
Abstract
The ability to distinguish the features of a chemical sedimentary rock that can only be attributed to biology is a challenge relevant to both geobiology and astrobiology. This study aimed to test criteria for recognizing petrographically the biogenicity of microbially influenced fabrics and fossil microbes in complex Quaternary stalactitic carbonate rocks from Caerwys, UK. We found that the presence of carbonaceous microfossils, fabrics produced by the calcification of microbial filaments, and the asymmetrical development of tufa fabrics due to the more rapid growth of microbially influenced laminations could be recognized as biogenic features. Petrographic evidence also indicates that the development of “speleothem-like” laminae was related to episodes of growth interrupted by intervals of nondeposition and erosion. The lack of any biogenic characteristics in these laminae is consistent with their development as a result of variation in the physicochemical parameters that drive calcite precipitation from meteoric waters in such environmental settings. Key Words: Microfossil—Stromatolite—Biogenicity—Carbonate—Tufa—Speleothem. Astrobiology 15, 883–900.
1. Introduction
T
To test whether existing biogenicity criteria for microfossils and stromatolites could distinguish biotic from abiotic features in chemical sedimentary precipitates, we applied them to the potentially complex specimens that grow in the so-called “twilight zone” at the entrance to modern caves (see Jones, 2010), and in caverns of Quaternary tufa deposits. Carbonate rocks that formed in a twilight zone where there is a transition from tufa to speleothem carbonate rocks include the Holocene barrage tufa deposit located at Caerwys, north Wales, UK (Figs. 1 and 2; Pedley, 1987). This tufa deposit contains several meter-scale cavities or caverns that are locally decorated with speleothem-like stalactitic calcite. The cavities are a primary feature that formed behind “curtains” of petrified moss that draped over tufa dams known as “barrages” (Figs. 2 and 3). We examined whether characteristics of the fabrics of specimens that formed in these twilight zone conditions could be differentiated from those formed biotically under the influence of photosynthesizers (“tufa-like”) and those formed abiotically in the dark (“speleothem-like”). We also determined which petrographic criteria developed for distinguishing biotic from abiotic precipitates were most successful when applied to these Holocene specimens, with potential implications for early Earth studies and astrobiology searches.
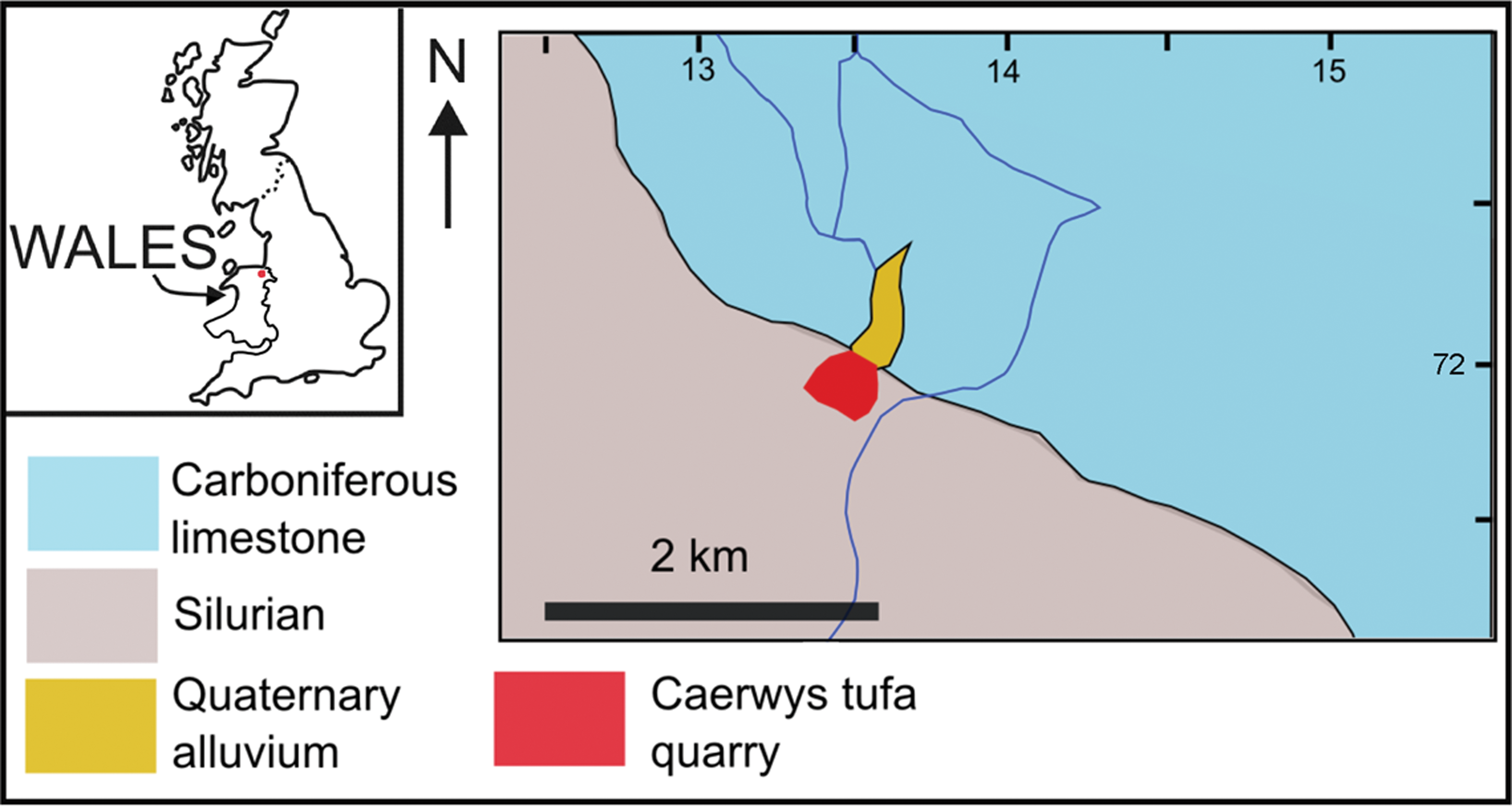
Map showing the location of the Caerwys tufa in the UK. Inset map shows the UK, with Wales labeled and Caerwys as a red dot. The larger map shows the location of the Caerwys tufa quarry (red) and its relationship to surrounding Quaternary alluvial sediments (orange) and underlying geology. Carboniferous limestone (blue) supplies the calcium and bicarbonate ions to groundwaters. These precipitate tufa calcite on emergence at springs associated with the contact between the limestone and underlying Silurian siliciclastic rocks (gray). Scale bar is 2 km. (Color graphics available at

Caerwys tufa quarry photographs. (

Active tufa stream at Alport, Derbyshire, UK, as an analogue for the Quaternary Caerwys tufa. (
1.1. Abiotic versus biotic origins of tufas and speleothems
Calcite precipitation in streams where tufas form was, until recently, most commonly viewed as a purely physicochemical process (e.g., Zhang et al., 2001) in which CO2 outgasses from fluids, causing them to become supersaturated with respect to calcite, as reflected in the net thermodynamic equilibrium equation
CO2 gas enrichment in meteoric waters occurs where they filter through biogenic CO2-rich soil zones. The partial pressure of CO2 builds in these environments; thus modern soil zone pCO2 commonly considerably exceeds that of atmospheric CO2. Calcite precipitation (Eq. 1) is typically associated with outgassing of this gas-rich fluid at spring effluents and where such fluids flow over obstacles like boulders or tree roots (e.g., Zhang et al., 2001). Recently, this model has been challenged by Hammer and coauthors (2010), who found that dissolved gas concentrations decrease very little by turbulence caused by flow over surface irregularities, which led them to conclude that the most important role of turbulence was to increase precipitation rates by bringing solutes to and from the calcite surface during turbulent mixing in the water column. Though the formation of stalactite and stalagmite speleothems in caves has most commonly been attributed to abiotic calcite precipitation driven by such physicochemical processes (e.g., Frisia and Borsato, 2010), field studies (Freytet and Verrecchia, 1998; Vazquez-Urbez et al., 2009; Glunk et al., 2011), and laboratory experiments with viable biofilms collected from modern stream settings (Rogerson et al., 2008; Pedley et al., 2009) has demonstrated that both abiotic and biotic processes influence calcite precipitation in streams.
