
Abstract
Habitability is a widely used word in the geoscience, planetary science, and astrobiology literature, but what does it mean? In this review on habitability, we define it as the ability of an environment to support the activity of at least one known organism. We adopt a binary definition of “habitability” and a “habitable environment.” An environment either can or cannot sustain a given organism. However, environments such as entire planets might be capable of supporting more or less species diversity or biomass compared with that of Earth. A clarity in understanding habitability can be obtained by defining instantaneous habitability as the conditions at any given time in a given environment required to sustain the activity of at least one known organism, and continuous planetary habitability as the capacity of a planetary body to sustain habitable conditions on some areas of its surface or within its interior over geological timescales. We also distinguish between surface liquid water worlds (such as Earth) that can sustain liquid water on their surfaces and interior liquid water worlds, such as icy moons and terrestrial-type rocky planets with liquid water only in their interiors. This distinction is important since, while the former can potentially sustain habitable conditions for oxygenic photosynthesis that leads to the rise of atmospheric oxygen and potentially complex multicellularity and intelligence over geological timescales, the latter are unlikely to. Habitable environments do not need to contain life. Although the decoupling of habitability and the presence of life may be rare on Earth, it may be important for understanding the habitability of other planetary bodies. Key Words: Habitability—Exoplanets—Habitat—Niche—Mars—Exoplanets. Astrobiology 16, 89–117.
1. Introduction
T
Astrobiologists have attempted to understand habitability and catalogue the requirements for its presence (Kasting and Catling, 2003; Gaidos et al., 2005; Nisbet et al., 2007; Zahnle et al., 2007; Lammer et al., 2009; Westall et al., 2013; Cockell, 2014a; Jaumann et al., 2014). Their focus has been on defining the basic requirements for life to be metabolically active or to reproduce in planetary environments and, in particular, describe processes that might be required for these conditions to be sustained over geological periods within the lifetimes of planetary bodies.
Efforts have also been made by ecologists to define what constitutes a habitat (e.g., Odum, 1971; Whittaker et al., 1973; Block and Brennan, 1993; Hall et al., 1997). This effort has been dominated by the study of multicellular organisms. Ecologists' motivation stems from an interest in advancing our knowledge of the interrelationships of life on Earth, improving our understanding of wildlife ecology, and enhancing our capacity to protect and preserve habitats.
Despite these parallel lines of thinking, there have been few attempts to synthesize them into a consistent view of habitability. This is unfortunate because the astrobiological view of habitability, which extends thinking more forcefully to microbial life and brings an extraterrestrial perspective to bear on the understanding of habitability at the planetary scale, has much to offer in helping to define the concepts of habitat and habitability. In a similar vein, well-established ecological thought has much to offer in refining astrobiological thinking, preventing a potentially wasteful reconsideration of long-debated topics and ensuring that a set of ideas and definitions emerges that is consistent across multiple scientific disciplines.
In this paper, to mark the 15th anniversary of the journal Astrobiology, we provide a review of habitability from an astrobiological perspective. This review has two purposes. First, it is an attempt to provide clarity in definitions and ideas using both astrobiological and ecological concepts. Second, it is a primer for anyone entering, or already in, the field of astrobiology who would value a review of this pervasive but often enigmatic term.
2. What Is Habitability?
We begin by proposing a working definition of habitability that we use to inform the subsequent discussions in this paper. Definitions are important for ensuring consistency in ideas and thus encouraging clarity in formulating scientific questions. However, they are, nevertheless, human constructs, and other authors are free to challenge, change, or ignore this definition.
The word habitability is derived from the Latin verb, habitare, “to live or dwell.” Habitat, which means, “it lives/dwells,” is the third-person singular, and habitability means a place in which “it” has the ability to live/dwell. The devil, however, is in the detail. What do we mean by “live”?
The definition of habitat that we adopt here is “an environment capable of supporting the activity of at least one known organism,” where “activity” (and thus “living”) is metabolic activity allowing for survival, maintenance, growth, or reproduction. This definition is binary. An environment either can or cannot sustain a given organism.
By “survival” we mean that life is able to use resources in the local environment to actively maintain a state of dormancy where the rate of molecular repair is at least equal to the rate of molecular damage (Price and Sowers, 2004). “Maintenance” means activity where a cell can carry out a range of other cell functions but has insufficient resources or energy to reproduce. “Growth” means increasing body biomass in multicellular organisms, but it is used synonymously with “reproduction” when applied to microorganisms. “Reproduction” is multiplication of an organism.
In astrobiology, an assessment is often performed as to whether an environment is habitable for “life” in general. By this we mean that a given environment can support “at least one known organism.”
The definition we adopt explicitly defines habitability with respect to a known life-form. Thus, habitability, as defined, is a conservative term, bounded by the current state of knowledge in biology. One may be able to assess certain requirements for the habitability of an environment independently of known life, such as energy availability. If we make the reasonable assumption that the Periodic Table is universal, then we can calculate the Gibbs free energy available in a given reduction-oxidation reaction or the energy available in light from a star and thereby assess whether an environment could theoretically support any organism, known or unknown, given certain assumptions about plausible energy demands (Hoehler, 2007). Some other speculative energy sources have been suggested, including gravitational, magnetic, kinetic, thermal, and radioactive energy; osmotic, ionic, or pressure gradients; and tectonic stress (Schulze-Makuch and Irwin, 2004). We do not know whether terrestrial life represents a universal norm (Pace, 2001; Bains, 2004; Benner et al., 2004). However, by constraining habitability to known life, we avoid the term becoming inextricably linked to the problem of defining life (Cleland and Chyba, 2002; Benner, 2010) or becoming defined by speculative capacities of unknown organisms.
To further explain why we adopt this working definition, it is useful to explore the existing ecological literature, since the problem of what constitutes a habitat and a habitable environment is not a new question. It has vexed ecologists for decades. Odum (1971) originally referred to a habitat as the “address” of an organism, its physical location. Although this unambiguously makes it clear that habitat is a physical space, it raises the question of what an organism must be doing for the space to count as an “address.” Whittaker et al. (1973) defined a habitat as the “range of environments or communities over which a species occurs,” although this usage leaves open the question of what the organism must be doing in a habitat for the word “occurs” to apply (merely being present there seems too wide a definition, as explained shortly).
In an attempt to provide clarity, Block and Brennan (1993) described habitat to be “the subset of physical environmental factors that a species requires for its survival and reproduction.” Hall et al. (1997) similarly defined a habitat as “the resources and conditions present in an area that produce occupancy—including survival and reproduction—by a given organism,” a definition reiterated by Krausman (1999). They go on to remark, “wherever an organism is provided with resources that allow it to survive, that is habitat.” These definitions are broadly consistent with ours, particularly when applied to multicellular life. To be minimally “surviving,” most multicellular organisms must be metabolically active. This is also the case for dormant vegetative microbial cells, which must have a minimum state of metabolic activity to repair macromolecular damage against molecular degradation (Price and Sowers, 2004). However, these definitions raise a problem in microbiology and astrobiology when microbial spores are considered, a particular type of survival state that is generally metabolically inactive.
To illustrate the problem, consider a thought experiment. Some bacterial spores are dropped deep into the nitrogen ices on Pluto, in some sort of drilling or penetrator mission. As they are protected from ionizing and UV radiation, they have a good chance to remain viable in the frozen ice. For a period of time they are surviving. Yet if mere survival of metabolically inactive spores is a sufficient requisite for a place to be a habitat in accordance with the definition of Hall et al. (1997), then Pluto has become a habitat for those spores and, by extension, Pluto is a habitable planetary body (it provides resources—cool temperatures and protection from radiation under the ice—for survival of the spores). This seems to be too wide a definition, since a habitat, and therefore a habitable environment, would then include any environment in the Universe where inactive bacterial spores could survive for some defined, and potentially very short (even seconds), time. Therefore, our working definition of a habitat has a minimum requirement for the environment to be able to support the metabolic activity of organisms.
Another end-member problem to address is the matter of reproduction. Must a habitat support reproduction? Many definitions of habitat include reproduction either as a possible or required characteristic of a habitat (e.g., the definitions of Block and Brennan, 1993, and Hall et al., 1997, as above). Aarts et al. (2013) defined a habitat as “a collection of resources and environmental conditions (abiotic and biotic) that determine the presence, survival, and reproduction of a population.”
If a habitat cannot sustain reproduction, then the organism (apart from a theoretical microbe that merely repairs itself indefinitely) has a limited future. Thus, there might be the inclination to include reproduction for a place to be habitable. To highlight the problem that lies in requiring reproduction to identify a place as habitable, another analogy is useful. The rabbit that visits the lead author's garden in Edinburgh eats grass, but it does not reproduce there. The garden is of insufficient size to host a rabbit warren. The rabbit must return to the hills outside Edinburgh to reproduce. However, the rabbit is using the garden to gain resources for growth and potentially for later reproduction. To require that a habitat should allow for reproduction would be to say that the garden is not a habitat for the rabbit and, by definition, it is uninhabitable to the rabbit. This seems too narrow a definition, since an area of a planetary surface that contains actively metabolizing life, but where reproduction is not occurring, would be classified as uninhabitable space. Hall et al. (1997) resolve this problem in stating, “Thus, migration and dispersal corridors and the land that animals occupy during breeding and non-breeding seasons are habitat.” As such, we consider a place that can support the metabolic activity of an organism, even if it does not reproduce in that place, to be a habitat. An obviously important caveat is that, if an organism does not reproduce in a habitat, it must have other habitats available to it during its life cycle that do allow for reproduction if its population is to persist.
In summary, the definition provided in this paper is broadly consistent with earlier definitions of habitat. However, we specify that “survival” must entail metabolic activity and that reproduction is a possible, but not a necessary, activity for an environment to be a habitat to an organism and therefore for a place to be habitable. What about a set of habitats that allow for an organism to be active but none of which allow for reproduction? These environments would be described as habitable to the organism, but clearly a population introduced into them will eventually go extinct. They sustain only instantaneous habitability but not long-term (continuous planetary) habitability according to the definitions we describe shortly.
Within the definition adopted here, a Special Region, as used for planetary protection concerns on Mars (Rummel et al., 2014) is a specific type of habitat in which reproduction (propagation) can occur.
Life itself can influence habitability by making physical space within which given organisms can be active. On a densely inhabited planet, biota may be habitat space. For example, the habitat of a bird species might be a particular forest, since the trees and their branches constitute physical space that defines the geographical distribution of the species (Sieving et al., 1996). Similarly, the layers of a microbial mat are habitat space for the organisms within them (Engel et al., 2003). However, on many planets, for example Mars, most habitat space, if it exists, is delimited by geological substrates (Southam et al., 2007).
Life can change the habitable space for other life-forms (Block and Brennan, 1993). As organisms weather rocks, cycle gases, and change the valence states of elements, for example, they change the potential for an environment to be habitable for other life-forms (Falkowski et al., 2008). On a densely inhabited planet, life becomes inextricably woven into defining the conditions for habitability for particular organisms. An example of how life can create new habitat for other types of life is successional changes in biota, where one type of organism, for example phototrophs or nitrogen-fixing bacteria on early lava flows, provides nutrients and conditions required for subsequent organisms (such as higher plants, Clarkson, 1997). An example of how life can cause deleterious conditions to habitability would be the large-scale oxygenation of a planetary atmosphere that causes the loss of habitable conditions for anaerobic organisms on the surface (Stolper et al., 2010).
The conditions required to produce organic molecules needed for an origin of life may occur in much wider conditions than those required for habitability. For example, temperatures in impacts or spark discharge required to produce amino acids from other compounds is likely to be higher than the maximal temperatures for microbial growth. However, the first organism to reproduce (and originate) must emerge in a habitable space. The conditions required for life to originate are a subset of the conditions for habitability. Abiogenesis therefore is likely to pass through a habitability bottleneck (Fig. 1).
