
Abstract
Earth is the most mineralogically diverse planet in our solar system, the direct consequence of a coevolving geosphere and biosphere. We consider the possibility that a microbial biosphere originated and thrived in the early Hadean-Archean Earth subseafloor environment, with fundamental consequences for the complex evolution and habitability of our planet. In this hypothesis paper, we explore possible venues for the origin of life and the direct consequences of microbially mediated, low-temperature hydrothermal alteration of the early oceanic lithosphere. We hypothesize that subsurface fluid-rock-microbe interactions resulted in more efficient hydration of the early oceanic crust, which in turn promoted bulk melting to produce the first evolved fragments of felsic crust. These evolved magmas most likely included sialic or tonalitic sheets, felsic volcaniclastics, and minor rhyolitic intrusions emplaced in an Iceland-type extensional setting as the earliest microcontinents. With the further development of proto-tectonic processes, these buoyant felsic crustal fragments formed the nucleus of intra-oceanic tonalite-trondhjemite-granitoid (TTG) island arcs. Thus microbes, by facilitating extensive hydrothermal alteration of the earliest oceanic crust through bioalteration, promoted mineral diversification and may have been early architects of surface environments and microcontinents on young Earth. We explore how the possible onset of subseafloor fluid-rock-microbe interactions on early Earth accelerated metavolcanic clay mineral formation, crustal melting, and subsequent metamorphic mineral evolution. We also consider environmental factors supporting this earliest step in geosphere-biosphere coevolution and the implications for habitability and mineral evolution on other rocky planets, such as Mars. Key Words: Origin of life—Mineral evolution—Bioalteration—Archean Earth—Crustal growth. Astrobiology 15, 922–939.
1. Introduction
C
For more than two decades, arguments have been made for a process called “bioalteration” or “bio-erosion” whereby microbes gain chemical energy and nutrients by dissolving basaltic glass in the subseafloor environment and form low-temperature alteration minerals and clays (e.g., Thorseth et al., 1992, 1995; Fisk et al. 1998; Furnes and Staudigel, 1999; Furnes et al., 2001; Banerjee and Muehlenbachs, 2003; Staudigel et al., 2008). It is important to point out here that our aim is not to prove that microbes were living in the Archean oceanic crust or that microbial bioalteration was operating in the Hadean–early Archean. The evidence for bioalteration in the early Archean subseafloor crust is highly controversial (see discussion in Grosch and McLoughlin, 2014). Rather, given the extensive evidence from the modern subseafloor environment and some sulfur isotope evidence for microbial sulfate reduction in early Archean subseafloor pillow lavas and the existence of a deep subsurface oceanic biosphere (McLoughlin et al., 2012), we explore and hypothesize on the implications of whether such a deep biosphere was active on early Earth. We discuss the implications of earliest geobiological processes and the possible role that subsurface microbial life may have had in dramatically shaping surface conditions and increasing mineralogical diversity on early Earth.
It is hypothesized that, in addition to well-known physical parameters that control the habitability of a planet (e.g., diameter/size, mantle temperature, distance from the sun, Fe/Mg ratio of the mantle, metallic core segregation), the early onset of fluid-microbe-rock interaction in subseafloor environments leads to formation of the first felsic crustal fragments and is critical to a planet becoming habitable for complex life. We further explore the geobiological interactions that may have occurred in the earliest mafic-ultramafic seafloor hydrothermal environments, which in turn led to increased mineralogical diversity with profound consequences for emerging tectonic processes, geochemical environments, and ultimately biological complexity to form the “Pale Blue Dot” we call home today. We compare and contrast the complexity of Earth's early coevolving geosphere and biosphere with possible scenarios on Mars and the implications for the extent of mineral evolution achieved by other rocky planets in our solar system and possibly beyond.
2. Earliest Geosphere-Hydrosphere Interaction and the Origin of Life
On modern Earth, formation of new continental crust occurs above subduction zones by dehydration melting of hydrothermally altered oceanic lithosphere. At mid-oceanic spreading centers, erupting mafic lava cools and hydrates through interactions with cold seawater along with hydrothermal fluid-rock interaction in the subseafloor beneath black smoker vents. On the Hadean Earth, 4.56–3.85 billion years ago, in situ oxygen isotope data from Hadean detrital zircons from Western Australia argue for cool liquid water on Earth's surface by 4.4–4.1 billion years ago (e.g., Bounama et al., 2001; Valley et al., 2002, 2014; Zahnle et al., 2007). As such, effective subseafloor hydrothermal alteration of the Hadean oceanic lithosphere was probably in operation at 4.4–4.1 billion years ago but may have been restricted to meteorite craters or relatively limited in some places possibly due to the lack of widespread proto-tectonic oceanic spreading processes or relatively shallow early oceans.
If relatively small landmasses of continental crust did exist, they were most likely short-lived on the surface and recycled back into the mantle, or they were destroyed by meteorite impacts, plumes, or possible proto-subduction. The oldest widely accepted, intact rocks preserved are the Acasta gneisses from the Slave Province in Canada dated at around 4.0 billion years old (Bowring et al., 1989; Bowring and Williams, 1999; Reimink et al., 2014).
The 3.85–3.70 Ga mafic oceanic lithosphere preserved in the Isua greenstone belt records evidence for hydrothermal alteration of basaltic oceanic crust (Nutman and Collerson, 1991; Maruyama et al., 1992; Nutman et al., 1997), suggesting that the onset of major interaction between the hydrosphere and geosphere may have occurred around 3.85 billion years ago. For example, preservation of the oldest paragneisses (metamorphic rocks of sedimentary protolith) dated at ca. 3.85 billion years old provides the first direct evidence for the onset of extensive hydrosphere-geosphere interaction forming sediments during the early Eoarchean (Nutman et al., 1997; Cates and Mojzsis, 2006; Mojzsis et al., 2014). Further evidence for extensive hydrosphere-geosphere interaction and proto-tectonic processes, involving shallow subduction dehydration melting along with tonalite-trondhjemite-granitoid formation (TTG-terrains), likely occurred on some parts of early Earth only during the mid-Archean around 3.45 billion years ago (e.g., Grosch et al., 2011, 2012). Prior to that transition, felsic continental crust, such as the tonalite sheets preserved in the ca. 4.02 Ga Acasta gneisses of Canada, most likely had its petrogenetic origin by direct melting above a plume in an Icelandic-type, extensional setting (Reimink et al., 2014).
The onset of subseafloor hydrosphere-geosphere interaction (e.g., in impact craters) on the Hadean to early Archean Earth was important for the emergence of life. Numerous workers have argued that life originated at hydrothermal seafloor vents on early Earth (e.g., Wächtershäuser, 1988a; Ferris, 1992; Russell et al., 1998; Lane et al., 2010; Russell et al., 2010; Sleep et al., 2011), because hydrothermal vents can provide a source of chemical energy for prolonged periods of time to sustain molecular reactions, provide a mechanism for the concentration of required reactants, and host numerous metals (Ni, Fe, Co) that could act as suitable catalysts for organic synthesis reactions to make the building blocks for life. For example, migration of hydrothermal fluids through the honeycomb pore networks of travertine-type vent structures on the seafloor may have concentrated nucleotide molecules and ultimately nucleic acid chains and thus provided conditions favorable for manufacturing the building blocks for life (Russell and Hall, 2009; Hazen and Sverjensky, 2010). As such, the onset of submarine hydrosphere-geosphere interaction on the Hadean to early Archean Earth may have been responsible for producing the first biogeochemical factories of life, allowing a transition from a geochemical early Hadean Earth to a biogeochemical early Archean world.
