
Abstract
Extracellular UV-screening pigments gloeocapsin and scytonemin present in the exopolysaccharide (EPS) envelopes of extremophilic cyanobacteria of freshwater and marine environments were studied by Raman spectroscopy and compared to their intracellular photosynthetic pigments. This Raman spectral analysis of the extracellular pigment gloeocapsin showed that it shared Raman spectral signatures with parietin, a radiation-protective pigment known in lichens. The UV-light spectra also show similarities. Gloeocapsin occurs in some cyanobacterial species, mostly with exclusion of scytonemin, indicating that these pigments have evolved in cyanobacteria as separate protective strategies. Both gloeocapsin and scytonemin are widely and species-specifically distributed in different cyanobacterial genera and families. The widespread occurrence of these pigments may suggest an early origin, while their detection by Raman spectroscopy makes them potential biosignatures for cyanobacteria in the fossil record and demonstrates the usefulness of nondestructive Raman spectroscopy analyses for the search for complex organics, including possible photosynthetic pigments, if preservable in early Earth and extraterrestrial samples. Key Words: Raman microspectroscopy—Biosignatures—Cyanobacteria—Gloeocapsin—Parietin—Scytonemin. Astrobiology 15, 843–857.
1. Introduction
P
Extracellular pigments seem to occur in some form in 2-billion-year-old microfossils Eoentophysallis belcherensis preserved in silicified stromatolites (Golubic and Hofmann, 1976; Hofmann, 1976; Golubic and Abed, 2010, Fig. 6). Thus, the analysis with nondestructive Raman spectroscopy of pigments that protect microorganisms from radiation and their diagenetic derivatives is especially relevant to the studies of early life and potentially to exobiology (e.g., Jorge Villar and Edwards, 2006; Marshall and Olcott Marshall, 2010; Jehlička et al., 2014).
Our research provides Raman microspectroscopy and chemical mapping of live cyanobacteria and investigates the nature of the gloeocapsin and other photoprotective pigments known in cyanobacteria and lichens, such as scytonemin, parietin, chlorophyll, and carotenoids, which are detectable with Raman spectroscopy. The present study focuses on epiliths and endoliths that were exposed to direct solar radiation, extreme variation in temperature and salinity, and water shortage (see Ercegović, 1932; Jaag, 1945; Golubic, 1967a, 1967b; Hoffmann, 1986).
2. Materials and Methods
2.1. Environmental settings and sampling
Epilithic and endolithic biofilms dominated by cyanobacteria were collected from two different habitats characterized by extreme conditions, including exposure to damaging solar radiation. (1) The dark epilithic biofilms periodically wetted by freshwater seepage over a limestone cliff at Pont de Bonne, Hoyoux Valley, Modave, Belgium (50°28′27″N, 5°23′05″E) (Fig. 1), were collected in February, June, and October, 2014. (2) Limestone fragments with endolithic populations of cyanobacteria that actively penetrate carbonate rocks in the upper intertidal and supratidal ranges of the limestone coast of Dalmatia, Korčula Island, Croatia (42°57′48″N, 16°43′21″E), which are periodically exposed to seawater by tides and wave spray (Fig. 2), were collected in July, 2013 and 2014. Both investigated habitats are dominated by cyanobacteria, which respond to solar irradiation with production of extracellular UV-screening pigments embedded in their gelatinous EPS envelopes, the subject of our investigation. These cyanobacteria also carried the bulk of the primary production in the examined habitats.