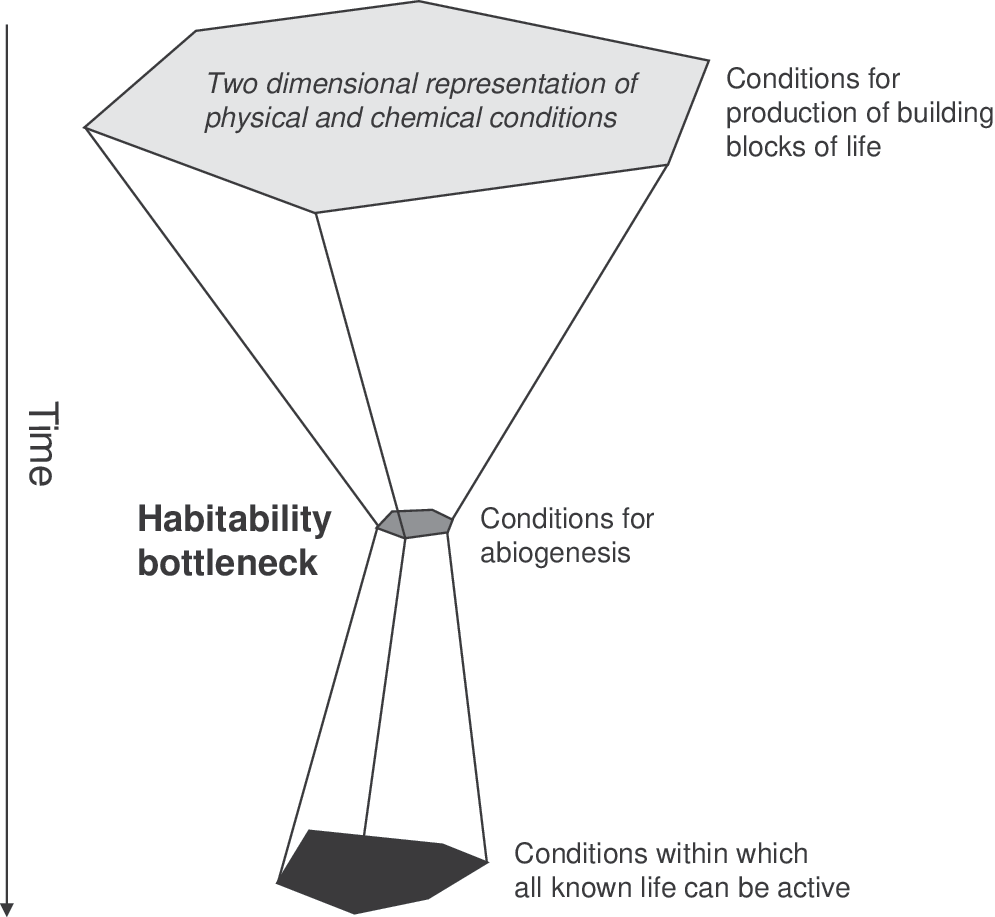
The physical and chemical conditions required to produce the building blocks of life are broader than the conditions for abiogenesis, which are presumed to be narrower than the total physical and chemical space that can be occupied by all life.
2.1. Habitability and niches
An important concept embedded within the conditions for habitability is the niche. Sometimes the word is used to mean a physical space, in other words as a synonym for “habitat.” However, this is not correct.
The ecological literature contains diverse opinions on exactly how the niche is defined. One approach is to consider the niche as the role that the organism occupies within a community (a group of interacting species within a given locality), including its functional role. This is consistent with Odum's view that, if the habitat is the “address” of the organism, the niche is the “profession” of an organism (Odum, 1971). The niche as a functional definition of the relationship of an organism to the rest of the biotic community was one adopted originally by Elton (1927)—the Eltonian niche.
A classic definition was provided by Hutchinson (1957). He considered the niche to be an n-dimensional abstract hyperspace made up of environmental conditions and resources that define the requirements of a known individual or a species to practice its mode of life. An important distinction in his concept is the “fundamental” niche, which is the niche that an organism can theoretically occupy, and the “realized” niche, which is the niche realized in practice. The former is more important for the theoretical assessment of an environment as being habitable.
Another type of niche was defined by Grinnell (1917). He considered it to be both the habitat that a species occupies and the adaptations that allow it to be successful in that environment (Grinnell, 1917)—the Grinnellian niche. The Grinnellian niche, in contrast to the Eltonian niche, emphasizes the way in which an organism uses resources to live in an environment rather than its functional role in the community (Devictor et al., 2010).
In astrobiology, the habitability of an environment is the capacity of a particular physical space to support the activity of an organism, that is, to provide the set of resources and conditions required for its way of life. Therefore, when astrobiologists speak about a habitable environment, their meaning conforms most closely to the Grinnellian niche concept. The Eltonian niche concept, at the current time, is less relevant because we have not found extraterrestrial biotic communities to be concerned about an organism's functional role in that community.
For microorganisms, their primary functions in the biosphere are usually linked to their mode of energy acquisition [e.g., using redox couples that mediate the biogeochemical cycling of elements such as iron, sulfur, carbon, and others through different valence states (Falkowski et al., 2008)]. Therefore, the niche is strongly linked to features of energy acquisition. By contrast, as most multicellular organisms use a single type of energy acquisition—aerobic respiration—it is not surprising that the existing ecological literature on niches has tended to focus on other functional roles of organisms vis-à-vis their behavior (foraging, competition, predation, etc).
The differences between niche definitions have also caused considerable controversy about whether a vacant niche can exist. The Eltonian niche considers an organism's place in a community in some sense independently of the organism itself, so that if a sulfate-reducing microorganism, for example, became extinct, it has been thought permissible to discuss the niche as being vacant to another sulfate-reducer. Similarly, the Grinnellian definition of a niche, as it includes the concept of the physical space or habitat, might also allow for a vacant niche, because it can be considered similar to a physical habitat that contains resources that are vacant for a species to use. However, Hutchinson's abstract definition of a niche focuses on a given species and defines its relationship to its environment and community. In this sense, a species is required to define a niche in the first place, making a vacant niche a logical impossibility.
The concept of a vacant niche is found in many instances in the ecological literature. If we allow a niche to be a set of known interactions and conditions that can provide the resources required for a given form of life, then we could plausibly identify a niche as being vacant, a view adopted by many ecologists (Colwell, 1992). An extreme end-member example of importance to astrobiology is a physical space that contains no life but can be shown to contain all the resources and have physical and chemical conditions required for a known organism to be active in that space. Such a place is an uninhabited habitat that contains a niche (or niches).
2.2. The spatial problem
The previous section raises a question on the spatial considerations of habitability. What extent of a planetary body must be habitable for the entire body to be defined as “habitable”? The working definition used here makes no prescription on habitat size. If a planetary body hosts a habitat of any kind on any scale, it is a habitable planetary body. The volume of Earth that is inhabited is less than 0.5% of its total volume (Jones and Lineweaver, 2010a, 2010b), but we describe Earth as a habitable planet. A reductio ad absurdum is to ask the question: “Does a planet that hosts a one-micron cube that is habitable to a single microorganism classify as a habitable planet?” Under our working definition, the answer is yes. However, a more realistic response is to point out that such a scenario is physically unlikely. A planetary body that has the required concatenation of conditions required for an organism to be active, including liquid water, is likely to have habitable conditions across macroscopic areas (even if only regional) if they exist at all.
2.3. Less habitable and more habitable worlds?
If a habitat is a place that can support the activity of at least one known organism, then it follows that it is meaningless to speak of more or less habitable places. The assessment is binary—either an environment can support the activity of a given organism or it cannot.
Heller and Armstrong (2014) discussed the concept of superhabitable worlds—worlds “generally more habitable” than the Earth. They recognized that habitability is a binary assessment and suggested that when discussing superhabitability they are considering the analogy of a “sow being pregnant with several furrows.” This notion of superhabitability probably comes closest to the traditional ecological concept of “habitat quality,” which is usually taken to be the ability of a habitat to sustain a certain biomass of organisms. Conditions that Heller and Armstrong (2014) identified that would lead to greater biomass on a planetary body compared to that of Earth include greater surface area, a larger number of water bodies, maintenance of plate tectonics over longer times, generally warmer temperatures, older planets, and planets around more long-lived (lower-mass) stars. These factors are discussed in Section 6.
Taken in the binary sense, a superhabitable world would be one that hosts a greater diversity of organisms or a greater number of organisms with different functional capabilities [e.g., if a planet had a more diverse range of combined physical and chemical environmental stressors than that on Earth (Harrison et al., 2013), it might host a more diverse set of niches]. We do not know the extent to which the capabilities of life have been fully explored on Earth. A large number of the theoretically available redox couples on Earth have a microorganism capable of using them to conserve energy for growth (Kim and Gadd, 2008). Theoretical redox couples have been successfully used to find novel microorganisms, for example, the thermodynamic prediction of anaerobic ammonium oxidation (anammox), which led to the subsequent discovery of organisms capable of carrying out this transformation (Broda, 1977; Mulder et al., 1995).
2.4. Habitability is decoupled from the presence of life
If the definition of a habitable environment is one that can support the activity of at least one known organism, then conceptually it is possible to have an uninhabited habitat—a place that can be shown to support the activity of a known organism (or organisms) but contains no such organism (Cockell et al., 2012a). These environments are rare on Earth but can be found, for example, in fresh lava flows (Cockell, 2014b). These environments could exist in greater abundance on other planets where the hydrological regime is less vigorous than on Earth, or where the atmosphere is more inclement (e.g., high UV radiation fluxes on anoxic planets), such that new habitats are disconnected from contemporaneous inhabited environments. Alternatively, if the origin of life does not occur on a planetary body because of the lack of suitable environments (or life is not transferred from another life-bearing planet), but habitable conditions exist, then the entire planet may be lifeless, despite hosting habitable conditions. We do not know the diversity of environments in which an origin of life can occur, and how inevitable its occurrence is, once habitable conditions emerge, to be able to assess the plausibility of this scenario. The planet Mars, where habitable conditions have been reported (Grotzinger et al., 2014) but deteriorating conditions throughout its history may have made these conditions localized, is one world where uninhabited habitats could plausibly have existed or exist today (Cockell, 2014a). Several schematic trajectories can be defined for the planet depending on whether it was inhabited or uninhabited (Fig. 2). The timing of changes in the relative abundance of different environments (uninhabitable, uninhabited habitat, inhabited habitat) is one of the key challenges in defining the history of martian habitability. The trajectories defined for Mars circumscribe the general set of trajectories for any Mars-like planet on which conditions deteriorate over time.
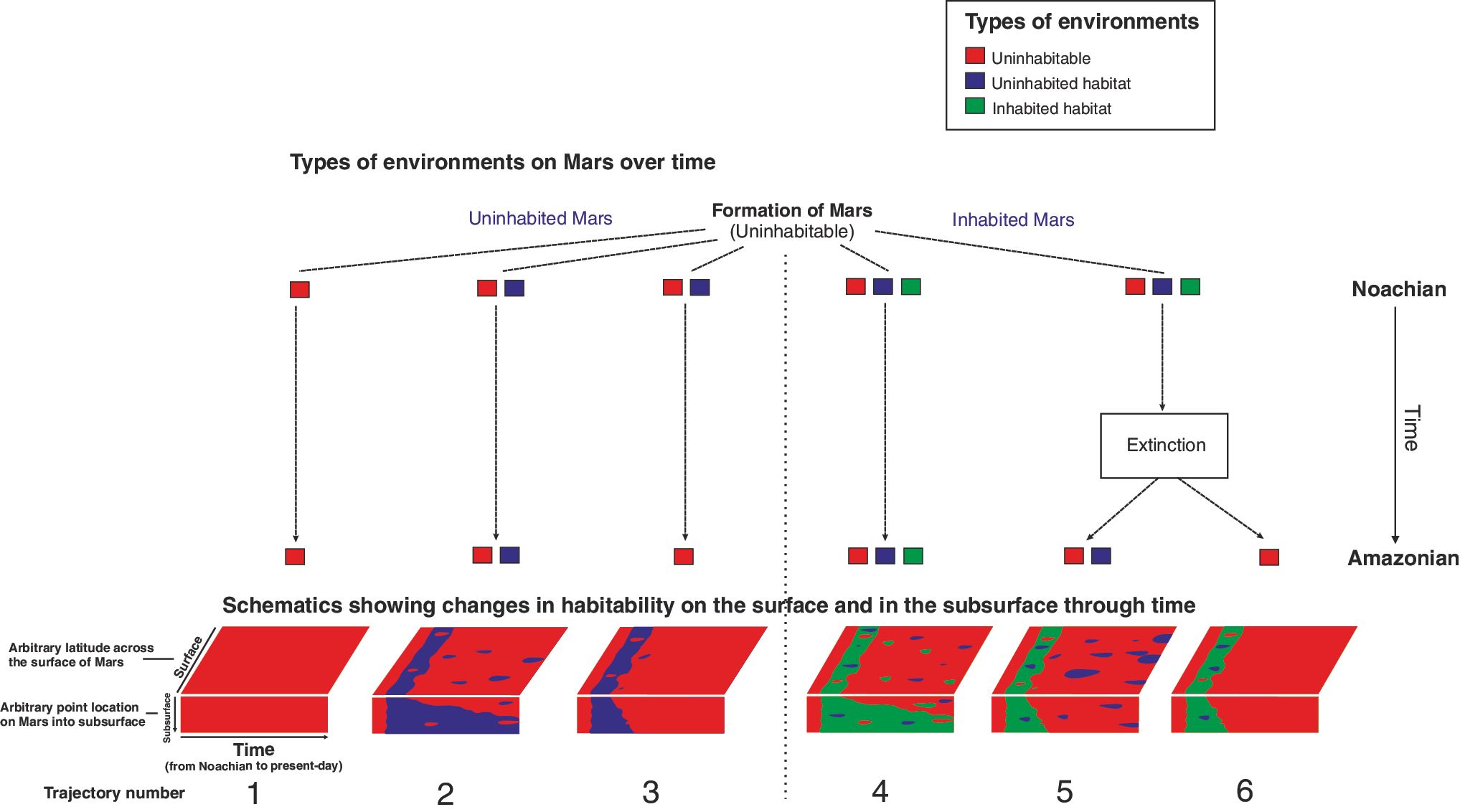
Martian habitability trajectories. Examples of different trajectories of the habitability of Mars through time, beginning with the branch point of an uninhabited and inhabited Mars. In the lower part of the diagram are shown schematics illustrating the trajectories. The “surface” section represents the surface of Mars. The “subsurface point” is an arbitrary point through the martian crust to an arbitrary depth of several kilometers. Color graphics available at