Recent experiments and field studies have also demonstrated that tufa precipitation can be significantly enhanced, even driven, by microbial extracellular polymeric substances (EPS) (e.g., Rogerson et al., 2008; Glunk et al., 2011; Pedley, 2014). The EPS of nonviable organisms has also been implicated in tufa formation (Rogerson et al., 2008). Biofilms of some species of cyanobacteria are reportedly associated with their own specific microfabrics (Freytet and Verrecchia, 1998), and such petrographic evidence would support the hypothesis that some cyanobacteria exert direct control on tufa formation.
Diagenetic alteration following deposition adds further complexity to the ability to determine the biogenicity of precipitated carbonate rocks like tufas and speleothems. In spring or stream carbonate deposits, primary biofabrics may be rapidly overprinted and recrystallized, generating textures that appear abiotic (Jones and Peng, 2012). For example, diagenetic changes transform primary tufas from finely crystalline (micritic) to more coarsely crystalline (sparry) calcite, the fabric changing either by aggrading neomorphism (Love and Chafetz, 1988; Janssen et al., 1999) or simply continued growth of favored crystals (Brasier et al., 2011). Yet characterizing all coarse, sparry calcite fabrics as either alteration products or abiotic would be an over-simplification (Brasier et al., 2011) that could lead to many microbial fossils, both ancient and modern, being overlooked.
2. Materials
2.1. The Caerwys tufa
The tufa deposit studied here was found to the south of the village of Caerwys in north Wales, UK (Figs. 1 and 2). The waters that formed the tufa were sourced from a spring in the Carboniferous limestone (Pedley, 1987). Tufa precipitation at Caerwys has been constrained via radiocarbon techniques to the Holocene and pre-Holocene “Late Glacial” intervals (Preece and Turner, 1990). Deposition has now ceased, but a small analogous active site is found nearby at Ddol (Preece and Turner, 1990). The Caerwys tufa forms a veneer of Quaternary terrestrial carbonate rock over the underlying geology. It grew in a steep stream that flowed down the scarp-face of a Carboniferous limestone outcrop, with a series of cascading pools developing behind arcuate tufa dams or “barrages” that were oriented transverse to flow (Fig. 2; Pedley, 1987). The Caerwys system would have resembled the currently active tufa-depositing stream at Alport, Derbyshire, UK (Fig. 3). In both cases (Caerwys and Alport), unconsolidated micrite muds are found in the pools between the barrages. Some of these pools contain isolated decimeter-scale thrombolitic heads constructed by calcifying cyanobacteria, green algae, and invertebrates. The barrage dams themselves comprise indurated, but vuggy, carbonate walls that include a similar assemblage of fossil organisms. Though extensively quarried, the internal fabric of the barrage system is visible (Fig. 2) and preserved as a site of special scientific interest (SSSI).
Lithostratigraphy and mollusc biostratigraphy of the quarry were described by Preece (1978) and Preece et al. (1982), and a facies model was compiled by Pedley (1987). Tufa oxygen and carbon isotopes were detailed and their Quaternary climate implications discussed by Garnett et al. (2006). The latter estimated summer water temperatures were in the range of 13–16.5°C on the basis of tufa and gastropod δ18O values.
2.2. Specimens exhibiting the continuum of tufa and speleothem fabrics
Pedley (1990) described calcitic tubes of tufa formed by fringe cements that encrusted larger plants (macrophytes). The samples analyzed in this study fit that basic description, and such calcite encrustations of macrophytes are identified by their morphology. Internally, they have a central longitudinal cavity that may remain open or may have been filled by clastic grains (commonly, but not necessarily, of tufa carbonate) or calcite cement. They are differentiated from hollow abiotic stalactites such as “soda straws” in that they contain preserved carbonaceous matter or recognizable impressions of organisms that became encrusted in calcite. In swampy paludal marshes and pool margins, most plant stems grow upward and, when encrusted by carbonate, become stalagmite-like deposits (Pedley, 1990). In barrage systems, however, many plants including bryophytes live on the overhangs of the barrage (dam) front, their stems and branches hanging downward. This leads to pendant, stalactite-like calcite-encrusted structures (Fig. 2). Externally, these calcitic encrustations appear indistinguishable from “speleothem” stalactites or stalagmites formed in caverns. Encrustations may be <10 mm to several tens of centimeters in width, and range from a few centimeters to meters in length. Such encrustations are common but under-reported for modern karst settings. The oldest known examples may be the pendant cavity-filling cements of the 2.75 Ga Fortescue Group, Australia, which were interpreted as biogenic by Rasmussen et al. (2009).
3. Methodology
Eleven different tubular specimens from Caerwys, each several centimeters in length, were collected. All were examined in hand specimen and with standard petrographic microscopy techniques. Specimens were injected with blue resin, and both longitudinal and transverse thin-sections were prepared and analyzed. All rocks were found to be minimally luminescent or nonluminescent when viewed under UV light. Cathodoluminescence microscopy revealed that none of the specimens luminesced. Thin-sections were examined and photographed with the use of polarized light microscopy. Carbonaceous filaments entombed in calcite were distinguished from dark micritic calcite by partially dissolving the specimen with acetic acid. Similar petrographic histories were elucidated from all 11 specimens, such that we could select two representative specimens, labeled S3.2 and Caerwys 1, for detailed study.
4. Results
The descriptions of hand-specimen-scale observations are followed by microscopic observations for specimen S3.2, followed by the same for specimen Caerwys 1. The terms micrite (crystals <4 μm diameter), microspar (4–10 μm diameter), and spar (>10 μm diameter) are used to indicate crystal sizes and do not reflect the origin of the grains.
4.1. Specimen S3.2: hand-specimen-scale observations
Specimen S3.2 is a downward-tapering stalactitic rock (Fig. 4) that was collected as a float specimen that was found adjacent to the central barrage outcrop in the quarry (Fig. 2B, 2C). Internally, there is strong asymmetry, and at least six separate components of the visible fabric of the sample, referred to here as Zones 1–6, were delineated (Fig. 5).