Location map of the sampling site on a limestone cliff at Pont de Bonne, Hoyoux Valley, Modave, Belgium, from general to local scale. Note the difference in color when dry in June (center, left) and wet in October (center, right); see also the immediate red coloration taken by Gloeocapsa alpina when treated with acetic acid (lower left). (Color graphics available at

Location map of the sampling site of the supratidal euendolithic community on the limestone coast at Vela Luka in Dalmatia (Korčula Island), Croatia. Note the blue color of small depressions (lower left) reflecting the populations of Hormathonema and Solentia contrasted by dark brown well-drained surfaces where Hyella spp. prevailed. (Color graphics available at
2.2. Light microscopy
The samples from the freshwater epilithic biofilms from the Belgian limestone cliff were preserved cold, examined directly following collection by compound light microscopy (Leica DM LB2), and photodocumented (Fig. 3). The marine euendolithic samples were extracted by using dilute (3%) hydrochloric acid to remove the carbonate matrix and were then mounted on microscope slides and examined and photodocumented. Light microscopy was used to provide and control the preparations for Raman spectroscopy analyses of HCl-extracted cyanobacteria and in culture control.

Cyanobacteria with UV-screening extracellular pigments invested in their EPS envelopes. Scale bars are 10 μm long. (
2.3. Extraction of gloeocapsin pigment and spectrophotometry
About 5 g of dark epilithic biofilms dominated by Gloeocapsa alpina were collected on the limestone cliff at Pont de Bonne, air-dried, and powdered. Gloeocapsin cyanobacterial pigment was extracted for 60 min with 100 mL of 100% acetone at room temperature. The solution was filtered and evaporated until the formation of small dark crystals occurred. Crystals were dissolved in 10 mL of 100% acetone and separated into six subsamples with different concentrations. One drop of 10% hydrochloric acid was added to three of the subsamples to test for the presence of gloeocapsin by changing the coloration of the solution from dark blue to red in acidic conditions (Fig. 4A). Spectrophotometry (230–750 nm) on acidified and non-acidified preparations was performed with a TECAN Infinite M200 PRO plate reader in a UV-Star 96-well microplate (Greiner Bio-One, NC, USA) in triplicate preparations with a read interval of 9 nm.

Gloeocapsin pigment of Gloeocapsa alpina extracted with acetone. (
2.4. Raman microspectroscopy
Raman analyses (Fig. 5) were performed with a Renishaw inVia Raman spectrometer coupled to a Leica DM 2500 confocal microscope and an Ar-ion 50 mW monochromatic 514.5 nm laser source. Laser excitation was adjusted to an on-sample intensity of maximum ca. 2 mW (measured with a Coherent LaserCheck analyzer) and focused through 50× (numerical aperture = 0.83) and 100× (numerical aperture = 0.74) objectives. Acquisitions were obtained with an 1800 grooves/mm grating with an air-cooled (-70°C) 1024 × 256 pixel CCD array detector. This method enabled a 2000 cm−1 spectral detection range. Beam centering and Raman spectra calibration were performed daily on a Si-Al microprocessor chip with a characteristic Si Raman band at 520.5 cm−1. Spectra were manipulated within Wire 4.1 software. Point and static line-scanning analysis measurements were made in static recording mode (fixed at 1150 cm−1). A high-resolution dynamic line-scanning Raman mapping technique (Renishaw HR StreamLine mode) was also used to detect the type and the distribution of UV-protecting pigments (Fig. 6). The map obtained is a true hyperspectral mapping (4958 acquisitions). These images cover a 96.2 × 87.1 μm area (1.3 μm step). The total measurement time is 461 s. Raman mapping images constructed from the “intensity at point” mapping tool at each sample point are shown in Fig. 6. Spectra have been baseline corrected by the use of “through fixed point” fitting mode. No interpolation mode are used to generate the chemical map.