Although many workers have argued that hydrothermal vents on the seafloor would make a good prebiotic address for life, the possibility that life may have emerged in metabasaltic glass beneath hydrothermal vents has not been previously considered. A rich variety of hollow and partially mineralized complex microtubular morphologies and granular textures have been reported from the in situ oceanic crust in low-grade metavolcanic glass from recent subseafloor environments (e.g., Thorseth et al., 1992, 1995, 2001; Furnes and Staudigel, 1999; Banerjee and Muehlenbachs, 2003; Fisk and McLoughlin, 2013). Many workers have advocated a biological origin for these microtextures as the consequence of microbes etching volcanic glass for nutrients and/or an energy source in a process called “bio-erosion” or bioalteration (Thorseth et al., 1995; Fisk et al. 1998; Furnes and Staudigel, 1999; Furnes et al., 2001; Banerjee and Muehlenbachs, 2003; Staudigel et al., 2008; Mayhew et al., 2013). Organic remains and DNA found in the hollow microtextures have been used to support the idea that microbes formed the microtunnels and left behind decayed organic remains (Torsvik et al., 1998; Banerjee and Muehlenbachs, 2003; Staudigel et al. 2008; Wacey et al., 2014). In addition, geochemical and genetic evidence from subseafloor lavas points to the existence of an active and extensive recent subseafloor biosphere (Bach and Edwards, 2003; Rouxel et al. 2008; Lever et al., 2013). However, several abiotic mechanisms have also been suggested to explain the possible origins of microtunnels in metavolcanic glass (e.g., Lepot et al., 2011; McLoughlin et al., 2012; McLoughlin and Grosch, 2014). For example, elevated fluid pressures in the subsurface can result in an abiotic mechanism where small metal-rich inclusions such as pyrite are propelled through the substrate, leaving behind a tubular trail (also known as ambient inclusion trails). Low-grade hydrothermal alteration at depth and elevated fluid pressures during diagenesis may cause crystalline (e.g., tiny pyrite or other sulfide crystals) or organic inclusions to migrate and thus create hollow tubular trails or complex microtunnels with a terminal crystal at one end (e.g., Lepot et al., 2011; McLoughlin et al., 2012). These tubular microtunnels often contain palagonite, zeolite, and clay minerals (e.g., Banerjee and Muehlenbachs, 2003; Staudigel et al., 2008; Izawa et al., 2010; Knowles et al., 2013; Wacey et al., 2014). For example, microtubular morphologies in volcanic glass have been found to contain fine-grained clay minerals (e.g., smectites, possibly montmorillonite), zeolites, and organic molecules (e.g., Torsvik et al., 1998; Staudigel et al., 2008; Knowles et al., 2013; Wacey et al., 2014). These alteration minerals, which formed by infiltrating fluids, would have provided important loci for prebiotic reactions (e.g., Wächtershäuser, 1988b; Weber, 1995) with chiral template surfaces for sorbtion of organic molecules and metal catalyst reactants. Rocks, minerals, and water involved in mafic-ultramafic rock hydrothermal alteration processes are potential sources of C, H, O, and N for organic molecule formation. Carbon dioxide (CO2 gas) and carbonates would provide C and H2. Water would provide O and H, whereas ammonia would have been generated by reduction of nitrogen-rich volcanic gases in oceanic hydrothermal systems on early Earth (Brandes et al., 1998, 2008; Smirnov et al., 2008). Low-temperature serpentinization would provide additional H2(g). It is likely that CO2, CO, and formaldehyde that formed under hydrothermal conditions would have been available as building blocks for larger organic compounds (e.g., Cody et al., 2000, 2004; McCollom, 2013; Shrenk et al., 2013). Abiotic microtunnels in subseafloor metavolcanic glass containing clay minerals and zeolites would thus have provided a favorable environment for concentrating organic and other prebiotic reactants required in the assembly of complex biomolecules during early prebiotic synthesis.
A range of different microenvironments have been argued to be favorable for the origins of life, including mineral surfaces and pore spaces between sulfide minerals in seafloor hydrothermal vent structures (Wächtershäuser, 1988b; Ferris, 1992; Russell et al., 1998; Hazen, 2005, 2006; Lane et al., 2010; Russell et al., 2010), zeolites in vesicles within floating pumice rafts (Brasier et al., 2011), and the internal structure of weathered feldspar grains on continents (Smith, 1998; Parsons et al., 1998). In addition, we contend that abiotic microtunnels in volcanic glass containing clay and zeolite minerals as well as organics could also be considered ideal sites for prebiotic synthesis, rather than only representing the preserved remains of microtunneling microbes. The metabasaltic subseafloor beneath vents represents a significantly greater reactive volume compared to the vent chimneys themselves. Conditions for concentration of biomolecule reactants in mineralized microtunnels of the subseafloor would have been possibly more suitable than the dilute conditions at exposed seafloor surface vents or in pumice floating at the early ocean surface. Fluid-rock ratios, as well as redox, chemical, and pH gradients, may all have been suitable at these subseafloor prebiotic reactor microsites. Serpentinization in the mafic to ultramafic suboceanic crust would have played a major role in providing the necessary redox disequilibrium for the origin of life in the subseafloor environment (Shrenk et al., 2013). Hydrothermal serpentinization processes in ultramafic aquifers would have been a common feature of the early Archean subseafloor and transitional upper mantle environments. Low-silica ultramafic rocks are rich in olivine and pyroxene. Low-temperature water-rock reactions result in the oxidation of ferrous iron from olivine and pyroxene, resulting in the precipitation of ferric iron in magnetite (Fe3O4) and other serpentine minerals, and in the release of diatomic hydrogen (H2) (e.g., Mayhew et al., 2013). At low temperatures (< ∼150°C), the serpentinization reaction results in alkaline conditions with extremely high pH, commonly above 10 (e.g., Russell, 2007). The aqueous fluids in serpentinization zones can be highly enriched in reductants and in oxidants. For example, typically 10–30 mM H2, with significant CO2. The zone of basalt alteration and serpentinization is highly dynamic, with constant circulation of ocean water through a complex system of cracks and fissures. That water is modified significantly by fluid-rock interactions, so it is in disequilibrium with the zones where metabolism is postulated to emerge. A number of authors have demonstrated synthesis of compounds to C6 and beyond in analogous environments, for example through FT synthesis and hydroformylation (e.g., Huber and Wächtershäuser, 1998; Brandes et al., 2000; Cody et al., 2000). The combination of H2 and CO2 or CO under highly reducing conditions leads to formation of methane and other hydrocarbons through Fischer-Tropsch-type synthesis (Shock et al., 1995; McCollom and Seewald, 2001; Charlou et al., 2002; Proskurowski et al., 2008; McCollom, 2013). These conditions would support abiotic organic synthesis in fractured mafic and ultramafic aquifers within the suboceanic crust of early Earth. The onset of subseafloor hydrosphere-geosphere hydrothermal interaction on the Hadean–early Archean Earth would have been important, not only for the start of hydration of the earliest oceanic crust but also for the fluid-rock alteration processes that formed the first subseafloor prebiotic reactor sites for the emergence of earliest microbial life.
3. Onset of Geosphere-Biosphere Interactions
Observations of modern oceanic crust have been used to argue that subseafloor volcanic glass may have been a likely substrate for early microbial life to thrive, as volcanic glass can provide energy and nutrients for chemolithotrophic microbes (e.g., Thorseth et al., 1992, 1995; Staudigel et al., 2008, and references therein). Several studies have attempted to distinguish between abiotic versus possible biotic alteration of basaltic glass (e.g., Thorseth et al., 1992; Staudigel and Furnes, 2004; Staudigel et al., 2008). In the modern oceanic crust, abiotic alteration of basaltic glass is typically assigned to the formation of a yellow-brown amorphous phase known as gel palagonite formed along angular glass fragments. This gel is the earliest abiotic alteration product, which is followed by a transition to fibropalagonite that hosts clay minerals, zeolites, and iron-oxy-hydroxides (Peacock, 1926; Stroncik and Schmincke, 2001). On the other hand, biotic alteration or “bioalteration” of subseafloor basaltic glass by microbes has been envisioned to form a wide range of microscopic granular and tubular morphologies (Thorseth et al., 1992, 1995; Staudigel et al., 1998, 2006; Furnes et al., 2001a; Banerjee and Muehlenbachs, 2003). These granular features are the most abundant morphological variety found on the subseafloor today and typically occur along microscopic fractures and veins. They consist of dense clusters of micron-sized, near-spherical voids or pits typically filled with authigenic minerals, such as clay minerals (smectites), zeolites, and sulfides (Fig. 1a). Granular alteration, as shown in Fig. 1, can form irregularly on both sides of a microvein or crack. Singular near-spherical pits may coalesce to form subspherical agglomerates on either side of a vein (Fig. 1a). Coccoid organisms, biofilms, and cell-like structures associated with granular textures in volcanic glass have all been used to support their origin as a direct consequence of etching of volcanic glass by microbes during possible bioalteration (Thorseth et al., 1992, 1995; Cockell, 2011).