3. What Are the Requirements for Habitability?
For any type of habitability to exist, there must be the possibility of habitable planetary bodies. In the case of our own galaxy, early star systems near the center may have been disrupted by intense supernova activity and star formation. Stars too far away from the center may exist in regions of low metallicity, where many elements are at too low abundance to form terrestrial-type rocky planets. Thus, the presence of planetary bodies of relevance to this article may be restricted to a galactic habitable zone. For the Milky Way, this has been estimated to be an annulus of 7–9 kpc (kiloparsecs) from the galactic center composed of stars formed between 8 and 4 billion years ago (Gonzalez et al., 2001; Lineweaver et al., 2004). However, this review focuses on star systems in which habitable planetary bodies are possible and does not review the conditions required to give rise to habitable planets in the first place. This topic was discussed, for instance, by Gaidos et al. (2005).
3.1. Instantaneous habitability and continuous planetary habitability
In the interests of ordering this discussion, two further definitions are used. Instantaneous habitability is defined as the set of conditions at any given place in an instant in time that will support habitability. Continuous planetary habitability is defined as the set of conditions on, or in, a planetary body that can support habitable conditions in at least part of the planetary body over geological time periods. In general, instantaneous habitability applies to localized (micron to macroscopic scale) conditions on a planetary body that support the activity of organisms. In contrast, continuous planetary habitability refers to the ability of an entire planetary body to support habitable conditions somewhere on its surface, or within its interior, over geological timescales. These definitions are not merely semantic. They allow the separation of two distinct discussions. The rest of this review is focused on considering the conditions that allow for habitable environments.
4. Instantaneous Habitability
At an instant in time in a particular location, a set of requirements for all known organisms to be active can be identified (Fig. 3). They are
(1) A solvent.
(2) Appropriate temperature conditions and other physicochemical conditions (such as water activity).
(3) Available energy.
(4) Major elements required by all known life (CHNOPS).
(5) Other elements required by a specific organism.
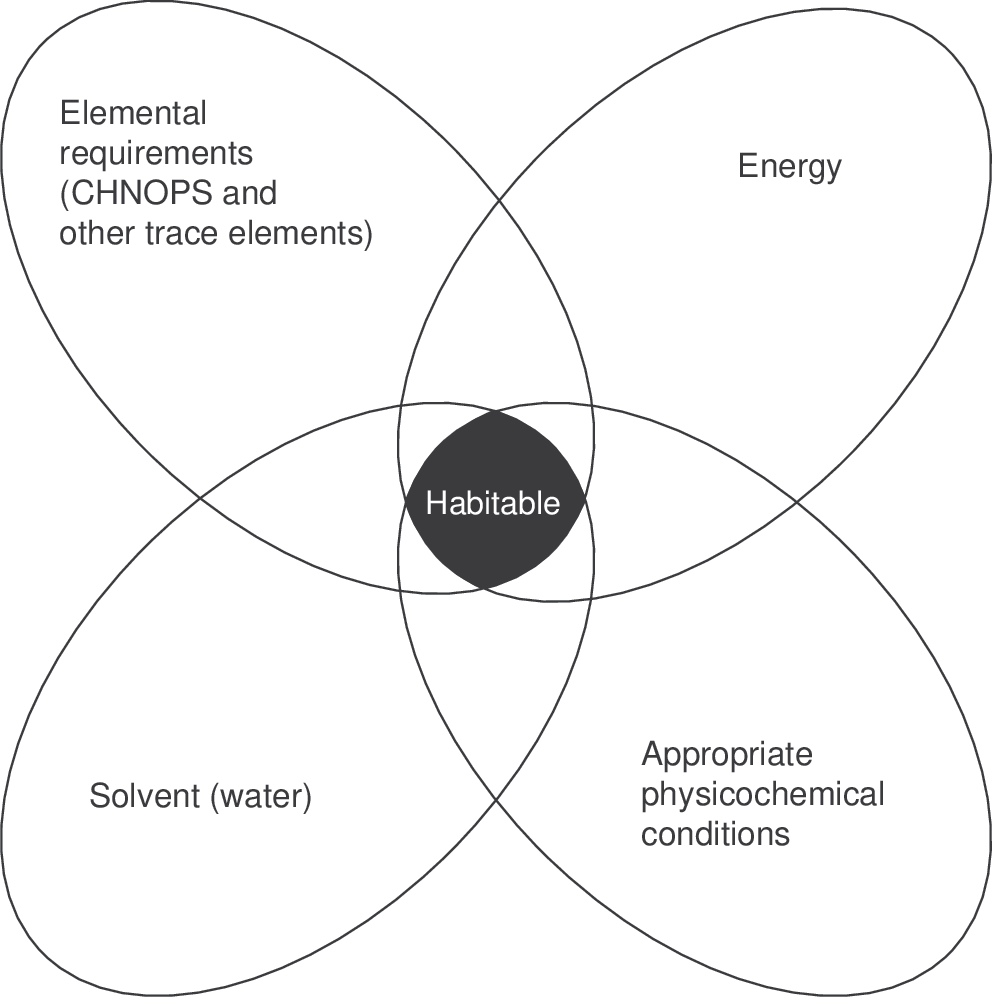
Instantaneous habitability. A series of physicochemical requirements must come together at the spatial scale of an organism to allow that organism to be active in a given environment (adapted from Hoehler, 2007).
These requirements can be assessed for any given environment or planetary body. For an environment to be habitable, they must be present and colocated at the scale of the organism (Fig. 3). However, it is also possible to more generically catalogue the presence of these requirements on planetary bodies as a first-order assessment of whether a planetary body is likely to host instantaneously habitable conditions on some part of its surface or interior. These Planetary Habitability Tables are shown for Earth, Mars, Europa, and Enceladus in Tables 1 –4.
Inventory of requirements for habitability on Earth (note that this table does not indicate whether the requirements for life are colocalized in any given environment for life). Given the vast range of minerals, valence states of elements, and redox couples that have been demonstrated, this table is non-exhaustive, but it illustrates the quantity of information on conditions for habitability compared to Tables 2 –4.
On Earth, the number of demonstrated redox couples is vast; thus this table does not represent all identified combinations. However, some combinations of electron donors and acceptors are more favorable over others. In addition to the electron acceptors listed above, humic acid, (per)chlorate, iodate, organic sulfonate, and organic nitro compounds can be used as electron acceptors by anaerobic bacteria.
2,4,6-TCP = 2,4,6-trichlorophenol; PCE = tetrachloroethene; PCP = pentachlorophenol; TCE = trichloroethene; 3-CB = 3-chlorobenzoate; 2-CP = 2-chlorophenol; 2,6-DCP = 2,6-dichlorophenol; DCE = dichloroethene.
Inventory of requirements for habitability on Mars (note that this table does not indicate whether the requirements for life are colocalized in any given environment for life). For energy sources, redox couple only shown if there is unequivocal evidence for both half reactions or for one half reaction and strong evidence or likelihood of the other (adapted from Cockell, 2014a).
Inventory of requirements for habitability in Europa (note that this table does not indicate whether the requirements for life are colocalized in any given environment for life). For energy sources, redox couple only shown if there is unequivocal evidence for both half reactions or for one half reaction and strong evidence or likelihood of the other.
Inventory of requirements for habitability in Enceladus (note that this table does not indicate whether the requirements for life are colocalized in any given environment for life). For energy sources, redox couple only shown if there is unequivocal evidence for both half reactions or for one half reaction and strong evidence or likelihood of the other.
4.1. A solvent
Liquid water is the solvent required for biochemical reactions to occur. At the current time it is the only compound known to be used by life as the primary biochemical solvent, although there have been speculations about the use of liquid ammonia, organic solvents (such as methane and ethane), formamide, and even sulfuric acid (Benner et al., 2004; Schulze-Makuch and Irwin, 2006). As we do not know of organisms that use these alternative liquids, we do not consider these possibilities further. For liquid water to be present in any location at a given time, there must be environmental conditions of temperature, pressure, and chemical impurities that allow water to fall within the liquid phase space determined by the equation of state. This is dependent on a set of planetary conditions that are discussed under continuous planetary habitability.
4.2. Appropriate temperature conditions and other physicochemical conditions
The presence of liquid water is a fundamental requirement for any form of known life, but its presence may not define an environment as suitable for life. Given the limitations to the activity of life, only part of the phase space of liquid water is habitable.
The lower limit for metabolic activity in microbes is thought to be ∼−25°C (Junge et al., 2004; Mykytczuk et al., 2013; Clarke, 2014). No convincing evidence for reproduction has been demonstrated below ∼−15°C (Breezee et al., 2004; Wells and Deming, 2006), although longer-term experiments and more sophisticated approaches may well change this assumption. Liquid water can exist at below this temperature at values that are applicable to planetary environments. For example, eutectic solutions of perchlorates have theoretical freezing points down to ∼ −65°C (Chevrier et al., 2009), well below the current lower limit for metabolic activity, although much work needs to be done to measure the freezing points of novel salt solutions under real environmental conditions. It remains to be seen whether the absolute lower temperature limit for microbial metabolic activity is coterminous with the lower limits for liquid water in the environment.
At the other extreme, liquid water can exist at temperatures well above the current upper limit for microbial growth (122°C, Kashefi and Lovley, 2003; Takai et al., 2008) at high pressures. As chemical reaction rates (including destructive processes) increase exponentially with temperature according to the Arrhenius equation, we might expect that a point is reached when destruction of cellular structures and noncovalent interactions caused by thermal energy exceeds the energy that can be harnessed from the environment by an organism to repair damage (Clarke, 2014; Corkrey et al., 2014). This limit may be something on the order of 140–150°C (Cowan, 2004). As liquid water can exist at greater than 300°C at pressures exceeding 10 MPa, it is possible that the upper temperature limit for life falls short of the upper limit for liquid water availability. The potential presence of supercritical water on exoplanets with dense atmospheres (Elkins-Tanton and Seager, 2008) suggests that planets could exist with liquid water above the upper temperature limit for life defined on Earth. This suggests that the temperature limits for life are an important first-order determinant of habitable conditions (McKay, 2014).