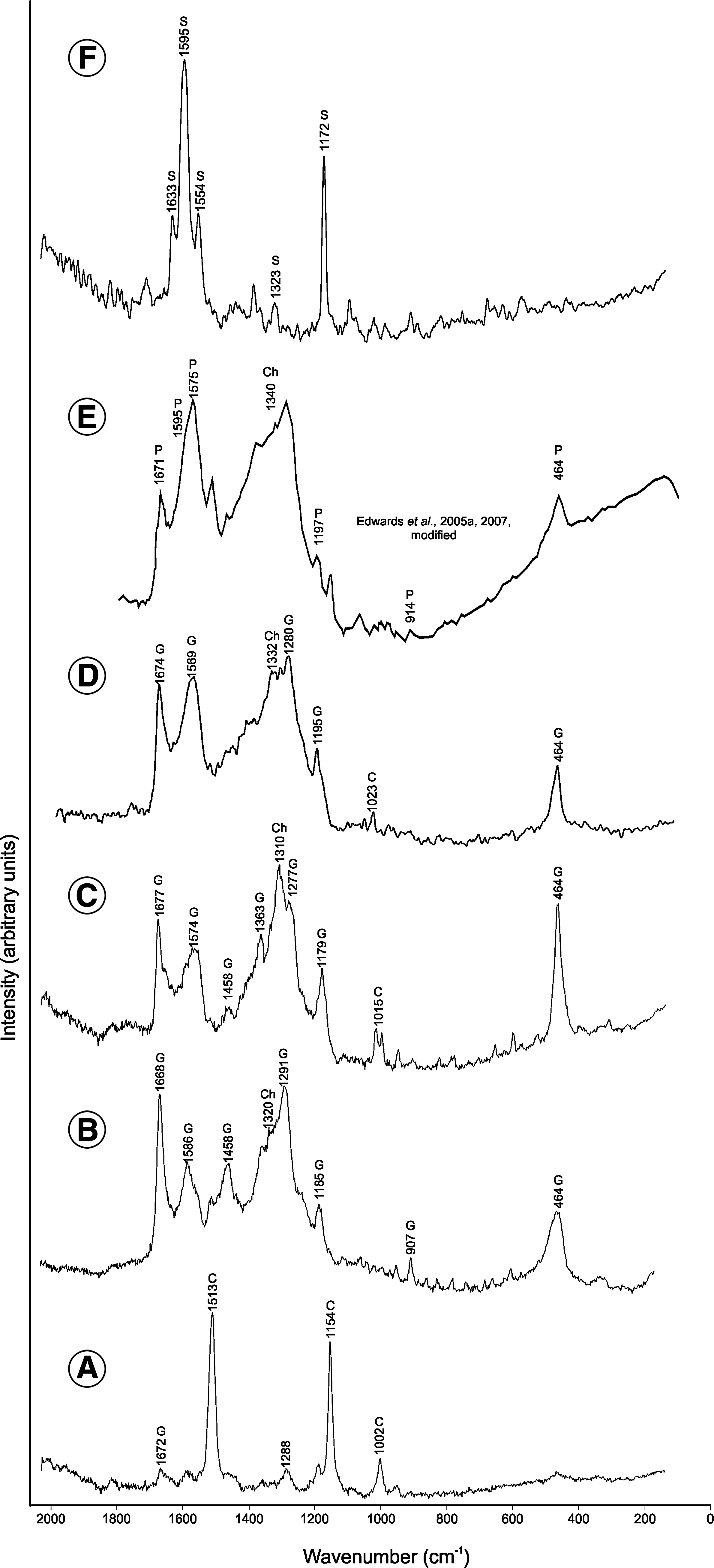
Confocal Raman spectra of cyanobacterial colonies. (

Confocal Raman StreamLine high-resolution mapping of a Gloeocapsa alpina colony. (
3. Results
3.1. Macroscopic appearance of collected biofilms
Soft epilithic biofilms collected from a limestone cliff at Pont de Bonne, Hoyoux Valley, Modave, Belgium, were of a loose crumbly consistency and were easily removed by scalpel. They were dark gray in color when dry and black when wet. They were distributed in vertical streaks along the water seepage down the cliff (Fig. 1, center left). The dark appearance of the biofilms could be changed in situ and in the laboratory to a bright red color by lowering the pH, for example, by adding acetic acid or dilute hydrochloric acid (Fig. 1, lower left), indicating the presence of gloeocapsin.