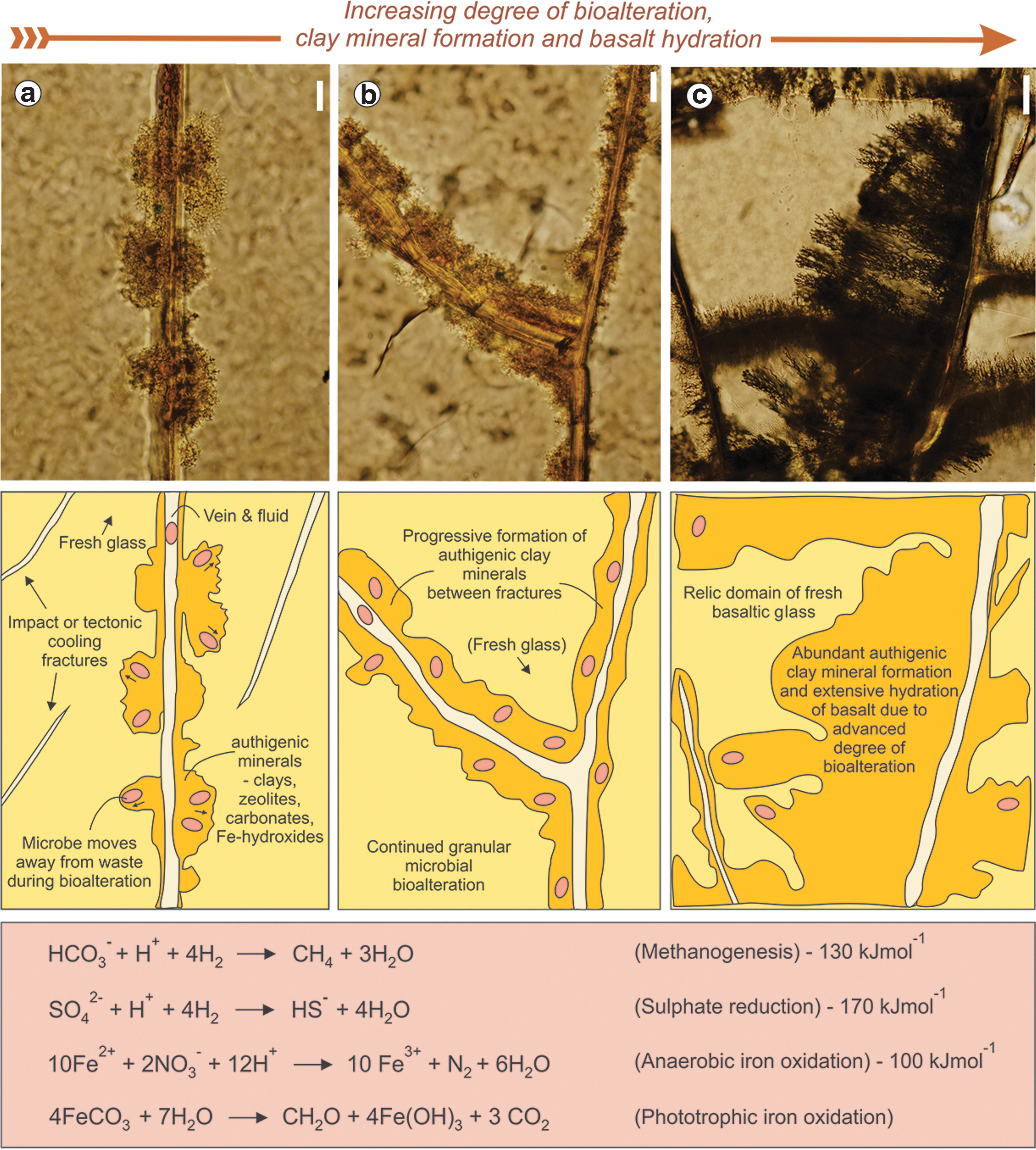
Progressive stages of alteration of subseafloor volcanic glass from the in situ oceanic crust: (
In the previous section, we entertained the hypothesis that life may have emerged in metavolcanic, partially mineralized microtunnels in the Hadean-Archean subseafloor environment. Once formed at these microscopic prebiotic reactor sites, microbial life would have sought to migrate into new subsurface metabasaltic niches, probably along veins and fractures. It is also possible that earliest microbial life was introduced by meteorites into the basaltic subsurface (lithopanspermia, e.g., Hoyle and Wickramasinghe, 1981; Olsson-Francis and Cockell, 2010). The Hadean–early Archean basaltic subsurface would have represented a permeable water-saturated regolith that was habitable, with extensive formation of fractures and reactivation along fault zones. (e.g., Sleep, 2012). Increased subsurface permeability would have resulted from heavy meteorite bombardment after the Moon-forming impact event (e.g., Cockell, 2006; Zahnle et al., 2007; Sleep et al., 2014). Several other physical processes on early Earth would have promoted subseafloor basaltic fracturing, faulting, and permeability. These mechanisms may include chemical brecciation and expansive cracking with crystal growth (Sawkins, 1969) and cracking related to thermal contraction of fresh rock during high-temperature hydrothermal alteration (Lister, 1974). Exothermic hydration reactions related to subseafloor serpentinization can also have rheological effects of lithospheric strength causing weaknesses, normal faulting, and deep seawater fluid circulation (e.g., Escartin et al., 1997; Lowell and Rona, 2002; Mével, 2003). Major tidal stresses and earthquakes due to a closer Moon on the Hadean–early Archean Earth would have also promoted major faults, fracturing on all scales, and increased subsurface permeability and thus habitability (Sleep, 2012; Sleep et al., 2014).
Abiotic hydrothermal alteration would have hydrated the oceanic crust along fractures, cracks, and faults forming palagonite. The permeable subsurface regolith formed by early Earth geophysical processes would have aided early microbial life (introduced extraterrestrially from meteorites or from within the oceanic crust) to migrate and thrive in various subseafloor basaltic niches by fluid flow. Geophysical forces and abiotic chemical fracturing mechanisms may have facilitated focused alteration and hydration of basalt restricted to narrow veins and faults. However, in this hypothesis it is argued that microbial bioalteration, if operating, would have had a much larger effect on increasing the hydration of basalt between fractures. For example, Staudigel and Furnes (2004) argued that progressive granular alteration along a fracture vein dramatically increases roughness along that fracture and thus increases the available surface area for fluid migration and alteration by a factor of approximately 200. Thus, abiotic chemical fracturing mechanisms may have been complementary to promoting extensive bioalteration and hydration of basalt.
In Fig. 1, the complementary mechanisms of abiotic fracturing (e.g., meteorite impacts, proto-tectonic forces, microfaulting, and hydraulic brecciation) and biological exploitation of such fracturing during bioalteration are illustrated. In Fig. 1, we illustrate with samples from the in situ oceanic crust possible stepwise stages in granular bioalteration that may have formed due to microbes progressively altering metavolcanic glass. Figure 1a shows a possible early stage of globular coalescing of granular textures on either side of a palagonite vein or fracture. Figure 1b illustrates more extensive formation and agglomeration due to further microbe-fluid-rock interaction in the subsurface, whereas during advanced stages of bioalteration an extended biogranular alteration network associated with clay minerals, zeolites, and organics develops extensively in the volume between veins. The corresponding cartoons in Fig. 1 illustrate a fluid-rock-microbe model forming granular bioalteration with increasing degrees of bioalteration and hydration of basalt with authigenic clay minerals (Thorseth et al., 1992, 1995). Microbes infiltrate with seawater and colonize the fresh subsurface basaltic glass along fractures and cracks (Fig. 1a). In this model, microbes at the onset of bioalteration are argued to excrete substances (e.g., acids, chelating agents, exo-enzymes, and molecular organics) that result in dissolving the glass. As the microorganisms dissolve the glass, they use Fe2+ or Mn as nutrients and/or as a source of energy. Excess dissolved glass components and waste in a thin biofilmlike solution have to be removed by exiting fluids from the space between the microbe and the glass. As the excess dissolved components and waste products are transported away, the microbes also move away from the sites where waste was produced to find other glassy areas with fresh nutrients and energy sources. As the solution carrying dissolved waste components flows out from the granular cavity, the changing pH conditions due to mixing with neutral seawater result in the precipitation of authigenic clays, zeolites, and Fe-hydroxides (e.g., Staudigel et al., 2008). A pH gradient across the granular texture also results in clay minerals precipitating inside the granular microstructure. This process of microbial dissolution of glass continues to form granular bioalteration textures and authigenic clays, and the microorganisms move away from microdomains where waste products are accumulating to new niches in search of nutrients and energy from fresh basalt (Fig. 1b). The final cartoon panel in Fig. 1c depicts a high degree of basalt alteration as a result of microbial bioalteration. Granular bioalteration and formation of extensive authigenic clay minerals spread extensively in the glass volume between fractures. The area (volume) of alteration and hydration of basalt caused by microbial bioalteration is argued to be orders of magnitude higher than that produced purely by abiotic hydrothermal processes along meteorite and/or chemical brecciation along fractures or veins. A list of chemical reactions that are important sources of metabolic energy (e.g., anaerobic iron oxidation) used by early subsurface lithoautotrophic microbes during glass bioalteration is also provided in Fig. 1 (after Edwards et al., 2005; Cockell, 2011).