Other physical and chemical conditions within an environment must also lie within the bounds defined by available energy (see below) and the biochemical limits of life. Life has been found growing in extremes of different physical and chemical stressors, such as ionizing and UV radiation, pressure, pH, salinity, aridity, and toxic metals (Rothschild and Mancinelli, 2001). The absolute limits for many of these stressors are not fully known, and some of them, for example, pH, may never achieve metabolically prohibitive extremes within planetary environments. Extremes can either work in synergy or compound energetic costs to organisms. We have very little information on how organisms adapt to multiple extremes (Harrison et al., 2013).
An example of one extreme other than temperature with well-defined limits is water activity, a w, for which the lower limit is currently thought to be 0.605 (Stevenson et al., 2014). Although such limits are unlikely to be met on a planetary scale, as on Earth, they could be localized to specific brines.
4.3. Available energy
Whether for maintenance, growth, or reproduction, organisms require energy. The process of using chemical energy from redox processes is named chemotrophy; if the energy is obtained by using inorganic electron donors, it is designated chemolithotrophy. If the energy is obtained from organic compounds as the electron donor, it is designated chemoorganotrophy (chemoheterotrophy), while if the organisms use sunlight as energy source, it is designated phototrophy.
The energy-yielding process in life involves the formation of a proton gradient across a membrane. This itself is generated by electron transport. An electron is transferred from a donor element or molecule to an acceptor element or molecule with or without the help of energy available in light. The movement of an electron through the electron transport chain drives the pumping of protons across a cell membrane, thus producing a proton gradient, or proton motive force, which can be used to do work (Mitchell, 1961). More exactly, this process produces adenosine triphosphate (ATP) from the covalent bonding of phosphorus (Pi) to adenosine diphosphate (ADP) in a complex protein called ATP synthase that sits across the membrane. The energetic anhydride bond so produced in ATP can be broken elsewhere in the cell to liberate energy for biochemical processes. Thus, what energy cells require is quantized in the energy available in the phosphate bond, analogous to voltage in electric systems (Hoehler, 2007). There are a great variety of different energy metabolisms such as the reduction or oxidation of iron, sulfur, and nitrogen species, manganese reduction, methanogenesis, internal reduction and oxidation of organic compounds (designated fermentation), and aerobic respiration making use of molecular oxygen as the final electron acceptor (Schulze-Makuch and Irwin, 2004; Kim and Gadd, 2008).
For an environment to have sufficient energy to make it habitable (assuming all other requirements are met), there must be sufficient electron donors and acceptors thermodynamically favorable to the organism, or sufficient light (the energy supply), to generate sufficient energy for the organism to carry out the activities it requires (maintenance, growth, or survival). These values should be calculable from first principles (Hoehler, 2007). The energy supply available from a given redox couple can be calculated as a Gibbs free energy based on chemical considerations. The energy required by the organism should be calculable if we know the energy required to respond or adapt to certain environmental factors. In reality, this calculation may be difficult. For example, on the supply side, Fe2+, which is used as an electron donor in biological iron oxidation, may be bound within minerals such as olivine. Its rate of supply will depend, inter alia, on the rate of water-rock interactions, paths of fluid flow through the rock, and the rate of reaction of the Fe2+ with other ions. As these conditions all feed back on one another, it may never be possible to calculate to great accuracy the energy available to organisms at micron scales. On the demand side, energy required to adapt to different physicochemical extremes is not merely additive, but extremes can act in a synergistic or antagonistic manner (e.g., an enhanced water activity tolerance when certain sea-ice bacteria are grown at suboptimal temperatures; Nichols et al., 1999). Given an environment with n permutations of extremes, it is difficult to calculate the exact energy required at any given moment by an organism; this may be impossible in highly complex dynamic environments. Finally, organisms are constrained by the evolutionary and, therefore, biochemical legacy of their ancestors. The mere presence of theoretically sufficient energy to overcome a given set of physical and chemical extremes may be confounded by a lack of biochemical machinery to address the damage caused by such extremes.
Despite this, it is possible to reach a state of knowledge where one can estimate the energy available in an environment and estimate the energy required by an organism, and determine whether the supply is sufficiently abundant to make it likely that a given organism known to use that energy supply could be active (Rogers et al., 2007). Nevertheless, a concrete determination of whether an environment is habitable requires knowledge of specific organisms' requirements.
4.4. Major and trace elements
Of all the elements in the Periodic Table, six are ubiquitous in the macromolecules of known life: C, H, N, O, P, and S. Carbon is required as the core element in a vast variety of macromolecules. Hydrogen is covalently linked to carbon and other atoms in macromolecules. Nitrogen is similarly to be found, particularly in the linkages of long-chained molecules such as proteins and in the base pairs of DNA. Oxygen is used in alcohols, sugars, and a variety of molecules. Phosphorus forms part of the backbone of DNA and is used in energy-rich molecules such as ATP. Sulfur is used in protein bridges and a variety of iron-sulfur clusters in molecules involved in energy acquisition. A variety of other elements, such as magnesium and even tungsten (in anaerobic taxa), are used by different organisms for different functions. In a simplistic way, one can view evolution as having used six elements to build the chassis of life and then rummaged through the Periodic Table to find and use other elements that have specific chemical properties of use in particular biochemical functions or environments (Wackett et al., 2004). The six CHNOPS elements are available to life in a variety of forms. Some examples of these on Earth are shown in Table 1. Their concentrations are determined by planetary conditions. For example, more oxidized conditions will favor compounds such as sulfate and ferric iron as potential sources of S and Fe, respectively.
The forms in which some of these elements are available are also forms that can be used as electron donors or acceptors in energy acquisition (Table 1). Therefore, there is a tight coupling, particularly in the microbial domains of life, between habitability with respect to the availability of CHNOPS and redox couples for energy acquisition.
The detection of diverse chemical compounds and elements on other planetary bodies allows for the link between CHNOPS elements and energy availability to be used in an assessment of the habitability of other planetary bodies (for Mars specifically, an example of this approach was addressed by Stoker et al., 2010). For example, in Tables 2 –4, detected and strongly inferred chemical species on Mars, Europa, and in the plumes of Enceladus are used to assess the possible presence of the components required for instantaneous habitability.
For any given organism there may be other elemental requirements (Wackett et al., 2004). For example, iron is used widely by organisms in electron transfer proteins. There is a reported instance of microbes not requiring iron, the lactobacilli (Sabine and Vaselekos, 1967; Bruyneel et al., 1989; Weinberg, 1997). Instead, they are thought to employ enzymes and proteins that use cobalt, magnesium, manganese, and other cations (Elli et al., 2000).
5. Continuous Planetary Habitability
What conditions are required for at least part of a planetary body to have instantaneous habitability over geological timescales? This section explores this question.
5.1. Water: Surface liquid water worlds and interior liquid water worlds
The distribution of liquid water can be used to recognize broadly two types of planetary bodies. The first type is a planet with liquid water on its surface as well as in its interior, where the liquid is sustained from a combination of internal heating and stellar energy; the latter in most cases will be the dominant form of energy for keeping water on the surface in a liquid state. These are “surface liquid water worlds.” Earth is an example. The second type of planetary body is one where stellar radiation is not sufficient to maintain surface liquid water but where liquid water exists in the interior. These are “interior liquid water worlds.” There is a variety of this type of planetary body. It includes icy moons with subsurface oceans (in our solar system, examples include Enceladus, Europa, and Ganymede) and some terrestrial-type rocky planets where stellar flux alone is not sufficient to maintain liquid water (on the surface), but internal sources of energy maintain liquid water (McMahon et al., 2013). Isolated lone planets in interstellar space have also been suggested as locations for liquid water (Stevenson, 1999; Abbot and Switzer, 2011).
Surface liquid water worlds can transition to interior liquid water worlds. The dry, desiccated surface of Mars, in contrast to its more water-rich past (Jakosky and Phillips, 2001), is an example of how conditions on the surface of a planet can deteriorate (for reasons discussed later).
The distinction between these two types of worlds from an astrobiological point of view may well be categorical.
In the case of surface liquid water worlds, the presence of liquid water may occur spatially colocated with the presence of light, thus allowing for photosynthesis. If we accept an assumption that the emergence of multicellularity and intelligence is linked to the presence of high concentrations of oxygen from oxygen photosynthesis that allows for aerobic respiration (Catling et al., 2005), then these worlds may be the only type capable of producing the conditions for instantaneous and continuous planetary habitability required by intelligent organisms. By contrast, interior liquid water worlds may have habitable conditions for a range of metabolisms but not conditions for photosynthesis. Some calculations, however, suggest that production of surface oxidants, if cycled into the interior, could lead to suboceanic oxygen concentrations as high as terrestrial surface waters (Hand et al., 2007). It is unknown whether such a scenario could support multicellular life, let alone intelligence. In the case of Mars or Mars-like planets, for instance, the transition from conditions capable of sustaining surface liquid water to those that cannot renders photosynthesis an implausible energy supply across most, if not all, the surface. This truncates the conditions for continuous planetary habitability required for the large-scale production of oxygen by oxygenic photosynthesis and thus multicellular life and intelligence.
5.1.1. Appropriate temperature conditions for surface liquid water worlds
The window within which life can operate is smaller than the total pressure-temperature phase space for liquid water (see above, Jones et al., 2011; Jones and Lineweaver, 2012). Thus, this smaller phase space must also be sustained over geological lifetimes. This requirement implies liquid water between ∼−25°C and 122°C over geological timescales.
Maintaining temperatures within the range required for biological activity on the surface of a planet depends on a sufficiently powerful greenhouse forcing. If the effective temperature of a planetary surface exceeds certain values, set by the energy received from its star and the greenhouse effect caused by the gases present in its atmosphere, then the water on the planet will evaporate at sufficient levels to supply the upper atmosphere with a moist greenhouse effect (Kasting, 1988). As the moist greenhouse becomes more effective at raising the temperature, a positive feedback may develop between the increasing levels of atmospheric humidity, the surface temperature, and rates of evapotranspiration. The moist greenhouse may eventually result in a runaway greenhouse effect (Walker et al., 1970; Nakajima et al., 1992). As observed on Venus, a runaway greenhouse will cause all loss of water from the planet, making it uninhabitable. The moist and runaway greenhouse effects therefore define an orbit that is too close to the star for liquid water to be sustained on the surface of a planet.
A planet too far from a star will suffer the effects that the carbon dioxide in the atmosphere can condense onto the surface, reducing the concentration of this greenhouse gas and contributing to low temperatures, resulting in a frozen surface. If CO2 is abundant enough, scattering can contribute to ineffective heating of the surface. This in itself will depend on the quantity of CO2 outgassed by the planet, linking habitability to planetary interior structure (Noack et al., 2014). These conditions define an outer limit for habitability, with the boundary conditions for life additionally depending on the star type. Hot stars, such as F stars, have boundary conditions for surface liquid water farther from the star compared to our own Sun (a G star). Cooler low-mass stars, such as K and M stars, have boundary conditions closer in (Fig. 4). The outer limit of the habitable zone may be considerably extended on planets with strong greenhouse gases such as hydrogen, which theoretically could expand the outer limit to ∼10 AU (Pierrehumbert and Gaidos, 2011). In extreme cases, free-floating planets not gravitationally bound to a star may even harbor surface habitable conditions (Stevenson, 1999).
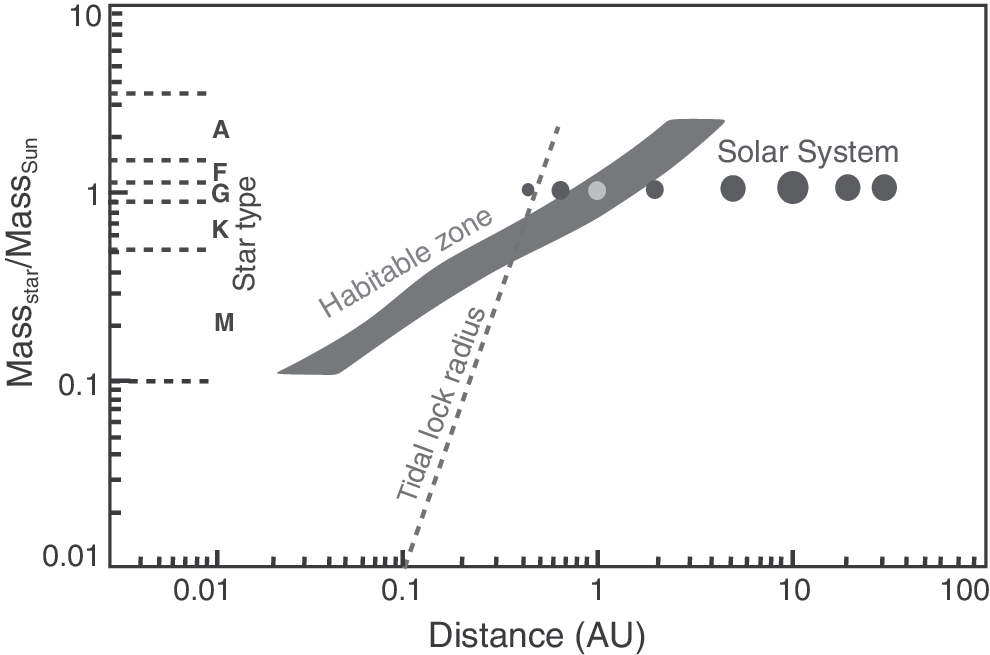
The habitable zone for different spectral types. Our own Solar System is shown. (diagram modified from Kasting et al., 1993).