The marine euendolithic (Golubic et al., 1981) biofilms adhered firmly to hard carbonate rock surfaces and were distributed in zones parallel to the sea level or irregularly, depending on local relief with different water drainage. They were dominated by Solentia paulocellulare (Ercegović) Le Campion-Alsumard and Golubic and Hyella balani Lehmann, both with extracellular pigments. The habitat also showed distinction in coloration when dry versus wet, as illustrated by Ercegović (1932). Surfaces dominated by Hormathonema violaceao-nigrum and Solentia appeared bright blue when dry and dark blue to black when wet, whereas surfaces dominated by Hyella spp. were dark brown in color (Fig. 2, lower left).
3.2. Light-microscopy observations
The microscopic examination of the biofilm from the limestone cliff at Pont de Bonne, Hoyoux Valley, Modave, Belgium, was dominated by Gloeocapsa alpina (Nägeli) Brand ( = G. sanguinea Kützing, sensu Jaag) with colonial unicellular cells (probably Chroococcidiopsis sp.) as a minor component. Gloeocapsa alpina (Nägeli) Brand shows blue-green-colored cells encapsulated within multiple EPS envelopes, which were distinctly blue on the light-exposed surface and colorless in the shaded interior parts of the biofilms. The effect of shading could be observed within individual colonies (Fig. 3A). When exposed to dilute HCl to remove incorporated carbonate particles, the envelope color turned distinctly red (Fig. 3B). Similar pH-dependent reactions and color changes were observed in marine euendolithic biofilms for Hormathonema violaceo nigrum Ercegović, Solentia paulocellulare (Ercegović) Le Campion-Alsumard and Golubic and Solentia foveolarum Ercegović, when moved from the slightly alkaline seawater (Fig. 3C) to dilute HCl to dissolve the carbonate matrix (Fig. 3D). However, this response did not occur in H. luteo-brunneum Ercegović and Hyella balani Lehmann (Fig. 3E), which both retained the yellow-brown coloration of their sheaths, turning greenish only when acidified to a very low pH.
3.3. Absorption spectra of extracellular sun-screening pigments
The extracellular gloeocapsin pigment was extracted in acetone and purified and recrystallized. Gloeocapsin extractions in acetone were subsequently analyzed at different concentration and pH (Fig. 4A). The ultraviolet-visible light (UV-vis from 230 to 750 nm) absorbance spectra of gloeocapsin shows λmax at 392 nm and 410 nm at pH ∼7 and pH <5, respectively (Fig. 4B).
3.4. Raman microspectroscopic analyses
Several Raman spectra of cyanobacterial specimens were obtained with the 514.5 nm laser source, the most favorable wavelength excitation for the observation of biosignatures in cyanobacteria according to Edwards et al. (2005a) and Varnali and Edwards (2010, 2014). The characteristic Raman biosignatures of photopigments, Raman band position (cm−1), and the approximate assignment of vibrational modes obtained by different authors and at different laser wavelengths (514.5, 785, and 1064 nm) are shown in Table 1.
Vibrational assignment following Edwards et al., 1999, 2003, 2005b; Parab and Tomar, 2012; Jehlička et al., 2014; Fernandes et al., 2015.
ν: stretching; δ: in-plane bending; ρ: rocking.