4. Role of Serpentinization in Early Earth Mineral Evolution
Hydrogen production from low-temperature alteration of mafic and ultramafic rocks provided a continuous source of redox potential for early life to be sustained. For earliest life, hydrogen (H2) was probably one of the most important electron donors arising from alteration of Fe2+ minerals in mafic-ultramafic rocks (e.g., Mayhew et al., 2013; Shrenk et al., 2013). Serpentinization and low-temperature alteration of ultramafic komatiites, basaltic komatiites, and tholeiitic basalts would have collectively fueled oceanic subsurface microbial communities, such as methanogens. Recent experimental studies have shown that serpentinization involving anoxic fluids at temperature conditions of between 55°C and 100°C can produce enough hydrogen to support modern-day oceanic microbial communities (e.g., Mayhew et al., 2013). Similar models have been invoked for the role of serpentinization in sustaining early subsurface chemolithoautotrophic communities on early Earth and Mars (e.g., Grosch et al., 2014, and references therein). Analysis of uranium (U) and potassium (K) content in palagonite combined with numerical modeling has recently shown that significant amounts of hydrogen can also be produced during low-temperature alteration of tholeiitic basalts (Türke et al., 2015, in this issue). The H2-rich suboceanic hydrothermal environments on early Earth were highly alkaline with pH of around 10–12, conditions favorable for sustaining bioalteration in altered subseafloor ultramafic and metabasaltic rocks (e.g., Russell, 2007). Serpentinization and associated redox gradients therefore not only played a major role in biosynthesis of organic compounds but also provided H2 as a source of chemical energy to support microbial bioalteration of basaltic and komatiitic basaltic glass in fractured subsurface hydrothermal environments. In our proposed model for the earliest onset of mineral evolution during subseafloor microbial alteration of crust, an open system is invoked for bioalteration microtextures (e.g., granular, tubular, and associated veins) that allows for the exchange of aqueous species. An open-system environment for the emergence of life (e.g., fracture network with microveins in the mafic/ultramafic subseafloor hydrothermal system) would have been much more conducive to biological processing of available free energy from the onset. This allows for excess chemical energy, continuous replacements of nutrients, and flushing of waste by open system fluid flow. Earliest microbial life would have thrived in such a hydrogen-rich environment. In addition to methanogenesis, other coexisting communities exploited other metabolic pathways such as microbial sulfate reduction. Similar to that observed in oceanic environments today, microbes thrived and exploited serpentinization and chemical disequilibrium that existed in the subsurface of early Earth. In turn, microbes would therefore have catalyzed alteration reactions in the metabasalt environments on early Earth, catalyzing clay mineral production in the oceanic crust and potentially accelerating melting and felsic crust production such that the earliest fragments of terrestrial continental lithosphere were formed.
5. Production of First Felsic Crust in Mineral Evolution
In the modern in situ oceanic crust, numerous lines of evidence exist for the presence of a deep subseafloor microbial biosphere (see Table 1 and references therein). Further back in the geological record, the evidence for bioalteration becomes less pronounced possibly due to a lack of preservation potential of textural and chemical biosignatures. A summary of the evidence for and against the presence of subseafloor bioalteration, including evidence in ophiolites (accreted sections of oceanic crust on continents) and Archean greenstone belts, is provided in Table 1. If granular microtextures and clay mineral formation in subseafloor metabasaltic glass are the result of bioalteration, this implies that microbes may play a major role in the alteration of oceanic crust today. Similarly, sulfur isotope data indicate the possible role of microbial sulfate reduction in the early Archean subseafloor environment (McLoughlin et al., 2012; Table 1). Considering these observations, it is hypothesized that microbial activity in the Hadean-Archean oceanic subseafloor may have similarly facilitated the alteration of basaltic volcanic glass. If the observations of Staudigel and Furnes (2004) are correct, bioalteration would have dramatically increased the surface area around veins by a factor of up to 200, allowing for extensive fluid flow, as well as the formation of alteration minerals, including smectites and other clay minerals, zeolites, and sulfides. Consequently, while abiotic vein-related alteration of the suboceanic lithosphere is more spatially limited, biotic alteration has the major potential to accelerate clay formation in volcanic glass of the oceanic lithosphere on a much larger scale. It is important to note here that rates of bioalteration and volume of bioaltered crust may possibly have been lower in the Hadean–early Archean (see full discussion of various planetary mineral evolution models in Section 6). However, it is postulated that microbes would have played a role in the alteration of oceanic crust. The implications of this accelerated alteration for crustal processes on early Earth are not trivial: microbes played a major role in promoting hydration of the Hadean-Archean oceanic lithosphere, possibly on a scale not seen prior to the first appearance of microbial life on our planet. Effective hydration as a direct consequence of this interaction between the geosphere, hydrosphere, and newly formed biosphere would lead to more efficient dehydration melting to form the earliest evolved felsic crust on our planet. We suggest that it is possible that suboceanic microbes were the earliest architects of the first sialic, continental crust.
References: (a) Thorseth et al., 1995; (b) Cockell, 2011; (c) Lever et al., 2013; (d) McLoughlin et al., 2011; (e) Edwards et al., 2005; (f) Staudigel et al., 2008; (g) Fisk et al., 2013; (h) Furnes et al., 2008; (i) McLoughlin et al., 2010; (j) McLoughlin et al., 2012; (k) Grosch and McLoughlin, 2014.
Hydrothermal vents at impact sites were commonplace on Hadean Earth, and microbes may have lived under those conditions during heavy impact bombardment (Cockell, 2006; Abramov et al., 2013). Consequently, granular bioalteration may have occurred locally at these impact sites. Extraterrestrial carbon or organic precursors for the origin of life may also have been supplied at such sites for prebiotic reactions (Chyba and Sagan, 1992). However, during secular cooling near the Hadean-Archean boundary, earliest mid-ocean spreading centers started to form, giving rise to new hydrothermal vent sites at spreading centers and an additional venue for microbial life to thrive (Fig. 2a). Granular textures may have formed beneath mid-oceanic spreading centers as a result of microbially mediated alteration of subseafloor basaltic glass (Fig. 2a, ii–iii). Advanced stages of microbial granular microtexture formation would have promoted extensive alteration and clay mineral formation (Fig. 2a, ii–iii). Progressively deeper into the mafic-ultramafic oceanic lithosphere, higher metamorphic grades from subgreenschist to greenschist facies conditions would have been achieved more easily by microbe-facilitated rock hydration (Fig. 2a, iii–v). Similarly, conditions further away in short-lived and aging crust would also have induced higher degrees of alteration. Extensive hydration of the oceanic crust would have effectively promoted dehydration melting of the late Hadean–early Archean oceanic lithosphere, thus forming the earliest stages of felsic crust accumulation (Fig. 2a, vi–vii). This crustal build-up at the earliest stages would probably have occurred above mantle plume heads but also in other proto-tectonic/geodynamic settings, such as crustal delamination, shallow subduction, or whole-scale convective recycling of oceanic crust into the mantle to give buoyant felsic proto-plutons (Fig. 2). Oxygen isotope evidence from Acasta gneiss zircons is consistent with the earliest evolved crust, which may have formed in an Icelandic-type oceanic spreading center by 4.02 billion years ago (Reimink et al., 2014). In summary, by facilitating major alteration of oceanic lithosphere at early spreading centers, microbes may have played a major role in promoting dehydration melting and generating the first felsic rocks on early Earth.