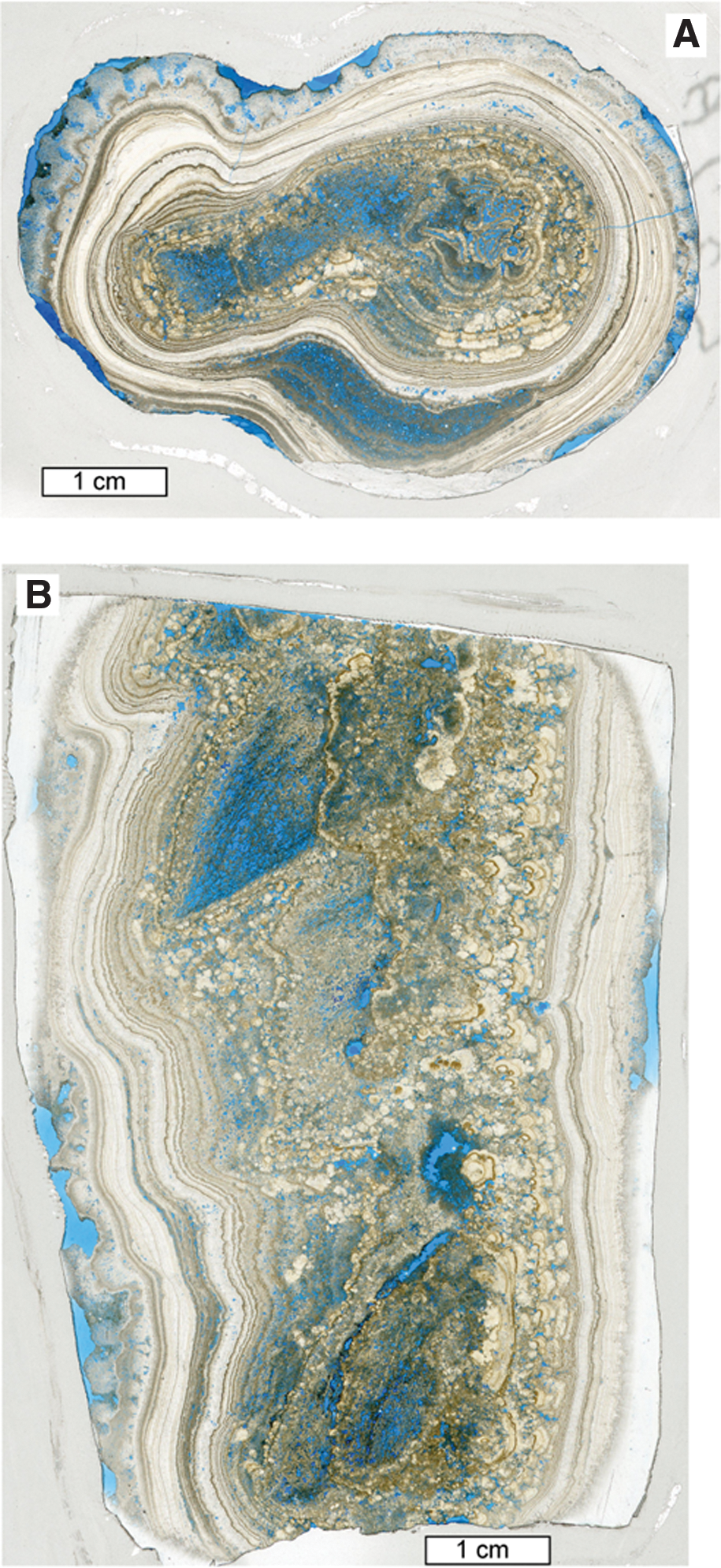
Scans of thin-sections of specimen S3.2. (
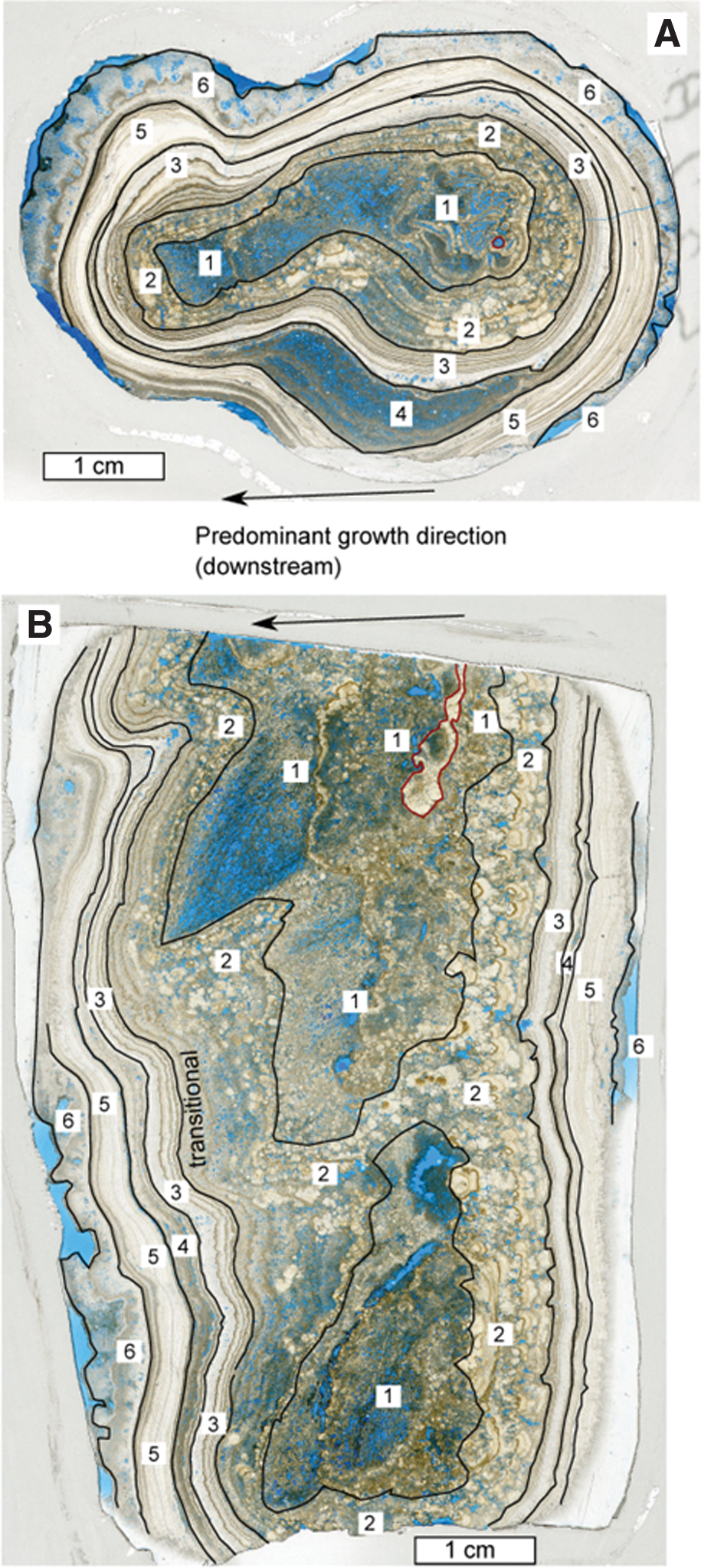
Interpreted scans of thin-sections of specimen S3.2. (
Zone 1 is located in the central section of the porous tufa, which is permeated by c. 50–100 μm diameter hollow tubes of calcite that form shrublike growths oriented downward at around 55 degrees from horizontal (Fig. 4B). From examination of longitudinal and transverse sections, it was possible to discern that these growths formed via micritic calcite encrustation of a biological (likely cyanobacterial) substratum. The asymmetrical nature of the sample, when viewed in cross section, is due to the more extensive outward growth of this primary depositional fabric in one direction away from the point of initiation. Denser-looking white patches of tufa, developed on one (likely the upstream) side of this sample, cover an area around 3 mm across and a centimeter in length.
