activity in the Paleoarchean subseafloor (Furnes et al., 2004; Banerjee et al., 2007; Fliegel et al., 2010a; Staudigel et al., 2015); thus filamentous titanite microtextures in metavolcanic glass have been advanced as a search image for life on Mars (e.g., Banerjee et al., 2009; Izawa et al., 2010). However, a recent study rejected the evidence proposed for the biogenicity and syngenicity of these microtextures, arguing that they are not reliable biosignatures (Grosch and McLoughlin, 2014, 2015). The new findings come from pillow lavas of the Barberton Scientific Drilling Project that targeted the original “biosignature” locality in the Hooggenoeg Formation (Grosch et al., 2009a, 2009b). Petrographic analysis revealed that the microtextures exhibit a morphological continuum that bears no similarity to candidate bioalteration textures found in the modern oceanic crust (Fig. 1 and S1 Grosch and McLoughlin, 2014). Moreover, the titanite filaments have a much larger range of diameters than previously appreciated, between 0.5 and 40 μm, and are typically an order of magnitude thicker compared to modern tubular microtextures (Fig. 1 Grosch and McLoughlin, 2014). Extensive in situ elemental mapping by nanoSIMS did not find any organic linings associated with the microtextures, despite the high spatial resolution and sensitivity of the nanoSIMS (Fig. 1 McLoughlin et al., 2012). Thus, previous apparent organic linings were re-interpreted as a result of subtle topographic variations in the samples (for a full explanation, see the work of McLoughlin et al., 2012). Indeed, the original authors have now acknowledged that carbonaceous linings are not preserved in the Hooggenoeg microtextures (Staudigel et al., 2015). Radiometric dating of the filamentous microtextures yielded a U–Pb titanite age of 2.819 ± 0.2 Ga, which is indistinguishable from thermal-related titanite growth dated at 2.913 ± 0.31 Ga that occurs adjacent to a local mafic sill in the drill core. These ages overlap with the estimated age of intrusion of the sill and are c. 650 Ma younger than the eruptive age of the Hooggenoeg pillow lavas (Grosch and McLoughlin, 2014, 2015). These new titanite ages allowed for rejection of a seafloor alteration origin for the titanite microtextures and hence their syngenicity. Furthermore, quantitative mapping and analysis of the metamorphic host rock revealed that the titanite filaments are best developed in relatively low-temperature microdomains, supporting a late Archean thermal contact metamorphic origin for the microtextures. Thus, Grosch and McLoughlin (2014, 2015) strongly questioned the syngenicity and biogenicity of these purported microbial biosignatures and argued rather for an origin as abiotic porphyroblasts of thermal contact metamorphic origin, which grew during late-stage retrograde cooling of the pillow lavas adjacent to a diabase sill (Grosch and McLoughlin, 2014, 2015).
In the hierarchical scheme proposed here, the Barberton titanite microtextures do not achieve Category 1 because they are not syngenetic to Archean c. 3.45 Ga seafloor alteration but rather are younger abiotic metamorphic porphyroblasts (Supplementary Table S4). In addition, the microtextures fail the size and shape criteria outlined above and lack organic linings and geochemical support for a biological origin. Their distribution is controlled by younger metamorphic events and not by primary environmental variables (Grosch and McLoughlin, 2014, 2015). More broadly, the authors contended that a full characterization of low-temperature metamorphic events and alternative biosignatures in greenstone belt pillow lavas is required before candidate traces of life can be confirmed in Archean subseafloor environments. It is thus time to revisit titanite microtextures from the Pilbara Craton of Western Australia and other Precambrian greenstone belts (e.g., Abitibi, Canada) and to apply techniques such as FIB-TEM and nanoSIMS to address the hierarchical biogenicity criteria proposed herein. We anticipate that these Precambrian candidate biosignatures will also fail several of the biogenicity criteria, especially concerning their syngenicity, and may also rank as Category 1 biosignatures or less, based upon published data and questions concerning the timing and origins of titanite growth. Thus, filamentous titanite is not a reliable search image for textural evidence of extraterrestrial life.