The so-called habitable zone is thus defined as the zone around a star where liquid water is stable at the surface of a planetary body (usually an Earth-mass planet) (Hart, 1979; Kasting et al., 1993; Kasting, 1997; Franck et al., 2000; Gonzalez, 2005; Kopparapu et al., 2013). The habitable zone is an old concept (Huang, 1959). The idea was developed by Dole in his book Habitable Planets for Man in which he elaborated the idea of the circumstellar habitable zone as well as various other determinants of planetary habitability (Dole, 1964).
In binary star systems, the presence of a second star can influence the habitable zone boundaries. Depending on the dynamics of the system, the boundaries can vary over time as a result of the gravitational interactions of the two stars (Haghighipour and Kaltenegger, 2013), or a bright second star can cause the boundaries to be further out than they would in a single star system (Kaltenegger and Haghighipour, 2013).
As a star's luminosity changes over time, generally increasing (a consequence of hydrogen burning on the main sequence), the habitable zone will move outward. Integrated over time, there is therefore a narrower band, the continuously habitable zone, within which conditions for surface liquid water are met (Hart, 1978; O'Malley-James et al., 2013; Rushby et al., 2013). As the boundaries of the habitable zone move outward, a planet can eventually cross the inner boundary, and surface temperatures will become too great to sustain liquid water. Therefore, a planet remains in the habitable zone for a given length of time, known as its “habitable lifetime.” This lifetime largely depends on stellar mass (but the physical properties of a planet and its atmosphere play a role). The more massive a main sequence star, the faster it burns through its hydrogen fuel supply, causing a more rapid increase in its luminosity. This pushes the habitable zone boundaries outward at a greater rate.
The situation is different for stars at the beginning of their lifetimes. Pre-main sequence stars are thought to have habitable zones that move inward as stellar luminosity initially decreases as they move toward the main sequence (Ramirez and Kaltenegger, 2015). This raises the possibility that planetary bodies that orbit a young star lose their water inventory because of an intense greenhouse effect (Luger and Barnes, 2015). They later end up in the habitable zone during the star's main sequence phase but would lack sufficient water to support life. Thus, an assessment of continuous planetary habitability must consider a planet's relationship to the changing location of the habitable zone from the pre-main sequence to the end of a star's life.
In the case of Earth, it has remained within the Sun's continuously habitable zone since its formation. It is expected to leave the habitable zone after another 2–3 billion years (O'Malley-James et al., 2013, 2014; Rushby et al., 2013). As the inner boundary of the habitable zone moves closer to Earth, environmental conditions on the planet are expected to change as the planet receives more solar energy. The planet will become hotter and drier, leading to environments that become increasingly hostile to life, setting into motion extinction sequences that begin with the eukaryotes and end with the extinction of extremophilic microorganisms (O'Malley-James et al., 2013, 2014). Hence, when assessing the habitability of a planet, it is necessary to consider the amount of time it has spent in the habitable zone, as well as its current position within the habitable zone.
While stellar evolution can cause the end of habitable conditions on a planet, it might also render previously uninhabitable worlds habitable. For example, in the later stages of the Sun's evolution, when it swells to become a Red Giant, the habitable zone will encompass Saturn's moon Titan—a moon that contains various organic compounds, which has been suggested to lead to a new habitable world (Lorenz et al., 1997).
A conceptually similar habitable zone can be formulated for a large moon orbiting a planet, making exomoons that have surface liquid water a viable possibility (Heller and Barnes, 2013). The inner circumplanetary habitable edge is defined as the orbit where a runaway greenhouse effect is caused by illumination from the planet and tidal heating (Heller and Barnes, 2013). The habitability of moons orbiting planets is dependent on orbital eccentricity (Forgan and Kipping, 2013). For example, large eccentricities can generate strong tidal heating, which might prevent the formation of stable bodies of water. Unlike tidally locked planets, moons tidally locked to their planets will experience variations in solar insolation, which might reduce the possibility of atmospheric freeze-out by providing combined planetary and solar insolation on different regions of the moon (Joshi et al., 1997).
5.1.2. Appropriate temperature conditions for interior liquid water worlds
The limitations of the habitable zone concept can be understood at once when we consider a planet with insufficient atmospheric conditions (or no appreciable atmosphere) to sustain surface liquid water but a sufficient internal source of energy to generate liquid water within the interior (interior liquid water worlds). This source of energy raises the possibility of planets outside the habitable zone with habitable conditions. In the case of icy moons, if a body has an eccentricity (noncircularity of orbit), obliquity (axial tilt), and/or nonsynchronous rotation state, then tidal effects can heat the interior. These conditions can melt internal ice over geological time periods (Reynolds et al., 1987; Scharf, 2006). This phenomenon is inferred from the observations of water plumes emanating from Enceladus (Waite et al., 2006, 2009) and inferred for the three Galilean moons Europa, Ganymede, and Callisto based on induced magnetic fields and thermodynamic calculations (Khurana et al., 1998; Vance et al., 2014; Saur et al., 2015).
The Galilean moons are instructive in demonstrating how the strength of tidal forces may play a role in regulating habitable conditions with respect to the availability of liquid water. Io, the closest of the moons, with a semimajor axis of 421,700 km, is so tidally active that it hosts active silicate and sulfur volcanoes (McEwen et al., 1998). This intense activity likely precludes liquid water. Europa, with a semimajor axis of 670,900 km, hosts substantial evidence for a subsurface ocean and a young geologically active surface (Schmidt et al., 2011). Ganymede, with a semimajor axis of 1,071,600 km, has a more ancient surface but is thought to host a subsurface ocean (Vance et al., 2014). Callisto, the farthest moon out, at 1,882,700 km from Jupiter, has an even more ancient surface, although an induced magnetic moment suggests an ocean deep within the moon (Khurana et al., 1998). Other factors aside from orbital radius, such as the size of a body and resonances between moons, also play a role in defining the strength of tidal interactions.
Tidal interactions enable liquid water to exist, but it remains unknown whether the other factors required for instantaneous habitability to exist and for these requirements to persist are met in these subsurface water bodies. What is crucial is whether the body of water is in contact with the surface or a subsurface core (Ruiz and Tejero, 2003; Hand and Chyba, 2007). From the surface, meteoritic matter, such as organics and various cations and anions, or other matter, such as radiation-produced oxygen and other oxidants, has a chance of being entrained within the ocean. In the subsurface, contact with a rocky core could provide cations and anions, which, as for the surface, may include not only required elements for growth but diverse half reactions required for energetic redox reactions such as iron and sulfate reduction and methanogenesis (Chyba and Phillips, 2001; Schulze-Makuch and Irwin, 2002; Pappalardo et al., 2009). Despite the theoretical possibilities, however, when the availability of CHNOPS elements and half reactions for redox couples is constrained to measured or strongly inferred elements and compounds, we still lack considerable knowledge about instantaneous habitability on Europa and Enceladus (Tables 3 and 4). For Europa, surface-interior interactions are inferred on account of such processes as putative plate tectonics (Kattenhorn and Prockter, 2014), although the extent to which these processes produce mixing between the surface and interior ocean is not known. By contrast, for Ganymede, the surface is ancient, and the internal liquid water is thought to be sandwiched between high-pressure ice layers (Vance et al., 2014), thus preventing, or at least minimizing, internal water interactions with both the surface and an internal rocky core. Such an arrangement might provide conditions that are habitable to a smaller range of organisms, if any, over geological timescales.
Saturn's moon Enceladus is thought to be a differentiated body with a rock-metal core surrounded by a liquid water–containing ice layer (Schubert et al., 2007). The extent of contact or circulation between the liquid water and the rocky core is not known. However, the detection of silica nanoparticles in Saturn's E-ring has been interpreted as evidence of present-day active hydrothermal activity in the moon in contact with the core (Hsu et al., 2015). Like Europa, tidal forces are thought to be responsible for ice melting, possibly aided by radioactive decay in the rocky core, which is thought to explain the lack of similar geological activity on Saturn's moon Mimas, which orbits closer to Saturn and has higher eccentricity but a small rock fraction (Schubert et al., 2007).
Even in rocky planets, internal heat from radioactive decay could generate temperature conditions sufficient to sustain liquid water in the subsurface far outside the boundaries of the traditional habitable zone (McMahon et al., 2013) and possibly in interstellar space (Abbot and Switzer, 2011).
5.2. Other physicochemical conditions
The physicochemical limits for life over geological times are identical to those defined for instantaneous habitability since the long-term limits are set by life's ability to cope with any given physical and chemical extreme at a point in time.
5.3. Available energy
For an environment or at least one location on a planet to be continuously habitable over geological time periods, it must provide sufficient energy for activity by a given organism. For photosynthesis to be a sustained energy source, the surface of the planet must sustain liquid water. For other forms of energy, the requirement is a set of chemical disequilibria that provides electron donors and acceptors for redox couples. This implies geochemical turnover or activity within the planetary crust to produce reduced and more oxidized compounds colocated at small scales for use by life over geological time periods.
5.4. Major elements
Over geological timescales, what are potential sources of CHNOPS?
Carbon may always be available as CO2 in an atmosphere throughout most of the lifetime of a terrestrial-type planet. Its presence in the atmospheres of Venus (96.5%), Earth (∼400 ppmv), and Mars (96%) shows that it is present on most types of terrestrial-type rocky planets. However, it is not necessarily the case that the CO2 can be fixed, for example by photosynthetic organisms, if its concentrations are too low for the carbon assimilation mechanisms used by a particular biota (e.g., the loss of photosynthesis at below 35–45 ppm CO2 in C3 plants; Bauer and Martha, 1981).
The long-term presence of hydrogen depends on its source. Serpentinization reactions (e.g., the reaction of the mineral fayalite with water) producing H2 cannot be sustained over geological time periods unless there is geological activity to sustain water flow through the crust (Okland et al., 2012). Hydrogen can be obtained from organic molecules which could be endogenously produced by Urey-Miller-type reactions (Bada, 2013). Hydrogen could also be obtained from diverse carbon compounds in meteoritic material (Sephton, 2002). Hydrogen is also pervasively present in water, the latter being required for habitable conditions to exist in the first place.
Nitrogen is present in the atmospheres of Venus (3.5%), Earth (78.1%), and Mars (1.9%). An environment can be habitable that contains N2 at a partial pressure suitable for biological nitrogen fixation (Mancinelli and Banin, 2003), although this presupposes the existence of the energetically expensive (16 molecules of ATP per N2 molecule) biochemical machinery to fix the gas. In the absence of biological nitrogen fixation, fixed nitrogen compounds such as ammonia, nitrite, and nitrate must be available. Abiotic processes such as impact events, hydrothermal activity, and lightning discharge can generate these compounds (Brandes et al., 1998; Segura and Navarro-González, 2005; Summers and Khare, 2007; Manning et al., 2009). This implies the presence of active atmospheric processes (lightning) or exogenously generated geological activity (impact events). On planets with reducing conditions in their interior, nitrogen is expected to be predominantly in the form of ammonia, which is sequestered within silicates. However, in more oxidizing conditions, such as Earth's mantle, the element is in the form of nitrogen gas, which is more readily degassed (Mikhail and Sverjensky, 2014). Although more oxidizing conditions may therefore favor a more nitrogen-rich atmosphere, the form of nitrogen is energetically less available to life.