In acidic conditions or during acid extraction, the extracellular gloeocapsin pigment changed its coloration from dark blue (or violet) to red (Figs. 1, 3B, and 4) (see also Radtke and Golubic, 2011, Fig. 1; Vincent, 2007). Raman spectra of single cells of Gloeocapsa alpina (Fig. 5A; Table 2) and associated extracellular pigments were explored at pH ∼7 (Fig. 5B; Table 2) and below pH 5 (Fig. 5C; Table 2). Specimens of Solentia paulocellulare, known to contain gloeocapsin (Radtke and Golubic, 2011), were also analyzed below pH 5 (Fig. 5D; Table 2). At pH ∼7, the spectra of the Gloeocapsa alpina cells show strong bands at 1513, 1154, and 1002 cm−1, which are assignable to carotenoids (Fig. 5A; Table 2). The spectra (in vivo) of the blue-violet-colored pigment, which correspond to gloeocapsin, exhibit bands at 1668, 1586, 1458, 1291, 1185, 907, and 464 cm−1 in neutral conditions (Fig. 5B; Table 2) and 1677, 1574, 1458, 1363, 1277, 1179, and 464 cm−1 in acidic conditions (Fig. 5C; Table 2). The broad and strong band centered about 1340 cm−1 could be assignable to chlorophyll (Edwards et al., 2005a; Tables 1 and 2). The extracellular location of the gloeocapsin pigment is confirmed by Raman StreamLine high-resolution mapping (4958 acquisitions) of a Gloecapsa alpina colony in acidic conditions (Fig. 6A). This mapping allowed for determination of the chemical distribution of organic and inorganic phases in a microscopic preparation, with a resolution of 0.83 μm (at 50×), including the spatial discrimination between intra- and extracellular pigments. Whereas the extracellular signal is assignable to gloeocapsin (Fig. 6B), the intracellular signal strongly corresponds to carotenoids (Fig. 6C) and chlorophyll (Fig. 6D). Band intensities at 1677 cm−1 (gloeocapsin; Raman band intensities at 1574 and 464 cm−1 generate an identical pigment distribution map—data not shown), 1154 cm−1 (carotenoids; Raman band intensities at 1513 and 1002 cm−1 generate an identical pigment distribution map—data not shown), and 1340 cm−1 (chlorophyll) were used to generate the Raman mapping. The spectra acquired on the extracellular pigment of a Solentia paulocellulare specimen, also known to contain extracellular gloeocapsin, gave Raman bands at 1674, 1569, 1280, 1195, 1023, and 464 cm−1 and a broad band at 1332 cm−1 assignable to chlorophyll (Fig. 5D; Table 2).
4. Discussion
4.1. Raman and photospectrometry characterization
Some of the bands attributed to the gloeocapsin pigment seem, however, also to be characteristic of parietin (Fig. 5F; Table 3; Edwards et al., 1998, 2005a, 2005b, 2005c, 2007). The similarity of the Raman signal for gloeocapsin and parietin in both Gloeocapsa and Solentia specimens confirms that the pigment gloeocapsin probably contains parietin, an anthraquinone component of a UV-protecting pigment type present in lichens, a major compound associated with chlorophyll. These data represent the first spectrochemical characterization of the UV-protective gloeocapsin pigment. The presence of chlorophyll with parietin was already observed by Edwards et al. (2005a) in some cyanobacteria (Gloeocapsa; Fig. 5E) of colonized selenite from the Haughton Crater (Canadian High Arctic). Parietin, an orange to red anthraquinone pigment (Edwards et al., 2003), which is known to be a biological accessory radiation-protective pigment, and β-carotene, have been previously identified in epilithic Antarctic lichens (Edwards et al., 2003, 2005a, 2005b, 2005c). In addition, Casanova Municchia et al. (2014) reported the presence of anthraquinone compounds from endolithic specimens from stone monuments and natural outcrops in temperate and Mediterranean climates.
Based on Edwards et al., 1998, 2003, 2004, 2005a, 2005b, 2005c, 2007; Jorge Villar et al., 2005, 2006; Jorge Villar and Edwards, 2006, 2013; Jehlička et al., 2014; Lepot et al., 2014; Casanova Municchia et al., 2014.
See Table 1 for references.
To differentiate between the blue to red gloeocapsin pigment and the yellow-brown scytonemin pigment, Raman spectra of Hyella balani, which are known to contain scytonemin, were also obtained (Fig. 5F; Tables 1 and 2). The Raman spectra, which exhibit bands at 1633, 1595, 1554, 1323, and 1172 cm−1 and correspond to the scytonemin signature, appear to be clearly different from the parietin/gloeocapsin Raman signature (Fig. 5B–5D). An interesting observation is the absence of a scytonemin signal in the Raman spectra of Gloeocapsa alpina and Solentia paulocellulare.