5.5. Altered olivines in the martian meteorite Yamato 000593
Microtunnels in altered olivines of the martian meteorite Yamato 000593 were recently reported along with indigenous carbonaceous remains (White et al., 2014a), and will now be evaluated by using the hierarchical criteria outlined here. Yamato 000593 was found in Antarctica in 2000 and belongs to the nakhlite group of martian meteorites (Imae et al., 2003). It is an unbrecciated clinopyroxenite, a cumulate igneous rock composed of coarse-grained augite (pyroxene) with accessory olivine and opaques. The crystallization age has been determined by Sm-Nd isochron as 1.31 ± 0.03 Ga (Shih et al., 2002). It is only lightly shocked, has a black fusion crust, and shows strong evidence for aqueous alteration with well-developed iddingsite veins that are truncated by the fusion crust, which suggests that the alteration may have occurred prior to atmospheric entry and is martian in origin (Treiman and Goodrich, 2002). In a recent study by White et al. (2014a), microtubular features were described emanating from the iddingsite veins into the surrounding olivines. These are curved or “s-shaped” and were compared to microtunnels described in another martian meteorite, Nakhla (Fisk et al., 2006), and to “bioerosion” textures from the in situ oceanic crust. There are two types of microtunnels: fine threadlike microtunnels that range from c. 100–200 nm in width and <1 to 4 μm in length, and a second larger type that are >0.5 μm wide. The much smaller average diameter, especially of the fine threadlike microtunnels, is significant, as this is an order of magnitude smaller than microtunnels reported from the recent in situ oceanic crust; this has been the subject of discussion as to whether this comparison is therefore valid (Seife et al., 2014; White et al., 2014b). We note, however, that the current microtunnel size range data come from volcanic glass of the upper oceanic crust and that there is scant size data available from olivine-hosted microtextures of the lower oceanic crust.
Our investigations of a sample of Yamato 000593 also found fine-scale alteration textures along iddingsite-filled fractures in the olivine (Fig. 6). These microtextures are so small that they require study by SEM-BSE and are typically 2–3 μm long, and the majority are less than 0.5 μm wide. They are straight to gently curved and sometimes taper toward their ends (Fig. 6). Investigation by FIB-SEM is required to determine their geometry in the third dimension and confirm whether all these microtextures are indeed tunnels or planar features. The microtextures are often parallel to one another, although not in all cases. Most commonly occur at high angles to the fractures, but examples have been seen at lower angles to the fractures (Fig. 6).
The iddingsite veins in Yamato 000593 are reported to contain indigenous organics, possibly kerogen that occurs as spheroidal structures less than 1 μm in diameter embedded in the iddingsite (Fig. 4 White et al., 2014a). The origin of these organics is unknown, and the authors postulate that it could be abiotic and exogenously sourced (e.g., from comets, asteroids, or interplanetary dust particles) or biogenic, in which case the organics may represent “fossilized microbes” and further work will be required to resolve these potential sources. Significantly, the spatial connection of these organics to the microtunnels is not yet clear and deserves further investigation by some of the techniques described above, for example, Raman spectroscopy, FIB-SEM, followed by TEM-EELS or nanoSIMS.
Considering the hierarchical biogenicity scheme proposed here, the Yamato features currently attain a Category 1 biosignature pending further study, given that (i) an early origin for the microtunnels is confirmed based on the cross-cutting fusion crust; (ii) the microtunnels are markedly different in size from terrestrial microtunnels from subseafloor dunnites, the significance of which is unknown; (iii) the microtextures need to be further investigated in the third dimension to confirm whether they are all tubular in cross section or some are more planar features (also they currently appear to lack ornament); and (iv) in situ geochemistry is lacking, particularly the relationship between the carbonaceous material and the microalteration textures (Supplementary Table S5). Therefore, the Yamato microtunnels currently score low on the hierarchical biogenicity scheme proposed here, that is, until further in situ, high-resolution imaging and geochemistry is undertaken.
6. Seeking a Category 5 Biosignature on Mars and Beyond
The possibility of a deep microbial biosphere on Mars is supported by widespread evidence for subsurface groundwater and hydrothermal fluid circulation (e.g., Ehlmann et al., 2011a; Michalski et al., 2013), the availability of potential electron donors and acceptors in the mafic-ultramafic crust, and redox gradients that could sustain life (e.g., Fisk and Giovannoni 1999; Varnes et al., 2003; Bishop et al., 2013a). There is now also abundant remote sensing data for low-temperature aqueous alteration in a range of environments on Mars that indicate the potential for production of clay minerals, phyllosilicates (e.g., Ehlmann et al., 2011b; Bishop et al., 2013a), and serpentine minerals (e.g., Bishop et al., 2013b). In addition, martian meteorites such as Nakhla and ALH84001 preserve evidence for near-surface alteration, including low-temperature clay minerals and carbonates, respectively, that formed under conditions that may have been viable for life (e.g., Bridge et al., 2001; Chatzitheodoridis et al., 2014). Thus, potential habitats for subsurface life are present on Mars and await proposed sample return missions (e.g., McLennan et al., 2012) to collect candidate biosignatures that will need to be subjected to rigorous biogenicity tests such as those proposed here before any robust claims for life in the martian subsurface can be made.