Oxygen is present in a wide diversity of compounds, such as oxides. However, many of these atoms, such as oxygen atoms bound to silicon in silicates, are not directly accessible to life. Suitable sources of oxygen atoms include compounds such as sulfates and iron oxides found ubiquitously on Earth and on Mars (Bibring et al., 2007). Oxygen exists in water, which is the source of this element for oxygen gas produced in oxygenic photosynthesis. Oxygen atoms also exist in organic molecules within, for example, alcohol and carboxylic acid groups. As for hydrogen, these organics can be produced endogenously or delivered exogenously.
On Earth, phosphate is available in igneous rocks as apatite and merrillite and therefore available in other rock types as a consequence of the rock cycle. Its detection on Mars illustrates its potentially ubiquitous availability on terrestrial-type rocky planets (e.g., Usui et al., 2008). Phosphorus can also be delivered to the surface of planets in meteoritic material such as in schreibersite [(Fe, Ni)3P] (Pasek et al., 2007).
Sulfur atoms are available in planetary bodies in diverse sources such as sulfides, sulfates, and compounds of intermediate oxidation state such as thiosulfate. The presence of water can enhance the diversity and abundance of these sources of sulfur. This is illustrated by Hesperian martian geochemistry. The production of sulfuric acid in low water-rock ratio interactions of water and SO2 has led to a diverse suite of sulfate minerals on Mars (Morris et al., 2006; Bibring et al., 2007).
In summary, there are diverse sources of CHNOPS elements available to life over geological time periods on, or within, planetary bodies. All of them can potentially be delivered to a surface by impact events. However, active geological turnover (such as plate tectonics, see Section 6.1.3) or atmospheric chemistry will increase the abundance and diversity of compounds in which these atoms are available, thus enhancing the likelihood of instantaneous habitability for given types of organisms. Two examples are the abiotic production of oxidized nitrogen compounds, whose diversity and abundance will be enhanced by greater atmospheric and geologically active processes, and hydrogen, whose availability can be enhanced by geologically active processes in planetary crusts such as serpentinization. Furthermore, the more geological turnover there is, the greater will be the number of environments in which CHNOPS elements, through mixing, are likely to be colocalized at small scales to be accessible to life. Geological turnover will ensure the constant reworking of supplies of these elements into environments where they may have become depleted by, for example, leaching in hydrological processes.
Most of the observations above refer to rocky terrestrial-type planets. The abundance and availability of CHNOPS in subsurface oceans, for example in icy moons (a type of interior liquid water world), is not well understood. As for rocky planets, impacts can deliver inventories of these elements provided the material they deliver reaches, or is cycled, to the interior of the oceans. The contact of water in such oceans with a rocky core could enhance the geochemical diversity and abundance of elements and compounds available to life.
5.5. Other elements
The same observations for CHNOPS apply to other elements required by life. A wide variety of cations (such as K+, Fe2+/3+, Mg2+) are available on a planetary surface and interior over geological time spans in materials such as igneous rocks. The combination of geological activity with a hydrological cycle circulates them within the surface and subsurface of a planetary body, making them available and colocated at small spatial scales in environments where previous water flow may have leached them and depleted their concentrations.
Life itself will change the availability of elements on a planetary body (including the CHNOPS elements). For example, the oxygenation of the atmosphere in the Paleoproterozoic ∼2.4 billion years ago and again in the Neoproterozoic ∼0.7 billion years ago undoubtedly vastly enhanced the mineral diversity of the planet by, for example, the production of mineral oxides (Hazen et al., 2008). Large-scale planetary oxidation can enhance the availability of some elements (such as fixed states of nitrogen), but concomitantly it can also reduce the availability of other elements (such as Fe, which is less soluble in the oxidized Fe3+ state compared to the reduced Fe2+ state at circumneutral pH).
6. Factors for Continuous Planetary Habitability
There are a range of planetary and astronomical factors that influence the availability of previously described requirements for continuous planetary habitability. These factors can influence habitability in one of two ways: (1) Existence of habitable conditions. Some factors are crucial for determining whether a planet is habitable to any type of life. (2) Extent of habitability. Some factors are likely to modify whether a planet is habitable for particular types of organisms and the time period over which continuous habitability for any given type of organisms can be sustained.
Understanding these factors is important because they may determine whether a planetary body retains continuous habitable conditions long enough to allow certain types of life. The comparison of some of these factors with those associated with Earth has been proposed as a basis for quantifying habitability (Schulze-Makuch et al., 2011).
In particular, astrobiologists are interested in factors that control the presence of surface liquid water on a planet and thus the potential for oxygenic photosynthesis leading to the emergence of multicellularity and intelligence. Factors that cause geological activity, and thus geochemical turnover, that result in the continuous presence of diverse redox couples and the elements required for life are also important. Here, these factors are broadly split into planetary factors (factors that result from the characteristics of the planetary body itself) and astronomical factors (factors that result from the astronomical environment of a planet). In this paper, we discuss four important examples of each. Most of these factors have a profound influence on the possibility of surface liquid water, but their relevance to interior liquid water worlds will also be discussed. In Table 5, an example of each of these factors and its influence on continuous planetary habitability is shown. Figure 5 shows an example of how some of these factors can interact.
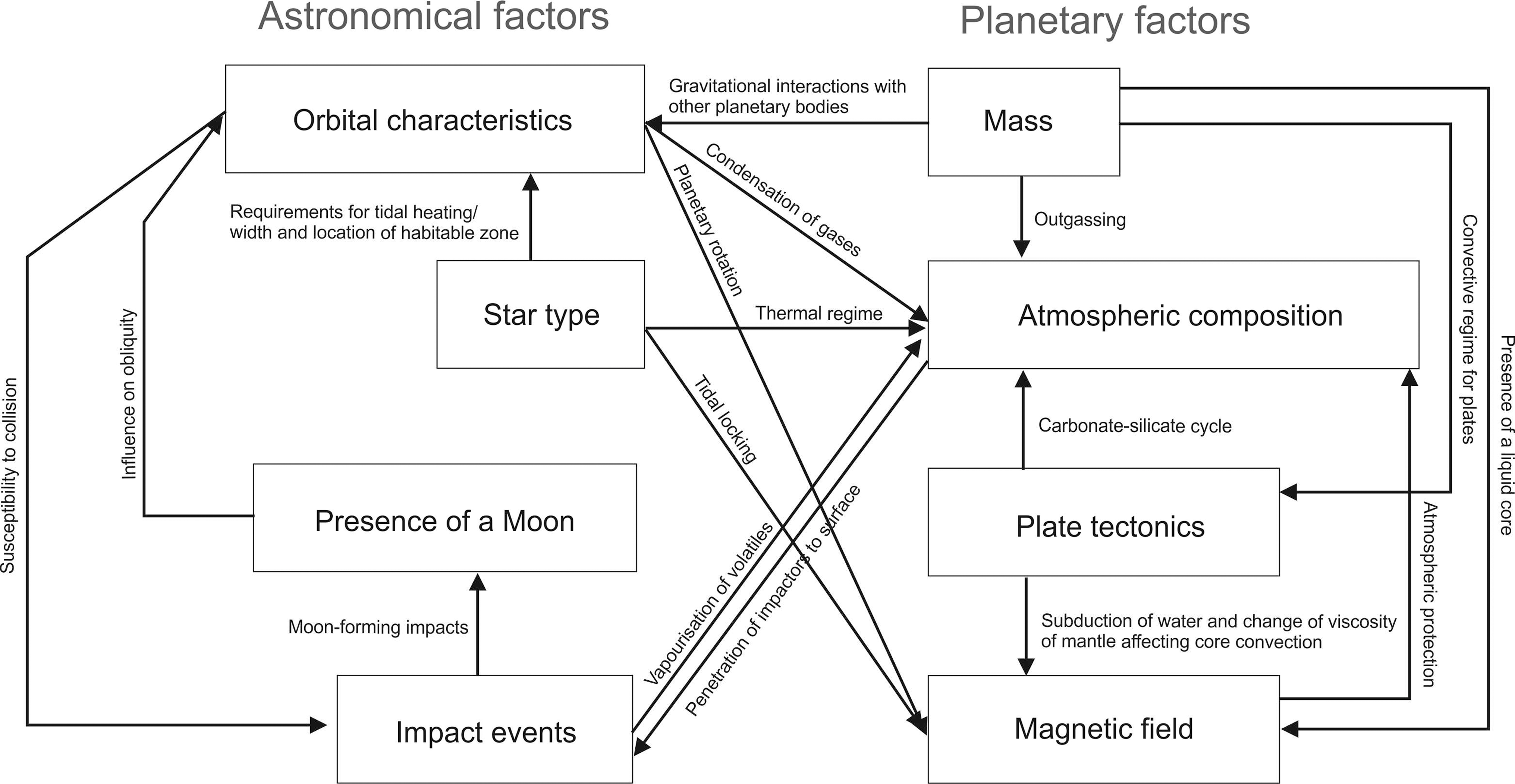
Examples of astronomical and planetary factors that influence the presence of liquid water on the surface of a planetary body. The figure shows examples of potential interactions that influence habitable conditions. This diagram does not include factors influencing the habitability of interior liquid water worlds (examples shown in Table 5).
Statements in italics, where appropriate, show factors that apply to habitability in interior liquid water worlds.
6.1. Planetary factors
6.1.1. Planetary mass/density
The mass of the planetary body influences habitability in a variety of ways. The mass will determine whether the object retains enough primordial heat or has enough radiogenic heat to maintain a liquid core, allowing for an atmosphere-protecting magnetic dynamo (Breuer and Spohn, 2003). The temperature gradient through a planetary body, which is influenced by its initial mass (influencing the energy released during accretion and differentiation), will influence whether plate tectonics can be initiated and sustained over its lifetime (Noack and Breuer, 2014). Both magnetic field and tectonic activity are discussed in more detail below.
Planetary mass will determine atmospheric composition by influencing both the degassing of volatiles and the extent to which a planet retains its primordial atmosphere. This will itself determine the concentration of different types of greenhouse gases and whether they are sufficient to sustain liquid water on at least part of the surface of a planet (Kasting et al., 1993; Kasting and Catling, 2003).
The study of exoplanets has revealed the considerable complexity in understanding how planetary mass influences the history of planetary atmospheres and thus the habitability of planetary surfaces.
For example, “super-Earths” are planets with a size that is larger than that of Earth and masses less than 10 Earth mass (10 M ⊕). However, many small planets have been found to have mean densities incompatible with rocky, Earth-like planets. In many cases, large hydrogen-dominated envelopes and/or large amounts (up to 100%) of water are necessary to explain the observations, which make such planets potentially “mini-Neptunes” instead of “super-Earths” (Barnes et al., 2009; Lammer, 2013; Lammer et al., 2014; Marcy et al., 2014; Rogers, 2014; Luger et al., 2015). This raises the question as to what fraction of low-mass planets in the habitable zone are indeed rocky and thus suitable for the evolution of life.
The early evolution of these is also important for the trajectories they take toward, or away from, being habitable. Theoretical studies indicate that rocky planets may accumulate large gaseous envelopes either by accretion of gas from the protoplanetary nebula or by outgassing (e.g., Hayashi et al., 1979; Elkins-Tanton and Seager, 2008; Elkins-Tanton, 2012; Lammer, 2013, and references therein). According to a number of studies (Ikoma and Hori, 2012; Bodenheimer and Lissauer, 2014; Lammer et al., 2014; Stökl et al., 2015), even Earth-like planets can accumulate up to 1000 Earth ocean equivalent amounts of hydrogen.
Modeling suggests that protoplanets with core masses that are ≤1 M ⊕ can lose their captured hydrogen envelopes during the active X-ray and extreme ultraviolet (XUV) phase of their young host stars (Luger et al., 2015), while rocky cores within the so-called “super-Earth” domain probably cannot get rid of their nebula-captured hydrogen envelopes during their lifetime (Lammer et al., 2014). These results indicate that the terrestrial planets in our solar system lost their nebula-based early atmospheres during the intense XUV activity phase of the young Sun or reached their final mass tens of millions of years after the nebula gas evaporated. It has been suggested that planets, like the terrestrial planets in our solar system, that can lose their nebula-captured hydrogen envelopes and keep their outgassed or impact-delivered secondary atmospheres inside the habitable zone of G-type stars most likely have core masses with 1 ± 0.5 M ⊕ and corresponding radii between about 0.8 and 1.15 R ⊕ (Lammer et al., 2014). Similar results have been presented by Kislyakova et al. (2013) and Luger et al. (2015).