Since cyanobacteria that produce scytonemin and those that produce gloeocapsin co-occur in the same radiation-exposed environments, these UV-shielding compounds may, in fact, be considered alternative strategies. Ultraviolet-visible spectra of gloeocapsin were compared to the spectra of scytonemin (Garcia-Pichel and Castenholz, 1991), parietin (Solhaug and Gauslaa, 1996), and scytonemin-3a-imine (Grant and Louda, 2013) (Fig. 4B). Gloeocapsin at pH ∼7 and scytonemin show conspicuous λmax peaks, 392 and 384 nm, respectively, in the UVA range (315–400 nm; Gao and Garcia-Pichel, 2011), while gloeocapsin at pH <5, thus in acidic conditions, shows high absorbance in the UVA but with a λmax (410 nm) shifted in the visible part of the UV-vis spectrum like parietin (434 nm) and scytonemin-3a-imine (437 and 564 nm). It is interesting to note that, in both gloeocapsin spectra, a small peak appears in the region between 650 and 700 nm, which corresponds to the chlorophyll a absorbance signature as noticed earlier for sunscreening pigments (Gao and Garcia-Pichel, 2011).
4.2. UV-screening pigments in cyanobacteria, lichens, and plants
Color is the most distinctive property of the cyanobacterial extracellular pigments, gloeocapsin and scytonemin. As mentioned earlier, these names were derived from the cyanobacterial genera in which the pigments were first observed, and conversely, the extracellular pigments have been traditionally applied in cyanobacterial taxonomic criteria. For example, the genus Gloeocapsa has been divided into the subgenera Hyalocapsa, Rhodocapsa, Cyanocapsa, and Chrysocapsa, depending one whether the envelopes were colorless, red, blue, or yellow-brown (e.g., Geitler, 1932). Jaag (1940) reviewed earlier observations and suggested the unification of several species that were originally separated on the grounds of the shape and color of extracellular envelopes. He noted that the production of scytonemin and gloeocapsin was triggered in response to intense light exposure and that the shift between red and blue pigmentation depended on the pH of the surrounding waters. He also noted that, in habitats with severe water shortage, the envelopes served as a water reservoir that produced a variety of states depending on the cycle of water availability. However, the distinction between gloeocapsin (red and blue) and scytonemin (yellow-brown) in sheath pigmentation appeared constant and apparently genetically fixed; thus it justified the distinction between the species with the yellow-brown envelope pigment scytonemin (“Chrysocapsa”) and those containing gloeocapsin (Jaag and Gemsch, 1940), thus distinguishing, for example, Gloeocapsa kützingiana from G. sanguinea. Similar distinction based on the presence of extracellular pigments gloeocapsin versus scytonemin was observed in sheathed filamentous cyanobacteria, for example, in genus Schizothrix, section “Chromosiphon.” In general, scytonemin has a wider distribution among coccoid and filamentous cyanobacteria than does gloeocapsin (Geitler, 1932).
Parietin is the most common UV-protecting pigment in lichens; it occurs in a number of species that belong to four lichen families: Teloschistaceae, Brigantiaceae, Letroutiaceae, and Psoraceae (Hauck et al., 2009). Parietin has been studied extensively, including with Raman spectroscopy, in the bright yellow lichen Xanthoria parietina (from which the compound derived its name) and in the bright yellow–colored Caloplaca (Solhaug and Gauslaa 1996; Edwards et al., 1997, 1998; Gauslaa and Ustvedt, 2003). Parietin, and probably gloeocapsin, belong to a group of naturally occurring anthraquinones (9,10-dioxoanthracene) that are characterized by three condensed rings in a linear arrangement (Edwards et al., 2003).
Parietin has also been extracted from the roots of some plants, particularly those of the genus Rumex, which has been used in traditional Chinese medicine. Parietin does not seem to occur in animals, and in fact, it acts as a deterrent to grazing. When identified in fungi, it was considered secondary, and its origin was traced back to the fungal components of lichens. In fact, parietin appears to have an antibiotic effect against parasitic fungi (Gazzano et al., 2013). According to our results, the origins of biosynthesis of these compounds point to cyanobacteria, and their further distribution may have occurred via endosymbiotic incorporation of chloroplasts to plants and via lichens to fungi.