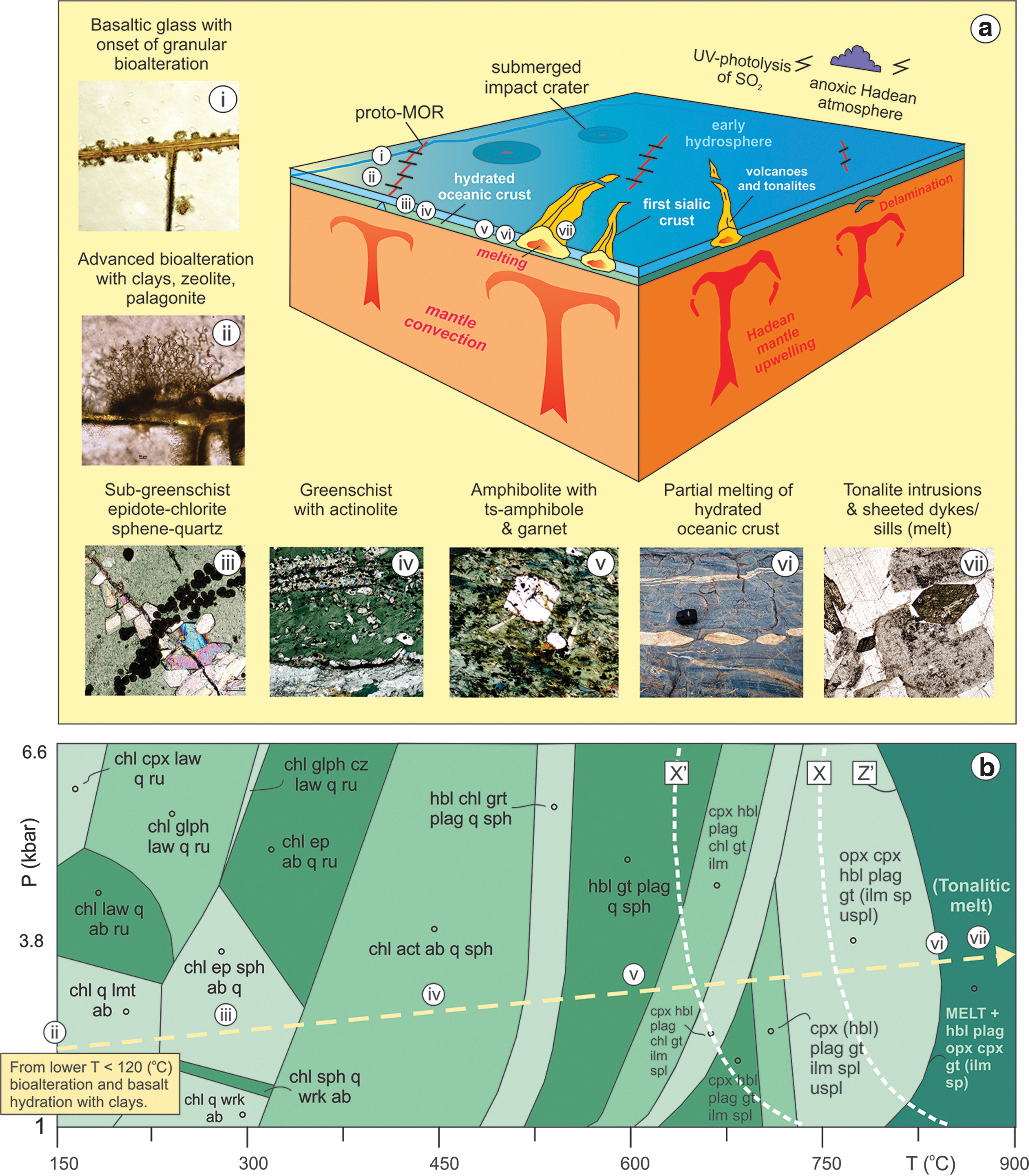
(
The progressive hydrothermal alteration and metamorphic mineral evolution with increasing temperature prior to “wet” melting of Archean tholeiitic basaltic crust can be modeled thermodynamically in pressure-temperature space (Fig. 2b). This has been performed by using a typical early Archean tholeiitic pillow basalt composition (representing early Archean oceanic crust) from the Barberton greenstone belt of South Africa (BGB-41a; SiO2 = 50.30, Al2O3 = 13.68, TiO2 = 1.95, FeO = 19.68, MgO = 7.14, CaO = 2.23, Na2O = 0.28). This mineral phase diagram calculated in the Na-Ca-Mg-Al-Si-Ti-H2O chemical system with the thermodynamic modeling software PERPLEX (Connolly, 2005) depicts mineral stability fields across increasing metamorphic facies from subgreenschist to upper amphibolite to melting conditions for the specific Archean bulk rock composition given above. For example, at point iii the assemblage chlorite-epidote-sphene-albite-quartz represents subgreenschist facies conditions, whereas the assemblage chlorite-actinolite-albite-sphene-quartz (iv) represents the pressure-temperature field of between around T = 300°C and 500°C for the greenschist facies (Fig. 2b). Mantle temperatures are argued to have been higher in the Archean eon at temperatures above ca. 1500–1550°C (see Johnson et al., 2014, and references therein). We therefore infer a flat geothermal gradient for melting. The dashed arrow in Fig. 2b thus indicates the direction of metamorphism with increasing temperature along a low-P, high-T (flat) Hadean–early Archean geothermal gradient, labels ii to vii marking pressure-temperature points and metamorphic facies changes along this geothermal gradient (e.g., above a mantle plume head). These labeled stability fields correspond to the hydrous metamorphic mineral assemblages in the petrographic images ii to vii provided in Fig. 2a.
The calculated pressure-temperature phase diagram indicates metamorphic stability fields defined by steep reaction curves. For example, the transition from the greenschist facies to the amphibolite facies marked by the appearance of hornblende (amphibole), garnet, and plagioclase is calculated from the bulk rock composition to occur at around T = 500°C (Fig. 2b) for the Archean basaltic rock sample. The reaction curve labeled Z’ marks the basalt “dry” melting reaction and represents the thermal decomposition of hornblende in the presence of quartz, to produce felsic tonalitic (trondhjemite-granitoid) melt at conditions of around T = 830°C. However, in this study we hypothesize hydration of metabasaltic crust by microbially mediated bioalteration and clay formation, with subsequent “wet” dehydration melting (loss of water from extensive sheet silicate formation during melting). Consequently, reaction X–X’ marking the positions of the experimentally determined wet tonalite solidus (Piwinskii, 1968; Johannes, 1978; Wyllie and Wolf, 1993) is more relevant for altered oceanic crust. For example, wet melting involving the decomposition of hornblende + plagioclase + quartz in the presence of water (derived from dehydration of earlier formed clay/sheet silicates in the protolith basaltic rock), under water activity conditions of around 0.5, results in a significant lowering of the melting temperature from T = 830°C to T = 750°C (position of reaction X). However, we consider a scenario that involves early extensive microbial bioalteration (Fig. 2b, ii) of the oceanic crust, dramatically increasing clay mineral formation and alteration surface area. Such an extensively altered basaltic protolith would produce plenty of water from thermal dehydration and create hydrous melting conditions with a H2O activity of 1. Conditions of melting would therefore, in this case, be dramatically lowered to around T = 660°C (reaction X’). An order-of-magnitude increase in the degree of hydration of a specific volume of Archean basaltic crust due to bioalteration could result in lowering of the melting temperature by around 200°C to form felsic tonalite crust. This extensive microbially mediated alteration and clay mineral hydration of the oceanic crust would have promoted extensive dehydration melting at lower temperatures, in a wider range of geodynamic thermal regimes, and resulted in more volumetrically extensive formation of felsic proto-continental crust. This may have occurred above hot mantle plume heads on the Hadean–early Archean Earth, possibly also during crustal thickening processes or when the microbially mediated oceanic crust was subducted into the mantle and melted to form TTG island arcs on early Earth as discussed further below (see Fig. 3).

Coevolution of the geosphere and biosphere before the Great Oxidation Event from the onset of possible subsurface bioalteration at around 4.0 billion years ago until modern-style tectonic environments operating on Earth's surface at ca. 3.0 billion years ago. A space-time vector (green arrow) indicates an increase in metamorphic and igneous mineral diversity on Earth's surface, progressive and stepwise increase in planetary habitability, and an increase in the complexity of the coevolving geosphere and biosphere. Panels on the right-hand side of the figure are pressure-temperature (PT) phase diagrams, with shaded red areas depicting stability fields of various metamorphic mineral assemblages that were realized in early Earth's crustal lithosphere at different times between 4.0 and 3.0 billion years ago. (
6. Hadean–Early Archean Crustal Growth and Geosphere-Biosphere Coevolution
The timing and nature of the first proto-tectonic and geodynamic processes that led to the generation of continental crust on early Earth are highly debated topics (e.g., Van Kranendonk et al., 2009; Grosch et al., 2011, 2012; Ganne et al., 2012; Cawood et al., 2013). Various geodynamic models range from modern-style plate tectonic processes operating as early as the Hadean-Archean boundary (e.g., Furnes et al., 2014) to plume-dominated processes as late as 3.45 billion years ago (Van Kranendonk et al., 2009). However, recent work has shown that during the late Hadean Eon dehydration melting probably occurred above mantle plume heads at Icelandic-type spreading centers (Reimink et al., 2014) in the absence of modern-style tectonics. We hypothesize that, if microbially mediated hydrothermal alteration of oceanic crust was operating in late Hadean–early Archean meteorite impact sites and at proto-oceanic spreading centers, then this process would potentially have had major implications for the formation of the earliest felsic crust, even prior to, and independent of, the onset of proto-tectonic processes.
Once microbial life had a foothold in the early subseafloor environment, it would have accelerated subseafloor metavolcanic alteration and hydration. Redox couples in basaltic glass produced by volcanic and hydrothermal processes would have provided a spectrum of energy sources on the anoxic early Earth (Jakosky and Shock, 1998; Edwards et al., 2005; Canfield et al., 2006; Cockell, 2011). Studies on modern-day basaltic glass from Icelandic basalts containing about 13% total Fe indicate that every 1 m3 of basaltic glass can potentially sustain about 2.5 × 1016 anaerobic Fe-oxidizing bacteria if all Fe is released by water-rock interaction. By using Icelandic basaltic glasses, the release rate of major cations from basaltic glass as a function of silica content at 25°C would correspond to the production of 2.8 × 1010 cells m−3 yr−1 (Wolff-Boenisch et al., 2004). Bach and Edwards (2003) estimated the amount of Fe-oxidizing bacteria biomass that can be generated from approximately 3 km3 yr−1 of basalt (7% Fe in rocks) at modern spreading centers to be approximately 1.6 × 1010 mol carbon yr−1. On early Earth, oceanic crustal spreading rates would have been higher due to significantly higher mantle geothermal gradient (Johnson et al., 2014), so biomass production in oceanic crust may have been higher, possibly at a maximum due to larger volumes of available basalt being generated.