However, even fast accreted Earth-like cores and “super-Earths” with up to a few percent of their mass in hydrogen and helium may still harbor conditions for liquid water oceans. However, at higher hydrogen envelope fractions, surface pressures in excess of more than 1 GPa (e.g., Choukroun and Grasset, 2007) would result in the formation of high-pressure ices, making the planet uninhabitable. These theoretical results show that the extent to which a terrestrial-type rocky planet keeps it primordial gas inventory may have a dramatic influence on its suitability for surface liquid water and thus habitability.
Catastrophic outgassing of H2O and CO2 is another process that may build up massive early atmospheres around rocky cores (Elkins-Tanton and Seager, 2008; Elkins-Tanton, 2011, 2012; Lammer, 2013). Because “super-Earths” have deeper magma oceans than Earth, they are likely to outgas more massive atmospheres than Earth-mass planets. Up to several 104 bar is possible (Elkins-Tanton, 2011). If the early atmosphere is removed from a planet, a secondary atmosphere may subsequently build up from these processes, and the planet may become habitable, provided the host star's activity has already decreased and the secondary atmosphere is not depleted significantly. Such a scenario has been suggested for early Earth (Sekiya et al., 1980). The high XUV fluxes of active young stars can lead to significant heating and expansion of the upper atmosphere (Tian et al., 2008; Lammer, 2013) so that the atmospheres are no longer protected against the stellar wind (Lichtenegger et al., 2010; Lammer et al., 2011; Lammer, 2013). A combination of strong planetary magnetic fields (see Section 6.1.4) and large quantities of a gas such as CO2 may be necessary to suppress loss of the atmosphere. If the atmospheric escape mechanisms during the early history of a planet are too inefficient and/or the early atmosphere is too massive, such planets may resemble “mini-Neptunes” rather than terrestrial planets (Lammer, 2013).
In the case of interior liquid water worlds, mass will influence the extent of tidal distortion and thus the source of energy for sustaining liquid water over geological time periods.
6.1.2. Atmospheric and surface characteristics
The atmospheric and surface characteristics of a planet will alter habitability by changing the effective temperature resulting from the balance between energy received from a star and the energy lost (Kasting et al., 1993), thus influencing the possibility of surface liquid water. Two of the most important factors are the atmospheric composition and the albedo.
Atmospheric composition will determine whether the concentration of greenhouse gases is sufficient to maintain liquid water on a planetary surface, thus categorically modifying whether a planet has any habitable conditions, or at least determining the extent of liquid water habitats. The presence of liquid water on the surface of early Earth, for instance, at a time when the Sun was less luminous (the “faint young Sun paradox”) motivates research into the type of greenhouse gases (e.g., NH3, CH4, and CO2) that could have maintained surface liquid water (Sagan and Mullen, 1972; Pavlov et al., 2001; Haqq-Misra et al., 2008). The removal of greenhouse gases from an atmosphere, for example, reduction of CH4 concentrations caused by increases in atmospheric O2 on Earth, could have had catastrophic effects on habitability by forcing the planet into an ice covered “Snowball” state (Hoffman et al., 1998).
Atmospheric composition will also be influenced by planetary mass. A large planet may retain its primordial hydrogen, or it may have a larger reservoir of reducing gases, frustrating the oxidation of the atmosphere and the possibility that photosynthesis can generate an oxygen-rich atmosphere required for multicellular life. Smaller planets could potentially make the transition to oxygen-rich conditions earlier in their history (McKay, 1996).
The quantity of water on a planet is thought to influence the effectiveness of the greenhouse effect. Drier planets have been shown by modeling to be less prone to a runaway greenhouse than wetter ones (Abe et al., 2011).
On small geological timescales (millions of years), atmospheric composition is one factor that influences surface heating and thereby the surface temperature of a planet, with implications for the frequency and severity of ice ages. Warmer periods may lead to stagnant oceans, with the rapidity of onset and emergence potentially causing extinctions (Huey and Ward, 2005) by changing the distribution of habitable conditions on a planetary surface for particular types of organisms over relatively short time periods.
Albedo influences surface temperatures in a number of ways. Cloud layers reflect radiation and can reduce the temperatures experienced on a planetary surface by, for example, expanding the habitable zone inward for tidally locked planets orbiting low-mass stars (Yang et al., 2013). Ice-snow albedo can be deleterious to habitability by providing a positive feedback loop in the onset of Snowball conditions, whereby a growth in ice and snow cover increases the reflection of radiation, causing temperatures to drop, thus enhancing snow and ice cover (Hoffman et al., 1998). The effects of ice-snow albedo on conditions for habitability are thought to become less important in the outer regions of the habitable zone where, if CO2 concentrations are high enough, they mask the climatic effects of the ice-snow albedo (Shields et al., 2013).
The albedo of the surface may be further influenced by the possible existence of life-forms that cover a large part of the surface (as trees and other plants do on Earth). A parable of an inhabited world inducing strong albedo feedbacks was described as so-called “Daisyworld” in the work of Watson and Lovelock (1983). In their theoretical model, two species of daisies of different colors absorb different amounts of radiation. The growth rate of the daisies depends directly on the surface temperature.
Daisies with a higher albedo survive and evolve at higher temperatures (and lead to a decrease of the surface temperature), whereas daisies with a low albedo (e.g., black daisies) have a higher growth rate at low temperatures (and lead to an increase of the surface temperature).
This theoretical system therefore self-regulates the global surface temperature based on albedo feedbacks and may even develop into a Darwinian system, where species adapt their internal physiology in response to environmental changes (e.g., Lenton and Lovelock, 2000).
6.1.3 Plate tectonics
It is not known whether plate tectonics as a mechanism of geological turnover is required to ensure that supplies of CHNOPS, other elements, and redox couples are sustained over geological time periods on, and within, a planetary body. Geological turnover might be achieved by magma upwellings (hot spot volcanism) or tidally driven circulation of water in a subsurface ocean through a rocky substrate (e.g., in icy moons) to sustain the circulation of essential elements and chemical disequilibria, even if only locally somewhere in, or on, a planetary body.
However, there are good reasons to suspect that plate tectonics is an important factor in sustaining conditions for surface liquid water over billions of years through its role in temperature regulation. By subducting rocks over large areas, plate tectonics provides a return pathway for CO2 in the atmosphere that has been sequestered in carbonate rocks. This is the carbonate-silicate cycle (Fig. 6), part of the carbon cycle. The negative feedback inherent in this process [higher CO2 in the atmosphere generally, though not always (Tyrell, 2014), leads to warmer conditions that enhance chemical reaction rates and increase rock weathering, thereby facilitating more effective drawdown of CO2 from the atmosphere] makes this carbonate-silicate cycle a long-term thermostat that regulates planetary surface temperatures within a range suitable for liquid water and biological activity (Walker et al., 1981; Berner et al., 1983). Both Mars and Venus illustrate the effects of the lack of this cycle. In the case of Venus, the loss of liquid water from the greenhouse effect not only prevents carbonate formation, but any buried carbon has long since been heated and returned to the planet's thick CO2 atmosphere. On Mars, on the other hand, lack of continuous volcanic outgassing leads to an atmosphere too thin to allow for an efficient greenhouse effect and hence conditions suitable for sustained surface liquid water.

The carbonate-silicate cycle. (
Planetary mass plays a role in determining the effectiveness of plate tectonics. Earth-mass planets or more massive planets have sufficient radiogenic heating to maintain conditions for plate tectonics over long time periods. Plate tectonics has been proposed for ocean-covered exoplanets (Valencia et al., 2007), with changing plate velocities linked to increasing planetary mass. While some studies (Valencia et al., 2007; van Heck and Tackley, 2011; Tackley et al., 2013) suggest that the greater shear stresses and thinner plates thought to be associated with planets of higher masses will favor subduction (the movement of plates into a planetary interior) by decreasing the overall resistance to plate motion, others maintain that a stagnant lid (an immobile planetary crust) or episodic tectonic regime may be a more realistic assumption because of increased internal heating and decreased lower mantle viscosity (Stein et al., 2011) or because of a modeled reduction in the ratio of driving to resistive forces and increased fault strength under high gravity (O'Neill and Lenardic, 2007; Noack and Breuer, 2014). An upper limit of planetary mass for plate tectonics to function may be defined by the mass at which high pressures in the mantle increase rock viscosity and prevent plate tectonics (Noack and Breuer, 2014). Too low a mass results in early cooling, a phenomenon seen on Mars. The small size of the planet means that the crust long ago became solidified into a single stagnant lid (Breuer and Spohn, 2003). There is therefore no process for carbonate rocks to be subducted and to return CO2 to the atmosphere in the carbonate-silicate cycle. Planetary masses on the order of one to five Earth masses may be optimum for plate tectonics (Noack and Breuer, 2014). In summary, the presence of plate tectonics on a planet will therefore depend upon a range of factors including mass, age, the inventory of water, the initial internal heat, the distribution of oceans and continents, all of which cannot be readily predicted for any given planet in a given star system at the current time.
Although plate tectonics is crucial in driving the carbon cycle on Earth, other cycles have been proposed for ocean-covered planets where CO2-induced rock weathering is less effective. The circulation of CO2 or CH4 through clathrate reservoirs has been proposed as alternative carbon cycles (Levi et al., 2013).
Plate tectonics may play a wider role in sustaining continuous habitability on the surface of planetary bodies. The subduction of water-containing rocks on Earth is thought to contribute to the fluidity of the silicate mantle, which leads to efficient cooling of the mantle. Thus, the induced temperature gradient at the core-mantle boundary triggers convection in the outer core as well as chemical convection due to continuous freezing of the inner core. Both are essential for the maintenance of long-term operation of the planetary dynamo (Olson and Christensen, 2006) that generates a magnetic field and protects the atmosphere from the solar wind, reducing its loss.
It remains uncertain how much plate tectonics, or the lack of it, plays a role in the lack of a magnetic field on Mars. There is evidence for remnant magnetism in the southern hemisphere of the planet (Connerney et al., 2005; Langlais et al., 2010), suggesting an early magnetic field that later shut down. The lack of plate tectonics on Mars (Nimmo and Tanaka, 2005) may be one factor that accounts for an insufficient convection to generate a dynamo.
Plate tectonics, by delivering water into the mantle, lowers the melting point of rocks, contributing to the production of silica-rich rocks from which the buoyant continents are made (Campbell and Taylor, 1983). Thus, the existence of continental land masses may itself be linked to plate tectonics and the existence of water.
Despite these observations, the presence of sedimentary rocks on Mars that are ∼3.7 billion years old and their potential habitability (Grotzinger et al., 2014) suggest that planets can sustain liquid water and thus surface habitable conditions over billion-year timescales in the absence of plate tectonics. It is possible to imagine a scenario in which Mars or a Mars-like planet has hot-spot volcanism and an active planetary-scale hydrological cycle generating movement in CHNOPS elements, other cations, and anions through rock weathering and chemical disequilibria, allowing continuous habitable conditions to be sustained in local environments.
Plate tectonics may play a role in interior liquid water worlds. For example, plate tectonics on icy moons (Kattenhorn and Prockter, 2014) could provide a pathway for exogenously delivered surface elements and redox couples to be mixed into a deep interior ocean.
6.1.4. Magnetic fields
Magnetic fields act to protect atmospheres from being sputtered away (Lammer et al., 2008). Planetary bodies orbiting low-mass stars, such as G and K stars, may experience intense atmospheric sputtering if there is no magnetic field (Lammer et al., 2009), which leads to atmospheric loss. Early K and M stars have extreme UV radiation emissions 3–4 and 10–100 times higher, respectively, than G stars (Ribas et al., 2005); and for M stars, this intense radiation can persist for up to a billion years (Lammer et al., 2009). In these cases, a magnetic dynamo may be indispensable for improving the longevity of a dense planetary atmosphere that can support liquid water at the surface of the planet. Compounding this problem, in the case of M stars, planets become tidally locked in the habitable zone. The loss of planetary rotation can weaken the magnetic field (Grießmeier et al., 2004). These planets may lose their magnetic field early in their history, which would result in inclement surface conditions and limit the duration of continuous planetary habitability for surface-dwelling organisms. Models suggest that the size of a planet can also influence the magnetic field, with larger planets potentially sustaining more intense magnetic fields (Lammer et al., 2009).