Zone 2 (Fig. 5) consists of micrite intergrown with dark brown sparry calcite fans, with the latter being dominant. Each fan is around 1–3 mm across, and all fans grew toward the outward edge of the specimen. This zone forms a band around 1 cm thick on one side of the specimen (the side toward which Zone 1 micrite prograded) and is traceable continuously around the perimeter, though it narrows to <1 mm thick on the opposite side of the sample.
Zone 3 in hand specimen consists of a 1–2 mm thick light brown band that envelops calcite of Zone 2, and is capped by a darker brown 1 mm thick band. Internally, these bands contain several very fine submillimeter-thick cream-colored laminae. These bands are thicker where they developed below centimeter-scale overhangs.
Zone 4 is recognizable in hand specimen only as a white band that separates brown calcite below it from very similar-looking brown calcite above, which is designated as Zone 5. Note that Zone 4 appears brown in thin-section (Fig. 5).
Zone 6, the outer casing of the specimen, consists of a layer of cream-colored botryoidal calcite, around 2 mm thick on one side and 6 mm thick on the other. This outer zone seems to be divided into two bands by a very thin dark lamina.
4.2. Specimen S3.2: microscopy
Two thin-sections of sample S3.2 were made from the cut specimen. Scans of these are shown in Fig. 4, one from the transverse sawn section (S3.2A) and the other from the longitudinal sawn section (S3.2B). Petrographic analysis of these thin-sections confirmed that Zone 1 comprises 6.5–26 μm (mostly c. 20 μm) diameter dominantly subvertically oriented biological filaments that were encrusted by 18–50 μm thick walls of calcite (Figs. 4, 5, and 6A). The carbonaceous filaments themselves have mostly been oxidized, leaving behind empty calcite tubes (Figs. 6A and 7A), though some tubes contain carbonaceous material that resembles hollow and partially decomposed cyanobacterial trichomes within a sheath (Fig. 7B, 7C). The central parts of each calcite tube that would have been in contact with the carbonaceous filament are comprised of an envelope of small microcrystalline crystals. The remaining outer part of each calcite tube wall is constructed of radiating c. 20 μm diameter spar crystals (Fig. 7A). Also in Zone 1 is a cluster of hollow calcite tubes that range from 250 to 500 μm across (Fig. 7D). These each have an inner zone of microspar (crystals c. 20 μm diameter) followed by two rings of larger sparry crystals, each c. 45 μm thick. Some of these tubes have sparry calcite fan growths on one side which marks the start of Zone 2.

Micrographs of specimen S3.2 thin-section A (transverse cut). (

Micrographs of specimen S3.2 showing biological fabrics. (
The first Zone 2 sparry calcite fans (Figs. 4, 5, and 6A) nucleated directly on the spar of Zone 1 encrusted tubes. These two spar types are, in some locations, in optical continuity, which indicates that Zone 1 spar acted as a template for precipitation and growth of Zone 2 spar. Some curved, dark micritic growth lines within the fans are traceable between fans (Fig. 6A). Fans interfered with each other where they touched during growth. Some fans terminated in smooth curved surfaces; others were flat-topped (e.g., Fig. 8), and several exhibited pointed euhedral crystal terminations. Growth of Zone 2 seemingly ceased for extended periods of time (perhaps months?) on at least three occasions. These cessations are marked by dark micritic layers from which new fans nucleated. The latter are indicated by their different crystallographic orientations from the underlying fans (Fig. 6A). A magnified view of one of the Zone 2 fans, shown in Fig. 8, reveals that the carbonaceous filaments interpreted as entombed cyanobacteria are dominantly, but not exclusively, oriented subparallel to the direction of crystal growth. They are c. 2 μm wide and 40 μm long, distinctly narrower than the carbonaceous filaments of Zone 1. These narrower filaments cross through some of the finer, micrometer-scale crystal growth bands within the fans (Fig. 8). The micrometer-scale growth bands were likely formed on approximately diurnal timescales (Andrews and Brasier, 2005). Pedley (1987) suggested spar-entombed carbonaceous filaments of the Caerwys quarry could be Schizothrix or Phormidium sp. cyanobacterial fossils on the basis of growth form and diameter.

A sparry calcite fan containing entombed cyanobacterial filaments of specimen S3.2 Zone 2. Note the filaments are dominantly oriented parallel to the direction of crystal growth. However, they also curve and cross each other and are not artifacts related to the crystal structure. Filaments also cross dark-colored growth banding (likely formed on ∼diurnal timescales) in the crystal. Scale bar is 0.1 mm across. (Color graphics available at
The contact between Zones 2 and 3 of this specimen (Figs. 4, 5, 6, and 9) is sharp in places, though more transitional in others, due to the optical continuity of subsequent crystal growth. At its base, Zone 3 comprises numerous (at least six) couplets of micrite and columnar sparite (Fig. 9B). A micrite band c. 5 μm thick forms the nucleation region for numerous crystals of spar (mostly c. 100 μm diameter) in several locations in the specimen. Petrography revealed that several of the latter columnar spar crystals, commonly with length-to-width ratios >6, developed through competitive growth of crystals that nucleated in this band (e.g., Fig. 9C, 9D). Other columnar crystals stem from lower horizons that include Zone 2 spar crystals (e.g., Fig. 9A). Inclusions in the columnar crystals appear as bands oriented perpendicular to the direction of growth and parallel to (and commonly in close proximity to) the bands of micrite (e.g., Fig. 9C, 9D). The inclusions were identified primarily within the calcite crystals rather than between them.