4.3. Role of radiation-protecting pigments in the survival of lithophytic extremophiles
Microbial colonization of bare rocky surfaces such as the exposed cliffs of various volcanic and sedimentary rocks can be observed on mountains in different parts of the world (e.g., Ercegović, 1925; Jaag, 1945; Friedmann et al., 1967; Golubic, 1967a, 1967b; Friedmann and Ocampo, 1976; Hoffmann, 1986; Büdel and Wessels, 1991; Büdel et al., 1994; Büdel, 1999). These organisms form differently pigmented epilithic biofilm communities (Bultel-Poncé et al., 2004) or penetrate carbonate and other soluble rocks as internal or euendolithic biofilms; they are among the first colonizer of newly exposed rock surfaces (Golubic and Schneider, 2003). These extremophilic microbial communities persist and survive for long periods of severe water shortage, excessive radiation impact, and intermittent nutrient shortages (Rothschild and Mancinelli, 2001), while being supported almost exclusively by cyanobacteria as primary producers.
Another set of extreme environments that are occupied by specialized microflora are the intertidal and supratidal (wave spray) ranges along the rocky sea coasts (Ercegović, 1932; Le Campion-Alsumard, 1966; Le Campion-Alsumard et al., 1996; Radtke et al., 1996; Radtke and Golubic, 2011), where inhabitants endure large fluctuations in salinity that range from freshwater to hypersaline (including mineral precipitation). Along limestone coasts, this microflora is dominated by cyanobacterial euendoliths that are engaged in bioerosion and landscape-shaping of geological significance (Schneider and Torunski, 1982; Tribollet et al., 2011).
The main strategy in these habitats appears to be survival rather than growth. The ability to slow down the physiological functions in response to water shortage and persist in a latent state for extended periods of time, as well as respond within a short period of time to achieve full phototrophic capacity upon wetting (Büdel and Lange, 1991), is an example of adaptations required for occupation of these habitats. Different species assemblages are arranged in zones parallel to water seepage and are controlled by supply and availability of water as it spreads from the seepage by capillarity (Rascher et al., 2003). The growth of cyanobacteria in these habitats proceeds at very slow rates (Le Campion-Alsumard, 1966, 1975), which poses difficulty in establishing cultures that usually become overgrown by opportunistic, fast-growing microorganisms (Palińska et al., 2012).
Another hazard these organisms face in these habitats is exposure to excessive radiation by UVA and UVB as well as by high-intensity visible light. These organisms, however, appear to be well adapted and protected by intracellular pigments, such as the carotenoids that are involved in the repair of radiation damage (Lakatos et al., 2001); the mycosporin-like amino acids (Garcia-Pichel et al., 1993); and most importantly extracellular deposition of radiation-shielding pigments (Rascher et al., 2003) such as scytonemin, gloeocapsin, and/or parietin.
4.4. Geological significance of gloeocapsin
Identifying cyanobacteria in the early fossil record requires unique biosignatures (Javaux and Lepot, 2015). Classically (e.g., Knoll and Golubic, 1992), such signatures were sought by using a combination of criteria such as unique morphology, cell dividing patterns, orientation of filaments toward light in the rock. These properties as well as the occurrence and distribution of microbial fossils in stromatolites were used to propose a cyanobacterial affinity. Microfossils of cyanobacteria occur through the geological record since the Paleoproterozoic and include silicified colonies and filaments of mat builders (such as Siphonophycus, Palaeolyngbya), probable akinetes (Archaeoellipsoides), probable planktonic colonies (such as Eomicrocystis), but also benthic and euendolithic clades (e.g., Polybessurus and Eohyella in the Neoproterozoic Draken Formation, Spitsbergen, as reviewed by Knoll and Golubic, 1992).