In the case of Mars, despite the shutdown of a dynamo in its early history, it is apparent that liquid water was sustained on, or just beneath, the planetary surface for a long time after this event, as seen in evidence for catastrophic outflows, some of which may be just a few million years old (Carr, 1986, 1996; Tanaka, 1986; Burr et al., 2002; Neukum et al., 2010). Indeed, brines may even exist on the surface today (Rennó et al., 2009; McEwen et al., 2011). However, the weak magnetic field on Mars is thought to have partly contributed to the loss of atmosphere (Luhmann et al., 1992) and ultimately the low atmospheric pressure that prevents sustained liquid water on its surface today.
6.2. Astronomical factors
6.2.1. Planetary rotational and orbital characteristics
Planetary rotation plays a role in defining the strength of the magnetic field (Grießmeier et al., 2004). Planets that rotate slower, for example tidally locked planets, generally have smaller magnetic fields and are therefore more prone to atmospheric loss (Kasting et al., 1993; Barnes et al., 2008). Although tidal locking might cause atmospheric freeze-out on the dark side, if atmospheric circulation is sufficient, this catastrophe can be averted (Joshi et al., 1997; Joshi, 2003). The presence of cloud layers can reduce temperatures on the star-facing side, potentially leading to wider habitable zones for tidally locked planets and reducing the thermal contrast between the light and dark side of the planet (Yang et al., 2013).
Planetary obliquity can influence habitability. There are end-member conditions of obliquity and obliquity variations of significance. Terrestrial planets with high obliquities at the outer regions of the habitable zone with thick CO2 atmospheres have been shown by modeling to undergo partial atmospheric collapse, which would influence the effectiveness of the carbonate-silicate cycle, potentially disrupting habitability (Spiegel et al., 2009) and increasing the extremity of climatic cycles (Williams and Kasting, 1997). However, other modeling studies suggest that high-amplitude and high-frequency obliquity variations can suppress the effects of the ice-albedo feedback and thus increase the outer limit of the habitable zone (Armstrong et al., 2014). A lack of obliquity variation in some cases may be deleterious to habitability. Planets orbiting close to low-mass stars may experience “tilt erosion” (Heller et al., 2011) that leads to permanently low obliquities, which with tidal locking would lead to uniform climatic conditions on a planet with unknown consequences for habitability.
Planetary eccentricity can influence habitability (Williams and Pollard, 2002). Eccentricity will influence the stellar flux received by a surface liquid water world. Planets need not be continuously within the habitable zone for liquid water to exist on the surface. Bodies in eccentric orbits may sustain liquid water if the average stellar flux during the orbit is sufficient (Williams and Pollard, 2002; Dressing et al., 2010), although temperature and climate variations in the orbit may be extreme. In the case of highly eccentric orbits (∼0.5), high obliquity can stabilize a planetary climate against it dropping into Snowball (ice-covered) conditions (Dressing et al., 2010).
For planets with large eccentricities orbiting M stars, circularization of the orbit associated with tidal locking might move the planet to within the inner edge of the habitable zone within a billion years (Barnes et al., 2008), rendering a planet uninhabitable. Thus, eccentricities initially allowing for habitable conditions coupled with the tendency toward tidal locking can categorically modify habitability by removing a planet from the habitable zone entirely.
In the case of icy moons (interior liquid water worlds), eccentricity will determine the degree of internal tidal heating and thus the extent of interior liquid water.
6.2.2. Star type
The type of star around which a planet orbits will determine the distance of the habitable zone and thus the semimajor axis that the planet requires to host liquid water on its surface. For M stars, the proximity of the habitable zone can lead to tidal locking, with implications for the strength of the magnetic field and the required efficiency of atmospheric circulation to prevent atmospheric freeze-out (Joshi et al., 1997; Scalo et al., 2007; Tarter et al., 2007).
Stellar type might influence the habitability of a planetary surface with respect to specific metabolic groups, particularly phototrophs on surface liquid water worlds. The different spectral quality of M stars compared to G stars, for instance, may require phototrophs that process pigments with different absorption characteristics to those on Earth to match the spectrum (Kiang et al., 2007a, 2007b). Alternatively, novel forms of photosynthesis may be required to capture lower-energy photons (Wolstencroft and Raven, 2002), for example, from M stars whose spectral peak is shifted toward the red end of the spectrum.
The plasma flow from host stars controls the planetary energy budget, the atmospheric photochemistry, and the atmospheric mass loss from the outer layers of planetary atmospheres (Ribas et al., 2005; Claire et al., 2012; France et al., 2013; Lammer and Khodachenko, 2015). Stellar optical and infrared radiation increases slowly during stellar evolution. Ultraviolet radiation, including the Lyman-α emission line (121.6 nm), that dominates the UV spectrum of M dwarf stars controls photochemical reactions of H2O, CO2, CH4, and NH3 molecules (e.g., Lammer, 2013, and references therein). The XUV and X-ray radiation from host stars ionizes, dissociates, heats, and expands the upper atmospheres, driving atmospheric escape that is high during the host star's early period, which can last from tens to hundreds of millions of years for young solar-like G-type stars to billions of years for M-type dwarf stars.
The power of the stellar UV and X-ray fluxes depends on stellar activity, which decays with time and is related to the host star's rotation period that also decreases during the star's evolution (Lammer and Khodachenko, 2015). Thus, the evolution of an exoplanet's atmosphere and its habitability are strongly related to the evolution of its host star. Planets within the habitable zones of active flaring stars are exposed to intense stellar short-wavelength irradiation and extreme particle and stellar wind conditions over long time periods.
Planets are not required to be in single star systems to be habitable. Stable, habitable orbits can be sustained in binary star systems, both in S-type orbits, where the planet orbits one of the binary stars, and in P-type (circumbinary) orbits, where the planet orbits both stars (Benest, 1988; Whitmire et al., 1998; Dvorak et al., 2003; Haghighipour and Kaltenegger, 2013; Kaltenegger and Haghighipour, 2013).
6.2.3 The presence of a moon
It has been suggested that the Moon plays a fundamental role in habitability. Modeling study results suggest that it plays a role in stabilizing the obliquity of Earth (Laskar et al., 1993). However, more recent model studies question this conclusion and suggest instead that a moonless Earth, although exhibiting greater obliquity changes, would still maintain obliquity variations within a 20–25° range (Lissauer et al., 2012). Regardless, by stabilizing obliquity to some extent, the Moon might influence variations in terrestrial climate (Waltham, 2004, 2011). Caution should be exercised in interpreting the consequences of these processes for habitability. One might argue that varying obliquity would select for more generalist organisms capable of coping with frequent climatic change caused by other perturbations. We could just as well speculate that the stabilization of obliquity leads to the evolution of extinction-prone specialist organisms.
6.2.4. Impact events
Highly energetic large impacts have the potential to vaporize a planetary ocean and disrupt the course of biological evolution. Impacts are likely to be a universal problem for life, since no solar system–forming process is known that remains free of leftover debris from planetary accretion (Gomes et al., 2005). However, the scale and frequency of impacts will depend upon orbital dynamics and debris in any given system. Some systems, such as τ Ceti, may have more debris than our own Solar System (Greaves et al., 2004). Giant planets can cause gravitational scattering of small bodies, mitigating impacts (Laakso et al., 2006). It has been suggested that the presence of Jupiter in our planetary system reduced the perturbing effects of impacts by mopping up comets (Wetherill, 1994; Horner et al., 2010). However, giant planets may even increase the impact flux of asteroids and other objects (Horner and Jones, 2008, 2009) and in some general cases provide minimum protection (Laakso et al., 2006).
Except for extremely large impactors capable of sterilizing an entire planet (Maher and Stevenson, 1988; Sleep et al., 1989), it seems likely that, for many planets, the impact frequency may change the types of organisms for which the planet is habitable (e.g., by creating a selection pressure for thermophiles and hyperthermophiles; Sleep et al., 1989) but not alter its long-term continuous habitability for at least some organisms (i.e., for life in general). Impact events can even improve conditions for life in the surface and subsurface by enhancing the availability of habitat space and/or nutrients and energy (Cockell et al., 2012b). For interior liquid water worlds, impacts may deliver essential elements or redox couples or cause geochemical turnover, thus improving conditions for life.
The extent to which impacts will prevent the emergence of multicellularity or intelligence depends on their frequency or energy on surface liquid water worlds. This is influenced by the thickness of the atmosphere, with thicker atmospheres disrupting impactors more effectively than thinner atmospheres (Schaber et al., 1992). If sufficiently large impacts are frequent throughout a planetary history and always periodically boil the oceans over intervals of millions of years, the lack of sufficient oxygenic photosynthesis on the surface to generate adequate O2 may frustrate the formation of a highly oxidized atmosphere and thus the emergence of multicellular life and intelligence. Frequent large impacts could also limit the persistence of surface-dwelling complex animal-like organisms even if they did originate. The extinction of the dinosaurs at the Cretaceous-Paleogene boundary is an example of how the diversity of multicellular life can be frustrated by impacts (Schulte et al., 2010).
6.3. Summary
In summary, the eight examples of factors that influence continuous planetary habitability show that we still have much to learn about which factors merely modify the habitability of planetary bodies with respect to given organisms and which factors categorically affect their ability to host any habitable conditions. The interconnectedness between them (Fig. 5) shows that more advanced atmosphere-geosphere models of planetary bodies in their astronomical environments, coupled with empirical observations of exoplanets, will allow us to better model and constrain how these factors interact on any given planetary body to influence continuous planetary habitability.
7. Technological Habitability
Environments at local or planetary scale could be changed from uninhabitable to habitable by technological intervention (Fogg, 1995). At the planetary scale, this process is sometimes called terraforming, although to be habitable an environment does not strictly have to be made as similar to Earth as possible. A planet on which such schemes may be viable is Mars. There are a number of proposed means by which the surface of the planet could be made habitable. The introduction of chlorofluorocarbons (CFCs) or perfluorocarbons (PFCs) into the atmosphere could be used to terraform the planet (McKay et al., 1991). It would require ∼40 billion tonnes of CFCs to meet a required warming of ∼60°C. The release of CO2 to create an atmosphere of ∼100 mb could make the surface suitable for plants. Over ∼100,000 years, plants would generate O2, ultimately leading to a potentially human-breathable atmosphere.
Technological habitability can be achieved on the local scale, for example by covering a small area on Mars or the Moon with a greenhouse structure, providing the requirements for instantaneous habitability for certain organisms such as crop plants (Boston, 1981).
Technological habitability raises a number of questions that bring habitability within the social sciences. Is it ethically acceptable to deliberately modify a planet where the presence of life is uncertain (McKay and Marinova, 2001)? Is it ethically acceptable to deliberately inoculate an uninhabited habitat that might eventually host life from an existing biosphere or eventually host life yet to originate (Cockell, 2011), particularly if the entire planetary surface is uninhabited?
8. Conclusion
Habitability is a commonly used word. Its usage is usually vague, yet it lies at the heart of our assessment of the conditions that are required to make Earth a planet suitable for life over its lifetime, and the potential for life elsewhere. Habitability is an artificial definition. While this type of discussion cannot be considered a goal in itself, having a clear idea of what habitability means is at the heart of understanding the limits of biology on Earth, and our ability to discover other habitable worlds. In this review, we have provided some proposed definitions while at the same time providing a review of this subject for the scientist new to astrobiology.
Footnotes
Acknowledgments
This paper was written with support from the Science and Technology Facilities Council (STFC), Grant No. ST/M001261/1. H. Lammer acknowledges support from the Austrian Research Foundation FWF NFN project S11601-N16 “Pathways to Habitability: From Disks to Active Stars to Planets and Life,” as well as the related FWF NFN subproject S11607-N16 “Particle/Radiative Interactions with Upper Atmospheres of Planetary Bodies Under Extreme Stellar Conditions.” L. Noack has been funded by the Interuniversity Attraction Poles Programme initiated by the Belgian Science Policy Office through the Planet Topers alliance.