Grosch et al. (2014) explored a range of potential low-temperature subsurface alteration environments in the mafic-ultramafic crust on Mars that may preserve candidate textural and geochemical biosignatures (see Fig. 5 Grosch et al., 2014). This study showed how quantitative EMP mapping combined with chlorite thermodynamic modeling can be used to constrain the pressure, temperature, and fluid conditions recorded by altered mafic-ultramafic rocks that host candidate biosignatures, all of which would aid in elucidation of the preservational history of target microtextures and evaluation of their biogenicity. This approach could be applied, for example, to microtextures in altered metavolcanic glass or olivines. Microtunnels have been reported from olivines in two martian meteorites: Yamato 000593 (White et al., 2014a) as discussed above and Nakhla (Fisk et al., 2006). Microtunnels of both meteorites have been compared to those from subseafloor volcanic glasses, although neither study drew strong conclusions about the biogenicity of the microtunnels on the basis of the current data available, the microtunnels thereby ranking low on our hierarchical scheme. Before such microtunnels can be advanced as reliable biosignatures on Mars, better size distribution data are required for microtunnels in both terrestrial and meteorite-hosted olivines, and further in situ geochemical and isotopic evidence is required, particularly from the associated organics.
Post-impact hydrothermal systems have also been proposed as potential microbial habitats on Mars and elsewhere (e.g., Cockell, 2006; Parnell et al., 2010). Recent studies of enigmatic tubular microtextures in impact glasses of the Reis impact crater in Germany have led to the hypothesis that the low-temperature alteration of impact glasses can produce microtubular biosignatures comparable to those from the subseafloor (Sapers et al., 2014). However, we caution that the role of abiotic mechanisms in forming microtubular structures during impact-induced melting of basement rocks to form glass has not been fully explored. In addition, the syngenicity of the organics associated with the Reis microtubes has been brought into question, because an origin from the pre-impact terrestrial ecosystem has not been fully excluded (McLoughlin and Grosch, 2014; Sapers et al., 2014). Therefore, impact-generated glass (pseudotachylite) is not a reliable lithology for seeking biosignatures, as the rheological properties of impact glasses and their alteration histories are not fully understood. This setting should be distinguished, however, from post-impact hydrothermal alteration of fractured basement basaltic glass or ultramafics, which may be a viable setting for candidate biosignatures on Mars.
Candidate bacterial encrustations in secondary alteration minerals have not yet been reported from Mars; nonetheless, zeolites and palagonite formed by low-temperature alteration of basaltic glass are important targets for Mars sample return. A large ovoid microtexture composed of clays that is 80 by 60 μm across and described as “biomorphic” was recently reported by Chatzitheodoridis et al. (2014) from the Nakhla meteorite. This microtexture is indigenous to the meteorite, has a concentric wall ultrastructure comprising several clay mineral and Fe-oxyhydroxide layers, contains organics of uncertain syngenicity, and was interpreted as abiogenic in origin. This study is instructive for illustrating the application of a wide array of in situ geochemical techniques to investigate the origins of an intriguing microtexture. The favored abiotic scenario for this ovoid body was the infilling of a preexisting vesicle in the meteorite by sequential mineral precipitation directly from a fluid to form an amygdale (Fig. 16 Chatzitheodoridis et al., 2014). This is not therefore a candidate bacterial encrustation.
In summary, we hope that the application of the hierarchical biogenicity scheme (Table 1) will help confidently assess candidate microtextural biosignatures that may one day be returned from Mars and will help toward the discovery of a Category 5 biosignature. This effort will be strengthened by ongoing work to better understand processes related to microbial activity in, and on, terrestrial volcanic glass and by the development of further microbiological and geochemical imaging techniques together with laboratory growth experiments. The probabilistic scheme (Fig. 2) provides an integrated approach to evaluating the relative merits of candidate traces of life reported from mafic-ultramafic meteorites, the martian crust, and perhaps rocky moons in our solar system.
Footnotes
Acknowledgments
We thank the Japanese Polar Institute for loan of sample Yamato 000593. Also thanks to our numerous colleagues and collaborators with whom we have discussed topics of volcanic glass alteration, subseafloor microbial versus abiogenic processes, and the search for life in the basaltic crust on Mars. The work was supported by a grant from the Bergen Research Foundation and the University of Bergen to N.M., also by the Research Council of Norway via the Centre for Geobiology.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.