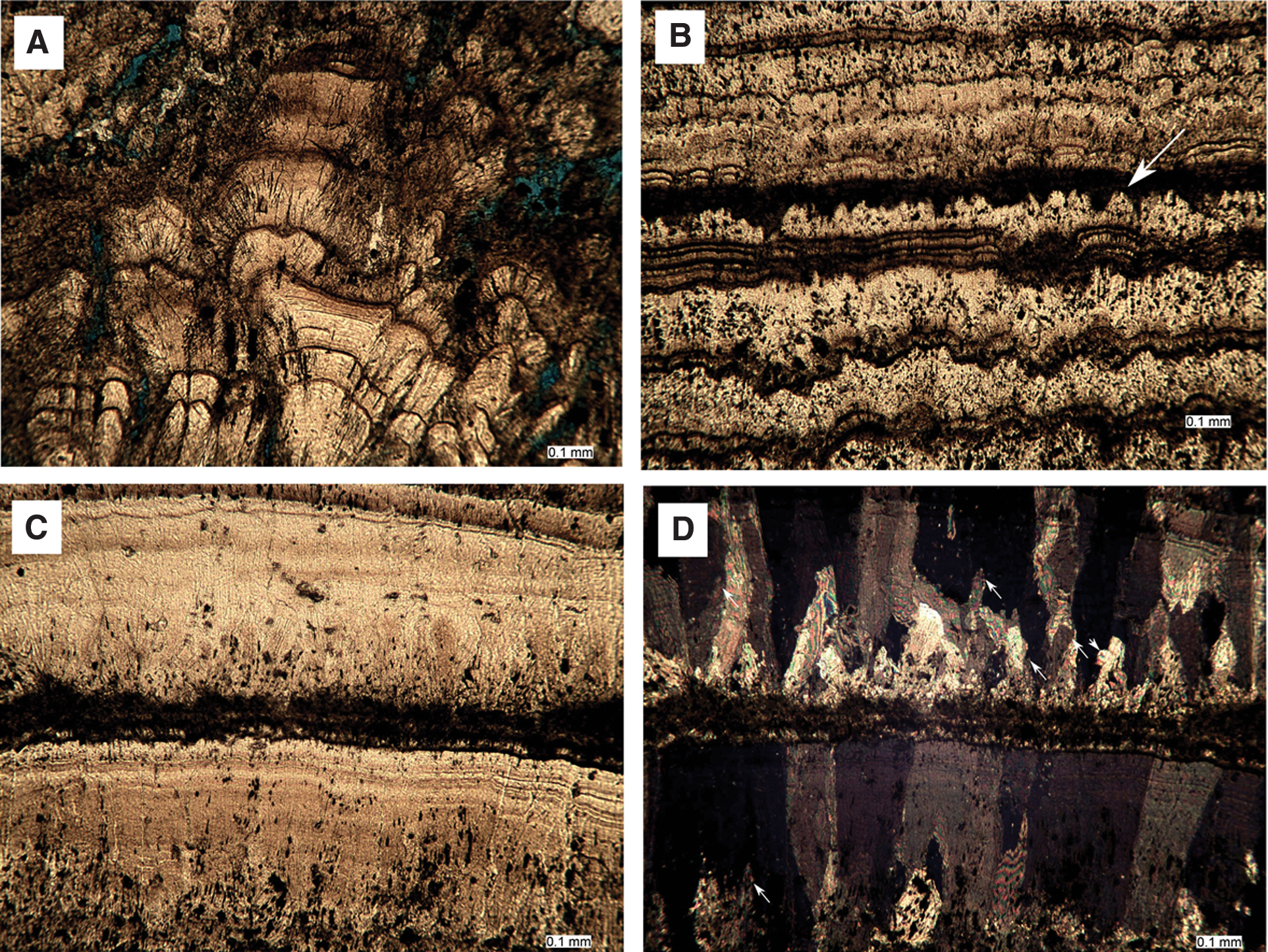
Sparry calcite of specimen S3.2 Zone 3. (
Most intriguing near the top of Zone 3 (Figs. 4, 5, and 10) is a patch a few millimeters across that includes a series of circular to oval pores that range from 100 to 240 μm in diameter (Fig. 10). The latter are most easily interpreted as transverse cross sections of hollow tubes. One tube oriented longitudinally in the thin-section extends for at least 2.2 mm. At higher magnification, it was determined that the walls of these tubes are thinly lined with dark micrite, surrounded by cylinders composed of spar crystals 100–400 μm in diameter (Fig. 10C, 10D). The latter are differently aligned from the columnar calcite host, radiating outward from the tube center. The crystals aligned parallel to the direction of growth of the columnar spar are elongated upward, forming flame-like growth shapes. These crystals evidently grew contemporaneously and in competition with the surrounding columnar spar.

Insect (likely chironomid) larval tubes in specimen S3.2 Zone 3 spar. These are seen as evidence that the spar is primary and not recrystallized from micrite. (
Zone 4 was recognized petrographically by the reappearance of micrite on one side of the specimen (Fig. 5B). This zone is laterally traceable into spar via two c. 5 μm thick subparallel micritic bands that mark its top and bottom. The micrite comprises a porous network of peloids that includes hollow tubes of c. 130 μm diameter.
Zone 5 spar crystal terminations range from pointed to flat or broken and are delineated by inclusions (see Figs. 5 and 11). In places, they are draped in continuous bands of dark, dustlike micrite that infilled the depressions and smoothed out the topography (Fig. 11A). Most spar crystals terminate at these micrite layers, though this was not always the case (Fig. 11B). For example, in one place a fragment of micrite 340 μm across adhered to the specimen surface (Fig. 11) and sits flat on a micritic layer. Spar crystals nucleated on this detrital micrite inclusion, though they were later out-competed by other columnar crystals. Further, the protrusion formed by this blob of micrite clearly affected development of the overlying layers (Fig. 11).

Caerwys specimen S3.2 Zone 5 in thin-section. (
The tops of Zone 5 columnar crystals formed the substrate for Zone 6 crystal growth. Zone 6 crystals (Figs. 5 and 12) are distinctly different from those of Zone 5 and appear as networks of needle-like laths. The latter are arranged along the faces of a crystal lattice (Fig. 12A, 12B). These laths are subcrystals (crystallites sensu Kendall and Broughton, 1978) that range individually in size up to c. 1 mm long and 50 μm wide and that link to form larger millimeter-to-centimeter-scale composite crystals (Frisia and Borsato, 2010). These composite crystals are sometimes in optical continuity with the Zone 5 columnar calcite on which they grew (Fig. 12).

Caerwys specimen S3.2 Zone 6 in thin-section. (
4.3. Specimen Caerwys 1: hand-specimen-scale observations
Caerwys 1 is an 8 cm long specimen that grew in a stalactitic fashion. It was collected from a cavern within in situ barrage deposits located in the center of the quarry. This specimen was collected from directly adjacent to the much larger stalactitic rock shown in Fig. 2C. In longitudinal section, it was possible to recognize the sample has a highly porous center surrounded by several c. 0.5–1 mm thick layers that alternate with much thinner (c. 100 μm thick), lighter-colored layers (Fig. 13). At least three layers were traced in the specimen, though in places the layers appear to be merged or truncated.