At ca. 4.02 billion years ago, extensive biologically mediated alteration of the oceanic crust due to microbes would have resulted in dehydration melting of basalt above mantle plume heads, generating early tonalite sheets and intrusions (Figs. 2 and 3a). These early, evolved or differentiated felsic terranes, which probably represented Earth's earliest continental entities, would have been much more buoyant than the underlying and surrounding oceanic crust and as a consequence would remain afloat on Earth's surface or possibly be recycled back into the mantle by plumes or meteorite impacts. In Fig. 4, estimates of continental crustal growth are provided across Earth's history using the model of Dhuime et al. (2012) based on hafnium and oxygen isotope data from zircons. These workers argued that modern-style tectonics (high-pressure metamorphism, steep subduction of cold slab, etc.) only started to operate around 3.0 billion years ago. Prior to this time, plume dynamics and proto-tectonic processes dominated and would have been involved in the formation of felsic continental crust. Continental crustal growth rates before 3.0 Ga are estimated to be around 3.0 km3 yr−1, whereas after that time net growth rates decreased to 0.8 km3 yr−1 (Dhuime et al., 2012). In this hypothesis, we postulate that microbially mediated alteration of subseafloor basaltic crust may have played a major role in catalyzing felsic crust generation on early Earth between ca. 4.4 and 3.0 Ga. By ca. 3.0 billion years ago, approximately 70–75% continental crust was already produced (Dhuime et al., 2012) in the absence of modern-style tectonics, and it is possible that subseafloor microbes may have been directly responsible for, or played a major role in, producing high felsic crustal growth rates. Pioneering microbes introduced from meteorites or originating from within the subseafloor would have thrived in the basaltic environment, using Fe oxidation and hydrothermal reactions for energy, which would have accelerated extensive basalt hydration and clay mineral formation. This would have represented a major habitable zone to exploit for these early microorganisms. Meteorite impact fracturing, major tidal stresses, and earthquakes (Sleep, 2012) created numerous and deep fractures in the subseafloor environment in which microbes would have colonized and ultimately promoted further bioalteration of basaltic glass for the first time and on an unprecedented scale in Earth's history. In the absence (or limited operation) of modern-style tectonic crustal recycling and reworking and under heavy impact bombardment, approximately 45% of Earth's total volume of continental crust was already produced by ca. 3.5 billion years ago (see Fig. 4), with microorganisms most likely playing a major role in felsic crust generation.
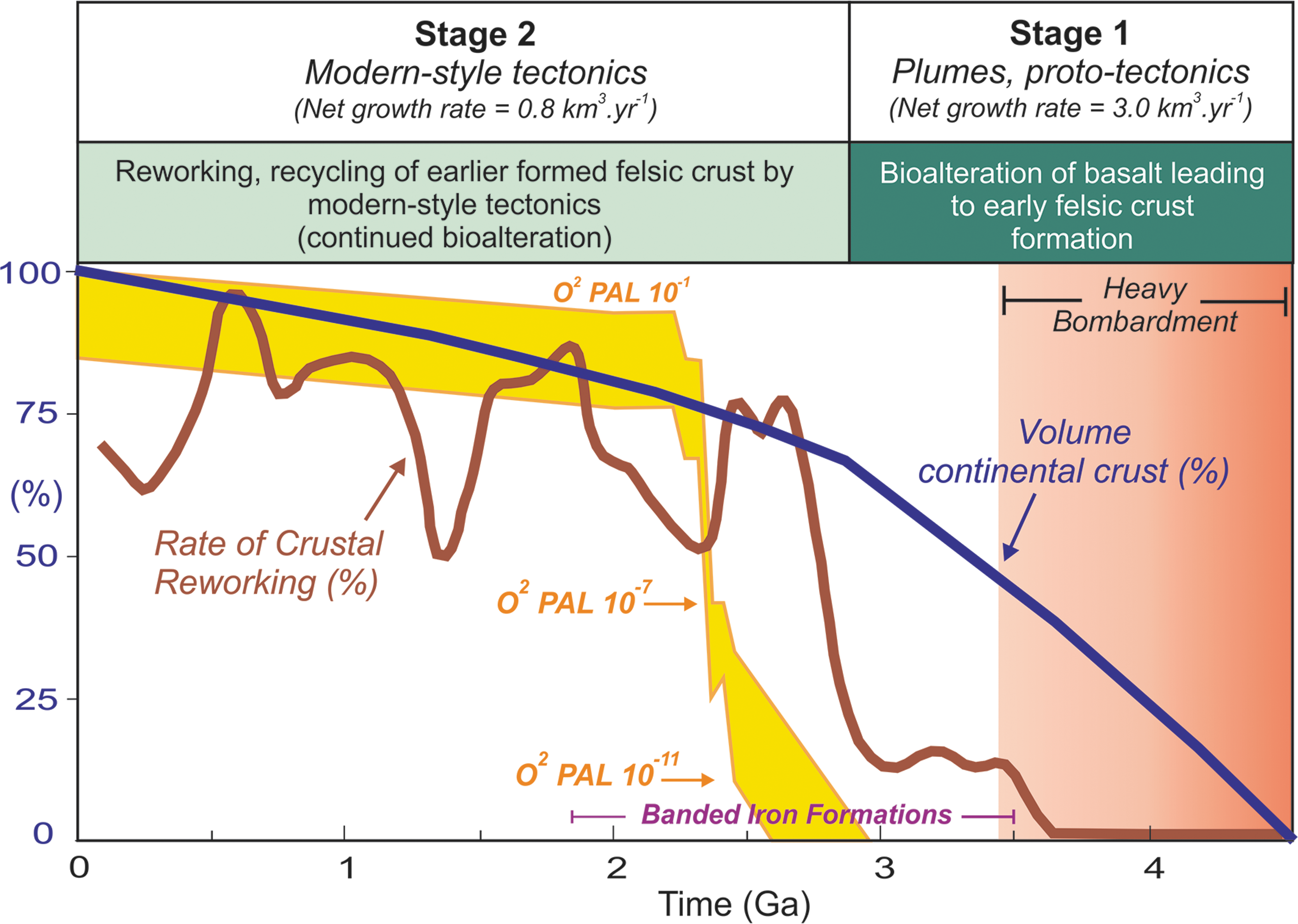
The possible role of subseafloor bioalteration in catalyzing the formation of early felsic continental crust under conditions of heavy impact bombardment. The continental crustal growth curves and rate of crustal reworking are modified after Dhuime et al. (2012). Stage 1 and Stage 2 reflecting change in crustal growth are also indicated (see Dhuime et al., 2012). The period of continuous heavy meteorite impact bombardment is taken from the work of Fernandes et al. (2013). The curve for atmospheric oxygen level over geological time is modified after Zahnle and Catling (2014) and references therein. Note that more than 45% of Earth's felsic continental crust was already produced by 3.5 billion years ago and occurred under the heavy bombardment conditions when microbial bioalteration may have been at its highest in the subseafloor of early Earth. “Whiffs” of oxygen appear around 2.5 billion years ago, but a major rise in oxygen atmospheric levels occurs at around 2.4–2.3 billion years ago (Great Oxidation Event), around ca. 600 million years after the onset of major crustal reworking in modern-style tectonics 3.0 billion years ago. Microbe-fluid-rock interaction and basalt hydration on early Earth may therefore have been a major driving force for the early formation of felsic crust on a plate-tectonic absent, or limited, Hadean–early Archean Earth with very low levels of crustal reworking and modern-style subduction to form granitoids.
The oldest oceanic rocks preserving evidence for extensive submarine hydrothermal activity, similar to modern-day activity at vents, are those of the 3.50–3.20 billion-year-old Barberton greenstone belt of South Africa (e.g., Paris et al., 1985; Hanor and Duchač, 1990; de Wit and Hart, 1993; Grosch et al., 2009a, 2009b; Philippot et al., 2009). Low-grade supracrustal sequences, including shallow marine sedimentary rocks in the Isua and Barberton greenstone belts, are also associated with TTG terrains. Evidence for life in the oceans, shallow marine sediments, and the oceanic subsurface has also been argued for from these greenstone belts (Rosing, 1999; Hofmann and Harris, 2008; McLoughlin et al., 2012; Grosch and McLoughlin, 2013). These observations suggest that between 3.50 and 3.20 billion years ago submarine proto-tectonic oceanic spreading centers were in operation as was an early form of shallow subduction and dehydration of descending oceanic lithosphere to form TTG intrusions. As such, major fluid-rock interaction between the geosphere and hydrosphere probably became more widespread and extensive across the Archean (Eo- to Paleoarchean), given the known evidence from the rock record. The onset of proto-tectonic processes and shallow subduction would have had a major influence on the diversity of new metamorphic minerals in Earth's early crust. Basaltic crust hydrated by microbially mediated alteration would have been subducted, releasing volatiles and modifying the local mantle, causing chemical and mineralogical heterogeneities. Proto-dehydration partial melting during shallow subduction would have led to the production of felsic arc magmas or tonalites (Fig. 3b). The earlier-formed buoyant felsic blocks produced by plume-melting would have been accreted together with later-formed TTGs from proto-subduction processes and volcanic arc-related felsic crust constituting significantly larger and more heterogeneous microcontinents (defined here in simple terms as an isolated fragment or small, early landmass of continental crust consisting of heterogeneous felsic rocks, i.e., tonalities, trondhjemites, granitoids, dacites, rhyolites, pegmatites, etc). As such, the early 4.0 Ga early-formed felsic sheets and crust would have been progressively incorporated as continental nuclei into the central parts of subduction-generated felsic arc terrains (Fig. 3b). It is thus possible that the earliest suboceanic microbial life was responsible for facilitating the earliest crustal differentiation and the formation of the earliest felsic, continental nuclei to microcontinents that are now preserved in the deepest roots of Archean cratons. By ca. 3.0 billion years ago, modern-style tectonics started to operate, producing continental crust in a manner similar to that seen above steep subduction zones beneath continental margins today (Fig. 3c).