The oldest specimens containing presumed extracellular pigmentation occur in silicified stromatolites that grew in tidal and shallow subtidal waters on a carbonate platform of the 1.9 Ga Belcher Group, from the Belcher Islands, Canada (Golubic and Hofmann, 1976; Hofmann, 1976). These cherts contain oriented, three-dimensionally preserved colonies of coccoidal (spheroidal) microfossils, with multiple envelopes that are darkened toward the top of the colony and interpreted as pigmentation. These microbial fossils, named Eoentophysalis belcherensis (Hofmann, 1976), display binary cell fission in three planes, which permitted relating them to the living cyanobacterial genus Entophysalis, as it occurs today in hypersaline Hamelin Pool, Shark Bay, Australia (Golubic and Abed, 2010).
These microfossils, together with elongated vesicles (Archaeoellipsoides) interpreted as akinetes (cysts) of cyanobacteria (Golubic et al., 1995) from the 2.1 Ga Francevillian Series in Gabon (Amard and Bertrand-Sarfati, 1997), represent some of the oldest remains of identified cyanobacteria. Possible akinetes are preserved also in cherts from the 1.65 Ga McArthur Group (Tomitani et al., 2006). These microfossils demonstrate the diversification of modern taxa of benthic coccoidal and filamentous cyanobacteria that were capable of cell differentiation (producing specialized cells: heterocyst and akinete) in oxygenated carbonate platforms during the Paleoproterozoic.
Planktonic cyanobacteria have mostly simple unicellular spheroidal morphologies and may occur as isolated individuals or colonies, so it is difficult to prove or disprove their fossil occurrence without detailed geochemical and ultrastructural analyses. To strengthen paleobiological interpretations of microfossils, we have investigated the preservation potential of cell ultrastructures, EPS, and pigments that could provide a new range of taxonomically informative signatures of cyanobacteria. In a study of cyanobacteria modern and fossil (∼3 ka) mats from an Antarctic lake, we previously demonstrated the preservation of scytonemin in situ, that is, inside cyanobacterial sheaths detected by autofluorescence, as well as by ultrastructure of thylakoids on the thousand-year timescale (Lepot et al., 2014). This study provided novel insights into the earliest stages of fossilization of organic components (cells and extracellular polymeric substances) and revealed a variety of mineral nanostructures entombing cells.
In the present study, we characterized the nature of gloeocapsin and parietin pigment in endolithic and epilithic cyanobacteria from freshwater and marine environments and found it to be distinct from scytonemin with regard to properties and taxonomic distribution. Further work will test the preservation potential of this signature as it relates to the identification of cyanobacteria in the fossil record and will track the evolutionary history of their UV protection strategies, which are crucial for colonizing intertidal zones and continents in the Precambrian.
5. Conclusions
The pigments gloeocapsin and parietin share a number of properties, which include similar (but not identical) UV-vis photoprotective characteristics and Raman spectral signal. The Raman spectra clearly identify the presence of parietin or anthraquinone in gloeocapsin-colored cyanobacterial sheaths. However, as parietin does not show the same color changes with pH as gloeocapsin shows (Gauslaa and Ustvedt, 2003), it is possible that gloeocapsin is a derivative of parietin, but with some additional groups or conformational changes that remain undetected by Raman spectroscopy. We thus await a complete molecular characterization of the gloeocapsin pigment. The presence of gloeocapsin and similar parietin-like pigments in cyanobacterial extremophiles as compounds known to be effective in radiation screening, which are detectable by Raman spectroscopy, adds another molecule to the array of potential chemical fossils used to detect the presence of living organisms and has the potential for significant application in studies of past life and astrobiology.
Footnotes
Acknowledgments
We acknowledge support from the FRS-FNRS for a sabbatical leave to S. Golubic and for a mobility postdoc (R. FNRS.1692) to D. Velázquez, as well as from FP7 ERC Stg Elite for a postdoctoral fellowship to J.-Y. Storme, and from FP7 ERC Stg Elite, Belspo IAP Planet Topers and the Francqui Foundation to E.J. Javaux. A. Wilmotte is a research associate of the FRS-FNRS. We thank E. Mignolet for field assistance and M. Renard and J. Laval for technical support. We thank the two anonymous reviewers for comments that greatly improved this manuscript.
Author Disclosure Statement
No competing financial interests exist.