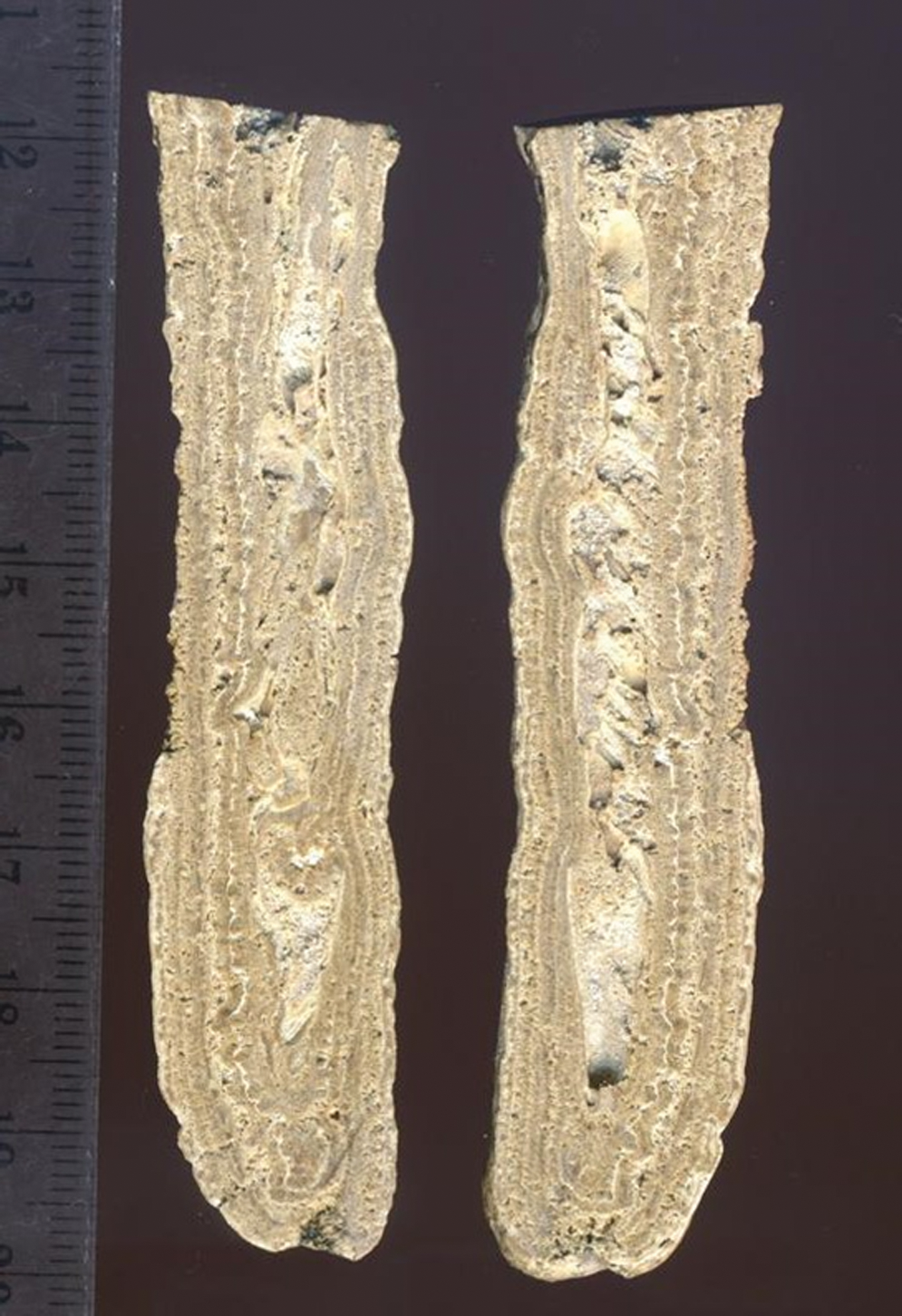
Stalactitic specimen Caerwys 1 (longitudinally cut hand specimen). The central cavity formed around a downward-hanging twig. Alternating micritic laminae (white) and sparry calcite fans (darker and thicker) grew on the outside. Ruler for scale (larger divisions are centimeters; smaller divisions are millimeters). (Color graphics available at
4.4. Specimen Caerwys 1: microscopy
A thin-section of Caerwys 1 was stained with Alizarin Red S and potassium ferricyanide. The pink color confirmed its nonferroan calcite composition. The central cavity (Fig. 13) of this specimen is highly porous, evidenced by the blue resin (Fig. 14), and divided into several empty pockets by walls of sparry calcite. One empty pocket is lined with 20 μm diameter microspar crystals (Fig. 14A) that are partially intergrown and form a porous network. On both sides of this initial cavity fill are crystal fans of sparry calcite, with crystal lengths of 80–150 μm. Adjacent to the sparry calcite fans at the top of the specimen is a second empty pocket, 3 mm across, that was progressively filled by 300 μm diameter sparry calcite fans (Fig. 14B). These grew inward from all sides toward the cavity center. Dark-colored inclusions within some of these fans are oriented along crystallite boundaries (Fig. 14C, 14D). The origin of these filamentous microfossil-like inclusions is discussed further below. The bulk of the specimen is comprised of layers of columnar sparry calcite that grew primarily outward as fans (Fig. 14E–14G). Each crystal is c. 500 μm long and 50–100 μm wide. These sparry layers correspond to the thick, lighter-colored layers observed in hand specimen that are separated by laminae of micrite c. 130 μm thick (Figs. 13 and 14F). There is evidence that columnar sparry calcite fans were partially dissolved prior to, or during, deposition of the micrite layers (Fig. 14E), as some of the micrite layers are a little thicker (c. 230 μm) and more porous, particularly one layer close to the outside of the specimen (Fig. 14G).

Photomicrographs of Caerwys 1 thin-section. (
5. Discussion
The criteria of Sugitani et al. (2007) and Wacey (2009) developed to establish the biogenicity of potential microbial fossils were applied to evaluate the biogenicity of the fossil-like objects in the two samples studied. For these specimens, their Quaternary age and sedimentary origins are not in doubt. These rocks have never been buried to any significant depth, so the characteristics of the precipitates and the fabrics were developed in the original sedimentary depositional environment.
5.1. Biotic versus abiotic growth of specimen S3.2
A complex growth history of specimen S3.2 was unraveled on the basis of the petrographic analysis, with different growth zones exhibiting different degrees of biotic influence. The oldest part of the specimen (in Zone 1; Fig. 5A, 5B) comprises dense, white-colored calcitic tufa that contains clusters of hollow carbonaceous tubes of fossilized cyanobacterial trichomes preserved inside sheaths. Zone 1 was formed by calcification of filamentous cyanobacterial shrubs that coated an overhanging leaf or twig. This photosynthesizing biofilm formed a substrate for subsequent growth that was necessarily focused away from the tufa wall toward the direction of light. Such biologically enhanced growth resulted in the asymmetrical, elongated form of the specimen. A network of 250–500 μm diameter hollow calcite tubes was formed by chironomid insect larvae, perhaps in the late spring season as waters warmed up (Janssen et al., 1999; Brasier et al., 2011). Filamentous microbes coated these tubes, separated by at least one and possibly two or three pauses of unknown duration (perhaps weeks or months, likely at most a few years). Zones 1 and 2 are at least part contemporaneous, with Zone 2 sparry calcite shrubs containing thinner entombed carbonaceous filaments dominantly forming on the side of the specimen that received less light. Their presence on the more illuminated side of the specimen toward the end of Zone 1 deposition, including in small crevices, is consistent with the onset of shaded, lower-light-intensity conditions that supported the growth of microbial species associated with spar formation.
The Zone 1 microfossils must have been syngenetic with the calcite deposition, as the calcite initially developed as tubes nucleated on the filaments (Figs. 4, 5, 6A, and 7). It is possible that the Zone 2 microfossils (Fig. 8) represent endoliths that bored into the tufa postdeposition, as they are oriented parallel to crystal growth planes and cut across the micron-scale growth lines within the sparry calcite crystal fans, features previously used as indicative of an endolithic habit (Knoll et al., 2013). However, the filaments in the Caerwys samples protrude across laminae that likely formed on diurnal timescales, well within the lifetime of entombed individual microorganisms. Further, endolithic organisms would also be expected to target earlier-formed tufa micrite as well as the crystal fans. The occurrence of the Zone 2 filaments close to the center of the specimen and not in the outer parts makes it unlikely that the cyanobacterial filaments preserved in Zone 2 were endoliths. More likely, and as Pedley (1987) inferred, the filaments were fossilized by syndepositional calcite entombment.