Recent work has shown that one of the most important biogeochemical cycles operating at early oceanic spreading centers, hydrothermal vents, and in the Paleoarchean subseafloor environments was the sulfur cycle (e.g., McLoughlin et al., 2012; Grosch and McLoughlin, 2013, and references therein). Some workers have argued for an oxygenated early Archean atmosphere with fluctuating (“yo-yo”) oxygen levels over the Archean eon (e.g., see Ohmoto et al., 2006). Early low-levels of oxygen or “whiffs” of O2 have also been reported as early as 2.5 billion years ago (Anbar et al., 2007). However, it is widely accepted that the Hadean-Archean atmosphere was anoxic before the Great Oxidation Event 2.4–2.1 billion years ago (see Fig. 4; Farquhar et al., 2007; Kamber and Whitehouse, 2007; Lyons et al., 2014; Zahnle and Catling, 2014, and references therein). Prior to 2.4 billion years ago, Earth's reduced atmosphere allowed UV-induced photochemical reactions to take place, notably the photolysis of sulfur dioxide (SO2; e.g., Farquhar, 2001, 2007). The sulfur by-products of these photolysis reactions were used by early sulfur-based microbes in both subseafloor and surface environments. We highlight that, by promoting felsic crust formation and volcanism, microbes would have facilitated the formation of volcanic sulfur species in the early atmosphere—species such as sulfur dioxide, carbonyl sulfide, and hydrogen sulfide with methane, all of which can be used as metabolites by subsurface life in anoxic environments. Once proto-tectonic processes fully emerged and began operating more efficiently, more felsic-related volcanism would have changed the atmospheric composition further. New crustal environments in the form of sedimentary basins and other ecological niches would also have become available for early chemolithoautotrophic life (Fig. 3c). Thus fluid-rock-microbe interaction created many new feedbacks between proto-tectonics and sulfur biogeochemical cycling that may have been in place as early as ca. 3.5 billion years ago (e.g., Grosch and McLoughlin, 2013).
7. Microbes, Mineral Evolution, and Mars
One of the principle findings of the theory of mineral evolution is that different rocky planets achieve different stages of mineral evolution (Hazen et al., 2008; Hazen and Ferry, 2010; Hazen, 2013). In this new framework for mineralogy, Earth is divided into 10 stages of mineral evolution, reflecting an increase in the mineralogical diversity of our planet's near-surface environment, particularly as a consequence of the coevolving geosphere and biosphere (Hazen et al., 2008). For example, subsequent to 2.4 billion years ago with the rise in atmospheric oxygen levels due to the evolution of oxygenic photosynthesis (Canfield, 2014; Lyons et al., 2014), more than 2500 minerals formed as a result of oxidation and weathering (Hazen et al., 2008; Hazen and Ferry, 2010). In the current study, we explore the most likely role of early life before 2.4 billion years ago and postulate that this interaction at its onset would have contributed to an increase in Earth's near-surface mineral diversity. Although weathering may have been limited and the Hadean-Archean atmosphere reduced, subsurface microbial life may still have played a major role in changing the diversity and distribution of Earth's crustal minerals. At the earliest stage of coevolution between the biosphere and geosphere, subseafloor microbial life would have effectively facilitated hydration of the suboceanic lithosphere, causing disequilibrium and formation of new clay minerals in metavolcanic glass. Ultimately, microbe-induced alteration and hydration of Hadean-Archean oceanic crust led to early dehydration melting with the formation of the first evolved felsic crust and further mineral evolution (Fig. 3a, 3b). These early tonalite sheets and intrusions, consisting of feldspar, hornblende, quartz, and biotite, would represent the earliest fragments of microcontinents. This resulting change in Earth's surface mineralogy with early buoyant proto-continental crust may also have promoted the onset of proto-tectonic processes on an early cooling planet (Fig. 3b). These early ca. 4.0 Ga felsic bodies would then be exposed to further seawater hydrothermal alteration, leading to formation of more alteration products, such as epidote, mica, and chlorite group minerals. Metamorphic environments would have increased gradually from low pressure–intermediate temperature environments to higher-pressure environments as proto-tectonic mechanisms shifted to more modern-style tectonic processes (Fig. 3b). Initially, low-pressure contact metamorphism and seafloor hydrothermal alteration would have dominated during the earliest microbially induced alteration of oceanic crust and dehydration melting to form sialic crust (Fig. 3a). Gradually, with modern-style mountain building processes developing around ca. 3.2–3.0 billion years ago, higher-pressure minerals would have become more common at, or near, Earth's surface, contributing to mineral diversity (Fig. 3c). Thus from its origins life may have played a major role in shaping Earth's surface environment, driving mineral evolution by creating environments that led to increasing mineral diversity on an anoxic early Earth. If this hypothesis is true, then bioalteration in mafic to ultramafic suboceanic crust could be argued to be the most important early step toward creating a habitable planet with felsic crust formation and new surface environments, such as sedimentary basins. We can then ask: Did bioalteration of basaltic glass occur on Mars and, if so, to what extent? Furthermore, what factors are important for sustaining a deep suboceanic/subsurface biosphere on other potentially habitable planets?
Some workers have argued that energy generated from photosynthesis by living organisms as early as 3.8 billion years ago would have modified Earth's geochemical cycles and promoted weathering and diagenetic rock alteration, thus leading to the generation of early granitoids (Rosing et al., 2006). However, we suggest that microbially mediated alteration of oceanic lithosphere, which promoted direct melting by mantle plumes and proto-tectonics to form felsic crust, is a more plausible driving mechanism for granitoid generation from the widespread hydration of oceanic crust, especially under an early atmosphere with low oxygen levels that may not have supported photosynthesis. We propose that development of an early sulfur biogeochemical cycle that sustained subseafloor microbial life under anoxic atmospheric conditions is an important requirement in the early stages of development of a habitable planet.
In Fig. 5a, we demonstrate three potential factors that might have led to extensive interaction of a coevolving geosphere and biosphere on Earth, and contrast these processes to events on Mars. The diameters of the planets and the sizes of their inner cores are illustrated to scale. Mars, being a smaller planet with a different Fe/Mg ratio and smaller core, cooled more quickly and did not experience plate tectonics. Surface oxidation occurred at an early stage ca. 3.7 billion years ago on Mars (Ehlmann et al., 2011; Michalski et al., 2013; Tomkinson et al., 2013; Tuff et al., 2013), unlike Earth's surface environment that remained mostly anoxic until approximately 2.4 billion years ago. Sulfate minerals were already abundantly formed and stable on Mars' oxidized surface at 3.7 billion years ago (e.g., Ehlmann et al., 2013; Michalski et al., 2013), whereas sulfides were abundant under Earth's reducing conditions. We postulate that the onset of bioalteration as the earliest step toward a coevolving geosphere and biosphere may have been an important requirement that led to Earth developing into a habitable planet with complex life and mineral diversity. In addition, a delicate balance with subsequent feedbacks between life and Earth's surface environments evolved later by (1) proto-tectonic processes due to planetary cooling at the required rate to support temperatures at oceanic hydrothermal vents; (2) prolonged delay in oxidation of Earth's atmosphere; and (3) establishment of a sulfur biogeochemical cycle on early Earth. A combination of these factors most likely resulted in Earth evolving into a mineralogically and biologically complex planet, in contrast to Mars or other rocky planets in our solar system and beyond. It is possible that subsurface microbial bioalteration of mafic-ultramafic crust occurred on Mars, for example, in the shallow subsurface of martian impact craters (e.g., Michalski et al., 2013; Grosch et al., 2014, and references therein), but conditions on early Mars did not favor prolonged or extensive bioalteration/hydration of newly formed mafic crust at new spreading centers, owing to the absence of tectonics, which offers an explanation for why granitoid or felsic crust is rare on the Red Planet (Fig. 4).
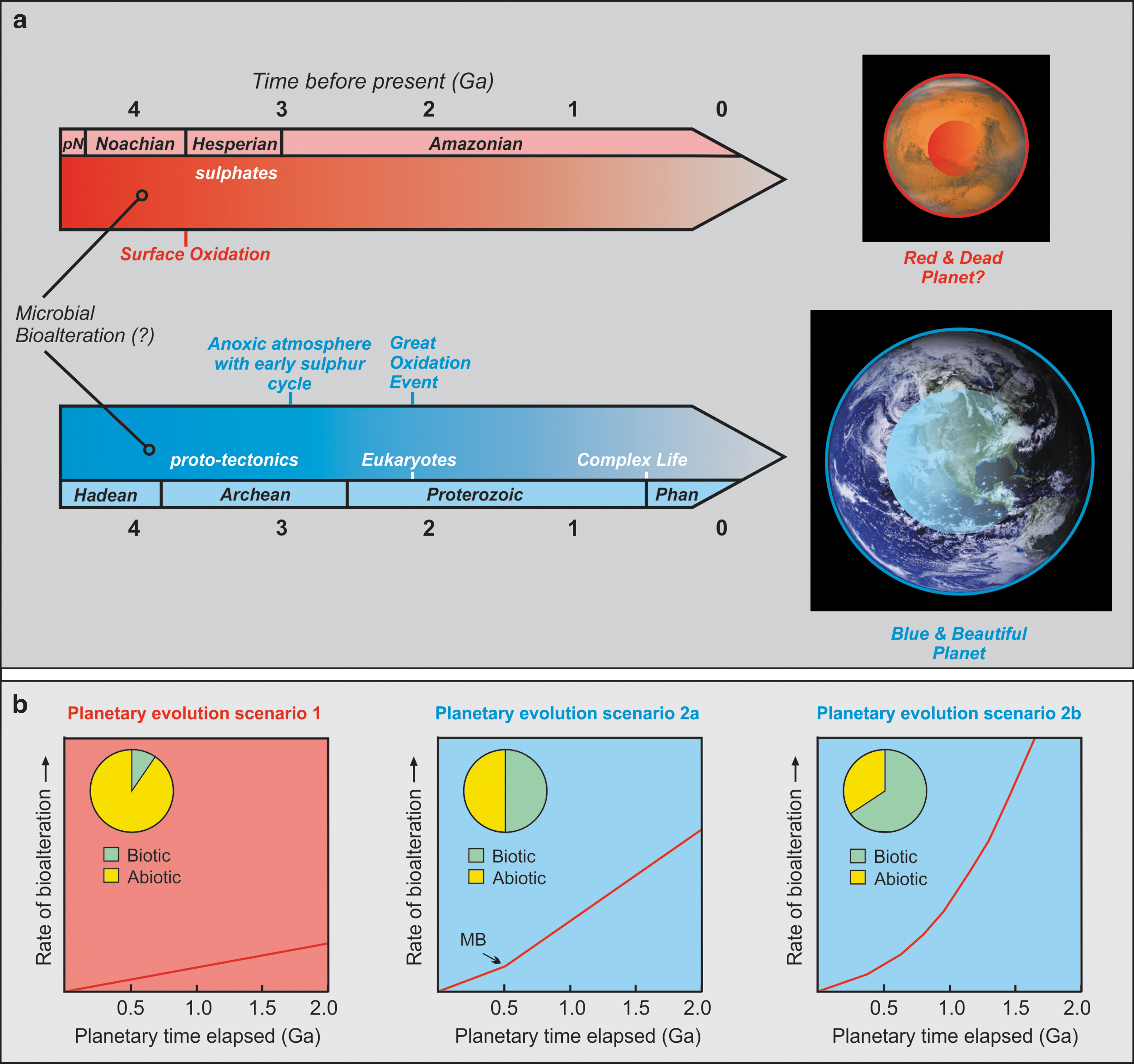
(
The current mineral-microbe coevolution hypothesis and its implications for planetary habitability may be extended and applied beyond Mars and to planetary bodies outside our solar system. In Fig. 5b, three planetary mineral evolution trajectories are presented, taking into account the relative rate of bioalteration, the planetary time elapsed, and the percentage of biotic versus abiotic alteration. There are currently no estimates on the rates of bioalteration from within the modern, in situ oceanic crust or from laboratory experiments (see Table 1 and references therein); thus we consider the following planetary evolution scenarios only as possible end-member cases to illustrate planetary mineral evolution trajectories (Fig. 5b). Planetary evolution scenario 1 depicts a low rate of bioalteration of subseafloor basalt and a high proportion of abiotic mineral alteration in the first 2 billion years of a planet's evolution. The result is a planetary body with limited felsic crust formation, similar to Mars. Planetary evolution scenario 2a depicts an Earth-like rocky planet that undergoes an increase in bioalteration rate after the first 500 million years of evolution due to a prolonged meteorite bombardment event that opens up new microbial ecological niches (hydrothermal impact craters, etc). In this scenario, 50% of the alteration of basaltic oceanic crust is represented to be biologically mediated. The probability of felsic crust formation and the onset of advanced mineral evolution in this case are much higher than in planetary scenario 1. On the other hand, planetary evolution scenario 2b illustrates a planetary body that experiences an exponential rate of basaltic subseafloor bioalteration and hydration (due to continuous meteorite bombardment, introduction of microbes by lithopanspermia, pioneering communities thriving in an FeO-rich basaltic environment, etc). In this end-member case, a much higher proportion of basalt hydration is due to microbially mediated bioalteration, and the planet has a very high probability of felsic crust generation, advanced mineral evolution, and the likelihood of becoming habitable to complex life. We propose planetary evolution scenario 2b for the first 2 billion years of early Earth mineral evolution, and that this may be a requirement (probably rare) in the formation of similar Earth-like habitable planets beyond our solar system.
8. Concluding Remarks
We hypothesize that, if abiotic microtunnels were present in Hadean subseafloor metavolcanic glass containing clay minerals, zeolites, and perhaps organic carbon, they may have provided a highly favorable site for prebiotic reactions. We postulate that the onset of interaction between the hydrosphere and geosphere would have been important for the origin of life in the subseafloor metabasaltic environment on the Hadean–early Archean Earth and the subsequent emergence of a subseafloor microbial biosphere. This early endolithic microbial biosphere thrived in the subseafloor environment and exploited fractures, hydrothermal fluids, and nutrients (e.g., Fe, Mn) released due to heavy meteorite impact bombardment. An unprecedented level of subseafloor bioalteration occurred at this early stage of Earth's history, whereby pioneering microbes formed granular alteration textures containing hydrated minerals analogous to those found in the modern oceanic crust. This early stage of bioalteration whereby endolithic microbes utilized energy from subseafloor rocks would have marked the onset of geosphere-biosphere coevolution. Advanced stages of granular microbial alteration would have promoted extensive alteration and effectively promoted dehydration melting of Hadean–early Archean oceanic lithosphere, leading to the earliest fragments of felsic crust. We conclude that microbial life may have been an essential agent in promoting continental crust formation and consequently initiating the extensive diversification of near-surface minerals on early Earth from subseafloor clay formation to felsic/granitoid minerals. Prior to 2.4 billion years ago, under anoxic atmospheric conditions, an early Paleoarchean sulfur biogeochemical cycle would have supported feedbacks between early subsurface life, the geosphere, hydrosphere, and atmosphere. We suggest that the onset of bioalteration was the earliest step toward a coevolving geosphere and biosphere, and an important prerequisite that led to Earth developing into a habitable planet with complex life and a correspondingly high degree of mineral diversity. In comparison to Mars, Earth's early evolution was different, as it retained an anoxic atmosphere for a longer period of time (from formation at 4.56 Ga up until 2.4 Ga). As a consequence, Earth established an early sulfur cycle that was most likely an important factor in the development of an environment that was capable of supporting extensive subseafloor microbial bioalteration. This biologically mediated alteration in turn promoted hydration melting and formation of the earliest felsic crust, thus increasing mineral diversity at, and near, Earth's evolving surface. Establishment of an early coevolving geosphere and biosphere that incorporated these critical factors may have been an important step that led to more widespread planetary habitability on Earth and, by analogy, perhaps necessary requirements prior to the evolution of complex life on any rocky planet. Thus, we conclude that the key factors that promoted the earliest evolution, diversification, and radiation of microbial life on Earth led to the evolution, diversification, and radiation of Earth's mineral kingdom, as well.
Footnotes
Acknowledgments
We thank Nicola McLoughlin for constructive discussions, also the Horizons lecture series committee of the Mathematics and Natural Sciences faculty at the University of Bergen, Norway, for helping to host Professor Robert Hazen as invited public speaker.