Evidence for microbial influence on the growth of Zones 1 and 2, consistent with the microfossil biogenicity criteria of Wacey (2009), includes
(1) Two different populations of carbonaceous filaments, each community associated with its own characteristic carbonate rock microfacies and not randomly distributed.
(2) Entombed clusters of hollow brown-colored carbonaceous filaments that are syngenetic with the carbonate rock. These microfossils include sheathed trichomes and are comparable in size and morphology with extant cyanobacteria.
(3) Calcitic molds of colonies of microbial filaments preserved in the carbonate rock, with calcification specifically on and around the filaments, consistent with microbial influence on crystal nucleation.
Zone 3 (Fig. 9) lacks the microfossils of Zones 1 and 2 (Figs. 6, 7, and 8). The thicker layering on the undersides of overhangs (Figs. 4B, 5B) is consistent with a dominantly abiotic growth process. However, as in Zone 1, the 100–240 μm diameter tubes (Fig. 10) are best interpreted as chironomid larval tubes (e.g., Brasier et al., 2011). Chironomid larvae are detrital feeders, which implies that Zone 3 formed in detritus-rich flowing stream water. Tubes at the top of Zone 3 could be directly associated with the micrite of Zone 4. This inference is based on the micritic peloids that could be of fecal origin and additional tubes of likely chironomid origin that occur in the Zone 4 micrite. This suggests that Zone 3 spar and Zone 4 micrite are partly contemporaneous, and indeed Zone 4 micrite merges laterally into sparry calcite.
Petrographic evidence that Zone 5 columnar spar is primary and not the result of recrystallization includes the aggradational fabrics that surround the micrite inclusion (Fig. 11). That the micrite inclusion sits flat on a thin micritic layer (Fig. 11A) is consistent with the latter representing a phase (perhaps a dry summer) during which little calcite crystal growth occurred. New columnar crystals nucleated on top of the micrite (Fig. 11B), presumably during a wetter phase. The resulting topography caused deflection of the subsequent growth laminae. Ultimately, the spar that nucleated on top of the micrite inclusion was out-competed by the surrounding larger columnar crystals (Fig. 11B). Columnar spar of Zone 5 likely formed within, or in very close proximity to, the active tufa-depositing stream. These conditions could have been found behind the downstream-facing accretionary surface of an actively accumulating tufa barrage. No obvious indicators of a biogenic cause for calcite deposition were identified in Zone 5. This does not necessarily imply that nucleation of Zone 5 spar was wholly abiotic; EPS produced by microbes were very likely present in the stream water, though there was no direct evidence that they contributed to the formation of Zone 5 spar.
Similar to the crystals of the Zone 5 spar, the crystals of Zone 6 also lacked carbonaceous microfossils (Fig. 12). The fabric of Zone 6 was reminiscent of the open dendritic speleothem texture described by Frisia and Borsato (2010). Such textures are known to form at cave entrances, in places subject to kinetic processes such as prolonged degassing phenomena. Such a scenario is consistent with Zone 6 developing as a late-stage cave speleothem-like meteoric growth within the cavern. Unlike the within-stream conditions that supported the growth of Zones 1–5, Zone 6 developed when the tufa stream system itself had become inactive.
In short, petrographic observations revealed that Zones 1 and 2 formed in the presence of microbes that included cyanobacteria. Zone 3 calcite growth was dominantly abiotic except for calcite tubes most likely constructed by chironomid larvae. Zone 4 is attributed to chironomid larvae that consumed microbes. There are no obvious preserved indicators of biological activity or control on calcite precipitation in Zones 5 and 6, and the latter formed in different environmental circumstances compared to those that supported the growth of Zones 1–5. Abundant petrographic evidence for syndepositional growth of columnar calcite spar is found in places with entombed cyanobacterial filaments (Zone 2), where crystal growth continued behind the aggrading outer surface layer (Fig. 6), where there is evidence of micrite bands capping spar layers and competitive crystal growth of spar crystals (Fig. 9), where insect larval tubes are found in sparry calcite layers (Fig. 10), and where detrital micrite stuck to the specimen surface and interfered with growth of overlying spar crystals (Fig. 11). There is no evidence for recrystallization that would have required dissolution. Despite the presence of cyanobacteria in Zones 1 and 2, the most clearly laminated (i.e., “stromatolitic”) zones are 3 and 5 (Figs. 4 and 5). The latter are the most cave speleothem-like and arguably abiotic sections. This lamination is discussed further below.
5.2. Biotic versus abiotic character of specimen Caerwys 1
Specimen Caerwys 1 is similar to specimen S3.2 in that it was nucleated on a downward-hanging biological substrate, overgrown by successive layers of sparry calcite that extend to the outside of the sample. Caerwys 1 is a thinner specimen than S3.2. This may be partly due to the absence of prograding cyanobacterial filaments in the core of Caerwys 1. Rather, the initial biological substrate seems to have been a stem or branch of a larger plant. The predominantly sparry calcite walls of Caerwys 1 are similar to Zone 2 of S3.2 (compare Figs. 6B and 8 with Fig. 14C–14F). Clusters of potential microfossil filaments in Zone 2 of specimen S3.2 (Fig. 8) and in the sparry calcite fans of Caerwys 1 (Fig. 14C) also share similar properties of size, coloring, and orientation parallel to the direction of crystal growth. However, the orientation of the dark-colored Caerwys 1 filaments was directly related to the orientation of the crystal structure. A comparison of the observations against the biogenicity criteria of Sugitani et al. (2007) and Wacey (2009) indicates that the Caerwys 1 filaments are much less convincing as microfossil candidates than those found in the S3.2 sample.
5.3. Biogenicity of the lamination?
The specimens described here are layered carbonate rocks that by some definitions could be identified as “tufa stromatolites” (Riding, 1991) or simply stromatolites (e.g., Semikhatov et al., 1979). Therefore, to evaluate whether the Caerwys tufa layering would be identified as biogenic when commonly used stromatolite biogenicity criteria are applied, we used the criteria of Buick et al. (1981) and Hofmann (2000) as critiqued in McLoughlin et al. (2013). A summary of the observations is given below.
The first group of biogenicity criteria pertain to the context of the lamination and include, for example, requirements that the structures are found in sedimentary or metasedimentary rocks and are synsedimentary with the deposit in which they are found. This is undoubtedly the case for these Quaternary tufas. Similarly, there is a criterion that brecciated mat chips should be found accumulated in depressions between convexly laminated mounds. Eroded chips of layered carbonate are found within the pool facies at Caerwys (Pedley, 1987). Such findings establish that the layered carbonate rocks are a primary sedimentary feature, but these contextual criteria do not differentiate biogenic from abiotic stromatolites.
The second group of biogenicity criteria pertain to the morphology of the lamination. A biogenic stromatolite should exhibit a preponderance of convex-upward structures; contain laminae that thicken over the crests of flexures; consist of laminations that are wavy, crinkled, or have several orders of curvature; and may be associated with thin, rolled-up fragments with coherent flexible laminae that can reasonably be interpreted as microbial mats. However, none of these criteria is itself diagnostic of a biogenic rock. Here we highlight, for example, that several tufa stromatolites of cyanobacterial origin exhibit isopachous layering (e.g., Janssen et al., 1999; Andrews and Brasier, 2005; Brasier et al., 2010, 2011). Spar layers in Caerwys tufa specimen S3.2 Zone 3 that might otherwise be interpreted as abiotic thicken over the crests of flexures, as do spar layers of Zone 5 (Fig. 11). Laminae of specimen Caerwys 1 seem thickest at the downward-pointing tip of the specimen (Fig. 13), which is consistent with the effects of gravity and hence abiotic growth. Likewise, the thin micritic layers of Caerwys 1 are crinkled and curved (Fig. 13), yet the petrographic observations suggest that these micrite bands are related to periods of exposure or nondeposition, as they cap spar fans that are partially dissolved or eroded (Fig. 14E). Despite being crinkled, these micrite laminae are unlikely to be biogenic.
The third group of biogenicity criteria are those that relate to the nature of microfossils and trace fossils within the lamination. Zones 1 and 2 of specimen S3.2 exhibit both entombed hollow carbonaceous filaments and calcitic external molds of filaments oriented perpendicular to the lamination, not parallel to it. Trace fossils of insect larvae are present within specimen S3.2 Zone 3. It is possible that these larvae were farming particular species of cyanobacteria above their tubes and that the cyanobacteria influenced crystal growth. Alternatively, the larvae might have produced their own growth-influencing organic substances (Brasier et al., 2011). However, there is no evidence for microbes directly causing the lamination of Zone 3. Microfossils are notably absent in the most distinctly laminated section, Zone 5.
Additional biogenicity criteria require that changes in the composition of microfossil assemblages should be accompanied by morphological changes in biogenic stromatolites. Where more than one microfossil assemblage is present in the samples studied, changes in micromorphology were found in these specimens. For example, the narrow filaments of specimen S3.2 Zone 2 are associated with sparry calcite fans (Fig. 8), whereas broader filaments in trichomes of Zone 1 are found in subvertically hanging tubes of calcite (Figs. 4, 5, 6, and 7).
The entombed microorganisms of specimen S3.2 are also organized and clustered in a fashion consistent with colonial photoautotrophic growth. We infer that their EPS did not simply bind sediment (e.g., Gerbersdorf and Wieprecht, 2014) but that it actively assisted calcite crystal nucleation (Rogerson et al., 2008; Glunk et al., 2011). This hypothesis is supported by the petrographic evidence for preferential growth of the carbonate rock toward the most light-illuminated direction. Insect larval tubes present in the Caerwys tufa (Fig. 10) would potentially meet a criterion that biogenic stromatolitic fossils must be organized in a manner that indicates trapping, binding, or precipitation of sediment, though whether the insect larvae actively trapped, bound, or precipitated the sediment themselves (e.g., Brasier et al., 2011) could not be discerned petrographically.
Though all the above criteria establish that the lamination is sedimentary in origin (Fig. 6A) and the cyanobacterial microfossils are consistent with evidence for photoautotroph-induced specimen growth, the cause of most of the lamination in the Caerwys specimens is related to alternating episodes of specimen growth, nondeposition, and erosion (e.g., Figs. 9D, 11, and 14D). Physicochemical parameters such as stream flow rates, pH, alkalinity, saturation, and temperature likely controlled the development of such lamination. Though microbially produced EPS may have exerted some influence on the laminae microstructure, direct evidence for microbial control on the lamination was absent.
In summary, the criteria consistent with biogenic features in the layered tufas at Caerwys are those that relate to fossils of the organisms themselves and not those of the lamination. Without the presence of microbial fossils, it would not be possible to identify the biogenicity of the laminae. Layering alone is not diagnostic of a biogenic structure in specimens like those characterized in this study.
6. Implications for Astrobiology
Carbonate rocks that may be targets in the search for martian microfossils were identified by Niles et al. (2013). As with Earth's deep-time stromatolites (McLoughlin et al., 2013), discriminating purely abiotic chemical sedimentary precipitates from biogenic rock structures of Mars will prove challenging. It is encouraging that simple criteria devised for ancient stromatolites and microfossils (Buick et al., 1981; Hofmann, 2000; Sugitani et al., 2007; Wacey, 2009) here enabled biogenic microfossils to be distinguished from abiogenic pseudofossils in a complex terrestrial case.
It is worth noting that recognition of microfossils requires detailed microscopic examination; definitively identifying biogenic structures in the examined Caerwys specimens would not have been possible at the outcrop or hand-specimen scales. Robotic exploration of Mars has already located potentially habitable streams (Grotzinger et al., 2014), structures consistent macroscopically with microbially induced sedimentary structures have been described (Noffke, 2015), and chemical signals consistent with martian life (Webster et al., 2015) have been reported. Potential lessons from the Caerwys quarry, however, are that the microbial structures visible under the microscope are not readily identifiable as biogenic at the hand-specimen scale, and the nature of the laminations were such that they could not be proven as biogenic at the hand-specimen or petrographic microscope scales.
7. Conclusions
The microfossils found in specimen S3.2 display at the petrographic scale the most convincing evidence for biogenicity: the layered stalactitic rock developed as a result of the calcification of filamentous cyanobacteria, and different populations of carbonaceous filaments were each associated with their own characteristic carbonate rock microfacies. The entombed clusters of hollow filamentous carbonaceous microfossils that included sheathed cyanobacterial trichomes demonstrated that the microfossil evidence is syngenetic with the carbonate rock. The influence of biology on the morphology of the deposit was identified by the strong asymmetry of the sample fabric; preferential growth on the most highly illuminated side of the barrage was associated with the presence of photoautotrophic microbes preserved as colonies of calcitic filament molds. Though some calcite spar fans in this sample formed in association with microbial filaments, the columnar spar that formed during a latter growth stage toward the outside of specimen S3.2 does not include entombed microfossils. In contrast to specimen S3.2, the filaments found in the specimen Caerwys 1 were associated with the crystal structure of the deposit, which according to the criteria of Sugitani et al. (2007) and Wacey (2009) indicate that these structures are less plausible microfossil candidates than those found in S3.2.
Petrographic evidence indicates that columnar spar is a primary fabric, and not due to secondary crystal growth. This spar lacks obvious microfossils and formed within the calcite-precipitating stream, but in poorly illuminated locations behind the barrage front. Lamination is related to episodes of growth interrupted by intervals of nondeposition and erosion. We interpret the petrographic evidence as indicative of lamination formation controlled by physicochemical parameters. Interestingly, we found that lamination in the samples studied was not an indicator of stromatolite biogenicity.
Footnotes
Acknowledgments
Natural Resources Wales and Steven Griffiths are thanked for access to Caerwys quarry and permission to work on the site. Sebastiaan Edelman and Thomas Logeman assisted with fieldwork and provided some of the field photographs. Bouke Lacet (Sedimentology laboratory, VU University Amsterdam) prepared the thin-sections. Three anonymous reviewers helped to sharpen the manuscript, and Sherry Cady provided valuable editorial advice and assistance. A.T.B. was inspired by Martin Brasier. He dedicates this manuscript to his father's memory.
Author Disclosure Statement
No competing interests exist.